关键词:玉米; 谷氨酰胺合成酶(GS); 表达; BNE; 聚合方式 Differential Expression and Assembly Mode of Glutamine Synthetase Isoenzymes in Different Tissues and Organs of Maize WANG Xiao-Chun1,2,3, ZHANG Hao-Ran3, WEI Yi-Hao1, JIA Xi-Ting3, GU Ming-Xin3, MA Xin-Ming1,* 1Collaborative Innovation Center of Henan Grain Crops / Henan Agricultural University, Zhengzhou 450002, China
2State Key Laboratory of Wheat and Maize Crop Science in China / Henan Agricultural University, Zhengzhou 450002, China
3Department of Biochemistry, College of Life Science / Henan Agricultural University, Zhengzhou 450002, China
Fund:The study was supported by the National Key Research and Development Program of China (2016YFD0300205) and State Key Laboratory of Wheat and Maize Crop Science (39990047) AbstractGlutamine synthetase (GS) is a key enzyme in nitrogen assimilation and recycling in cereals. In this study, the expression characteristics of GS isoenzymes in different tissues and organs of maize in grain-filling period were analyzed, and the assembly of GS isoenzymes were indentified. The GS isoforms expressed differentially in different organs were shown by Western-blot obviously; GS1 with a molecular weight of about 40 kD expressed highly in all tissues, and GS1 with a molecular weight of about 39 kD was merely expressed in the node of ear position and pedical, and GS2 with a molecular weight of about 44 kD was weakly expressed in the photosynthtic tissue such as leaf. With a modified blue naive PAGE (BNE) technique and in-gel activity analysis, the size of GS holoenzyme was calibrated; combined the 2-D gel with western-blot analysis, the subunits composition of GS isoenzymes were identified. Three GS isoenzymes with different sizes were identified in maize. GS2 holoenzyme was about 460 kD and likely a decamer, GS1 holoenzyme existed two kinds of assembly state, one was about 410 kD and likely a decamer, another was about 240 kD and more likely a pentamer; therefore, the expression of GS isoenzymes exists diversity in maize.
Keyword:Maize; Glutamine synthetase (GS); Expression; Blue native PAGE (BNE); Assembly Show Figures Show Figures
图1 玉米不同组织GS同工酶表达和活性分析(A) Western blot鉴定GS同工酶亚基的表达。12% SDS-PAGE分离玉米蛋白, 转膜后利用小麦GS多克隆抗体检测GS亚基。(B)胶内GS酶活性检测。以5% Native-PAGE分离组织蛋白, 胶内GS酶活性分析检测GS同工酶活性。Roots: 根; Lower 1th leaf: 穗下第1叶; Sheath: 穗位叶鞘; Ear leaf: 穗位叶; Vein: 穗位叶叶脉; Internode: 穗位节间; Node: 穗位节; Pedicel: 果穗柄; Husk leaves: 苞叶; Grain: 籽粒; Top leaf: 顶部功能叶。Fig. 1 Analysis of the expression and activity of GS isoforms from different tissues of maize(A) Expression of GS isoforms detected by Western blot. Protein extracts separated by 12% SDS-PAGE, and then probed with GS ployclonal antibodies against wheat after electrobloting. (B) In-gel detection of GS activity. Proteins from different tissues were separated by 5% Native-PAGE, and GS isoforms were detected with the in-gel GS activity analysis.
