 ,1,2,*, 马胜1,2, 李建武1,2, 黄伟1,2
,1,2,*, 马胜1,2, 李建武1,2, 黄伟1,2Effects of over-expression of AtDREB1A gene on potato growth and abiotic stress resistance gene expression
JIA Xiao-Xia1,2, QI En-Fang1,2, LIU Shi1,2, WEN Guo-Hong,1,2,*, MA Sheng1,2, LI Jian-Wu1,2, HUANG Wei1,2通讯作者:
收稿日期:2018-12-11接受日期:2019-04-15网络出版日期:2019-05-13
| 基金资助: |
Received:2018-12-11Accepted:2019-04-15Online:2019-05-13
| Fund supported: |
作者简介 About authors
E-mail:jiaxx0601@163.com。

摘要
关键词:
Abstract
Keywords:
PDF (3319KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
贾小霞, 齐恩芳, 刘石, 文国宏, 马胜, 李建武, 黄伟. AtDREB1A基因过量表达对马铃薯生长及抗非生物胁迫基因表达的影响 [J]. 作物学报, 2019, 45(8): 1166-1175. doi:10.3724/SP.J.1006.2019.84166
JIA Xiao-Xia, QI En-Fang, LIU Shi, WEN Guo-Hong, MA Sheng, LI Jian-Wu, HUANG Wei.
转录因子DREB1A/CBF3可以调控一系列与干旱、高盐和低温胁迫相关的功能基因的表达[1,2], 将DREB1A/CBF3基因转入农作物可以明显提高受体作物的抗逆性[3,4,5,6]。目前, 研究者已将DREB1A/CBF3基因通过转基因技术导入马铃薯的不同品种中, 并进行了相关的研究。许昆朋[7]将拟南芥CBF3基因转化马铃薯鲁引1号块茎, 对温室生长6周的转基因植株光合生理指标及RT-PCR的测定分析表明, 转基因马铃薯中CBF3的表达可以直接或间接地诱导多种抗氧化胁迫基因的表达, 以此来提高对低温胁迫的抗性。Dou等[8]以AtCBF3转基因马铃薯为试验材料, 研究AtCBF3基因表达与高温逆境的相关性。通过对温室生长6周的植株抗逆相关生理指标的测定及qRT-PCR分析表明, 高温可以诱导转基因植株中AtCBF3的表达, AtCBF3基因的表达明显提高了抗氧化基因(StGLDH、StGMpase、StDHARl和StDHAR2)、光合基因(iStaldolase、StsFBPase)以及蔗糖和淀粉合成基因(StSPS、StADPase)的表达量, 明显提高了转基因马铃薯的耐热性。本课题组通过农杆菌介导法将AtDREB1A 基因导入陇薯系列马铃薯中[9,10], 以不同浓度的PEG溶液模拟不同程度的干旱胁迫条件, 通过对试管苗的电导率, 超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)和抗坏血酸过氧化物酶(ASP)活性的测定及转基因植株中AtDREB1A基因的表达情况分析表明, AtDREB1A基因的转入提高了抗氧化酶活性, 增强了转基因马铃薯试管苗在干旱环境下的耐受能力。多项研究表明, AtDREB1A/CBF3基因的转入, 明显提高了马铃薯的抗逆性, 但关于AtDREB1A/CBF3提高马铃薯抗逆机制的研究主要集中在生理学和个别基因的表达变化层面, 不能全面分析其调控机制, 研究结果具有一定的局限性。
本研究在严格控水条件下, 试图在明确AtDREB1A基因过量表达对马铃薯植株表型、叶片相对含水量、丙二醛含量和抗氧化酶活性影响效应的基础上, 掌握马铃薯叶片抗非生物胁迫相关基因对AtDREB1A基因过量表达的响应效应, 为解析AtDREB1A基因过量表达对马铃薯生长和基因表达的影响机制, 进一步为阐明AtDREB1A提高马铃薯抗旱性的机制奠定基础。
1 材料与方法
1.1 试验材料与处理
以马铃薯品种陇薯3号(L3)及其转AtDREB1A基因株系T2为试验材料, 于2017年4月25日将统一繁殖且大小一致的各株系微型薯以同样的深度播种于试验盆(31 cm × 27 cm)中, 每盆1株, 每个株系种24盆, 置抗旱棚中常规管理, 待马铃薯出苗并生长到盛花期(7月14日)时, 选择大小和长势均匀一致的盆栽马铃薯植株12盆, 并分为2组, 每组6盆, 一组正常浇水, 一组进行水分胁迫, 胁迫时保持盆土含水量为田间最大持水量(FWC)的45%~50%, 试验期间采用土壤水分仪和称重法监测土壤水分, 及时补水, 保证土壤含水量处于设定的干旱胁迫范围。胁迫20 d后, 于8月3日早晨10:00-11:00统一选取同一层上部叶片带回实验室, 用于生理指标测定、AtDREB1A基因表达情况和转录组分析, 取样时, 各指标均为3次重复。1.2 生理指标测定
用硫代巴比妥酸法[11]测定丙二醛(MDA)含量, 氮蓝四唑(NBT)光化还原法[11]测定超氧化物歧化酶(SOD)活性, 愈创木酚比色法[11]测定过氧化物酶(POD)活性, 称重法[12]测定叶片相对含水量(RWC)。1.3 转录组测序
