 ,*山西农业大学农学院, 山西太谷 030801
,*山西农业大学农学院, 山西太谷 030801Allohexaploid synthesis and its characteristic identification between cotton species Gossypium hirsutum and G. sturtianum
SHEN Zhuang-Zhuang, LI Yu-Ying, RONG Er-Hua, WU Yu-Xiang,*College of Agriculture, Shanxi Agricultural University, Taigu 030801, Shanxi, China通讯作者:
收稿日期:2018-06-22接受日期:2019-01-12网络出版日期:2019-02-03
| 基金资助: |
Received:2018-06-22Accepted:2019-01-12Online:2019-02-03
| Fund supported: |
作者简介 About authors

摘要
关键词:
Abstract
Keywords:
PDF (7739KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
申状状, 李昱樱, 荣二花, 吴玉香. 陆地棉和野生斯特提棉种间异源六倍体的合成与性状鉴定[J]. 作物学报, 2019, 45(4): 628-634. doi:10.3724/SP.J.1006.2019.84086
SHEN Zhuang-Zhuang, LI Yu-Ying, RONG Er-Hua, WU Yu-Xiang.
远缘杂交伴随着多倍化是新物种形成的重要途径[1,2]。棉花的远缘杂交有利于促进种属间的基因交流, 比种内杂交更容易获得新的种质资源。种质资源是棉花育种的重要基础, 长期以来, 棉花育种家做了大量的远缘杂交研究, 希望创造出近缘杂交所无法得到的优异特性。陆地棉(G. hirsutum)是世界上重要的栽培棉种, 然而现有的栽培品种由于亲缘关系太近, 难以拓宽陆地棉的遗传基础[3]。野生棉含有栽培棉所缺乏的许多优质基因, 挖掘野生棉资源是棉花育种的一个重要方向[4]。野生斯特提棉(G. sturtianum)具有许多优良性状, 如耐低温、抗黄萎病、生长势强、纤维优质以及种子腺体延缓发育等[5,6]。将这些优良基因通过远缘杂交转移到陆地棉, 对棉花育种和生产有重要作用。1935年开始, 国外****[5,6,7]和我国的姜茹琴等[8]、李育强等[9]、崔荣霞等[10]和梁正兰等[5]先后报道了陆地棉和斯特提棉的杂交研究。这些研究证明陆地棉与斯特提棉的三倍体杂种不育, 减数分裂过程中单价体数量多, 染色体同源性低, 两亲本亲缘关系远, 通过染色体加倍和回交技术恢复其育性, 获得了新的种质资源和品种, 在棉花育种方面取得显著成就。但目前尚未获得陆地棉与斯特提棉种间染色体数完整、可以稳定遗传的异源六倍体种质, 该种质对棉花远缘杂交、染色体加倍和物种进化研究具有重要价值, 有助于更好地利用棉花野生种质资源开展育种研究。
本实验室在2015年亦报道了利用陆地棉品种中棉所12和斯特提棉进行种质创新探索的研究, 获得的杂种植株可结铃但无可育种子[6]。本研究重新选择陆地棉品种中棉所16作母本与野生斯特提棉杂交, 对杂种幼苗进行秋水仙碱染色体加倍, 杂种植株育性恢复, 此世代植株为混倍体, 倍性与性状不稳定。将收获的种子继续种植, 对可育植株进行细胞遗传学和流式细胞倍性鉴定, 对其叶片、花、纤维和种子腺体发育等性状进行初步评价, 以期筛选出染色体数完整可以稳定遗传的异源六倍体新种质。
1 材料与方法
1.1 试验材料
母本陆地棉品种中棉所16来源于中国农科院棉花研究所, 父本野生斯特提棉来源于国家种质三亚野生棉圃。1.2 试验设计
陆地棉(AADD)作母本P1, 野生斯特提棉(CC)作父本P2, 进行远缘杂交, 获得三倍体杂种F1 (ADC), 对F1幼苗进行多倍体诱变得到M0代(AADDCC)植株, 将M0代植株所得种子播种得到M1代(AADDCC)植株, 对4个不同世代P1、P2、F1和M1进行比较鉴定。所用材料均以盆栽的方式保存于山西农业大学农学院温室。1.3 试验方法
1.3.1 远缘杂交 以陆地棉为母本, 早上6:00左右选择即将开放的花蕾去雄, 套上蜡管待用。上午9:00以后, 取当天开放的父本斯特提棉的花粉给去雄的陆地棉授粉, 继续套上蜡管, 在苞叶内滴加25 mg L-1的赤霉素保铃。1.3.2 多倍体诱变 将棉种播于营养钵, 待幼苗子叶展开当天, 用0.2%秋水仙碱凝胶包裹幼苗生长点, 盖上透明罩杯保证湿度, 48 h后去除凝胶并用水冲洗干净; 待幼苗恢复生长后移栽到花盆, 冬天转移到温室保存。
