关键词:SA; Cu胁迫; 烟草; 微量元素 Effect of Salicylic Acid on Growth and Nutrient Uptake in Hydroponic Tobacco under Cu Stress XU Gen-Di1,2, GE Shu-Fang1,2, ZAHNG Yi3, WU Yu-Huang4, MEI Xiao-Man5, LIU Peng1,2,* 1 Research Institute of Ecology, Zhejiang Normal University, Jinhua 321004, China
2 Key Laboratory of Botany / Zhejiang Normal University, Jinhua 321004, China
3 Tourism College of Zhejiang, Hanzhou 311231, China
4 College of Life and Environmental Sciences, Hangzhou Normal University, Hangzhou 310036, China
5 College of Teacher Education, Hanzhou Normal University, Hangzhou 330036, China
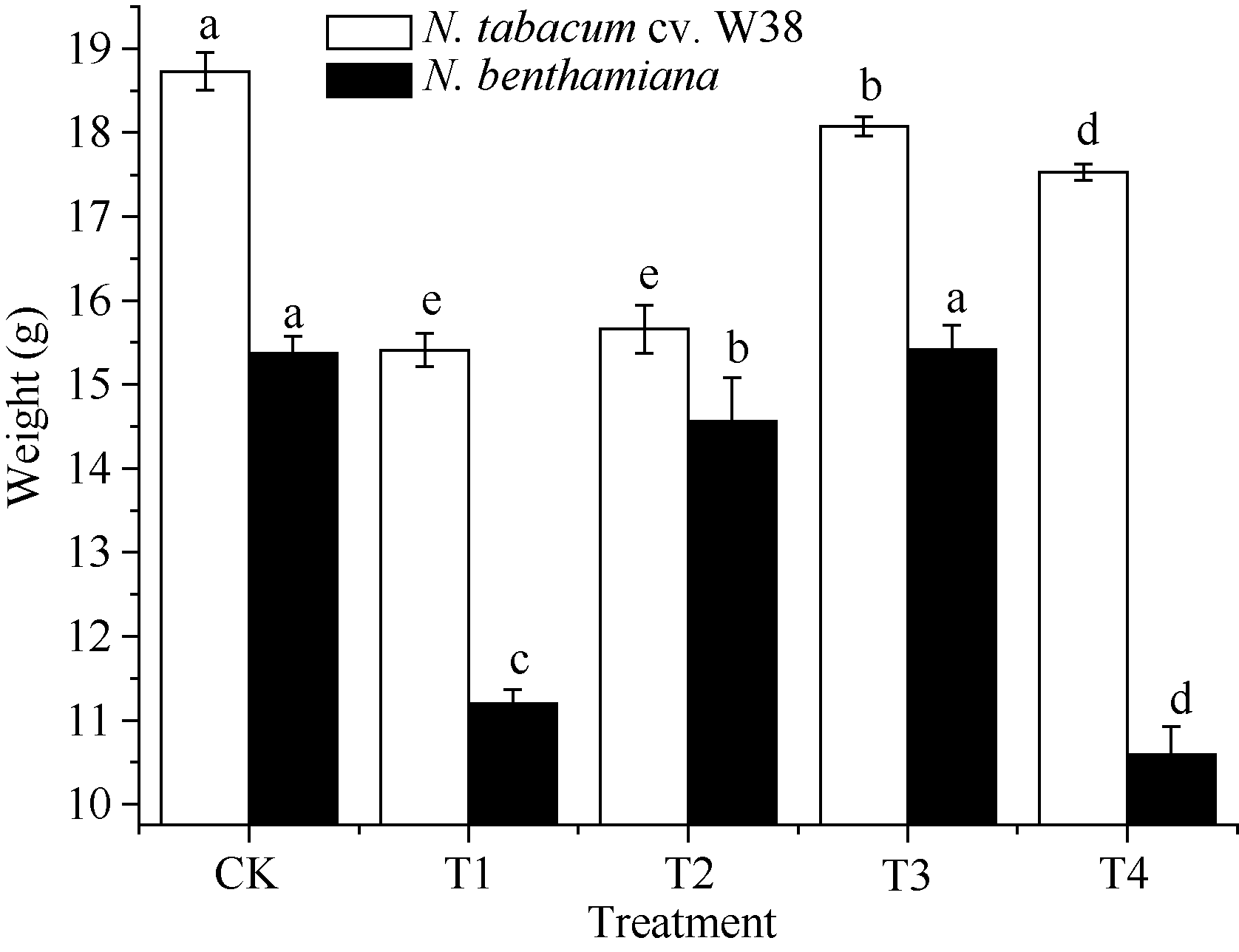
AbstractA hydroponic experiment was conducted to study the effects from treatmeats including CK ( Cu2+ 0 mg L-1, SA 0 μmol L-1 as control), T1 (Cu2+ 4 mg L-1, SA 0 μmol L-1), T2 (Cu2+ 4 mg L-1, SA 100 μmol L-1), T3 (Cu2+ 4 mg L-1, SA 300 μmol L-1), T4 (Cu2+ 4 mg L-1, SA 500 μmol L-1) on growth characteristics, elements absorption and accumulation in two tobacco cultivars, N. tabacum cv .W38 (Cu resistant genotype) and N. benthamiana (Cu sensitive genotype) under copper stress. The result showed that the growth of two tobacco cultivars under 4 mg L-1Cu stress for 15 days was inhibited to some extent, meanwhile, the content of Cu in their organs was increased while the absorptions of K, Ca, Mg, Fe, Zn, B, and Mn were inhibited. The addition of exogenous SA could alleviate the inhibition of plant height, root length and fresh weight, and reduce Cu content of tobacco, but increase the absorption of nutritional elements K, Ca, Mg, Fe, and Mn. Compared with treatment T1, contents of Cu in root, stem and leaf were increased by 25.05%, 39.78%, 22.91% in N. benthamiana, and by 23.27%, 37.30%, 28.88% in N. tabacum cv. W38, respectively. In conclusion, appropriate concentration of SA promotes the growth and elements absorption in tobacco under copper stress. In this experiment, 300 μmol L-1 SA could significantly alleviate copper inhibition to tobacco.