采集2个马铃薯材料上部相同部位叶片在液氮中速冻4 h, 然后保存于-80℃备用。使用Qiagen的RNeasy Plant MiniKit提取总RNA。分别采用Nanodrop、Qubit 2.0和Aglient 2100检测RNA的纯度、浓度和完整性, 检测合格后用于后续试验。用oligdit磁珠富集mRNA, 以此为模板合成cDNA, 纯化后进行PCR扩增, 得到最终的文库。库检合格后, 委托北京百迈客生物信息科技有限公司用HiSeq 2500进行高通量测序。1.4 差异表达基因的筛选、功能注释和富集分析
为了保证信息分析质量, 必须将含有带接头的、低质量的Raw reads过滤掉, 得到Clean reads, 后续分析都基于Clean reads。采用Bowtie软件将各样品测序得到的Reads与参考序列比对, 根据比对结果, 结合RSEM估计表达量水平。使用FPKM (Fragments Per Kilobase of transcript per Million mapped reads)法计算基因的表达量, 找出不同样本之间差异表达的基因(FDR<0.01且|Fold Change|≥2)。将Unigene在相应的数据库(nr、Swiss-Prot、KEGG和COG)中进行序列比对, 获得该Unigene序列对应的注释信息。利用topGO软件对注释到GO数据库的差异表达基因(differentially expressed genes, DEGs)进行富集分析。1.5 实时荧光定量PCR
为分析AtDREB1A基因表达情况、验证RNA- Seq结果的可靠性, 以qRT-PCR检测AtDREB1A基因和6个转录组鉴定出的上、下调表达基因。用RNA simple Total RNA Kit (TIANGEN)试剂盒提取胁迫后转基因植株和非转基因对照上部相同部位叶片总RNA, 采用First strand cDNA Synthesis Kit (THERMO)试剂盒反转录成cDNA。以反转录的cDNA为模板, 用ABIPRISMR 7300实时荧光定量PCR仪(ABI, USA)进行实时荧光定量分析, 试验设3次重复。反应体系为25 μL, 按照SYBR Premix Ex Taq (TaKaRa)反应系统, 以马铃薯EF1α基因为内参, 用AtDREB1A基因特异引物和DEGs序列引物(表1)进行扩增, 反应条件为95℃预变性3 min; 95℃变性10 s, 56.9℃退火30 s, 40个循环。用Rotor-Gene软件分析CT值, 采用2-ΔΔCT计算差异表达水平。Table 1
表1
表1用于qRT-PCR表达验证的基因及其引物
Table 1
| 基因 ID Gene ID | 正向引物 Forward primer (5′-3′) | 反向引物 Reverse primer (5′-3′) |
|---|---|---|
| EF1α | ATTGGAAACGGATATGCTCCA | TCCTTACCTGAACGCCTGTCA |
| AtDREB1A | GCCGATCAGCCTGTCTCAAT | TCTGCCATATTAGCCAACAAACTC |
| PGSC0003DMG400016489 | AAATGTGTCTTTCAAGCGAA | TCCATCTTCCCACATTTACCGTA |
| PGSC0003DMG400011977 | CTTTGCCTGAGAATATTGACT | CTTTGGGATTACTACTAGCCTT |
| PGSC0003DMG400000534 | CCGACGATACAAACATATACTGA | TGAGTTCCTTCCAGCGATT |
| PGSC0003DMG402019042 | CCAACTCAGAACTGGTACGGACA | CGTAAACATCCGTTGGGACCAT |
| PGSC0003DMG400000242 | AACAGCCAGAAGAAGTTGAT | ATTCCCATCACAGGCATCTCA |
| PGSC0003DMG401018057 | AGGATCCATTAAAGTTCAAGCCA | AATCCTTGAAGAAGCCTAGCA |
新窗口打开|下载CSV
1.6 数据统计
用 Microsoft Excel 2010软件整理数据, 用DPS V3.01软件进行单因素方差(One-way analysis, ANOVA)统计分析, Duncan’s法分析试验结果的差异显著性。所有数据均为“各重复的平均值±标准差”。2 结果与分析
2.1 干旱胁迫下马铃薯的生长状况
在正常浇水条件下, 转基因植株T2与对照L3均生长状态良好且大致相同, 表明AtDREB1A基因的转入并未对马铃薯的生长发育造成不良影响。盛花期开始水分胁迫, 20 d时对照L3植株上部和中心叶片明显萎焉卷曲、基部叶片发黄枯萎(图1-A), 而转基因植株T2除基部极个别叶片黄化外, 基本保持良好的生长状态(图1-B), 表明T2对干旱的耐受性比L3强。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1干旱胁迫下转基因及其对照马铃薯表型分析
Fig. 1Phenotypes of transgenic and no-transgenic potato under drought stress
A: 未转基因植株L3; B: 转基因植株T2。A: no-transgenic plant L3; B: transgenic plant T2.