1.3.3 流式细胞检测 取2 cm × 3 cm大小的幼嫩叶片, 将其切碎, 置400 μL裂解液中以提取完整细胞核, 用30 μm滤网过滤裂解液至样品管中, 加入1600 μL染液, 上机测试(CyFlow Space, Sysmex Partec)。
1.3.4 花粉母细胞减数分裂观察 选取3~5 mm大小的花蕾, 浸泡在卡诺氏固定液(无水乙醇∶氯仿∶冰乙酸 = 5∶3∶2) 24 h, 蒸馏水冲洗浸泡20 min, 滤纸吸干, 取3~5粒花药于载玻片上, 滴加卡宝品红, 用镊子将花药压破, 去除杂质, 盖上盖玻片, 轻轻压紧, 显微镜下观察。
1.3.5 气孔叶绿体计数 选取生长良好的倒四叶或倒五叶冲洗干净; 切取叶片主脉附近1 cm × 2 cm部分, 浸入卡诺氏固定液脱色处理48 h以上; 将脱色的叶片用蒸馏水冲洗浸泡10 min以上, 切取约2 mm × 2 mm叶片于载玻片上, 下表皮朝上, 用1%的I2-KI溶液染色5~10 min, 盖上盖玻片; 在显微镜下统计气孔保卫细胞中的叶绿体数, 每株统计30个气孔。
1.3.6 花粉粒直径测定 选取当天开放的花, 取3~5粒花药置载玻片上, 滴加卡宝品红, 并用镊子压碎, 盖上盖玻片, 在显微镜下测量花粉粒直径。取每株棉花2~3朵花, 共测量150个花粉粒。
1.4 数据统计分析
本研究所得数据使用Microsoft Excel和SPSS软件进行统计分析和作图。2 结果与分析
2.1 细胞学鉴定
2.1.1 流式细胞检测结果 从M1中选取6株(M1-1至M1-6)植株鉴定倍性, 以四倍体母本P1和二倍体父本P2做参照, 用流式细胞技术检测F1以及M1-1至M1-6的相对DNA含量。结果显示, P2、F1和P1的峰值分别位于98.81、122.89和140.44, 表明F1的DNA含量为P1和P2两亲本之和的一半; M1代中, M1-4和M1-5的峰值位于239.88和239.92, 表明其DNA含量接近P1与P2之和, M1-1、M1-2和M1-3的DNA含量介于三倍体和六倍体之间, M1-6的DNA含量大于六倍体(表1)。这些结果表明, F1为陆地棉和斯特提棉的真杂种, 所测M1代中有2株为异源六倍体(M1-4和M1-5), 有4株为非整倍体(M1-1、M1-2、M1-3和M1-6)。Table 1
表1
表1不同世代流式细胞倍性检测结果
Table 1
| P2 | F1 | P1 | M1-1 | M1-2 | M1-3 | M1-4 | M1-5 | M1-6 | |
|---|---|---|---|---|---|---|---|---|---|
| X-mean | 98.81 | 122.89 | 140.44 | 153.02 | 184.70 | 190.86 | 239.88 | 239.92 | 254.28 |
新窗口打开|下载CSV
2.1.2 花粉母细胞减数分裂观察 根据流式细胞检测结果, 对2株DNA含量完整的异源六倍体植株进行花粉母细胞减数分裂观察, 并与P1和F1比较。结果显示, P1减数分裂行为正常, 四分体时期正常四分体比例为100%; F1减数分裂行为异常, 主要表现在减数分裂后期I, 染色体向多极不均等分离, 并且分离不同步, 出现大量染色体桥和无着丝粒的染色体片段, 最终形成的正常四分体比例仅占4.24%; M1代异源六倍体植株的减数分裂行为已趋于正常, 在终变期可以清晰观察到39对二价体, 在减数分裂中期II, 可观察到均等分离的78条染色体, 在四分体时期, 正常四分体比例达94.5% (图1)。这些细胞学结果表明, 本研究获得了陆地棉与斯特提棉的异源六倍体新种质, 并且其减数分裂行为已趋于正常。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1花粉母细胞减数分裂表现
A: P1中期II; B: P1后期II; C: P1四分体; D: F1后期I; E: F1后期I; F: F1异常四分体; G: M1终变期; H: M1中期II; I: M1四分体。
Fig. 1Meiosis performance of pollen mother cells