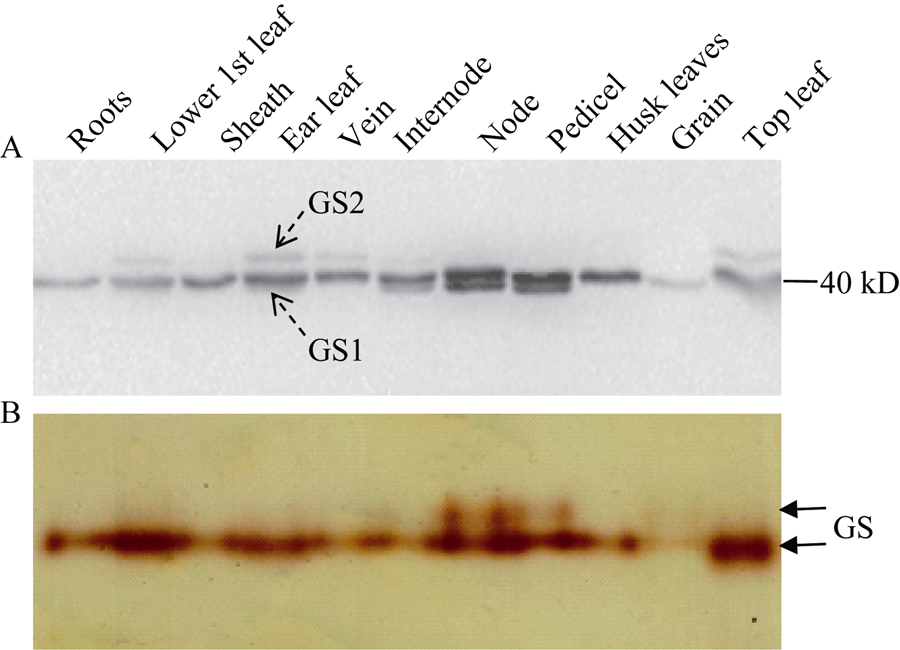
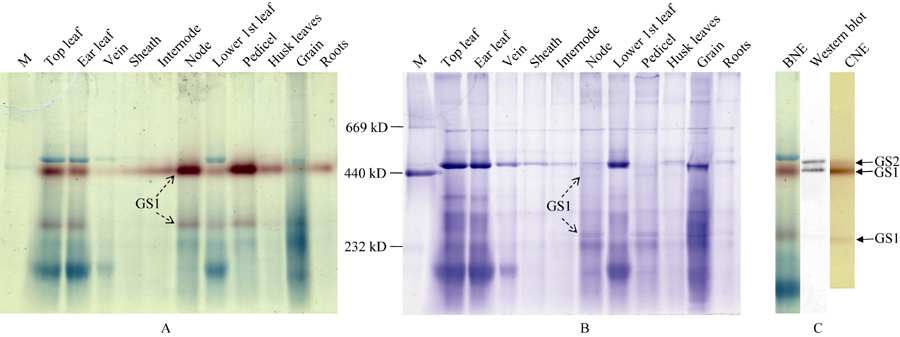
图2 玉米GS同工酶全酶大小鉴定(A)以BNE分离玉米不同组织可溶蛋白, 以胶内转移酶活性检测GS同工酶。(B) BNE胶考马斯亮蓝染色, 参照蛋白marker, 利用Gel-Pro analyzer计算GS同工酶全酶大小。(C)以BNE/CNE分离玉米穗位叶可溶蛋白, 转膜后进行Western blot检测。Fig. 2 Identification of the molecular weight of GS holoenzymes(A) Soluble proteins from different tissues of maize were separated by BNE, and GS isoforms were detected with an in-gel transferase activity assay. (B) CBB-staining after BNE, the molecular weight of GS holoenzymes were estimated using Gel-Pro analyzer with the calibration of the protein marker. (C) Proteins from the ear leaf of maize were separated by BNE/CNE, and GS isoforms were detected by Western blot.
图3 玉米GS同工酶亚基组成的鉴定(A)玉米穗位叶GS同工酶亚基组成鉴定, 利用BNE分离叶片可溶蛋白, 切取相应泳道进行变性处理, 利用SDS-PAGE分离复合体亚基, Western blot检测GS亚基。(B)穗柄中GS同工酶亚基鉴定, 方法步骤同(A)。Fig. 3 Identification of the subunit composition of GS isoenzymes in maize(A) Identification of the subunit composition of GS isoenzymes in the ear leaf of maize. Proteins were separated by BNE, the corresponding gel lane was cut out and denaturalized, and the protein complexes were separated by SDS-PAGE, then the GS subunits were identified by Western blot. (B) Identification of the subunit composition of GS isoenzymes from the pedicel of maize, the procedure was the same as that of (A).