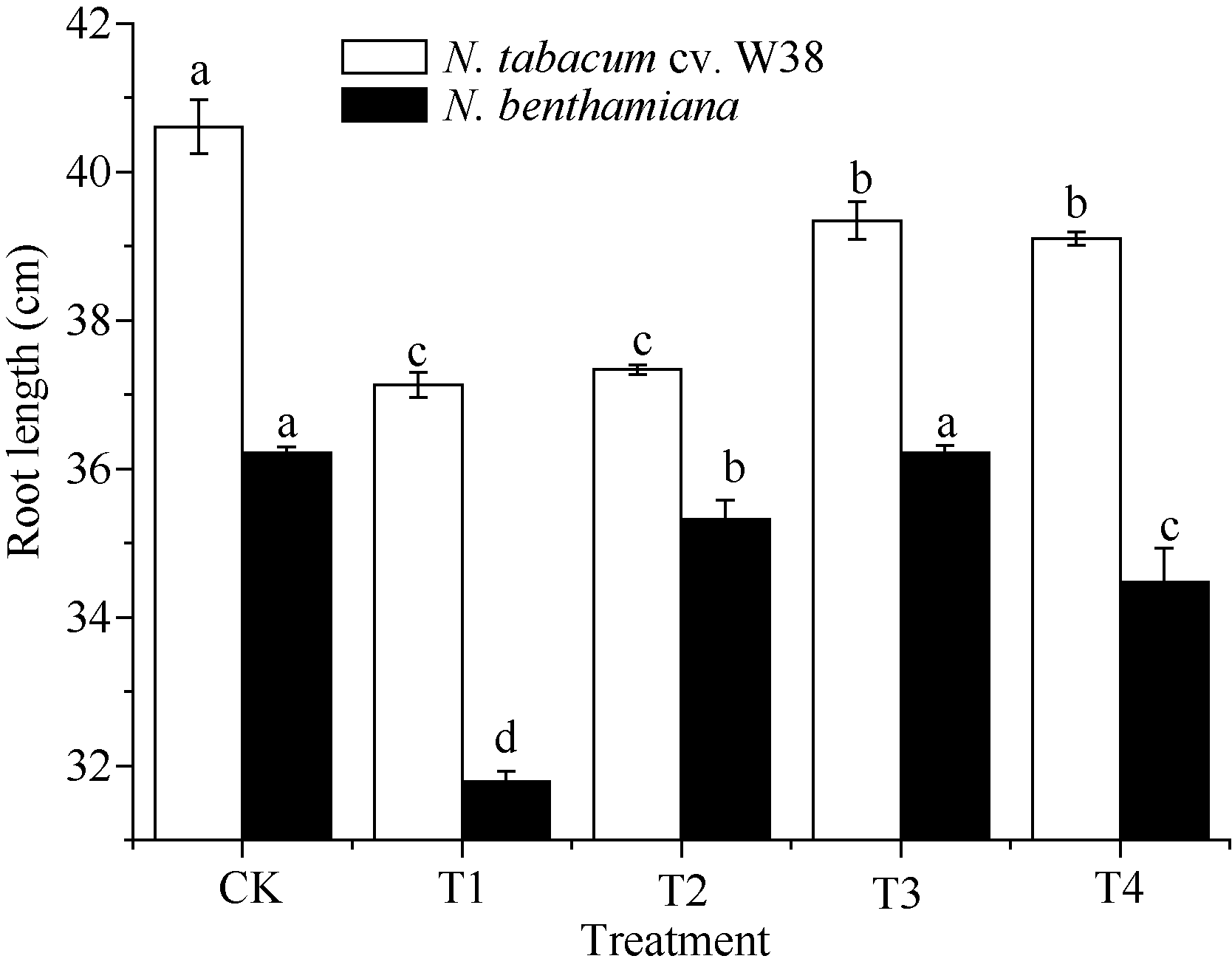
Effects of SA on the root length (mean± SE) of tobacco under copper stress Bars superscripted by different letters are significantly different at the 5% probability level. CK: Cu2+ 0 mg L-1, SA 0 μ mol L-1, T1: Cu2+ 4 mg L-1, SA 0 μ mol L-1; T2: Cu2+ 4 mg L-1, SA 100 μ mol L-1, T3: Cu2+ 4 mg L-1, SA 300 μ mol L-1, T4: Cu2+ 4 mg L-1, SA 500 μ mol L-1.
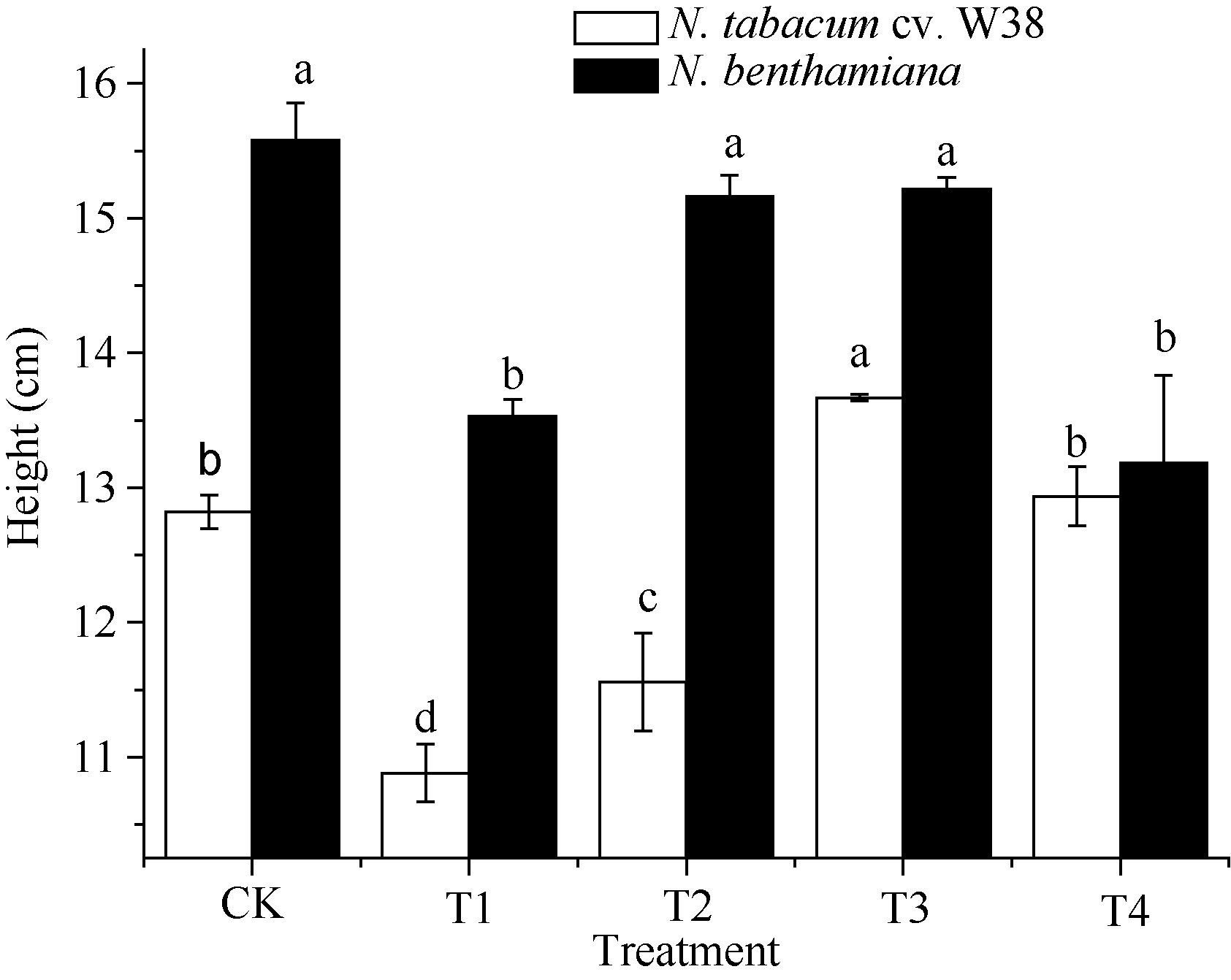
图2 SA对铜胁迫下烟草株高的影响Fig. 2 Effects of SA on plant height (mean± SE) of tobacco under copper stressBars superscripted by different letters are significantly different at the 5% probability level. Treatment codes are the same as those given in Fig. 1.
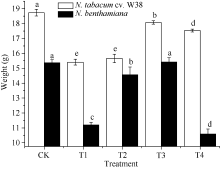
图3 SA对铜胁迫下烟草鲜重的影响Fig. 3 Effects of SA on fresh weight (mean± SE) of tobacco under copper stressBars superscripted by different letters are significantly different at the 5% probability level. Treatment codes are the same as those given in Fig. 1.
表1 SA对铜胁迫下烟草Cu含量的影响 Table 1 Effects of SA on the content of Cu (mean± SE) in tobacco under copper stress (μ g g-1DW)
品种 Variety
部位 Part
CK
T1
T2
T3
T4
W38 N. tabacumcv. W38
根Root
20.47± 0.51 e
150.52± 0.44 a
139.11± 0.49 b
115.49± 0.71 d
130.52± 0.67 c
茎Stem
3.11± 0.10 d
20.16± 0.17 a
18.22± 0.18 b
12.64± 0.36 c
17.84± 0.52 b
叶Leaf
8.38± 0.15 d
60.61± 0.62 a
55.84± 0.19 b
43.10± 0.89 c
42.29± 1.24 c
本氏烟 N. benthamiana
根Root
23.27± 0.35 d
172.87± 0.72 a
129.57± 0.55 c
145.14± 0.12 b
144.89± 0.75 b
茎Stem
7.18± 0.15 d
21.26± 0.85 a
16.78± 0.38 c