A: metaphase II of P1; B: anaphase II of P1; C: tetrad of P1; D: anaphase I of F1; E: anaphase I of F1; F: abnormal tetrad of F1; G: diakinesis of M1; H: metaphase II of M1; I: tetrad of M1.
2.2 表型鉴定
2.2.1 叶片性状统计 不同世代植株的叶片大小、叶形、叶色以及叶柄颜色等表型差异大(图2)。对P1、P2、F1和M1的叶面积、叶形指数(叶长/叶宽)和叶片下表皮气孔保卫细胞的叶绿体数目(以下简称叶绿体数)统计显示, F1与M1在叶面积、叶形指数和叶绿体数均存在极显著差异(表2)。随着倍性增大, 不同世代叶形指数均呈下降趋势, 叶面积和叶绿体数目呈上升趋势, 亲本因自身基因组差异而不完全符合该变化趋势(图3)。这些结果证明六倍体叶形介于双亲之间, 叶面积增大, 保卫细胞中叶绿体数明显增多, 这对远缘杂种的光合作用等生理功能具有重要意义。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同世代叶片形态比较
Fig. 2Leaf morphology comparison among different generations
Table 2
表2
表2不同世代间叶片性状的差异显著性分析
Table 2
| 材料 Material | 倍性 Ploidy | 叶面积 Leaf area (cm2) | 叶形指数(叶长/叶宽) Leaf index (leaf length/leaf width) | 叶绿体数 Chloroplast number |
|---|---|---|---|---|
| P1 | 4 | 39.47 ± 12.32 bcBC | 0.77 ± 0.09 aA | 13.57 ± 1.59 bB |
| P2 | 2 | 16.38 ± 3.20 aA | 1.07 ± 0.06 dC | 12.92 ± 1.55 bB |
| F1 | 3 | 32.70 ± 5.82 bB | 0.93 ± 0.06 cB | 11.64 ± 1.77 aA |
| M1 | 6 | 43.97 ± 17.11 cC | 0.82 ± 0.06 bA | 19.95 ± 2.97 cC |
新窗口打开|下载CSV
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3叶面积、叶形指数和叶绿体数在不同倍性间的变化趋势
Fig. 3Trends of leaf area, leaf index and chloroplast number in different ploidies
2.2.2 花的表型性状和花粉粒直径统计 远缘杂种与亲本在花瓣颜色和大小、花斑的有无、花药颜色和数量、柱头长短以及苞叶形态等方面均存在明显差异; 在F1和M1之间, 花的表型性状整体相近, 但M1的花药数量明显少于F1(图4)。花粉粒直径频率直方图显示, P1花粉粒直径集中在100~120 μm之间; P2花粉粒直径集中在90~110 μm之间; F1花粉粒直径明显小于双亲, 且变异范围大, 集中在20~100 μm之间; M1花粉粒直径最大, 集中在100~140 μm之间(图5)。
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同世代花的表型性状比较(标尺: 20 mm)
Fig. 4Phenotype comparison of flowers among different generations (bar: 20 mm)
图5
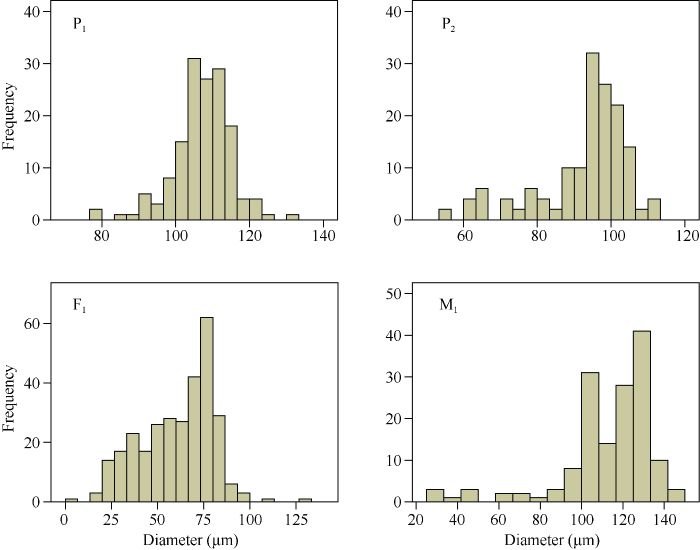 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5不同世代花粉粒直径的频率直方图
Fig. 5Frequency histogram of pollen grain diameter among different generations
2.2.3 棉纤维和种子腺体观察 陆地棉与斯特提棉种间异源六倍体棉纤维为白色, 纤维长度明显小于陆地棉。