2.2 干旱胁迫下超氧化物歧化酶(SOD)和过氧化物酶(POD)活性的变化
在正常浇水条件下, 转基因材料T2和对照L3叶片中SOD和POD活性较低, 并且各株系差异不显著。胁迫20 d后, 各株系的SOD和POD活性均增强, 且T2的2种酶活性显著高于对照L3 (P<0.05) (图2)。表明在遭受干旱胁迫时, AtDREB1A过表达可提高抗氧化酶的活性, 增强植株的抗旱性。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2干旱胁迫下马铃薯叶片SOD和POD活性的变化
相同处理标以不同字母的柱值在P < 0.05水平上差异显著。
Fig. 2Changes of SOD and POD activities in potato leaves under drought stress
Bars superscripted by different letters are significantly different at P < 0.05 within the same treatment.
2.3 干旱胁迫下丙二醛含量(MDA)和叶片相对含水量(RWC)的变化
在正常浇水条件下, 转基因材料T2和对照L3叶片中MDA含量较低, 并且各株系差异不显著。胁迫20 d后, 各株系的MDA含量明显上升, 但转基因材料T2的MDA含量显著低于对照L3 (P<0.05)(图3-A)。在正常浇水条件下, 转基因材料T2和对照L3叶片RWC无显著差异, 均在93%以上。水分胁迫20 d后, 各株系的叶片RWC都降低, 但转基因材料T2的降低幅度小于对照L3, 且T2植株的RWC (85.31%)显著高于对照L3 (79.20%)(P<0.05)(图3-B)。
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3干旱胁迫下马铃薯叶片MDA含量和RWC的变化
相同处理标以不同字母的柱值在 P < 0.05 水平上差异显著。
Fig. 3Changes of MDA content and relative water content in potato leaves under drought stress
Bars superscripted by different letters are significantly different at P < 0.05 within the same treatment.
2.4 干旱胁迫下AtDREB1A基因表达分析
qRT-PCR分析结果表明, 在正常浇水条件下, 转基因材料T2和对照L3叶片中AtDREB1A基因相对表达量差异不显著, 差异倍数仅为2.34。胁迫20 d后, 转基因材料T2叶片中AtDREB1A的相对表达量显著高于对照L3, 差异倍数高达37.56 (P<0.05)(图4)。图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4马铃薯叶片中AtDREB1A基因表达的实时荧光定量分析
相同处理标以不同字母的柱值在 P < 0.05 水平上差异显著。
Fig. 4qRT-PCR analysis on AtDREB1A gene in potato leaves
Bars superscripted by different letters are significantly different at P < 0.05 within the same treatment.
2.5 干旱胁迫下马铃薯叶片差异表达基因分析
由图5可知, 干旱胁迫下, 2个样品的基因表达水平存在一定的差异。转基因材料T2相对于对照L3的差异表达基因为430个, 其中上调表达基因287个, 下调表达基因143个。功能注释和显著性富集结果表明, 这些差异表达基因富集涉及GO功能分类体系中生物过程、细胞组分和分子功能3个大类别, 且大部分差异表达基因集中在细胞内和膜上, 主要涉及信号传导、氧化还原、生物调解、应激反应、发育过程、系统免疫过程、核酸和蛋白结合转录因子活性、转运活性及催化活性(图6)。其中非生物胁迫相关基因主要涉及PPR蛋白家族、细胞色素P450蛋白家族、热激蛋白(HSP)家族和MLO蛋白家族。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5干旱胁迫后马铃薯叶片中差异基因表达量火山图
横坐标代表基因在不同样品中表达倍数变化; 纵坐标代表基因表达量变化的统计学显著程度。图中每个基因均由一个点表示, 黑色圆点表示无显著性差异的基因, 左侧绿色圆点表示有显著性差异的下调基因, 右侧红色圆点表示有显著性差异的上调基因。
Fig. 5Volcano plot of gene expression in potato leaves after drought stress
The abscissa shows the log2 (fold change) between the two samples; the ordinate shows the statically significance of the difference. Each gene is represented by one point on the graph. Differential genes with no significant difference were represented by black dots, those left green points and right red points represent significantly different down-regulated and up-regulated genes, respectively.
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6差异表达基因GO注释聚类图
横坐标为GO各分类内容, 纵坐标左边为基因数目所占百分比, 右边为基因数目。
Fig. 6Differentially expressed gene GO annotation clustering map
The abscissa is the content of each category of GO. The left side of the ordinate is the percentage of the gene number, and the right side is the gene number.