18.86± 0.17 b
19.65± 0.35 b
叶Leaf
8.47± 0.19 d
81.79± 0.56 a
63.05± 0.18 c
63.24± 0.27 c
65.90± 0.27 b
Values followed by different letters within a row are significantly different at the 0.05 probability level (P< 0.05). Treatment codes are the same as those given in Fig. 1. 同行数据后的不同字母表示在0.05水平上差异显著。处理代码同图1。
表1 SA对铜胁迫下烟草Cu含量的影响 Table 1 Effects of SA on the content of Cu (mean± SE) in tobacco under copper stress (μ g g-1DW)
表2 SA对铜胁迫下烟草K含量的影响 Table 2 Effects of SA on the content of K (mean± SE) in tobacco under copper stress (μ g g-1 DW)
品种 Variety
部位 Part
CK
T1
T2
T3
T4
W38 N. tabacumcv. W38
根Root
15.32± 0.27 a
8.27± 0.07 c
13.55± 0.20 b
15.32± 0.22 a
15.14± 0.11 a
茎Stem
51.23± 0.22 a
38.35± 0.24 c
46.51± 0.24 b
51.44± 0.58 a
50.85± 0.65 a
叶Leaf
30.59± 0.45 a
19.62± 0.51 d
25.52± 0.25 c
28.42± 0.19 b
28.12± 0.19 b
本氏烟 N. benthamiana
根Root
17.37± 0.93 a
8.75± 0.87 c
17.49± 0.75 a
17.58± 1.25 a
13.90± 0.28 b
茎Stem
45.97± 0.27 a
31.00± 0.70 d
38.45± 0.51 b
45.28± 0.36 a
35.32± 0.28 c
叶Leaf
30.03± 0.65 a
19.43± 0.18 d
28.17± 0.18 b
28.42± 0.19 b
25.72± 0.52 c
Values followed by different letters within a row are significantly different at the 0.05 probability level (P< 0.05). Treatment codes are the same as those given in Fig. 1. 同行数据后的不同字母表示在0.05水平上差异显著。处理代码同图1。
表2 SA对铜胁迫下烟草K含量的影响 Table 2 Effects of SA on the content of K (mean± SE) in tobacco under copper stress (μ g g-1 DW)
表3 Table 3 表3(Table 3)
表3 SA对铜胁迫下烟草Ca、Mg含量的影响 Table 3 Effects of SA on contents of Ca and Mg (mean± SE) in tobacco under copper stress (μ g g-1 DW)
微量元素 Trace element
处理 Treatment
W38 N. tabacumcv. W38
本氏烟 N. benthamiana
根Root
茎Stem
叶Leaf
根Root
茎Stem
叶Leaf
Ca
CK
17.19± 0.05 a
20.45± 0.49 a
25.08± 0.39 a
15.36± 0.16 a
22.33± 0.32 a
30.06± 0.19 a
T1
13.01± 0.77 c
