Activity and gene family expression of β-amylase in Brassica napus differing in harvest index
JIN Shu-Rong1,2,**, WANG Yan-Mei1,2,**, CHANG Yue1,2, WANG Yue-Hua1,2, LI Jia-Na1,2, NI Yu,1,2,*1 College of Agronomy and Biotechnology, Southwest University, Chongqing 400715, China 2 Academy of Agricultural Sciences, Southwest University, Chongqing 400715, China
This study was supported by the National Natural Science Foundation of China.31771694 the Chongqing Basic and Advanced Research Project.cstc2018jcyjAX0263 the Chongqing Basic and Advanced Research Project.cstc2016jcyjA0170 the Fundamental Research Funds for the Central Universities.XDJK2017B028 the China Agriculture Research System.CARS-12
Abstract The transferring efficiency of photosynthate from source organs to grains is the key process to increase the harvest index in Brassica napus, and amylase activity in source organs affects the transport intensity of assimilates to grains. The relationship between β-amylase (BAM) and the formation of high harvest index in rapeseed had not been clear. In this study, three different inbred lines, high-yield and high-harvest index rapeseed, high-yield and low-harvest index rapeseed, and low-yield and low-harvest index rapeseed were selected, and stem, leaf, silique pericarp and seed were sampled at 5, 10, 15, 20, and 25 d after the final flowering to analyze the activity of β-amylase and the expression level of its gene family members. The activity of β-amylase increased with the development of source organs. The activity of β-amylase in leaves and silique pericarps of high-harvest index rapeseed was significantly higher than that of low-harvest index rapeseed. In the β-amylase gene family, the expression of BAM1, BAM4, and BAM5 in the stem, leaf and silique pericarp of rapeseed increased with the development of organs. At 25 d after the final flowering, the expression of BAM1 and BAM3 in leaves and silique pericarps of high-harvest index rapeseed was significantly higher than that of low harvest index rapeseed. The expression of BAM4 and BAM5 in the silique pericarps of rapeseed with high harvest index was significantly higher than that of rapeseed with low harvest index at 15 d and 20 d after the final flowering. Taken together, BAM1 and BAM3 may enhance the transport intensity of photosynthate to grains by promoting starch degradation in leaves and silique pericarps in rapeseed with high harvest index, and BAM4 and BAM5 may regulate the transport of photosynthate to grains mainly by acting on starch degradation in silique pericarps. BAM4 and BAM5 may also be involved in the regulation of starch in rapeseed seeds. Keywords:Brassica napus;harvest index;β-amylase;;gene expression
PDF (576KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 靳舒荣, 王艳玫, 常悦, 王月华, 李加纳, 倪郁. 不同收获指数甘蓝型油菜β-淀粉酶活性及其基因家族成员的表达分析[J]. 作物学报, 2019, 45(8): 1279-1285. doi:10.3724/SP.J.1006.2019.94001 JIN Shu-Rong, WANG Yan-Mei, CHANG Yue, WANG Yue-Hua, LI Jia-Na, NI Yu. Activity and gene family expression of β-amylase in Brassica napus differing in harvest index[J]. Acta Agronomica Sinica, 2019, 45(8): 1279-1285. doi:10.3724/SP.J.1006.2019.94001
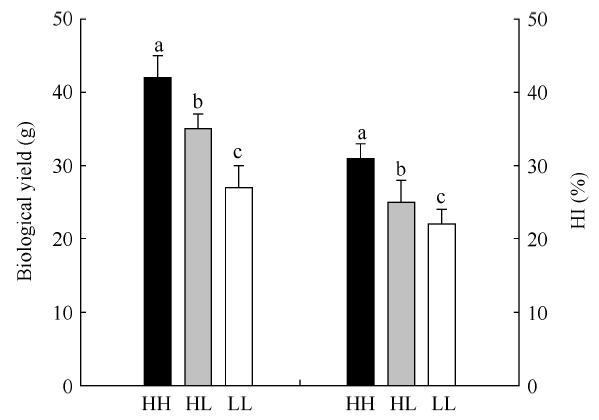
HH: 高产高收获指数型油菜; HL: 高产低收获指数型油菜; LL: 低产低收获指数型油菜; HI: 收获指数。图中柱形图上方不同小写字母表示品种间差异显著(P < 0.05)。 Fig. 1Biological yield and harvest index in different B. napus varieties
HH: high-yield and high-harvest index rapeseed; HL: high-yield and low-harvest index rapeseed; LL: low-yield and low-harvest index rapeseed; HI: harvest index. Different letters above the column indicate significant difference at P < 0.05 among varieties.
