关键词:小麦; 穗部性状; 90k基因芯片; QTL定位 QTL Mapping for Spike Traits of Wheat Using 90k Chip Technology WU Bing-Jin, JIAN Jun-Tao, ZHANG De-Qiang, MA Wen-Jie, FENG Jie, CUI Zi-Xia, ZHANG Chuan-Liang, SUN Dao-Jie* College of Agronomy, Northwest A&F University, Yangling 712100, China Fund:This study was supported by the National Key Basic Research Program of China (2014CB138100), the Natural Science Foundation of Shaanxi Province (2015JM3094), and the Key Scientific and Technological Innovation Team of Shaanxi Province (2014KCT-25). AbstractSpike traits are important to grain yield in wheat. Molecular markers associated with genes/QTLs controlling spike traits are highly valuable to marker-assisted breeding. A recombinant inbred line (F8) population derived from Zhou 8425B ×Xiaoyan 81 were evaluated in three environments, and QTLs for spike length, spikelet number per spike, sterile spikelet number, grain number per spike and thousand-grain weigh were mapped into a high-density genetic map built by 90k chip. A total of 71 QTLs were located on 19 chromosomes, and the phenotype variation explained (PVE) by a single locus ranged from 2.10% to 45.25%. Thirty-seven loci were considered as main-effect QTLs owing to the PVE larger than 10%. QTLs QSl.nafu-6A.2 for spike length, QSl.nafu-7A for spike length, QSsn.nafu-2A.1 for sterile spikelet number, QSsn.nafu-2D for sterile spikelet number and QGns.nafu-2B for grain number per spike were identified repeatedly in different environments with the LOD value higher than 10 and PVE larger than 20%. QSl.nafu-6A.2 for spike length, QGns.nafu-6A for grain number per spike and QTgw.nafu-6A for thousand-grain weight were mapped in a cluster on chromosome 6A and might be applicable in marker-assisted selection because they have been detected in multiple environments and close to the loci reported.
Keyword: Triticumaestivum; Spike-related traits; 90k gene chip; QTL mapping Show Figures Show Figures
表1 穗部性状表型分析 Table 1 Phenotypic analysis of panicle traits
环境 Environ.
亲本 Parent
RIL群体 RIL population
遗传力 Heritability
周8425B Zhou 8425B
小偃81 Xiaoyan 81
均值 Mean
标准差 SD
最大值 Max
最小值 Min
峰度 Kurtosis
偏度 Skewness
变异系数 CV (%)
穗长 Spike length (mm)
E1
12.67
7.67
11.60
1.15
14.50
8.67
0.14
-0.11
10.06
0.82
E2
12.38
8.76
10.28
0.95
12.96
8.48
0.77
0.20
10.85
E3
10.56
7.64
9.41
0.78
11.16
8.16
0.26
0.09
12.02
AE
11.87
8.02
10.43
0.81
14.58
8.64
-0.01
0.39
12.88
小穗数 Spikelet number per spike
E1
25.00
20.33
22.05
1.39
23.00
18.67
-0.42
-0.19
15.84
0.65
E2
19.40
19.00
18.87
1.75
22.06
14.20
-0.04
-0.21
10.81
E3
19.75
18.25
18.00
1.15
20.75
14.50
0.22
-0.06
15.60
AE
21.38
19.19
19.64
1.06
21.67
14.12
0.61
-0.41
18.57
穗粒数 Grain number per spike