16.37± 0.16 d
23.23± 0.38 c
12.30± 0.08 c
18.23± 0.18 d
25.89± 0.39 c
T2
13.36± 0.22 c
17.10± 0.17 c
23.40± 0.23 c
14.38± 0.19 b
20.27± 0.23 b
27.32± 0.07 b
T3
16.12± 0.15 b
19.19± 0.16 b
25.25± 0.17 a
13.91± 0.94 b
19.62± 0.32 c
27.10± 0.32 b
T4
15.77± 0.18 b
17.38± 0.41 c
24.04± 0.38 b
11.44± 0.47 c
18.39± 0.22 d
24.45± 0.52 d
Mg
CK
7.30± 0.26 a
5.230± 0.10 a
3.61± 0.35 a
7.41± 0.35 a
5.26± 0.06 a
3.27± 0.27 a
T1
6.30± 0.02 bc
4.55± 0.20 b
2.34± 0.27 b
6.23± 0.35 c
4.43± 0.28 cd
2.30± 0.30 bc
T2
6.60± 0.25 b
4.48± 0.12 bc
2.62± 0.04 b
6.83± 0.15 b
4.47± 0.14 c
2.72± 0.25 b
T3
6.42± 0.28 bc
4.60± 0.28 b
2.65± 0.82 b
6.52± 0.20 bc
4.83± 0.16 b
2.68± 0.06 b
T4
6.15± 0.15 c
4.21± 0.10 c
2.37± 0.07 b
6.08± 0.21 c
4.14± 0.15 d
2.18± 0.20 c
Values followed by different letters within a row are significantly different at the 0.05 probability level (P< 0.05). Treatment codes are the same as those given in Fig. 1. 同列数据后的不同字母表示在0.05水平上差异显著。处理代码同图1。
表3 SA对铜胁迫下烟草Ca、Mg含量的影响 Table 3 Effects of SA on contents of Ca and Mg (mean± SE) in tobacco under copper stress (μ g g-1 DW)
表4 SA对铜胁迫下烟草Fe、Zn、B、Mn含量的影响 Table 4 Effects of SA on contents of Fe, Zn, B, and Mn (mean± SE) in tobacco under copper stress (μ g g-1 DW)
微量元素 Trace element
处理 Treatment
W38 N. tabacumcv. W38
本氏烟 N. benthamiana
根Root
茎Stem
叶Leaf
根Root
茎Stem
叶Leaf
Fe
CK
524.59± 1.42 a
120.51± 0.44 a
132.85± 0.72 a
510.51± 0.43 a
102.62± 0.40 a
95.52± 0.50 a
T1
410.84± 1.01 e
62.48± 0.46 d
59.58± 0.55 d
333.13± 0.16 e
82.38± 0.67 d
67.31± 0.29 c
T2
494.36± 0.72 c
97.21± 0.20 b
117.22± 0.08 b
474.61± 0.52 c
91.44± 0.45 b
86.26± 0.29 b
T3
499.44± 0.50 b
97.73± 0.63 b
132.14± 1.01 a
492.51± 0.44 b
87.44± 0.50 c
59.90± 0.28 d
T4
444.12± 0.19 d
79.83± 0.18 c
100.41± 0.53 c
379.18± 0.07 d
81.76± 0.19 d
