Genetic analysis of plant height related traits in Ricinus communis L. with major gene plus polygenes mixed model
CUI Yue1,**, LU Jian-Nong2,**, SHI Yu-Zhen3, YIN Xue-Gui,2,*, ZHANG Qi-Hao21 Lingnan Normal University, Zhanjiang 524048, Guangdong, China 2 College of Agricultural Sciences, Guangdong Ocean University, Zhanjiang 524088, Guangdong, China 3 College of Chemistry and Environment, Guangdong Ocean University, Zhanjiang 524088, Guangdong, China
This study was supported by the National Natural Science Foundation of China.31271759 the Guangdong Provincial Science and Technology Projects.2013B060400024 the Guangdong Provincial Science and Technology Projects.2014A020208116 the Guangdong Provincial Science and Technology Projects.2016A020208015 Project of Enhancing School with Innovation of Guangdong Ocean University.GDOU2013050206
Abstract In this study, plant height related traits in Ricinus communis L. were analyzed using the mixed major gene plus polygenes genetic model with two groups of six-generation populations (P1, P2, F1, B1, B2, and F2) derived from the cross YC2×YF1. The results revealed that the plant height was controlled by a pair of major gene and polygenes. The major-gene heritability in B1, B2, and F2 populations was 37.05%/49.57% (group I/group II), 30.51%/34.48%, and 43.98%/43.64%, respectively. The bearing height of primary raceme and the node number of main stems were all controlled by two pairs of major genes and polygenes, with the importance of major-gene genetic components in the order of epistasis >dominance >additive. In the three generations the heritability of major genes conferring the bearing height of primary raceme was 67.91%/92.72%, 86.89%/92.13%, and 60.18%/66.87%, respectively, and that of major genes conferring the node number of main stem were 91.83%/91.50%, 35.22%/63.37%, and 85.76%/94.58%, respectively. As for the length of main stem internode, it was fully controlled by polygenes, the heritability was 47.64%/47.64%, 38.87%/38.87%, and 25.25%/52.71%, respectively. The above genetic models explained the positive transgressive inheritance of plant height, the bearing height of primary raceme and the length of main stem internode as well as the similar performance to the low value parent of the node number of main stem in F1 generation. It suggested that the bearing height of primary raceme and the node number of main stem should be used as indirect selection indexes for plant height at early stage and the node number of main stem of lower value parent should not be too little in high yield breeding. Keywords:Ricinus communis L.;plant height related traits;major gene plus polygene model;genetic analysis