2.6 干旱胁迫下马铃薯叶片中抗非生物胁迫基因表达情况
2.6.1 差异表达的PPR蛋白家族基因 PPR (Pentatricopeptide repeat)蛋白家族是植物中最大的基因家族之一, 直接或间接参与植物抵抗低温、干旱等逆境胁迫。本研究中, 共有9个PPR蛋白家族基因表达量上调(图7), 其中基因PGSC0003DMG4 00016489、PGSC0003DMG400012057和PGSC0003 DMG400006026上调倍数较大, 分别为37.39、25.93和14.11倍; PGSC0003DMG400005692、PGSC0003 DMG400026708、PGSC0003DMG402025785和PGSC 0003DMG400011199上调倍数次之, 分别为9.40、9.14、8.53和6.19倍; PGSC0003DMG401003570和PGSC0003DMG400031153上调倍数最小且差异不大, 分别为4.99和4.47。2.6.2 差异表达的HSP蛋白家族基因 热激蛋白(heat shock protein, HSP)又称热休克蛋白或应激蛋白, 在生物体的抗逆反应中起着非常重要的作用。本研究中, 共有13个HSP家族基因表达量上调(图8), 其中PGSC0003DMG401028907编码HSP90, 上调6.22倍, PGSC0003DMG400014835编码HSP70, 上调4.10倍; 其余11个均为HSP20蛋白相关基因, 表达量上调倍数的顺序为PGSC0003DMG 400024476 (17.86) > PGSC0003DMG400003219 (14.91) > PGSC0003DMG400030340 (10.52) > PGSC 0003DMG400009255 (9.64) > PGSC0003DMG4000 28624 (9.18) > PGSC0003DMG400012619 (7.22) > PGSC0003DMG400002928 (6.58) > PGSC0003DMG 400030339 (6.24) > PGSC0003DMG400017098 (6.21) > PGSC0003DMG400030427 (5.50) > PGSC0003 DMG400011977 (4.54)。
图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7干旱胁迫下PPR蛋白家族基因表达模式聚类图
图中连线表示各个基因变化趋势的相近程度, 线越短越相近。红色表示上调表达水平, 绿色表示下调表达水平。
Fig. 7Cluster diagram of gene expression patterns of PPR protein family under drought stress
The lines in graph show the degree of similarity in the variation trend of each gene, and the shorter the line is, the more similar the genes are. Red indicates the up-regulated expression level, while green indicates the down-regulated expression level.
图8
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8干旱胁迫下HSP蛋白家族基因表达模式聚类图
图中连线表示各个基因变化趋势的相近程度, 线越短越相近。红色表示上调表达水平, 绿色表示下调表达水平。
Fig. 8Cluster diagram of gene expression patterns of HSP protein family under drought stress
The lines in graph show the degree of similarity in the variation trend of each gene, and the shorter the line is, the more similar the genes are. Red indicates the up-regulated expression level, while green indicates the down-regulated expression level.
2.6.3 差异表达的P450蛋白家族基因 细胞色素P450家族蛋白在植物抗病虫防卫反应和抗非生物胁迫中都具有很重要的作用。本研究中, 共有5个编码细胞色素P450家族蛋白基因表达量上调(图9), 其中PGSC0003DMG400000565上调了271.06倍, PGSC0003DMG400000534上调了19.09倍, PGSC 0003DMG400000148、PGSC0003DMG40000756和PGSC0003DMG400028096分别上调了9.11、5.41和4.39倍。
图9
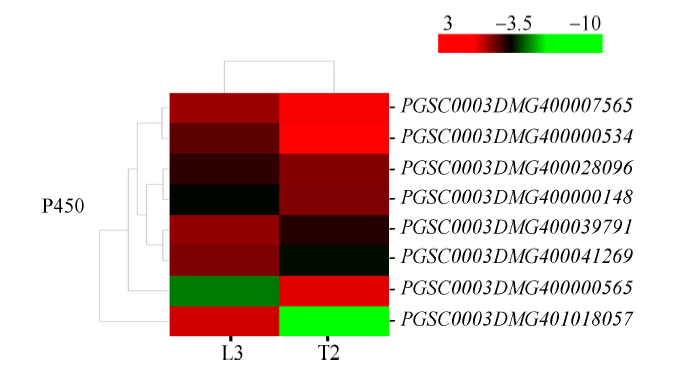 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9干旱胁迫下P450蛋白家族基因表达模式聚类图
图中连线表示各个基因变化趋势的相近程度, 线越短越相近。红色表示上调表达水平, 绿色表示下调表达水平。
Fig. 9Cluster diagram of gene expression patterns of P450 protein family under drought stress
The lines in graph show the degree of similarity in the variation trend of each gene, and the shorter the line is, the more similar the genes are. Red indicates the up-regulated expression level, while green indicates the down-regulated expression level.