异源六倍体表现出与斯特提棉相似的种子腺体延缓发育性状, P1和F1种子表面有腺体, P2和M1种子表面无肉眼可见腺体, 进一步在体视显微镜下观察发现, F1种子腺体密度明显小于P1, P2种子表面无腺体, M1种子表面有极少数颜色较淡的腺体, 种子表面腺体密度总体趋势为P2<M1<F1<P1 (图6)。随着种子萌发, M1和P2种子表面腺体逐渐增多, 子叶完全展开后, P1子叶腺体密度最大, P2子叶腺体形成速度最慢(图7)。种子腺体表型在不同世代间的差异对于研究种子腺体延缓发育基因的遗传规律和表达机制具有重要意义, 同时为该性状向陆地棉的转移提供了有价值的种质材料。
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6不同世代种子腺体性状比较
Fig. 6Comparison of seed gland traits among different generations
图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7不同世代子叶腺体性状比较
Fig. 7Comparison of cotyledon gland traits among different generations
3 讨论
3.1 陆地棉与斯特提棉的远缘杂交探讨
陆地棉农艺性状优良, 栽培范围广, 长期以来, 棉属远缘杂交研究多以陆地棉做母本, 希望将棉属其他种的优良性状转移到陆地棉中。斯特提棉具有耐低温、抗黄萎病、生长势强、纤维优质以及种子腺体延缓发育等多种优良性状[5,6], 是棉花远缘杂交育种关注度较高的一个野生种。我国关于陆地棉和斯特提棉的远缘杂交研究起步较晚, 但也获得了显著成效。姜茹琴等[8]用陆地棉与斯特提棉杂交, 对杂种进行胚胎培养和试管内染色体加倍, 获得的杂种后代多为混倍体, 根尖染色体数39~50条不等, 用可育株与陆地棉回交, 选育出一批棉花新品种。李育强等[9]以陆地棉和海岛棉为母本分别与斯特提棉杂交获得F1代, 观察其花粉母细胞减数分裂行为, 认为棉属A、C和D三个染色体组间同源性极低。崔荣霞等[10]在获得陆地棉与斯特提棉杂种F1代后, 对其枝条进行诱变加倍和扦插繁殖, 但无法获得可育植株, 继续与陆地棉回交获得BC1, 亦没有可育植株, 花粉母细胞减数分裂结果显示F1和BC1为非整倍体。梁正兰等[5]通过陆地棉和斯特提棉杂交, 胚的离体培养, 诱变加倍和回交等技术方法获得了新的种质资源和品种, 但并未获得真正的异源六倍体植株。本研究用陆地棉与斯特提棉进行杂交和染色体加倍, 通过流式细胞技术和花粉母细胞减数分裂观察, 筛选出2株染色体数完整的异源六倍体植株, 对斯特提棉在棉花育种中的开发利用和棉属远缘杂交育种研究具有重要价值。3.2 棉花种子腺体延缓发育性状探讨
种子无腺体而植株有腺体, 是棉花育种家想要转移到陆地棉中的一个重要性状, 但至今没有获得可用于生产的棉花品种。目前已发现的具有种子腺体延缓发育性状的棉种为分布在澳洲的几个野生二倍体棉种, 如斯特提棉、比克氏棉、澳洲棉、纳尔逊氏棉、南岱华棉等。李炳林等[11]报道了其获得的亚洲棉和比克氏棉的异源四倍体带有种子腺体延缓发育性状, 并且可以稳定遗传。张天真等[12]对具有种子低棉酚、植株有腺体(但低于常规品种)的陆地棉突变体分析发现, 其突变基因是Gl3的复等位基因。Zhu等[13]用亚洲棉与比克氏棉的异源四倍体与陆地棉杂交后不断回交和自交, 获得了带有种子腺体延缓发育性状的陆地棉种质, 遗传分析表明来自比克氏棉的腺体发育基因位于Gl2基因位点。关于棉属不同种所携带的腺体发育基因之间的互作关系, 祝水金等[14,15]用5个带有种子腺体延缓发育性状的澳洲野生棉种与A染色体组亚洲棉、D染色体组戴维逊氏棉以及AD染色体组的陆地棉4种不同色素腺体基因型(Gl2Gl2Gl3Gl3、Gl2Gl2gl3gl3、gl2gl2Gl3Gl3和gl2gl2gl3gl3)杂交, 初步明确5个澳洲棉野生种的腺体性状表达有个别差异, 但控制该性状的基因位于同一基因位点; 该基因对A染色体组亚洲棉为显性, 对D染色体组戴维逊氏棉为隐性, 对陆地棉Gl2为显性上位, 对陆地棉Gl3为隐性上位。另外, Liu等[16]在对草棉和澳洲棉杂交加倍后发现, 其获得的异源四倍体与母本草棉相比, 种子腺体数大大减少, 但仍有少量腺体, 这表明控制目标性状的非等位基因间并不是单纯的显隐性互作关系。这些发现尚有矛盾之处, 需要进一步的研究来证明控制棉花腺体发育基因之间的互作关系。本研究获得了陆地棉与斯特提棉的种间异源六倍体新种质, 带有与斯特提棉相似的种子腺体延缓发育性状, 种子腺体密度表现出P2<M1<F1<P1的梯度变化, 这为进一步研究棉花腺体基因间的互作关系提供了很好的材料, 我们将在随后的研究中对控制腺体延缓发育性状基因及在后代中的表达进行系统分析。
