关键词:芝麻; 含油量; 蛋白质含量; 芝麻素含量; QTL QTL Mapping for Oil, Protein and Sesamin Contents in Seeds of White Sesame WU Kun1,2, WU Wen-Xiong1, YANG Min-Min1, LIU Hong-Yan1, HAO Guo-Cun1, ZHAO Ying-Zhong1,* 1 Oil Crops Research Institute, Chinese Academy of Agricultural Sciences / Key Laboratory of Biology and Genetic Improvement of Oil Crops, Ministry of Agriculture, Wuhan 430062, China
2 Biotechnology Research Center of Southwest University, Chongqing 400715, China
Fund:The study was supported by the National Natural Science Foundation of China (31201243), the National Key Basic Research Program of China (973 Program) (2011CB109304), and the China Agriculture Research System (CARS-15). AbstractSeed oil content, protein content and sesamin content are three important targets for sesame breeding. The objective of this study was to explore the genetic model and detect quantitative trait loci (QTLs) for three quality traits using a mapping population containing 224 recombinant inbred lines (RILs, F9). The three quality traits of seeds were measured by near infrared reflectance spectroscopy in three environments. Correlation analysis showed oil content was negatively correlated with protein content but positively correlated with sesamin content, whereas, protein content was negatively correlated with sesamin content. Using mixed composite interval mapping (MCIM) method, eight QTLs for the three seed quality traits were detected with heritability of additive effect ranging from 0.41% to 14.55%. Using multiple interval mapping (MIM) method, 13 QTLs were mapped on nine linkage groups (LGs), and a single QTL explained 5.2%-18.6% of the total phenotypic variation. Among them, five loci had at least one marker identical to the QTL identified with MCIM, indicating they might be located in the same region. Two QTLs were detected in multiple environments. QTLs Qoc-5 and Qsc-5 were located in the same region on LG5, both had positive effects, whereas Qpc-5 in the adjacent region had a negative effect. Similar loci exist on LG2 and LG1, probably caused by pleiotropism or close linkage of several QTLs. These results suggest that the oil and sesamin contents in seed can be improved together, while the protein content needs to be selected negatively with the oil or sesamin content.
表3 利用MCIM模型同时检测3个环境下RIL群体品质性状相关QTL Table 3 QTL for quality traits detected by MCIM using the RILs in multi-trials
性状 Trait
QTL
连锁群 LG
标记区间 Marker interval
区域 Region (cM)
位置 Position
加性效应a Additive effect a
遗传率b ha2(%)
广义遗传率c H2(%)
含油量 Oil content
Qoc-5
LG5
SBN3585-SBN1490
49.4-52.1
51.6
0.2616* * *
3.47
40.38
蛋白质含量 Protein content
Qpc-2
LG2
SBN2776-SBN2749
11.9-16.2
13.5
-0.2606* * *
4.26
22.76
Qpc-5
LG5
SBI007-SBN795
63.5-65.8
64.5
-0.2343* * *
3.31
芝麻素含量 Sesamin content
Qsc-5
LG5
ZHY01-SBN3585
48.4-51.6
51.4
0.0160* * *
14.55
14.68
Qsc-6
LG6
SBI043-SBN636
95.0-102.4
100.4
0.0051* * *
2.01
Qsc-8
LG8
SBN1735-SBN1407
35.4-37.0
36.4
0.0088* * *
5.05
Qsc-11
LG11
SBN3764-SBN1798
16.9-18.2
17.9
0.0053* * *
0.41
Qsc-16
LG16
SBN1927-SBN3232
22.6-29.1
23.6
-0.0091* * *
3.34
aPositive and negative values indicate additive effect coming from the alleles of Zhongzhi 14 and Miaoqianzhima, respectively; * , * * , * * * Significant at the 0.05, 0.01, and 0.001 probability levels, respectively; bContribution ratio of QTL additive effect, c The broad-sense heritability (H2). a正值或负值分别代表加性效应来自亲本中芝14或庙前芝麻的等位变异, * , * * , * * * 分别表示在0.05, 0.01, 0.001显著水平; bQTL加性效应遗传率; c广义遗传率H2。
表3 利用MCIM模型同时检测3个环境下RIL群体品质性状相关QTL Table 3 QTL for quality traits detected by MCIM using the RILs in multi-trials
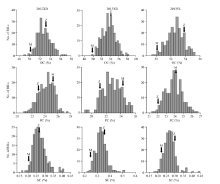
图1 RIL群体3个品质性状在3个环境下的频率分布(OC、PC和SC分别代表油分、蛋白质和芝麻素含量, 箭头所指为各环境中亲本测定值, Z和M分别代表亲本中芝14和庙前芝麻。)Fig. 1 Frequency distribution of three quality traits of RIL population in three environments(OC: oil content; PC: protein content; SC: sesamin content. Arrows indicate determined values of two parents under environments. Z and M stand for Zhongzhi 14 and Miaoqianzhima, respectively.)