2.6.4 差异表达的MLO蛋白家族基因 MLO是高等植物特有的一类抗病家族, 在响应非生物胁迫中发挥重要作用。本研究中, 共有3个MLO蛋白家族基因表达量上调(图10), 其中PGSC0003DMG 400020605上调12.43倍、PGSC0003DMG400030134上调9.13倍、PGSC0003DMG400008112上调7.52倍。
图10
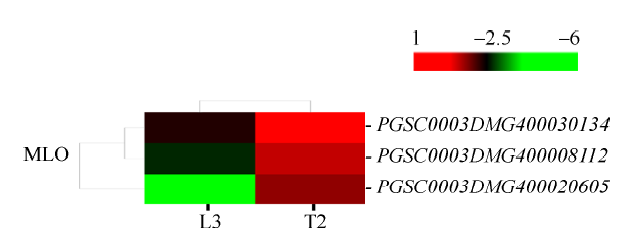 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图10干旱胁迫下MLO蛋白家族基因表达模式聚类图
图中连线表示各个基因变化趋势的相近程度, 线越短越相近。红色表示上调表达水平, 绿色表示下调表达水平。
Fig. 10Cluster diagram of gene expression patterns of MLO protein family under drought stress
The lines in graph show the degree of similarity in the variation trend of each gene, and the shorter the line is, the more similar the genes are. Red indicates the up-regulated expression level, while green indicates the down-regulated expression level.
2.7 差异表达基因的qRT-PCR验证
随机选取6个差异表达基因进行qRT-PCR验证。其中PGSC0003DMG400016489、 PGSC0003DM G400011977和PGSC0003DMG400000534为上调表达基因, PGSC0003DMG402019042、PGSC0003DMG 400000242和 PGSC0003DMG401018057为下调表达基因。以马铃薯EF1α为内参基因, 计算6个目的基因的相对表达量, 并与DGE 测序分析的结果比较。DGE测序分析中表达量上调的3个基因, 在胁迫后叶片中相对表达量均较对照上升; 而在DGE测序分析中表达量下调的3个基因, 在胁迫后叶片中相对表达量均低于对照, 即qRT-PCR 中6个基因表达量的变化趋势与测序分析结果相一致(图11), 证明RNA-seq测序结果具可靠性。2种分析方法检测的6个差异基因的表达变化趋势相同, 但表达量存在一定差异, 这可能由2种检测方法的检测范围和灵敏度不同所致。图11
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图11干旱胁迫下L3 vs. T2差异表达基因的qRT-PCR验证
Fig. 11Verification of DEGs between L3 vs. T2 after drought stress using qRT-PCR
3 讨论
当处于逆境时, 植物不仅会在形态和生理上产生适应性反应, 同样会通过调节基因表达来适应新的不良环境[13]。DREB/CBF由逆境胁迫诱导后, 在转基因植物中的过量表达可以激活其他一系列抗逆功能基因同时表达, 从而增强植物对干旱、低温及高盐等逆境的抗性[7-8,14]。本研究对盛花期的马铃薯进行控水处理, 将土壤含水量控制在田间最大持水量的45%~50%, 胁迫处理20 d时, 对照L3植株的叶片萎蔫卷曲及黄化程度比AtDREB1A基因过表达植株T2严重的多, 说明AtDREB1A基因过表达明显增强了马铃薯对干旱的耐受性。RWC能体现植株叶片水势, 是评价植株抗旱性的重要指标, 受到干旱胁迫后, 跟耐旱性差的植株相比, 耐旱性强的植株能保持较高的RWC[15,16]。本研究中, 在正常浇水条件下, 转基因材料T2和对照L3叶片RWC无显著差异; 水分胁迫20 d后, 各株系的叶片RWC都降低, 但转基因材料T2显著高于对照L3, 说明转基因植株耐旱性较对照强。逆境胁迫会加速植物体内活性氧的积累, 过多的活性氧会对细胞膜脂造成严重的损伤。MDA含量是评价叶片细胞质膜受干旱胁迫损伤程度的重要生理指标[17]。植物细胞内的酶促和非酶促体系形成一系列反应机制来清除活性氧, 从而维持整个细胞防御系统的动态平衡。作为酶促体系的重要成员, SOD和POD常被用来衡量逆境胁迫条件下细胞活性氧清除能力的大小[18]。本研究发现, 在干旱胁迫条件下, 转基因植株的丙二醛含量显著低于对照, 抗氧化保护酶SOD和POD活性均显著高于对照, 表明AtDREB1A基因过表达, 提高了抗氧化保护酶SOD和POD活性, 有效地增强了细胞清除活性氧的能力, 进而减轻了活性氧对质膜造成的伤害, 这与过表达AtDREB1A植株受干旱影响程度较轻相一致。通过转录组测序及生物信息学方法分析转基因马铃薯与未转基因对照在转录水平上基因表达的差异, 发现转基因材料T2相对于L3的差异表达基因有430个, 其中上调表达基因287个, 下调表达基因143个。上调表达基因中与抗非生物胁迫相关的基因主要包括PPR蛋白家族、HSP蛋白家族、P450蛋白家族和MLO蛋白家族基因。PPR蛋白家族不仅在RNA的编辑加工、细胞器形成、细胞核与细胞器之间信号传递、恢复细胞质雄性不育和调节植物生长发育方面发挥重要作用, 而且直接或间接参与植物抗逆境胁迫, 在马铃薯抗旱[19]、水稻抗低温[20]、水稻和毒麦抗盐[21,22]、拟南芥抗氧化[23,24]中均发挥着重要作用。