3.3 异源六倍体种质创新及利用价值
本研究获得异源六倍体可育后代, 其减数分裂行为已经趋于正常, 但不同六倍体植株间育性差异明显, 这为研究基因互作、表达调控及植株生理状况等因素对远缘杂种育性的影响提供了有价值的材料。异源六倍体具有种子腺体延缓发育特性, 种子腺体密度表现为P2<M1<F1<P1的梯度变化特性, 为研究棉花腺体基因互作和斯特提棉种子腺体延缓发育性状向陆地棉的转移提供了种质材料。在对本研究全部材料的统一管理中, 母本陆地棉在温室无法过冬, 父本斯特提棉以及远缘杂种F1、M0和M1全部正常过冬, 表现出远优于母本的耐低温特性, 可作为棉花耐低温育种的种质材料。在异源六倍体后代群体中, 易出现非整倍体植株, 需采取措施使其倍性稳定遗传。李炳林等[11]合成的亚洲棉与比克氏棉的异源四倍体, 初代胚根异常, 无法栽培成活, 通过嫁接和诱导生根进行多代繁殖, 获得了可正常生长且育性恢复良好的异源四倍体。吴玉香等[17]对亚洲棉、草棉、陆地棉和海岛棉四元杂种的减数分裂染色体构型和花粉粒活性多年测定证明, 活体保存对染色体组之间的协调和远缘杂种育性恢复有重要作用。本研究可以通过活体保存和多代繁殖的方法进一步稳定异源六倍体的倍性和育性。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1007/s00425-015-2450-xURLPMID:26715561 [本文引用: 1]

Main conclusion This article provides an up-to-date review concerning from basic issues of polyploidy to aspects regarding the relevance and role of both natural and artificial polyploids in plant...
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 5]
[本文引用: 5]
[本文引用: 4]
[本文引用: 4]
DOI:10.2135/cropsci1969.0011183X000900010009xURL [本文引用: 1]
Forty plant species were grown in pots containing viruliferous Xiphinema diversicaudatum (Micol.) for 15 wk to assess the host range of the nematode in relation to infection with arabis mosaic (AMV) and strawberry latent ringspot (SLRV) viruses. Host status for the nematode was determined mainly from changes in total populations, but the presence of eggs in the uteri of females and changes in the numbers of adults provided additional criteria. The nematode multiplied on relatively more woody perennials than on herbaceous crop plants or weeds. Chrysanthemum coronarium was the only plant on which numbers declined significantly below those on the controls. Most plant species became infected with either AMV or SLRV. Neither virus was detected in eight out of thirteen species of trees and shrubs although four were good hosts for the nematode. Galling or distortion of the terminal region of fine feeder roots, associated with X. diversicaudatum feeding, was seen on many of the experimental plants.