表4 利用MIM模型分别检测3个环境下RIL群体品质性状相关QTL Table 4 QTL for quality traits detected by MIM using the RILs in three environment traits
性状 Trait
试验地 Location
QTL
连锁群 LG
标记区间 Marker interval
区域 Region (cM)
位置 Position
LOD
加性效应a Additive a
表型变异贡献率bR2 (%)
含油量 Oil content
2012徐东 2012XD
Qoc-1
LG1
SBN2389-SBN2485
37.0-40.6
39.6
4.67
-0.4274
10.1
2013徐东 2013XD
Qoc-2
LG2
SBN2776-SBN1045
13.5-15.2
14.5
3.70
0.4009
6.5
2013阳逻 2013YL
Qoc-5
LG5
SBN3585-SBN1490
41.5-52.1
44.4
4.44
0.3189
7.7
2013徐东 2013XD
Qoc-9
LG9
SBN1388-SBN1459
6.9-9.7
9.5
4.68
-0.4567
8.6
2012徐东 2012XD
Qoc-16
LG16
SBN1927-SBN3232
22.6-26.6
23.6
3.36
-0.3740
7.6
蛋白含量 Protein content
2012徐东 2012XD
Qpc-1
LG1
SBN741-SBN2389
20.0-28.7
20.4
3.28
0.3293
7.6
2013徐东 2013XD
Qpc-2
LG2
SBN2776-SBN2749
13.5-15.2
14.8
3.38
-0.3290
6.2
2013阳逻 2013YL
Qpc-3
LG3
HS184-SBI013
6.3-9.7
7.7
3.63
-0.2356
5.7
2013阳逻 2013YL
Qpc-5
LG5
SBI007-SBI057
62.8-68.4
64.5
5.75
-0.3280
10.9
2013阳逻 2013YL
Qpc-6
LG6
SBN3210-SBN636
94.0-102.4
99.0
4.00
-0.2470
6.4
芝麻素含量 Sesamin content
2013阳逻 2013YL
Qsc-4
LG4
SBI050-SBN1100
70.7-73.7
73.7
3.36
0.0082
5.4
2012徐东 2012XD
Qsc-5
LG5
ZHY01-SBN1548
49.4-56.2
51.4
5.71
0.0132
11.1
2013徐东 2013XD
Qsc-5
LG5
SBN3568-SBI007
40.5-62.9
52.1
11.23
0.0194
18.6
2013阳逻 2013YL
Qsc-5
LG5
SBN3568-SBI007
40.5-62.9
51.6
8.29
0.0130
14.1
2013徐东 2013XD
Qsc-8
LG8
SBN1735-SBN2668
35.4-44.6
36.4
3.96
0.0134
8.9
2013阳逻 2013YL
Qsc-8
LG8
SBN1407-SBN1140
37.0-38.2
37.4
3.09
0.0100
5.2
aPositive and negative values indicate additive effect coming from the allele of Zhongzhi 14 and Miaoqianzhima, respectively. bProportion of phenotypic variation explained by individual QTL. a正值或负值分别代表加性效应来自亲本中芝14或庙前芝麻的等位变异; b单个QTL解释的表型变异率。
表4 利用MIM模型分别检测3个环境下RIL群体品质性状相关QTL Table 4 QTL for quality traits detected by MIM using the RILs in three environment traits
张书芬, 傅廷栋, 朱家成, 王建平, 文雁成, 马朝芝. 甘蓝型油菜产量及其构成因素的QTL定位与分析. 作物学报, 2006, 32: 1135-1142Zhang SF, Fu TD, Zhu JC, Wang JP, Wen YC, Ma CZ. QTL mapping and epistasis analysis for yield and its components in Brassica napus L. Acta Agron Sin, 2006, 34: 1135-1142 (in Chinese with English abstract)[本文引用:1]
[3]
Yan XY, Li JN, Fu FY, Jin MY, Jin MY, ChenL, Liu LZ. Co-location of seed oil content, seed hull content and seed coat color QTL in three different environments in Brassica napus L. Euphytica, 2009, 170: 355-364[本文引用:1]
[4]
金梦阳, 李加纳, 付福友, 张正圣, 张学昆, 刘列钊. 甘蓝型油菜含油量及皮壳率的QTL分析. 中国农业科学, 2007, 40: 677-684Jin MY, Li JN, Fu FY, Zhang ZS, Zhang XK, Liu LZ. QTL Analysis of oil and hull content in Brassica napus L. Sci Agric Sin, 2007, 40: 677-684 (in Chinese with English abstract)[本文引用:1]