PDF (432KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 崔月, 陆建农, 施玉珍, 殷学贵, 张启好. 蓖麻株高性状主基因+多基因遗传分析[J]. 作物学报, 2019, 45(7): 1111-1118. doi:10.3724/SP.J.1006.2019.84127 CUI Yue, LU Jian-Nong, SHI Yu-Zhen, YIN Xue-Gui, ZHANG Qi-Hao. Genetic analysis of plant height related traits in Ricinus communis L. with major gene plus polygenes mixed model[J]. Acta Crops Sinica, 2019, 45(7): 1111-1118. doi:10.3724/SP.J.1006.2019.84127
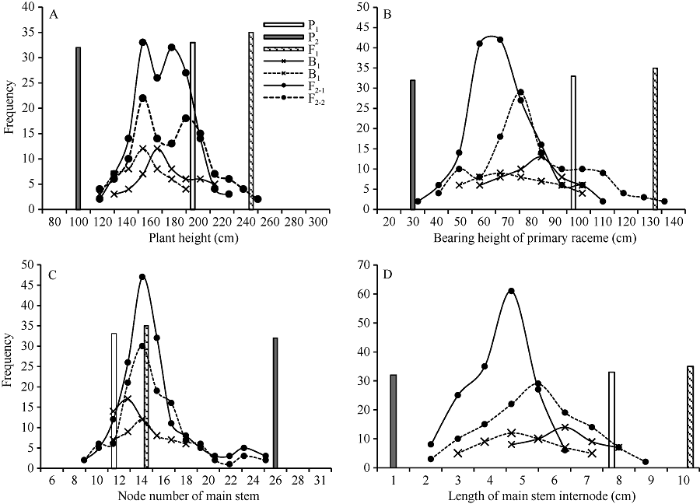
亲本为连续套袋自交7代的纯合自交系, 其中YC2 (P1)为高秆雌性系, 株高212.3 cm, YF1 (P2)为矮秆两性系, 株高87.6 cm。F1群体中有1株主穗表现为单雌, 其余均为两性。从F1中选取主穗单雌株和两性株分别自交, 创建了2个F2群体, 即F2-1和F2-2, 与同一组P1、P2、F1、B1 (F1×P1)和B2 (F1×P2)构成2组六世代群体(I组和II组)。2013年10月20日, 于广东海洋大学实验基地同一地块种植所有群体, 试验地前茬为水稻, 地势平坦, 肥力均匀。行株距为1 m × 1 m, 穴播, 每穴3粒, 三叶时随机定苗一株。P1、P2、F1、B1和B2群体分别为33、32、35、51和48株。由于F2群体中有少数几株在其他植株二级分枝穗成熟2个月后仍不现蕾, 继续保持营养生长状态而未能调查其株高性状, 最终F2-1群体162株, F2-2群体121株。
1.2 性状测定
2014年5月二级分枝穗成熟时, 按照《蓖麻种质资源描述规范和数据标准》[44]测量各群体单株株高(plant height, PH)、主穗位高(bearing height of primary raceme, PRH)、主茎节数(node number of main stem, MSNN), 并计算主茎节长(length of main stem internode, MSIL),
Table 1 表1 表1六世代群体株高性状表型特征值 Table 1Characteristics of plant height related traits in six generations
性状 Trait
世代 Generation
单株数 No. of plants
最大值 Max
最小值 Min
极差 Rage
平均值 Mean
标准差 SD
变异系数 CV (%)
株高 PH (cm)
P1
33
226.00
184.00
42.00
208.12
9.03
4.34
P2
32
110.00
74.00
36.00
98.03
8.22
8.39
F1
35
245.00
224.00
21.00
230.84
1.53
0.66
B1
51
238.00
100.00
138.00
171.12
6.74
3.94
B2
48
198.00
77.00
121.00
135.82
6.21
4.57
F2-1
162
260.00
60.00
200.00
159.51
38.14
23.91
F2-2
121
281.00
80.00
201.00
174.10
45.96
26.40
主穗位高 PRH (cm)
P1
33
112.00
87.00
25.00
101.69
5.35
5.26
P2
32
38.00
24.00
14.00
31.33
3.17
10.13
F1
35
129.00
118.00
11.00
125.01
3.48
2.78
B1
51
100.00
49.00
51.00
75.65
15.80
20.89
B2
48
94.00
39.00
55.00
63.42
16.02
25.26
F2-1
162
105.00
12.00
93.00
57.52
22.18
37.35
F2-2
121
137.00
29.00
108.00
69.04
34.05
45.90
主茎节数 MSNN
P1
33
15
9
6
12.12
1.96
16.17
P2
32
31
21
10
26.89
2.38
8.85
F1
35
15
10
5
12.96
1.51
11.65
B1
51
15
9
6
10.68
1.65
15.45
B2
48
25
9
16
14.84
3.14
21.16
F2-1
162
31
4
27
14.56
4.48
30.77
F2-2
121
29
5
24
14.99
4.86
32.42
主茎节长MSIL (cm)
P1
33
11.20
5.40
5.80
8.14
1.36
16.71
P2
32
1.60
0.90
0.70
1.19
0.14
11.76
F1
35
12.10
9.00
3.10
10.25
0.91
8.88
B1
51
9.00
4.45
4.55
6.52
1.24
19.02
B2
48
7.58
2.81
4.77
5.14
1.22
23.74
F2-1
162
7.30
1.58
5.72
4.17
1.16
27.82
F2-2
121
9.55
2.00
7.55
4.99
1.46
29.26
PH: plant height; PRH: bearing height of primary raceme; MSNN: node number of main stem; MSIL: length of main stem internode; F2-1: F2 population in group I; F2-2 : F2 population in group II. F2-1: I组F2群体; F2-2: II组F2群体。
Table 2 表2 表2入选模型的适合性检验结果 Table 2Test for goodness-of-fit of selected genetic models
性状 Trait
模型 Model
模型含义 Implication of model
极大对数似然函数值 Max log likelihood value
AIC 值 AIC value
适合性检验a Test of goodness-of-fita
I组
株高 PH
D-0
MX1-AD-ADI
-1215.34
2458.23
0/0/0/0/0
Group I
主穗位高 PRH
E-1
MX2-ADI-AD
-1033.05
2097.31
0/0/0/0/0
主茎节数 MSNN
E-1
MX2-ADI-AD