HSP是干旱胁迫物质形成的必需要素, 通过提高自身的表达量增强植物对胁迫的适应能力, 维持胁迫调节下蛋白的稳定性和维持植物的生存与正常生长[25,26]。细胞色素P450家族蛋白在植物抗病虫防卫反应和抗非生物胁迫中都具有非常重要的作用。崔会婷等[27]研究发现, 模拟干旱胁迫条件下, 苜蓿MtCYP450基因表达量明显上调, 转MtCYP450基因拟南芥比野生型表现出较好的耐旱性, 表明MtCYP450基因与干旱胁迫的调节机制有很大的相关性。MLO是高等植物特有的一类抗病家族, 在响应非生物胁迫中也发挥着重要作用, 很多MLO基因的表达会受到各种生物和非生物胁迫的影响[28,29]。辣椒CaMLO2基因的表达受到脱落酸和干旱的诱导, 说明CaMLO2基因与干旱胁迫有关[30]。本研究中, 相对于未转基因对照L3, 转基因材料T2中抗非生物胁迫相关蛋白PPR、HSP、P450和MLO等家族基因表达量上调幅度较大, 上调倍数最低的都不低于4, 大部分都上调十几倍甚至几十倍, 说明这些基因在转AtDREB1A基因马铃薯抵御干旱过程中发挥着非常重要的作用。 本研究为进一步解析AtDREB1A基因提高马铃薯抗旱性的调控网络奠定了基础。
4 结论
在增强细胞清除活性氧能力、减轻活性氧对质膜伤害程度的同时, AtDREB1A基因过量表达引起430个基因表达变化, 其中上调表达基因287个, 下调表达基因143个, 主要涉及信号传导、氧化还原、生物调解、应激反应、发育过程、系统免疫过程、核酸和蛋白结合转录因子活性、转运活性及催化活性, 其中抗非生物胁迫相关蛋白PPR、HSP、P450和MLO等家族的大量基因表达量发生较大的变化, 说明这些基因在转AtDREB1A基因马铃薯抵御干旱过程中发挥着非常重要的作用。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1023/A:1009652326121URL [本文引用: 1]
Magsci [本文引用: 1]

<p>以小麦品种H6756和藁城8901作为基因枪转化的靶材料,取其护颖至雌雄蕊原基形成期的幼穗,用含逆境诱导转录因子DREB1A和bar基因的质粒pAHC25轰击胚性愈伤组织,在分别含有5mgL和10mgLBasta溶液的培养基上进行筛选。得到的抗性愈伤组织在不含Basta溶液的培养基上再生培养,获得218棵再生植株。田间涂抹浓度为100mgL的Basta溶液检测后,对抗性植株作PCR检测,获得54棵再生植株。通过对其中20株T1代的PCR和Southern杂交分析,已获得14株含DREB1A和bar基因的转基因小麦植株,其中H675613株,藁城89011株。</p>
Magsci [本文引用: 1]
<p>以小麦品种H6756和藁城8901作为基因枪转化的靶材料,取其护颖至雌雄蕊原基形成期的幼穗,用含逆境诱导转录因子DREB1A和bar基因的质粒pAHC25轰击胚性愈伤组织,在分别含有5mgL和10mgLBasta溶液的培养基上进行筛选。得到的抗性愈伤组织在不含Basta溶液的培养基上再生培养,获得218棵再生植株。田间涂抹浓度为100mgL的Basta溶液检测后,对抗性植株作PCR检测,获得54棵再生植株。通过对其中20株T1代的PCR和Southern杂交分析,已获得14株含DREB1A和bar基因的转基因小麦植株,其中H675613株,藁城89011株。</p>
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
DOI:10.11686/cyxb20140312Magsci [本文引用: 1]
马铃薯在生长发育过程中受各种生物和非生物胁迫的影响,干旱是其中最常见和危害最严重的非生物胁迫因素之一,常常导致单产不高,总产不稳,严重影响着马铃薯产业的发展。本研究以改善马铃薯抗旱性为目的,采用PCR方法从拟南芥中克隆了转录因子DREB1A基因和诱导型启动子rd29A。利用DNA重组技术成功构建了诱导型启动子rd29A驱动转录因子DREB1A基因和CaMV35S启动子驱动Bar基因的双价植物表达载体pBI121-rd29-BDR,并通过农杆菌介导法对马铃薯进行遗传转化,PPT筛选得到22株抗性苗,PCR和RT-PCR检测证明DREB1A基因已整合到陇薯10号马铃薯基因组中并在转基因植株中转录表达,有望提高转基因马铃薯的抗旱性。目前,作者正在进行转基因马铃薯的抗旱性分析研究。
DOI:10.11686/cyxb20140312Magsci [本文引用: 1]
马铃薯在生长发育过程中受各种生物和非生物胁迫的影响,干旱是其中最常见和危害最严重的非生物胁迫因素之一,常常导致单产不高,总产不稳,严重影响着马铃薯产业的发展。本研究以改善马铃薯抗旱性为目的,采用PCR方法从拟南芥中克隆了转录因子DREB1A基因和诱导型启动子rd29A。利用DNA重组技术成功构建了诱导型启动子rd29A驱动转录因子DREB1A基因和CaMV35S启动子驱动Bar基因的双价植物表达载体pBI121-rd29-BDR,并通过农杆菌介导法对马铃薯进行遗传转化,PPT筛选得到22株抗性苗,PCR和RT-PCR检测证明DREB1A基因已整合到陇薯10号马铃薯基因组中并在转基因植株中转录表达,有望提高转基因马铃薯的抗旱性。目前,作者正在进行转基因马铃薯的抗旱性分析研究。
[本文引用: 1]
.