[5]
Zhang WK, Wang YJ, Luo GZ, Zhang JS, He CY, Wu XL, Gai JY, Chen SY. QTL mapping of ten agronomic traits on the soybean ( Glycine max(L. ) Merr. ) genetic map and association with EST markers. Theor Appl Genet, 2004, 108: 1131-1139[本文引用:1]
[6]
郑永战, 盖钧镒, 卢为国, 李卫东, 周瑞宝, 田少君. 大豆脂肪及脂肪酸组分含量的QTL定位. 作物学报, 2006, 32: 1823-1830Zheng YZ, Gai JY, Lu WG, Li WD, Zhou RB, Tian SJ. QTL mapping for fat and fatty acid composition contents in soybean. Acta Agron Sin, 2006, 32: 1823-1830 (in Chinese with English abstract)[本文引用:1]
[7]
廖伯寿, 雷永, 王圣玉, 李栋, 黄家权, 姜慧芳, 任小平. 花生重组近交系群体的遗传变异与高油种质的创新. 作物学报, 2008, 34: 999-1004Liao BS, LeiY, Wang SY, LiD, Huang JQ, Jiang HF, Ren XP. Genetic diversity of peanut RILs and enhancement for high oil genotypes. Acta Agron Sin, 2008, 34: 999-1004 (in Chinese with English abstract)[本文引用:1]
[8]
Hui YH(编), 徐生度, 裘爱泳(译). 油脂化学与工艺学(第5版, 第3卷). 北京: 中国轻工业出版社, 2001. pp 474-513Hui Y Hed. , Xu S D, Qiu A Y trans. Industrial Oil & Fat Products, 5th edn, Vol. 3. Beijing: China Light Industry Press, 2001. pp 474-513(in Chinese)[本文引用:1]
[9]
梅鸿献, 魏安池, 刘艳阳, 王春弘, 杜振伟, 郑永战. 芝麻种质资源芝麻素蛋白质脂肪含量变异及其相关分析. 中国油脂, 2013, 38(4): 87-90Mei HX, Wei AC, Liu YY, Wang CH, Du ZW, Zheng YZ. Variation and correlation analysis of sesamin, oil and protein contents in sesame germplasm resources. China Oils Fats, 2013, 38(4): 87-90 (in Chinese with English abstract)[本文引用:2]
[10]
张秀荣, 李培武, 汪雪芳, 王旭. 芝麻种子木质素组分、粗脂肪、粗蛋白含量及相关性分析. 中国油料作物学报, 2005, 27(3): 88-90Zhang XR, Li PW, Wang XF, WangX. Studies on relationship among lignans, oil and protein content in sesame seed. Chin J Oil Crop Sci, 2005, 27(3): 88-90 (in Chinese with English abstract)[本文引用:1]
[11]
Wang LH, Zhang YX, Li PW, Wang XF, ZhangW, Wei WL, Zhang XR. HPLC analysis of seed sesamin and sesamolin variation in a sesame germplasm collection in China. J Am Oil Chem Soc, 2012, 89: 1011-1020[本文引用:1]
[12]
金青哲, 刘元法, 王兴国, 戴洪平. 国产芝麻中芝麻素含量分析. 中国粮油学报, 2005, 20(4): 85-87Jin QZ, Liu YF, Wang XG, Dai HP. Analysis of sesamin in domestic Sesamum indicum L. J Chin Cereals Oils Assoc, 2005, 20(4): 85-87 (in Chinese with English abstract)[本文引用:1]
[13]
危文亮, 张艳欣, 吕海霞, 王林海, 黎冬华, 张秀荣. 芝麻资源群体结构及含油量关联分析. 中国农业科学, 2012, 45: 1895-1903Wei WL, Zhang YX, Lyu HX, Wang LH, Li DH, Zhang XR. Population structure and association analysis of oil content in a diverse set of Chinese sesame ( Sesamum indicum L. ) germplasm. Sci Agric Sin, 2012, 45: 1895-1903 (in Chinese with English abstract)[本文引用:2]