-648.77
1326.46
0/0/0/0/0
主茎节长 MSIL
C-0
PG-ADI
-404.12
827.38
0/0/0/0/0
II组
株高 PH
D-0
MX1-AD-ADI
-898.87
1821.42
0/0/0/0/0
Group II
主穗位高 PRH
E-1
MX2-ADI-AD
-776.23
1585.13
0/0/0/0/0
主茎节数 MSNN
E-1
MX2-ADI-AD
-475.89
981.14
0/0/0/0/0
主茎节长 MSIL
C-0
PG-ADI
-324.56
668.02
0/0/0/0/0
a Five digitals in test of goodness-of-fit column denote numbers of significant statistic parameters among U12, U22, U32, nW2, and Dn. MX: mixed major gene and polygene model. A: additive effect; D: dominance effect; I: interaction; PG: polygene model. PH: plant height; PRH: bearing height of primary raceme; MSNN: node number of main stem; MSIL: length of main stem internode. a “适合性检验”中5个数字分别指在U12、U22、U32、nW2和Dn适合性检验中达到显著水平的统计量个数。MX: 主基因+多基因遗传模型; A: 加性效应; D: 显性效应; I: 上位性; PG: 多基因遗传模型。
Table 3 表3 表3株高性状遗传参数估计值 Table 3Estimates of genetic parameters for plant height related traits
世代 Generation
遗传参数 Genetic parameter
株高 PH
主穗位高 PRH
主茎节数 MSNN
主茎节长 MSIL
(D-0) I
(D-0) II
(E-0) I
(E-0) II
(E-0) I
(E-0) II
(C-0) I
(C-0) II
da
35.87
41.63
-9.39
-3.88
-4.44
-4.58
db
-9.39
-3.88
-4.44
-4.58
ha
-4.85
-8.27
17.78
10.22
-6.60
-7.27
hb
17.78
10.16
-8.17
-7.68
i
42.42
42.72
-0.07
-1.34
jab
-11.82
0.77
2.30
0.28
jba
-11.82
-16.41
3.88
3.12
l
40.39
29.78
9.12
7.79
[d]
16.54
27.51
-0.57
1.95
[h]
95.17
83.79
-3.08
-0.47
ha/da
-0.14
-0.20
-1.89
-2.64
1.49
1.59
hb/db
-1.89
-2.64
1.84
1.68
B1
σ2p
1243.83
1235.34
255.84
255.84
2.94
2.94
1.91
1.91
σ2mg
460.84
612.36
173.74
237.22
2.70
2.69
σ2pg
718.38
561.23
64.80
1.32
0.00
0.00
0.91
0.91
h2mg (%)
37.05
49.57
67.91
92.72
91.83
91.50
h2pg (%)
57.86
45.31
25.40
0.53
0.00
0.00
47.64
47.64
B2
σ2p
899.84
900.96
282.33
282.33
13.46
13.46
1.65
1.65
σ2mg
274.54
310.65
245.32
260.11
4.74
8.53
σ2pg
561.03
524.54
20.07
5.35
5.42
2.90
0.64
0.64
h2mg (%)
30.51
34.48
86.89
92.13
35.22
63.37
h2pg (%)
62.21
58.15
7.07
1.86
40.32
21.55
38.87
38.87
F2
σ2p
1448.34
2086.94
380.38
779.11
20.09
23.63
1.35
2.12
σ2mg
636.98
910.74
228.91
520.99
17.23
22.35
σ2pg
754.25
1138.15
135.22
241.46
0.00
0.00
0.34
1.12
h2mg (%)
43.98
43.64
60.18
66.87
85.76
94.58
h2pg (%)
51.54
53.47
35.45
30.94
0.00
0.00
25.25
52.71
da: additive effect of the first major gene; ha: dominant effect of the first major gene; db: additive effect of the second major gene; hb: dominant effect of the second major gene; i: epistatic effect value between da and db; jab: epistatic effect value between da and hb; jba: epistatic effect value between ha and db; l: epistatic effect value between ha and hb; [d]: additive effect of polygene; [h]: dominant effect of polygene; σ2p: phenotypic variance; σ2pg: polygene variance; σ2mg: major gene variance; h2mg (%): heritability of major gene; h2pg (%): heritability of polygene. PH: plant height; PRH: bearing height of primary raceme; MSNN: node number of main stem; MSIL: length of main stem internode. da: 第1对主基因的加性效应; ha: 第1对主基因的显性效应; db: 第2对主基因的加性效应; hb: 第2对主基因的显性效应; i: 加性×加性互作; jab: 加性×显性互作; jba: 显性×加性互作; l: 显性×显性互作; [d]: 多基因加性效应; [h]: 多基因的显性效应; σ2p: 表型方差; σ2pg: 多基因方差; σ2mg: 主基因方差; h2mg (%): 主基因遗传力; h2pg (%): 多基因遗传力。