34.62± 0.40 e
Zn
CK
20.28± 0.20 b
13.10± 0.17 a
18.45± 0.06 a
22.83± 0.56 ab
17.26± 0.17 a
20.10± 0.43 ab
T1
19.46± 0.06 c
12.69± 0.50 b
18.07± 0.20 b
21.82± 0.42 b
16.64± 0.40 b
19.19± 0.47 b
T2
19.60± 0.06 c
12.82± 0.17 ab
18.20± 0.10 ab
22.95± 0.80 a
16.72± 0.23 b
19.76± 0.33 ab
T3
21.29± 0.27 a
11.54± 0.23 d
18.20± 0.13 ab
22.19± 0.59 ab
16.42± 0.10 b
20.80± 0.86 a
T4
19.51± 0.16 c
12.06± 0.24 c
17.33± 0.31 c
17.68± 0.30 c
15.10± 0.10 c
17.27± 0.83 c
B
CK
32.47± 0.10 a
17.35± 0.19 a
25.30± 0.05 a
30.13± 0.45 a
19.43± 0.18 a
21.49± 0.20 a
T1
29.73± 0.44 d
15.22± 0.10 c
23.46± 0.22 c
28.66± 0.69 b
17.43± 0.12 c
20.80± 0.39 b
T2
30.82± 0.34 c
16.40± 0.10 b
23.85± 0.23 b
28.75± 0.16 b
17.24± 0.06 c
20.57± 0.21 b
T3
31.69± 0.39 b
16.40± 0.32 b
25.20± 0.16 a
29.76± 0.20 a
18.65± 0.18 b
21.68± 0.34 a
T4
32.04± 0.08 ab
17.28± 0.14 a
25.39± 0.32 a
28.59± 0.08 b
16.38± 0.15 d
19.26± 0.27 c
Mn
CK
624.52± 0.43 a
181.87± 0.57 a
165.35± 0.10 a
611.48± 0.48 a
165.00± 0.61 a
115.24± 0.07 a
T1
354.25± 0.08 d
74.20± 0.22 e
69.73± 0.45 d
401.51± 0.43 d
69.39± 0.78 e
74.16± 0.05 d
T2
580.32± 1.77 b
169.67± 0.34 c
149.90± 0.29 b
584.92± 0.34 c
149.49± 0.45 b
112.27± 0.06 b
T3
581.53± 0.43 b
172.36± 0.16 b
117.78± 0.67 c
587.33± 0.09 b
117.78± 0.67 c
108.32± 0.30 c
T4
427.62± 0.53 c
95.51± 0.15 d
117.31± 0.10 c
401.25± 1.03 d
73.29± 1.01 d
60.74± 0.46 e
Values followed by different letters within a row are significantly different at the 0.05 probability level (P< 0.05). Treatment codes are the same as those given in Fig. 1. 同一元素、同列数据后的不同字母表示在0.05水平上差异显著。处理代码同图1。
表4 SA对铜胁迫下烟草Fe、Zn、B、Mn含量的影响 Table 4 Effects of SA on contents of Fe, Zn, B, and Mn (mean± SE) in tobacco under copper stress (μ g g-1 DW)