Yuan WZ, Guan CY . Regulation of assimilates transportation by amylase activity in rapeseed pods J Nat Sci Univ Norm Hunan, 1995,18(3):74-79 (in Chinese with English abstract). [本文引用: 2]
Yuan WZ, Guan CY . Harvest index in rapeseed affected by a few physiological factors Acta Agron Sin, 1997,23:580-586 (in Chinese with English abstract). [本文引用: 2]
Yu TS, Zeeman SC, ThorneycroftD, Fulton DC, DunstanH, Lue WL, HegemannB, Tung SY, UmemotoT, ChappleA, Tsai DL, Wang SM, Smith AM, ChenJ, SmithS M . alpha-Amylase is not required for breakdown of transitory starch in Arabidopsis leaves J Biol Chem, 2005,280:9773-9779. DOI:10.1074/jbc.M413638200URL [本文引用: 1]
Weise SE, Kim KS, Stewart RP, Sharkey TD . β-Maltose is the metabolically active anomer of maltose during transitory starch degradation Plant Physiol, 2005,137:756-761. DOI:10.1104/pp.104.055996URL [本文引用: 2]
ScheidigA, Fr?hlichA, SchulzeS, Lloyd JR, KossmannJ . Down regulation of a chloroplast-targeted β-amylase leads to a starch-excess phenotype in leaves Plant J, 2002,30:581-591. DOI:10.1046/j.1365-313X.2002.01317.xURL [本文引用: 2]
Shen GZ . Balance and Regulation of Hormones of High Harvest Index Rapeseed (Brassica napus L.) in Different Environments MS Thesis of Southwest University, Chongqing, China, 2016 (in Chinese with English abstract). [本文引用: 2]
Li JN, LuK, Jian HJ, LiangY, Lu JH, PengL, Shen GZ, ZhangY, ZhangC, YangB, ZhangL . Research advances on harvest index of Brassica napus L Chin J Oil Crop Sci, 2018,40:640-648 (in Chinese with English abstract). [本文引用: 1]
Allen EJ, Morgan DG, Ridgman WI . A physiological analysis of the growth of oilseed rape J Agric Sci, 1971,77:339-341. DOI:10.1017/S0021859600024515URL [本文引用: 2]
Chapman JF, Daniels RW, Scarisbrick DH . Field studies on 14C assimilate fixation and movement in oil-seed rape (B. napus) J Agric Sci, 1984,102:23-31. [本文引用: 1]