Table 4 表4 表4株高相关性状间的相关系数 Table 4Correlation coefficients between plant height related traits
性状Trait
株高PH
主穗位高PRH
主茎节数MSNN
主茎节长MSIL
株高 PH
0.844**
0.474**
0.709**
主穗位高 PRH
0.725**
0.726**
0.613**
主茎节数 MSNN
0.309**
0.569**
-0.242**
主茎节长 MSIL
0.508**
0.600**
-0.254**
** Significance at the 0.01 probability level. PH: plant height; PRH: bearing height of primary raceme; MSNN: node number of main stem; MSIL: length of main stem internode. **表示在0.01概率水平差异显著。
Li JQ, Zhu GL, Wu GL, He ZB, Li JX, Zhang CH, Tian FD, Jia JX . Correlation and path analysis between seed oil content and major quantitative characters in Castor Chin J Oil Crop Sci, 2004,26(2):43-46 (in Chinese with English abstract). [本文引用: 1]
FoissetN, DelourmeR, BarretP, RenardM . Molecular tagging of the dwarf BREIZH (Bzh) gene in Brassica napus. Theor Appl Genet, 1995,91:756-761. [本文引用: 1]
Sun DS, Li WB, Zhang ZC . Quantitative trait loci analysis for the developmental behavior of soybean (Glycine max L. Merr.). Theor Appl Genet, 2006,112:665-673. [本文引用: 1]
Sun XL, Wu ST, Song XE . Studies on characters and genetic effects of determinate cucumber Acta Hortic Sin, 1990,17:59-64 (in Chinese with English abstract). [本文引用: 1]
Gao FM, JiangY, Kong DW, Li SG . Genetic control of plant height and its utilization in rice Mol Plant Breed, 2005,3:87-93 (in Chinese with English abstract). [本文引用: 1]
ZhaoL . Cloning and Functional Analysis of the PAY1 Gene Controlling Plant Architecture and Grain Yield of Rice (Oryza sativa L.). PhD Dissertation of China Agricultural University, Beijing, China, 2015 (in Chinese with English abstract). [本文引用: 1]
Chen GF, Chen JS, Tian JC . Genome-wide association analysis between SNP markers and plant height related traits in wheat Acta Agron Sin, 2015,41:1500-1509 (in Chinese with English abstract). [本文引用: 1]
Yang SJ, Zhang XK, He ZH, Xia XC, ZhouY . Distribution of dwarfing genes Rht-B1b and Rht-D1b in Chinese bread wheats detected by STS marker. Sci Agric Sin, 2006,39:1680-1688 (in Chinese with English abstract). [本文引用: 1]
林泽川, 曹立勇 . 水稻株型相关基因的定位与克隆研究进展 中国稻米, 2014,20(1):17-22. DOI:10.3969/j.issn.1006-8082.2014.01.004Magsci [本文引用: 1] 水稻株型相关性状包括分蘖数、分蘖夹角、株高及穗部性状。株型相关性状都是重要的农艺性状,是水稻产量因素的重要组成部分。株型形成涉及到一系列基因的表达和表达产物行使功能,因此对这些基因的挖掘和功能分析具有重要意义。目前,虽尚未完全了解控制株型相关性状的基因及其功能,但也已取得了日新月异的进展。本文针对各个株型相关性状,综述了已鉴定的基因及其功能特点,以为分子育种运用及进一步遗传生理研究作理论基础。 Lin ZC, Cao LY . Advances in mapping and cloning plant type related genes on rice Chin Rice, 2014,20(1):17-22 (in Chinese). DOI:10.3969/j.issn.1006-8082.2014.01.004Magsci [本文引用: 1] 水稻株型相关性状包括分蘖数、分蘖夹角、株高及穗部性状。株型相关性状都是重要的农艺性状,是水稻产量因素的重要组成部分。株型形成涉及到一系列基因的表达和表达产物行使功能,因此对这些基因的挖掘和功能分析具有重要意义。目前,虽尚未完全了解控制株型相关性状的基因及其功能,但也已取得了日新月异的进展。本文针对各个株型相关性状,综述了已鉴定的基因及其功能特点,以为分子育种运用及进一步遗传生理研究作理论基础。