Poll EA, SchzendbelA. Hevey metals ignaling in plants: linking celluar and organismic responses. Topics Curr Genet, 2003, 4: 187-202[本文引用:1]
[2]
高培培, 章艺, 吴玉环, 徐根娣, 刘鹏. 外源水杨酸对铝胁迫下栝楼光合特性及耐铝性的影响. 水土保持学报, 2012, 26(6): 268-273Gao PP, ZhangY, Wu YH, Xu GD, LiuP. Effect of exogenous SA on photosynthesis and Al tolerance of Trichosanthes kirilowii Maxim under aluminum stress. J Soil Water Conserv, 2012, 26(6): 268-273 (in Chinese with English abstract)[本文引用:1][JCR: 1.265]
[3]
张营, 李法云, 严霞. 外源K+和水杨酸在缓解融雪剂对油松幼苗生长抑制中的效应与机理. 生态学报, 2012, 32: 4300-4308ZhangY, Li FY, YanX. Alleviation effect and mechanism of exogenous potassium nitrate and salicylic acid on the growth inhibition of Pinus tabulaeformis seedlings induced by deicing salts. Acta Ecol Sin, 2012, 32: 4300-4308 (in Chinese with English abstract)[本文引用:1]
[4]
HayatQ, HayatS, IrfanM. Effect of exogenous salicylic acid under changing environment. Environ Exp Bot, 2010, 68: 14-25[本文引用:1][JCR: 2.578]
[5]
郝敬虹, 易旸, 尚庆茂, 董春娟, 张志刚. 干旱胁迫下外源水杨酸对黄瓜幼苗膜脂过氧化和光合特性的影响. 应用生态学报, 2012, 23: 717-723Hao JH, YiY, Shang QM, Dong CJ, Zhang ZG. Effects of exogenous salicylic acid on membrane lipid peroxidation and photosynthetic characteristics of Cucumis sativus seedlings under drought stress. Chin J Appl Ecol, 2012, 23: 717-723 (in Chinese with English abstract)[本文引用:1][CJCR: 1.742]
[6]
易小林, 杨丙贤, 宗学凤, 李名扬. 信号分子水杨酸减缓干旱胁迫对紫御谷光合和膜脂过氧化的副效应. 生态学报, 2011, 31: 67-74Yi XL, Yang BX, Zong XF, Li MY. Signal chemical salicylic acid mitigates the negative effects of drought on photosynthesis and membrane lipid peroxidation of purple majesty. Acta Ecol Sin, 2011, 31: 67-74 (in Chinese with English abstract)[本文引用:1]
[7]
Wang XM, Yuan XJ, HanS, Zha HW, Sun XC, Huang JH, Liu YN. Aniline modified hypercrosslinked polystyrene resins and their adsorption equilibriums, kinetics and dynamics towards salicylic acid from aqueous solutions. Chem Eng J, 2013, 233: 124-131[本文引用:1][JCR: 3.473]