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.11686/cyxb20100205Magsci [本文引用: 1]
<FONT face=Verdana>为了进一步明确冷季型草坪草形态、水分生理和碳分配模式对干旱及复水处理的反应,本试验以草地早熟禾草的3个栽培种:新歌来德、午夜和奖金为材料,进行人工模拟干旱处理21d,然后再恢复浇水21d。期间测定了草坪质量、叶片相对含水量(RWC)、总的非结构性碳水化合物(TNC)含量及<SUP>14</SUP>C 同化物在根、茎、叶的分配率。试验结果表明,干旱胁迫造成3个品种的草坪质量和RWC 明显下降,变化幅度因品种和处理时间不同而存在差异;复水后,草坪质量和RWC 不同程度地得到恢复,其中奖金表现出较强的恢复能力。干旱胁迫初期,3 个品种根部<SUP>14</SUP>C分配率明显增加,干旱14d 时,3个品种<SUP>14</SUP>C分配模式为:根>茎>叶。随着干旱胁迫的进行,<SUP>14</SUP>C由根部向茎和叶片转移,其茎和叶片中<SUP>14</SUP>C 分配率明显升高。复水21d后,3个品种中<SUP>14</SUP>C分配率模式为:茎> 叶> 根;干旱胁迫期间,3个品种的叶片、茎和根之间TNC 积累量一直发生变化,干旱胁迫21d时,3个品种TNC 含量在根、茎、叶中的积累模式为:叶>茎>根。复水后,3个品种根、茎、叶中TNC 积累量较干旱胁迫21d时各自对应器官中TNC积累量有不同程度下降,但TNC 在根、茎、叶中的积累模式与干旱胁迫期间保持一样。试验结果表明,在干旱和复水处理中碳同化物在植物体内不同器官不同分配率及碳水化合物在植物体内不同器官中积累是草坪草对干旱胁迫及复水处理的一种生理调节反应,将为今后研究草坪草在其他逆境胁迫下的抗性提供了一定的科学参考。</FONT>
DOI:10.11686/cyxb20100205Magsci [本文引用: 1]
<FONT face=Verdana>为了进一步明确冷季型草坪草形态、水分生理和碳分配模式对干旱及复水处理的反应,本试验以草地早熟禾草的3个栽培种:新歌来德、午夜和奖金为材料,进行人工模拟干旱处理21d,然后再恢复浇水21d。期间测定了草坪质量、叶片相对含水量(RWC)、总的非结构性碳水化合物(TNC)含量及<SUP>14</SUP>C 同化物在根、茎、叶的分配率。试验结果表明,干旱胁迫造成3个品种的草坪质量和RWC 明显下降,变化幅度因品种和处理时间不同而存在差异;复水后,草坪质量和RWC 不同程度地得到恢复,其中奖金表现出较强的恢复能力。干旱胁迫初期,3 个品种根部<SUP>14</SUP>C分配率明显增加,干旱14d 时,3个品种<SUP>14</SUP>C分配模式为:根>茎>叶。随着干旱胁迫的进行,<SUP>14</SUP>C由根部向茎和叶片转移,其茎和叶片中<SUP>14</SUP>C 分配率明显升高。复水21d后,3个品种中<SUP>14</SUP>C分配率模式为:茎> 叶> 根;干旱胁迫期间,3个品种的叶片、茎和根之间TNC 积累量一直发生变化,干旱胁迫21d时,3个品种TNC 含量在根、茎、叶中的积累模式为:叶>茎>根。复水后,3个品种根、茎、叶中TNC 积累量较干旱胁迫21d时各自对应器官中TNC积累量有不同程度下降,但TNC 在根、茎、叶中的积累模式与干旱胁迫期间保持一样。试验结果表明,在干旱和复水处理中碳同化物在植物体内不同器官不同分配率及碳水化合物在植物体内不同器官中积累是草坪草对干旱胁迫及复水处理的一种生理调节反应,将为今后研究草坪草在其他逆境胁迫下的抗性提供了一定的科学参考。</FONT>
DOI:10.11686/cyxb20120116Magsci [本文引用: 1]