E1
32.60
40.40
41.12
5.96
65.56
30.00
0.24
0.33
6.90
0.57
E2
43.70
46.57
47.57
8.90
72.60
24.80
0.16
0.26
5.35
E3
41.35
39.25
45.11
5.18
71.00
34.75
0.77
0.14
8.71
AE
39.22
42.07
44.60
4.60
64.14
25.52
0.43
0.59
9.69
不育小穗数 Sterile spikelet number
E1
2.67
3.00
2.35
1.08
4.67
0.00
-0.34
0.06
2.18
0.75
E2
2.20
2.20
1.97
0.78
4.20
0.00
0.32
0.04
2.52
E3
1.75
1.50
1.61
0.59
3.00
0.00
0.05
-0.34
2.75
AE
2.21
2.23
1.98
0.65
3.32
0.15
0.10
-0.56
3.03
千粒重 Thousand-grain weight (g)
E1
54.55
40.75
40.35
9.68
61.15
29.95
-0.62
-0.15
4.17
0.76
E2
60.67
43.43
47.92
10.32
61.08
30.81
-0.74
-0.25
4.64
E3
61.36
42.66
49.42
5.90
61.34
30.67
0.57
-0.47
8.37
AE
58.86
42.28
45.97
6.99
61.02
32.48
0.22
-0.24
6.58
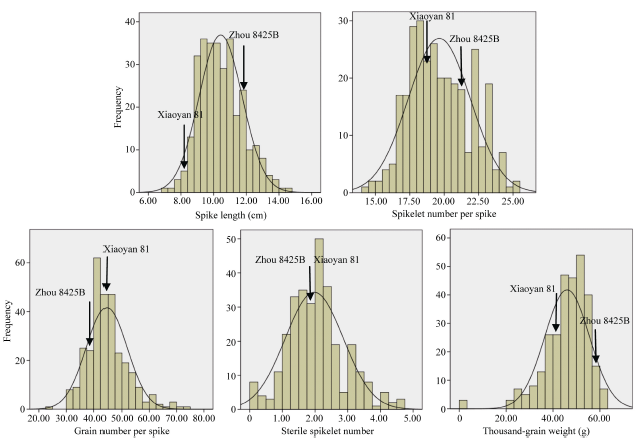
E1, E2, E3, and AE refer to environments of 2013-2014 Yangling, 2014-2015 Anyang, 2015-2016 Yangling, and the average, respectively. E1、E2、E3和AE分别表示2013-2014陕西杨凌、2014-2015河南安阳、2015-2016陕西杨凌和平均环境。
表1 穗部性状表型分析 Table 1 Phenotypic analysis of panicle traits
图2 4A、6A和7B染色体上定位的穗部性状QTL(穗长、小穗数、不育小穗数、穗粒数和千粒重相关QTL分别用黑、红、绿、蓝和棕色标注。)Fig. 2 QTLs for spike traits on chromosomes 4A, 6A, and 7B(QTLs for spike length, spikelet number per spike, sterile spikelet number, grain number per spike, and thousand-grain weight were marked in black, red, green, blue, and brown, respectively.)
Huang XQ, KempfH, Canal MW, Roder MS. Advanced backcross QTL analysis in progenies derived from a cross between a German elite winter wheat variety and a synthetic wheat ( Triticum aestivum L . ). Theor Appl Genet, 2004, 109: 933-943[本文引用:1]
[2]
Ma ZQ, Zhao DM, Zhang CQ, Zhang ZZ, Xue SL, LinF, Kong ZX, Tian DG, Luo QY. Molecular genetic analysis of five spike-related traits in wheat using RIL and immortalized F2 populations. Mol Genet Genomics, 2007, 277: 31-42[本文引用:1]
[3]
DengS, WuX, WuY, ZhouR, WangH, JiaJ, LiuS. Characterization and precise mapping of a QTL increasing spike number with pleiotropic effects in wheat. Theor Appl Genet, 2011, 122: 281-289[本文引用:1]
[4]
CuiF, Zhao CH, Ding AM, LiJ, WangL, Li XF, Bao YG, Li JM, Wang HG. Construction of an integrative linkage map and QTL mapping of grain yield-related traits using three related wheat RIL populations. Theor Appl Genet, 2014, 127: 659-675[本文引用:1]
[5]
CuiF, Ding AM, LiJ, Zhao CH, WangL, Wang XQ, Qi XL, Li XF, Li GY, Gao JR, Wang HG. QTL detection of seven spike-related traits and their genetic correlations in wheat using two related RIL populations. Euphytica, 2012, 186: 177-192[本文引用:1]
[6]
卢翔, 张锦鹏, 王化俊, 杨欣明, 李秀全, 李立会. 小麦-冰草衍生后代3558-2穗部相关性状的遗传分析和QTL定位. 植物遗传资源学报, 2011, 12: 86-91LuX, Zhang JP, Wang HJ, Yang XM, Li XQ, Li LH. Genetic analysis and QTL mapping of wheat spike traits in a derivative line 3558-2 from wheat× Agropyron cristatum offspring. Plant Genet Resour, 2011, 12: 86-91 (in Chinese with English abstract)[本文引用:1]