Pechan PA, Morgan DG . Defoliation and its effects on pod and seed development in oil seed rape (Brassica napus L.) J Exp Bot, 1985,36:458-468. [本文引用: 1]
Tayo TO, Morgan DG . Factors influencing flower and pod development in oil-seed rape (Brassica napus L.) J Agric Sci, 1979,92:363-373. [本文引用: 1]
Smith SM, Fulton DC, ChiaT, ThorneycroftD, ChappleA, DunstanH, HyltonC, Zeeman SC, Smith AM . Diurnal changes in the transcriptome encoding enzymes of starch metabolism provide evidence for both transcriptional and posttranscriptional regulation of starch metabolism in Arabidopsis leaves Plant Physiol, 2004,136:2687-2699. DOI:10.1104/pp.104.044347URL [本文引用: 1]
Monroe JD, Storm AR . The Arabidopsis β-amylase (BAM) gene family: Diversity of form and function Plant Sci, 2018,276:163-170. DOI:10.1016/j.plantsci.2018.08.016URL [本文引用: 1]
Fulton DC, StettlerM, MettlerT, Vaughan CK, LiJ, FranciscoP, GilM, ReinholdH, EickeS, MesserliG, DorkenG, HallidayK, Smith AM, Smith SM, Zeeman SC . β-amylase 4, a noncatalytic protein required for starch breakdown, acts upstream of three active beta-amylases in Arabidopsis chloroplasts Plant Cell, 2008,20:1040-1058. DOI:10.1105/tpc.107.056507URL [本文引用: 3]
Lao NT, SchoneveldO, Mould RM, Hibberd JM, Gray JC, Kavanagh TA . An Arabidopsis gene encoding a chloroplast- targeted β-amylase Plant J, 1999,20:519-527. DOI:10.1046/j.1365-313X.1999.00625.xURL [本文引用: 2]
ValerioC, CostaA, MarriL, Issakidis-BourguetE, PupilloP, TrostP, SparlaF . Thioredoxin-regulated β-amylase (BAM1) triggers diurnal starch degradation in guard cells, and in mesophyll cells under osmotic stress J Exp Bot, 2011,62:545-555. DOI:10.1093/jxb/erq288URL [本文引用: 1]