Zhao YM . Analysis of genetic effect and genetype by environment of ear height on maize Maize Sci, 2009,17(2):12-14 (in Chinese with English abstract). [本文引用: 2]
Lan JH, ChuD . Study on the genetic basis of plant height and ear height in Maize (Zea mays L.) by QTL dissection. Hereditas, 2005,27:925-934 (in Chinese with English abstract). [本文引用: 1]
Yang XJ, LuM, Zhang SH, ZhouF, Qu YY, Xie CX . QTL mapping of plant height and ear position in maize (Zea mays L.). Hereditas, 2008,30:1477-1486 (in Chinese with English abstract). [本文引用: 1]
BaiW, ZhangH, ZhangZ, TengF, WangL, Tao Y. ZhengY . The evidence for non-additive effect as the main genetic component of plant height and ear height in maize using introgression line populations Plant Breed, 2010,129:376-384. [本文引用: 1]
Yuan LZ, Tang JH, Wang XP, Li CH . QTL analysis of shading sensitive related traits in maize under two shading treatments PLoS One, 2012,7:e38696. DOI:10.1371/journal.pone.0038696URL [本文引用: 1]
Cai HG, ChuQ, Gu RL, Yuan LX, Liu JC, Zhang XZ, Chen FJ, Mi GH, Zhang FS . Identification of QTLs for plant height, ear height and grain yield in maize (Zea mays L.) in response to nitrogen and phosphorus supply. Plant Breed, 2012,131:502-501. [本文引用: 1]
PengJ, Cai YL, Xu DL, Wang GQ . Genetic analysis of plant type traits in joint multi-generational on maize J Mathematical Biol, 2009,24:149-156 (in Chinese). [本文引用: 1]
ZhaoG, Wu ZK, Wang BW . Major gene plus polygene inheritance of plant height and ear height in microedosperm super-high oil corn J Anhui Agric Sci, 2007,35:5096-5098 (in Chinese with English abstract). [本文引用: 1]
Wang TG, MaJ, Zhang HS, Chen SL . Analysis of major gene plus polygenes genetic model for ear height of maize Guizhou Agric Sci, 2012,40(4):10-13 (in Chinese with English abstract). [本文引用: 4]
Qiu ZG, Wang GX, YangH, Qi ZY, Ge ZH, Zhang SH, Cai ZR . Air mutagenesis mutant waxy corn plant height raceme bits of genetic models J Southwest Univ (Nat Sci Edn), 2008,30(3):60-65 (in Chinese with English abstract). [本文引用: 1]
Cai LN . Studies on the Major Gene Plus Polygene Interitance of Principal Agronomy Traits and Quality Traits in Soybean. MS Thesis of Jilin Agricultural University, Changchun, Jilin, China, 2012 (in Chinese with English abstract). [本文引用: 1]
Cao YQ, Dong LJ, Lyu GL, Sun XG, Wang WB, Song SH, Xie FT . Different parents soybean reciprocal crosses F2, F3, BC1F2 stem node number genetic research Soybean Sci Technol, 2009, ( 1):18-21 (in Chinese with English abstract). [本文引用: 2]
Li YQ . Soybean interspecific hybridization main agronomic traits of maternal effect at the beginning of the study Crops, 2006, ( 6):28-30 (in Chinese). [本文引用: 2]