Bernard SM, Habash DZ. The importance of cytosolic glutamine synthetase in nitrogen assimilation and recycling. New Phytol, 2009, 182: 608-620[本文引用:2]
[2]
McNally SF, HirelB, GadalP, Mann AF, Stewart GR. Glutamine synthetases of higher plants: evidence for a specific isoform content related to their possible physiological role and their compartmentation within the leaf. Plant Physiol, 1983, 72: 22-25[本文引用:3]
[3]
HirelB, GadalP. Glutamine synthetase isoforms in pea leaves: intracellular localization. Zeitschrift Fü Pflanzenphysiologie, 1981, 102: 315-319[本文引用:1]
[4]
HirelB, Lea P J. Ammonia Assimilation. Plant Nitrogen. Springer Berlin Heidelberg, 2001. pp 79-99[本文引用:1]
[5]
Zozaya-HinchliffeM, PotenzaC, Ortega JL, Sengupta-GopalanC. Nitrogen and metabolic regulation of the expression of plastidic glutamine synthetase in alfalfa (Medicago sativa). Plant Sci, 2005, 168: 1041-1052[本文引用:1]
[6]
CrenM, HirelB. Glutamine synthetase in higher plants regulation of gene and protein expression from the organ to the cell. Plant Cell Physiol, 1999, 40: 1187-1193[本文引用:1]
[7]
Robert FM, Wong PP. Isozymes of glutamine synthetase in Phaseolus vulgaris L. and Phaseolus lunatus L. root nodules. Plant Physiol, 1986, 81: 142-148[本文引用:1]
[8]
CarvalhoH, PereiraS, SunkelC, SalemaR. Detection of a cytosolic glutamine synthetase in leaves of Nicotiana tabacum L. by immunocytochemical methods. Plant Physiol, 1992, 100: 1591-1594[本文引用:1]
[9]
IshiyamaK, InoueE, Watanabe-TakahashiA, ObaraM, YamayaT, TakahashiH. Kinetic properties and ammonium-dependent regulation of cytosolic isoenzymes of glutamine synthetase in arabidopsis. J Biol Chem, 2004, 279: 16598-16605[本文引用:1]
[10]
LlorcaO, BettiM, González JM, ValenciaA, Márquez AJ, Valpuesta JM. The three-dimensional structure of an eukaryotic glutamine synthetase: functional implications of its oligomeric structure. J Structural Biol, 2006, 156: 469-479[本文引用:3]
[11]
Seabra AR, Vieira CP, Cullimore JV, Carvalho HG. Medicago truncatula contains a second gene encoding a plastid located glutamine synthetase exclusively expressed in developing seeds. BMC Plant Biol, 2010, 10: 183[本文引用:1]
[12]
Singh KK, GhoshS. Regulation of glutamine synthetase isoforms in two differentially drought-tolerant rice (Oryza sativa L. ) cultivars under water deficit conditions. Plant Cell Rep, 2013, 32: 183-193[本文引用:1]
[13]
MartinA, LeeJ, KicheyT, GerentesD, ZivyM, TatoutC, DuboisF, BalliauT, ValotB, DavantureM, Tercé-LaforgueT, QuilleréI, CoqueM, GallaisA, Gonzalez-Moro M B, Bethencourt L, Habash D Z, Lea P J, Charcosset A, Perez P, Murigneux A, Sakakibara H, Edwards K J, Hirel B. Two cytosolic glutamine synthetase isoforms of maize are specifically involved in the control of grain production. Plant Cell, 2006, 18: 3252-3274[本文引用:3]
[14]