[7]
WangJ, LiuW, WangH, LiH, WuJ, YangX, LiX, GaoA. QTL mapping of yield-related traits in the wheat germplasm 3228. Euphytica, 2011, 177: 277-292[本文引用:1]
[8]
LiS, JiaJ, WeiX, ZhangX, LiL, ChenH, FanY, SunH, ZhaoX, LeiT, XuY, JiangF, WangH, LiL. An intervarietal genetic map and QTL analysis for yield traits in wheat. Mol Breed, 2007, 20: 167-178[本文引用:1]
[9]
WangS, WongD, ForrestK, AllenA, ChaoS, Huang BE, MaccaferriM, SalviS, Milner SG, CattivelliL, Mastrangelo AM, WhanA, StephenS, BarkerG, WiesekeR, PlieskeJ, International Wheat Genome Sequencing Consortium, Lillemo M, Mather D, Appels R, Dolferus R, Brown-Guedira G, Korol A, Akhunova A R, Feuillet C, Salse J, Morgante M, Pozniak C, Luo M C, Dvorak J, Morell M, Dubcovsky J, Ganal M, Tuberosa R, Lawley C, Mikoulitch I, Cavanagh C, Edwards K J, Hayden M, Akhunov E. Characterization of polyploid wheat genomic diversity using a high-density 90000 single nucleotide polymorphism array. Plant Biotechnol J, 2014, 12: 787-796[本文引用:1]
[10]
李立会. 小麦种植资源描述规范和数据标准. 北京: 中国农业出版社, 2006. pp 8-45Li LH. Descriptors and Data Stand ard for Wheat (Triticunm aestivum L. ). Beijing: Chinese Agricultural Science and Technology Press, 2006. pp 8-45(in Chinese)[本文引用:1]
[11]
王建康. 数量性状基因的完备区间作图方法. 作物学报, 2009, 35: 239-245Wang JK. Inclusive composite intervalmapping of quantitative trait genes. Acta Agron Sin, 2009, 35: 239-245 (in Chinese with English abstract)[本文引用:1]
[12]
You GX, Zhang XY, Wang LF. An estimation of the minimum number of SSR loci needed to reveal genetic relationships in wheat varieties: information from 96 rand om accessions with maximized genetic diversity. Mol Breed, 2004, 14: 397-406[本文引用:1]
[13]
AlamerewS, ChebotarS, HuangX, RöderM, BörnerA. Genetic diversity in Ethiopian hexaploid and tetraploid wheat germplasm assessed by microsatellite markers. Genet Resour Crop Evol, 2004, 51: 559-567[本文引用:1]
[14]
郝晨阳, 王兰芬, 张学勇, 游光霞, 董玉琛, 贾继增, 刘旭, 尚勋武, 刘三才, 曹永生. 我国育成小麦品种的遗传多样性演变. 中国科学: 生命科学, 2005, 35: 408-415Hao CY, Wang LF, Zhang XY, You GX, Dong YC, Jia JZ, LiuX, Shang XW, Liu SC, Cao YS. Changes of genetic diversity of wheat varieties released in China. Sci China: Life Sci, 2005, 35: 408-415 (in Chinese with English abstract)[本文引用:1]
[15]
RousselV, KoenigJ, BeckertM, BalfourierF. Molecular diversity in French bread wheat accessions related to temporal trends and breeding programmes. Theor Appl Genet, 2004, 108: 920-930[本文引用:1]
[16]
贾继增, 张正斌. 小麦21条染色体RFLP作图位点遗传多样性分析. 中国科学: 生命科学, 2001, 31: 13-21Jia JZ, Zhang ZB. Genetic diversity analysis of 21 chromosomes of wheat with RFLP marker. Sci China: Life Sci, 2001, 31: 13-21[本文引用:1]