HorrerD, FlütschS, PazminoD, Matthews JS, ThalmannM, NigroA, LeonhardtN, LawsonT, SanteliaD . Blue light induces a distinct starch degradation pathway in guard cells for stomatal opening Curr Biol, 2016,26:362-370. DOI:10.1016/j.cub.2015.12.036URL [本文引用: 1]
KaplanF, Guy CL . RNA interference of Arabidopsis beta- amylase 8 prevents maltose accumulation upon cold shock and increases sensitivity of PSII photochemical efficiency to freezing stress Plant J, 2005,44:730-743. DOI:10.1111/tpj.2005.44.issue-5URL [本文引用: 1]
Monroe JD, Storm AR, Badley EM, Lehman MD, Platt SM, Saunders LK, Schmitz JM, Torres CE . β-amylase 1 and β-amylase3 are plastidic starch hydrolases in Arabidopsis that seem to be adapted for different thermal, pH, and stress conditions Plant Physiol, 2014,166:1748-1763. DOI:10.1104/pp.114.246421URL [本文引用: 1]
Monroe JD, Breault JS, Pope LE, Torres CE, Gebrejesus TB, Berndsen CE, Storm AR . Arabidopsis β-amylase 2 is a K+-requiring, catalytic tetramer with sigmoidal kinetics Plant Physiol, 2017,175:1125-1135. [本文引用: 1]
Laby RJ, KimD, Gibson SI . The ram1 mutant of Arabidopsis exhibits severely decreased β-amylase activity Plant Physiol, 2001,127:1798-1807. [本文引用: 1]
HuangL, Tao SS, ZhangM, JiangL, Peng YL . Differences analysis of harvest and varieties of Brassica napus L Jiangsu Agric Sci, 2011, (1):95-97 (in Chinese with English abstract). [本文引用: 1]
 ,1,2,*
,1,2,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT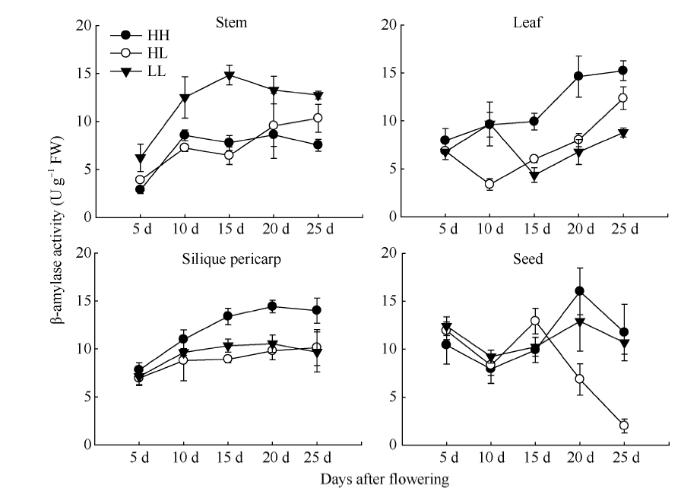 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT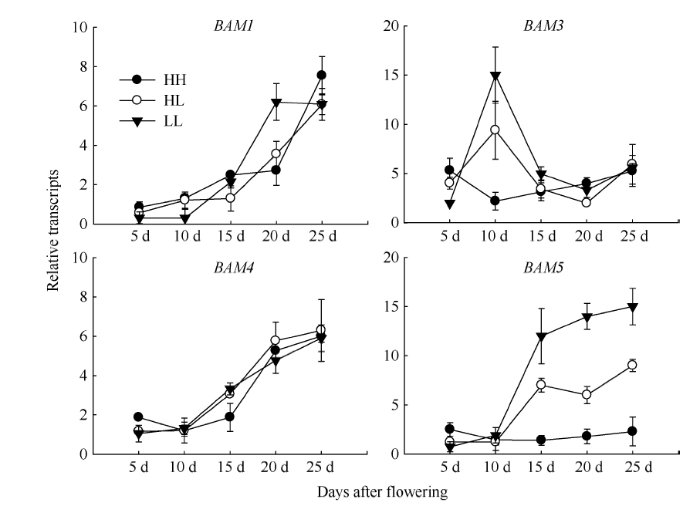 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT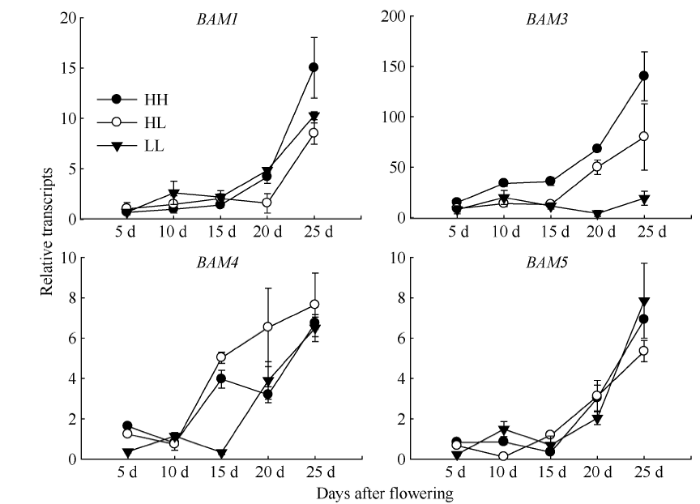 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT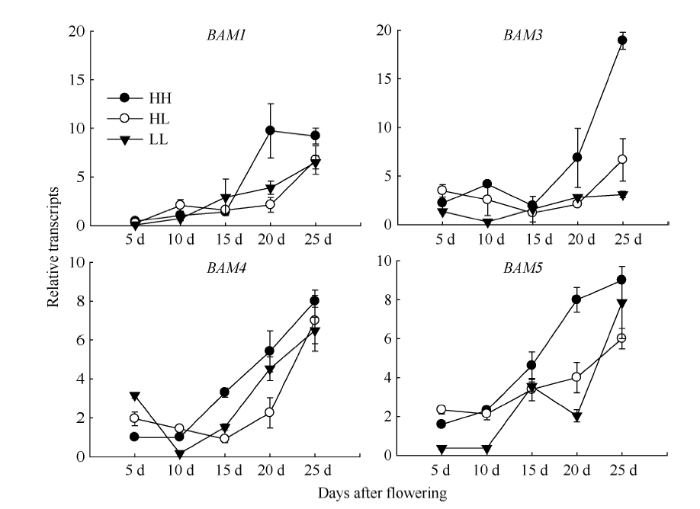 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT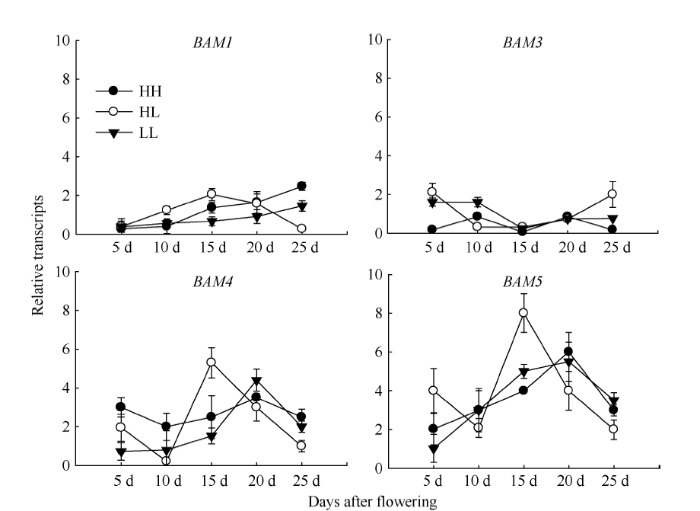 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}