通过长期继代(24个月)假俭草胚性愈伤组织,诱发体细胞无性系变异的发生,从再生植株中筛选抗旱变异体,获得了5个变异体植株。生理检测结果表明,干旱处理后所有植株叶片的相对含水量下降,相对电导率和丙二醛(MDA)含量升高,但抗旱变异体植株的相对含水量显著高于对照植株,相对电导率和MDA 含量显著低于对照植株,表明变异体植株受干旱伤害程度较轻,提高了抗旱性。干旱处理前变异体植株与其对照植株间脯氨酸含量、SOD 活性和CAT 活性差异不大,干旱处理后所有植株的脯氨酸、SOD 活性和CAT 活性均升高,但变异体植株的脯氨酸含量低于对照植株,SOD 和CAT 活性高于对照植株。结果表明,变异体植株的抗旱性与在干旱条件下维持更高的SOD 和CAT 活性有关。
DOI:10.11686/cyxb20120116Magsci [本文引用: 1]
通过长期继代(24个月)假俭草胚性愈伤组织,诱发体细胞无性系变异的发生,从再生植株中筛选抗旱变异体,获得了5个变异体植株。生理检测结果表明,干旱处理后所有植株叶片的相对含水量下降,相对电导率和丙二醛(MDA)含量升高,但抗旱变异体植株的相对含水量显著高于对照植株,相对电导率和MDA 含量显著低于对照植株,表明变异体植株受干旱伤害程度较轻,提高了抗旱性。干旱处理前变异体植株与其对照植株间脯氨酸含量、SOD 活性和CAT 活性差异不大,干旱处理后所有植株的脯氨酸、SOD 活性和CAT 活性均升高,但变异体植株的脯氨酸含量低于对照植株,SOD 和CAT 活性高于对照植株。结果表明,变异体植株的抗旱性与在干旱条件下维持更高的SOD 和CAT 活性有关。
DOI:10.11686/cyxb20100411Magsci [本文引用: 1]
<p><font face="Verdana">以中苜1号为对照,对来自俄罗斯的18份紫花苜蓿种质的耐盐性进行了综合评价,并对盐胁迫下的生理反应进行了初步研究。结果表明,运用标准差系数赋予权重法进行综合评价,不但考虑了不同指标的权重,还定量的鉴定出了每份材料的耐盐能力,比聚类分析的结果更具科学合理性,试验共筛选出M7、M9、M15、810共4份耐盐性强的种质;在此基础上,进一步探讨了盐胁迫下的生理反应,除叶水势随着盐浓度的增加呈下降趋势外,游离脯氨酸含量、可溶性糖含量、细胞质膜透性、丙二醛含量、水分饱和亏缺则呈上升趋势,且不同材料变化幅度不同。可溶性糖含量、细胞质膜透性、丙二醛含量、水分饱和亏缺和叶水势等指标很好的反映了材料间的耐盐性,可直接作为耐盐评价的鉴定指标。</font></p>
DOI:10.11686/cyxb20100411Magsci [本文引用: 1]
<p><font face="Verdana">以中苜1号为对照,对来自俄罗斯的18份紫花苜蓿种质的耐盐性进行了综合评价,并对盐胁迫下的生理反应进行了初步研究。结果表明,运用标准差系数赋予权重法进行综合评价,不但考虑了不同指标的权重,还定量的鉴定出了每份材料的耐盐能力,比聚类分析的结果更具科学合理性,试验共筛选出M7、M9、M15、810共4份耐盐性强的种质;在此基础上,进一步探讨了盐胁迫下的生理反应,除叶水势随着盐浓度的增加呈下降趋势外,游离脯氨酸含量、可溶性糖含量、细胞质膜透性、丙二醛含量、水分饱和亏缺则呈上升趋势,且不同材料变化幅度不同。可溶性糖含量、细胞质膜透性、丙二醛含量、水分饱和亏缺和叶水势等指标很好的反映了材料间的耐盐性,可直接作为耐盐评价的鉴定指标。</font></p>
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1074/mcp.M500251-MCP200URL [本文引用: 1]
[本文引用: 1]
DOI:10.1093/jxb/erj098URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1002/(ISSN)1615-9861URL [本文引用: 1]
DOI:10.1016/j.tplants.2004.03.006URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}