Lin GQ, Xu SC, Huang JC, Chen ZX . Analysis of genetic parameters of main characteristics of F2 in different parent forms in soybean J Plant Genetic Resour, 2001,2(2):12-15 (in Chinese with English abstract). [本文引用: 2]
Qi ZY, Li JX, Zou XX, Cao LW, Rao LL, Yu JL, Chen LP . Genetic analysis of plant architecture traits in melon (Cucumis melo L.). J Agric Biotechnol, 2015,23:302-310 (in Chinese with English abstract). [本文引用: 1]
ZhangJ H . Plant Height, Internodes, Ear traits, General Combining Ability Analysis and Their QTLs Mapping Using Double Haploid Lines of Maize. PhD Dissertation of Agricultural University of Hebei, Baoding, Hebei, China, 2009 (in Chinese with English abstract). [本文引用: 2]
Severino LS, Auld DL, BaldanziM, CandidoM J D, ChenG, CrosbyW, TanD, HeX, LakshmammaP, LavanyaC, MachadoO L T, MielkeT, MilaniM, MillerT D, MorrisJ B, MorseS A, NavasA A, SoaresD J, SofiattiV, WangM L, ZanottoM D, ZielerH . A review on the challenges for increased production of castor Agron J, 2012,104:853-870. DOI:10.2134/agronj2011.0210URL [本文引用: 1]
YaoY, Li FS, Chen YS, Li JQ, Huang FL, Wang YJ . Research progress on castor J Inner Mongolia Univ Nationalities, 2009,24(2):172-174 (in Chinese with English abstract). [本文引用: 1]
Deng CH, SunQ . A correlation and path coefficient analysis of main quantitative characters in castor Jilin Agric Sci, 1992, ( 3):25-28 (in Chinese with English abstract). [本文引用: 1]
Li JQ, Zhang ZY, He ZB, Jia JX, Qiao WJ . Dwarf castor-oil plant hybrids yield and the main agronomic characters of correlation and multiple regression analysis J Inner Mongolia Univ Nat(Nat Sci Edn), 2010,25(1):40-43 (in Chinese with English abstract). [本文引用: 1]
Li JQ, Zhu GL, He ZB, Zhang ZY, Jia JX, Qiao WJ, Li JX . Study on the genetic regularity of castor bean dwarf characters gene Inner Mongolia Agric Sci Technol, 2010, ( 1):54-56 (in Chinese with English abstract). [本文引用: 1]
LiuC, Lu JN, Yin XG, BiC, Wen DY, ZhengJ, LiuS, Shi ZX, Cheng YX . Genetic analysis of traits related to plant height in Ricinus communis L. based on QTL mapping. Acta Agron Sin, 2014,40:751-759 (in Chinese with English abstract). [本文引用: 3]
Zhang ZY, NiN, WangJ, Zhu GL, MoD L T, QiaoW J, JiaJ X, HeZ B, GanM J . Major gene plus poly-gene inheritance analysis of the main raceme flowing period in castor (Ricinus communis L.). Inner Mongolia Agric Sci Technol, 2015,43(1):10-12 (in Chinese with English abstract). [本文引用: 1]
Zhang YM, Gai JY . The IECM algorithm for estimation of component distribution parameters in segregating analysis of quantitative traits Acta Agron Sin, 2000,26:699-705 (in Chinese with English abstract). [本文引用: 1]
Wang CW, Gai JY, Fu SX, Yu DY, Chen SY . Inheritance and QTL mapping of tofu and soymilk output in soybean Sci Agric Sin, 2008,41:1274-1282 (in Chinese with English abstract). [本文引用: 1]
 ,2,*, 张启好2
,2,*, 张启好2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}