[14]
王蕾, 黎冬华, 齐小琼, 张艳欣, 丁霞, 王林海, 危文亮, 高媛, 张秀荣. 芝麻核心种质芝麻素和芝麻酚林的关联分析. 中国油料作物学报, 2014, 36: 32-37WangL, Li DH, Qi XQ, Zhang YX, DingX, Wang LH, Wei WL, GaoY, Zhang XR. Association analysis of sesamin and sesamolin in the core sesame ( Sesamum indicum L. ) germplasm. Chin J Oil Crop Sci, 2014, 36: 32-37 (in Chinese with English abstract)[本文引用:2]
[15]
WuK, Liu HY, Yang MM, TaoY, Ma HH, Wu WX, ZuoY, Zhao YZ. High-density genetic map construction and QTL analysis of grain yield-related traits in sesame ( Sesamum indicum L. ) based on RAD-Seq techonology. BMC Plant Biol, 2014, 14: 274[本文引用:2]
[16]
Schlotzhauer SD, Littell RC. SAS System for Elementary Statistical Analysis. SAS institute, Cary, North Carolina. 1997[本文引用:1]
[17]
WangS, Basten CJ, Zeng ZB. Windows QTL cartographer version 2. 5. Department of statistics, North Carolina State University, Raleigh, NC. 2006. http://statgen.ncsu.edu/qtlcart/WQTLCart.htm[本文引用:1]
[18]
YangJ, HuC, HuH, YuR, XiaZ. QTLNetwork: mapping and visualizing genetic architecture of complex traits in experimental populations. Bioinformatics, 2008, 24: 721-723[本文引用:1]
Kamal-EldinA, Appelqvist LA, YousifG. Lignan analysis in seed oils from four Sesamum species: comparison of different chromatographic methods. J Oil Fat Ind, 1994, 71: 141-147[本文引用:1]
[21]
Zhang SF, Ma CZ, Zhu JC, Wang JP, Wen YC, Fu TD. Genetic analysis of oil content in Brassica napus L. using mixed model of major gene and polygene. Acta Genet Sin, 2006, 33: 171-180[本文引用:2]
[22]
Zhao JY, Becker HC, Zhang DQ, Zhang YF, EckeW. Oil content in a European × Chinese rape seed population QTL with additive and epistatic effects and their genotype-environment interactions. Crop Sci, 2005, 45: 51-59[本文引用:1]
[23]
马珍珍, 李加纳, WittkopB, FrauenM, 阎星颖, 刘列钊, 肖阳. 甘蓝型油菜籽粒含油量、蛋白质、纤维素及半纤维素含量QTL分析. 作物学报, 2013, 39: 1214-1222Ma ZZ, Li JN, WittkopB, FrauenM, Yan XY, Liu LZ, XiaoY. QTL Mapping for oil, protein, cellulose, and hemicellulose contents in seeds of Brassica napus L. Acta Agron Sin, 2013, 39: 1214-1222 (in Chinese with English abstract)[本文引用:1]
SiP, Rodney JM, NickG, David WT. Influence of genotype and environment on oil and protein concentrations of canola ( Brassica napus L. ) across southern Australia. Aust J Agric Res, 2003, 54: 397-407[本文引用:1]
[26]
禹山林, 杨庆利, 潘丽娟, 薄文娜. 花生种子含油量的遗传分析. 植物遗传资源学报, 2009, 10: 453-456Yu SL, Yang QL, Pan LJ, Bo WN. Genetic analysis for oil content of peanut seeds. J Plant Genet Resour, 2009, 10: 453-456 (in Chinese with English abstract)[本文引用:1]



{kind=link}