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
A tri-specific hybrid with delayed pigment gland morphogenesis was obtained by crossing the amphidiploid of (G. arboreum×G. bickii) F1 and an upland cotton germplasm with pigment gland genotype of Gl2Gl2gl3gl3. The tri-specific hybrid was a typical interspecific hybrid with high sterile, and the chromosome configuration at meiosis MI of PMC was 2n=52=41.04[+4.54Ⅱ+0.57Ⅲ+0.04.The crossover value of bivalent was 1.19. Two fertile plants with objective character were obtained in BCs population by continuously backcrossing with Gl2Gl2gl3gl3 as recurrent parent to the tri-specific hybrid, and a new upland cotton germplasm, named ABH-0318, with delayed pigment gland morphogenesis trait was developed through selfing and screening. The pigment gland trait of ABH-0318 was stable,and there were almost no pigment glands observed in the dormant seeds, although there were a few pigment glands confined to cotyledon edges, and the gossypol content in the dormant seeds was 0.017% only, being a typical low gossypol cotton type. However, a large quantity of pigment glandsemerged in cotyledons and other main organs of plant after seed germination, and the gossypol contents in the upper parts of the plant were similar to that of ordinary glanded cotton types. Genetic analysis demonstrated that the delayed pigment gland morphogenesis trait of this germplasm was controlled by the interaction of the genes located in two pigment gland loci, Gl2 and Gl3. Among them, the gene located in locus of Gl2, derived from Gbickii, was dominance to upland cotton pigment gland alleles, Gl2 and gl2, but was recessive epistatic to another glanded gene Gl3, which was named GI2b temporarily. While the gene located in the locus of Gl3 was a recessive gene come from upland cotton.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}