[8]
张玉秀, 柴团耀. 植物耐金属机理研究进展. 植物学报, 1999, 41: 453-457Zhang YX, Chai TY. Research advances on the mechanisms of heavy metal tolerance in plants. Acta Bot Sin, 1999, 41: 453-457 (in Chinese with English abstract)[本文引用:1][JCR: 0.599]
[9]
AshrafM, IrisF, ManfredG. Salicylic acid alleviates the cadmium toxicity in barley seedlings. Plant Physiol, 2003, 132: 272-281[本文引用:1][JCR: 6.555]
[10]
刘素纯, 萧浪涛, 廖柏寒, 匡逢春, 黄运湘. 铅、镉胁迫对黄瓜幼苗内源水杨酸的影响. 湖南农业科学, 2006, (3): 25-28Liu SC, Xiao LT, Liao BH, Kuang FC, Huang YX. Effects on endogenous salicylic acid of cucumber seedling sunder lead and cadmium stress. Hunan Agric Sci, 2006, (3): 25-28 (in Chinese with English abstract)[本文引用:1][CJCR: 0.451]
[11]
Freeman JL, GarciaD, KimD, HopfA, Salt DE. Constitutively elevated salicylic acid signals glutathione-mediated nickel tolerance in thlaspinickelhyper accumulators. Plant Physiol, 2005, 137: 1082-1091[本文引用:1][JCR: 6.555]
[12]
DrazicG, MihailovicN. Modification of cadmium toxicity in soybean seedlings by salicylic acid. Plant Sci, 2005, 168: 511-517[本文引用:1][JCR: 2.922]
[13]
张艳英, 周楠, 刘鹏, 徐根娣, 陈文荣, 蔡妙珍. 铜胁迫对烟草幼苗氮代谢的影响. 生态学报, 2009, 29: 6779-6784Zhang YY, ZhouN, LiuP, Xu GD, Chen WR, Cai MZ. Effect of Cu stress on nitrogen metabolism of Nicotiana tabacum L. seedling. Acta Ecol Sin, 2009, 29: 6779-6784 (in Chinese with English abstract)[本文引用:1]
[14]
张小梅, 王智鑫, 李广录. 水杨酸对铬胁迫下绿豆幼苗生长抑制的缓解效应. 河南农业科学, 2012, 41(11): 52-54Zhang XM, Wang ZX, Li GL. Mitigating effects of SA on inhibition of Cr2+ stress to mung bean seedling growth. Henan Agric Sci, 2012, 41(11): 52-54 (in Chinese with English abstract)[本文引用:1]
[15]