McParland RH, Guevara JG, Becker RR, Evans HJ. The purification and properties of the glutamine synthetase from the cytosol of soya-bean root nodules. Biochem J, 1976, 153: 597-606[本文引用:1]
[15]
UnnoH, UchidaT, SugawaraH, KurisuG, SugiyamaT, YamayaT, SakakibaraH, HaseT, KusunokiM. Atomic structure of plant glutamine synthetase: a key enzyme for plant productivity. J Biol Chem, 2006, 281: 29287-29296[本文引用:4]
[16]
Seabra AR, CarvalhoH, Pereira PJ. Crystallization and preliminary crystallographic characterization of glutamine synthetase from Medicago truncatula. Acta Crystallographica, 2009, 65: 1309-1312[本文引用:2]
[17]
TorreiraE, Seabra AR, MarriottH, ZhouM, LlorcaÓ, Robinson CV, Carvalho HG, Fernández-TorneroC, Pereira PJ. The structures of cytosolic and plastid-located glutamine synthetases from Medicago truncatula reveal a common and dynamic architecture. Acta Crystallographica, 2014, 70: 981-993[本文引用:2]
WittigI, KarasM, SchäggerH. High resolution clear native electrophoresis for in-gel functional assays and fluorescence studies of membrane protein complexes. Mol Cell Proeom, 2007, 6: 1215-1225[本文引用:3]
[20]
Kimata-ArigaY, HaseT. Multiple complexes of nitrogen assimilatory enzymes in spinach chloroplasts: possible mechanisms for the regulation of enzyme function. PLoS One, 2014, 9(10): e108965[本文引用:2]
[21]
Seabra AR, Silva LS, Carvalho HG. Novel aspects of glutamine synthetase (GS) regulation revealed by a detailed expression analysis of the entire GS gene family of Medicago truncatula under different physiological conditions. BMC Plant Biol, 2013, 13: 137-142[本文引用:2]
[22]
Wang XC, Wei YH, Shi LX, Ma XM, Theg SM. New isoforms and assembly of glutamine synthetase in the leaf of wheat (Triticum aestivum L. ). J Exp Bot, 2015, 66: 6827-6834[本文引用:7]
[23]
Mann AF, Fentem PA, Stewart GR. Identification of two forms of glutamine synthetase in barley (Hordeum vulgare). Biochem Biophys Res Commun, 1979, 88: 515-521[本文引用:1]
[24]
郭玉朋. 植物光呼吸途径研究进展. 草业学报, 2014, 23: 322-329Guo YP. A study on advances in plant photorespiration. Acta Pratac Sin, 2014, 23: 322-329 (in Chinese with English abstract)[本文引用:1]
[25]
Bernard SM, Møller AL, DionisioG, KicheyT, Jahn TP, DuboisF, BaudoM, Lopes MS, Tercé-LaforgueT, Foyer CH, Parry MA, Forde BG, Araus JL, HirelB, Schjoerring JK, Habash DZ. Gene expression, cellular localisation and function of glutamine synthetase isozymes in wheat (Triticum aestivum L. ). Plant Mol Biol, 2008, 67: 89-105[本文引用:1]
[26]
HirelB, AndrieuB, Valadier MH, RenardaS, QuilleréI, ChellebM, PommelB, FournierC, Drouetb JL. Physiology of maize: II. identification of physiological markers representative of the nitrogen status of maize (Zea mays) leaves during grain filling. Physiol Plant, 2005, 124: 178-188[本文引用:1]
 , 张浩然
, 张浩然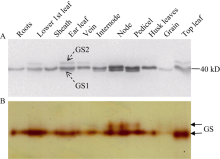


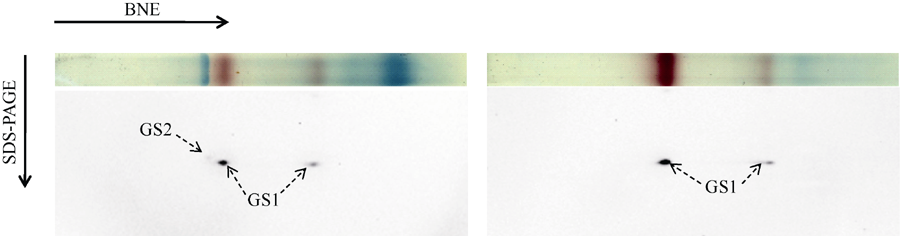

{kind=link}
{kind=link}
{kind=link}