[17]
JiaJ, ZhaoS, KongX. Aegilops tauschii draft genome sequence reveals a gene repertoire for wheat adaptation. Nature, 2013, 496: 91-95[本文引用:2]
[18]
吴秋红, 陈娇娇, 陈永兴, 周升辉, 傅琳, 张德云, 肖尧, 王国鑫, 王振忠, 王立新, 韩俊, 袁成国, 尤明山, 刘志勇. 燕大1817/北农6号重组自交系群体穗部性状的QTL定位. 作物学报, 2015, 41: 349-358Wu QH, Chen JJ, Chen YX, Zhou SH, FuL, Zhang DY, XiaoY, Wang GX, Wang ZZ, Wang LX, HanJ, Yuan CG, You MS, Liu ZY. Mapping quantitative trait loci related to spike traits using a RILs population of Yand a 1817 × Beinong 6 in wheat ( Triticum aestivum L. ). Acta Agron Sin, 2015, 41: 349-358 (in Chinese with English abstract)[本文引用:4]
[19]
高尚, 莫洪君, 石浩然, 王智强, 林宇, 武方琨, 邓梅, 刘亚西, 魏育明, 郑有良. 利用SNP基因芯片技术进行小麦遗传图谱构建及重要农艺性状QTL分析. 应用与环境生物学报, 2016, 22: 85-94GaoS, Mo HJ, Shi HR, Wang ZQ, LinY, Wu FK, DengM, Liu YX, Wei YM, Zheng YL. Construction of wheat genetic map and QTL analysis of main agronomic traits using SNP genotyping chips technology. Chin J Appl Environ Biol, 2016, 22: 85-94 (in Chinese with English abstract)[本文引用:1]
[20]
LiC, BaiG, Carver BF, ChaoS, WangZ. Mapping quantitative trait loci for plant adaptation and morphology traits in wheat using single nucleotide polymorphisms. Euphytica, 2016, 208: 299-312[本文引用:1]
[21]
刘凯, 邓志英, 李青芳, 张莹, 孙彩铃, 田纪春, 陈建省. 利用高密度SNP遗传图谱定位小麦穗部性状基因. 作物学报, 2016, 42: 820-831LiuK, Deng ZY, Li QF, ZhangY, Sun CL, Tian JC, Chen JS. Mapping QTLs for wheat panicle traits with high density SNP genetic map. Acta Agron Sin, 2016, 42: 820-831 (in Chinese with English abstract)[本文引用:4]
[22]
CuiF, Ding AM, LiJ, Zhao CH, WangL, Wang XQ, Qi XL, Li XF, Li GY, Gao JR, Wang HG. QTL detection of seven spike-related traits and their genetic correlations in wheat using two related RIL populations. Euphytica, 2012, 186: 177-192[本文引用:1]
[23]
LiC, BaiG, Carver BF. Single nucleotide polymorphism markers linked to QTL for wheat yield traits. Euphytica, 2015, 206: 1-13[本文引用:3]
[24]
KatoK, MiuraH, SawadaS. Mapping QTLs controlling grain yield and its components on chromosome 5A of wheat. Theor Appl Genet, 2000, 101: 1114-1121[本文引用:1]
[25]
张坤普, 徐宪斌, 田纪春. 小麦籽粒产量及穗部相关性状的QTL 定位. 作物学报, 2009, 35: 270-278Zhang KP, Xu XB, Tian JC. QTL Mapping for grain yield and spike related traits in common wheat. Acta Agron Sin, 2009, 35: 270-278 (in Chinese with English abstract)[本文引用:1]
[26]
丁安明, 李君, 崔法, 赵春华, 马航运, 王洪刚. 利用小麦关联RIL 群体定位产量相关性状QTL. 作物学报, 2011, 37: 1511-1524Ding AM, LiJ, CuiF, Zhao CH, Ma HY, Wang HG. QTL mapping for yield related traits using two associated RIL populations of wheat. Acta Agron Sin, 2011, 37: 1511-1524 (in Chinese with English abstract)[本文引用:1]





{kind=link}
{kind=link}