计汪栋, 施国新, 张慧, 徐勤松, 许晔, 杜开和. 菹草对Hg2+胁迫的生理和结构应答反应. 生态学报, 2007, 27: 2856-2863Ji WD, Shi GX, ZhangH, Xu QS, XuY, Du KH. Physiolgical and ultrastructural responses of Potamogeton crispus to Hg2+ stress. Acta Ecol Sin, 2007, 27: 2856-2863 (in Chinese with English abstract)[本文引用:1]
[16]
DelhaizeE. Uptake on environmental stress: aluminum toxicity and tolerance in plants. Plant Physiol, 1995, 107: 315-321[本文引用:1][JCR: 6.555]
[17]
郑国红, 胡婵娜, 刘鹏. 外源钾对铁胁迫下水稻元素吸收运输规律的影响. 水土保持学报, 2010, 24(5): 141-145Zheng GH, Hu CN, LiuP. Effects of exogenous kalium on elements absorption and transportation of rice under iron toxicity. J Soil Water Conserv, 2010, 24(5): 141-145 (in Chinese with English abstract)[本文引用:1][JCR: 1.265]
[18]
PeterR. Interaction between aluminum toxicity and calcium uptake at the root apex in near-isogenies lines of wheat (Triticum aestivan L. ) differing in aluminum tolerance. Plant Physiol, 1993, 102: 975-982[本文引用:1][JCR: 6.555]
[19]
DrewmC. Oxygen deficiency and root metabolism: injury and acceliation under hypoxia and anoxia. Annu Rev Plant Physiol Mol Biol, 1997, 48: 223-250[本文引用:1]
[20]
Link HR, Weng CH, Lo HF. Study of the root antioxidative system of tomatoes and eggplants under water logged conditions. Plant Sci, 2004, 167: 355-366[本文引用:1][JCR: 2.922]
[21]
WangC, Zhang SH, Wang PF, HouJ, Zhang WJ, LiW, Lin ZP. The effect of excess Zn onmineral nutrition and antioxidative response in rapeseed seedlings. Chemosphere, 2009, 75, 1468-1476[本文引用:1][JCR: 3.137]
[22]
周媛. 水杨酸对铝胁迫下栝楼生长的调控机理研究. 浙江师范大学硕士学位论文, 浙江金华, 2012ZhouY. The Mechanism of Salicylic Acid Regulating Aluminum Stress in Trichosathe skirilowii Maxim. MS Thesis of Zhejiang Normal University, Jinhua, China, 2012 (in Chinese with English abstract)[本文引用:1]
 , 葛淑芳
, 葛淑芳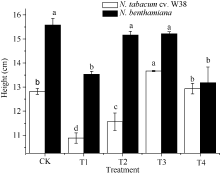

{kind=link}
{kind=link}