关键词:籼粳杂交稻; 甬优系列; 根系形态生理 Root System Morphological and Physiological Characteristics of Indica- japonica Hybrid Rice of Yongyou Series JIANG Yuan-Hua, XU Jun-Wei, ZHAO Ke, WEI Huan-He, SUN Jian-Jun, ZHANG Hong-Cheng*, DAI Qi-Gen, HUO Zhong-Yang, XU Ke, WEI Hai-Yan, GUO Bao-Wei Innovation Center of Rice Cultivation Technology in the Yangtze Valley, Ministry of Agriculture / Key Laboratory of Crop Genetics and Physiology of Jiangsu Province, Yangzhou University, Yangzhou 225009, China
AbstractA field experiment was conducted to compare the root system morphological and physiological characteristics of four types of rice populations including indica-japonica hybrid rice of Yongyou series (A), japonica hybrid rice (B), conventional japonica rice (C), and indica hybrid rice (D) under high-yielding cultivation condition in the rice-wheat cropping system. Results were as follows: (1) The root dry weight, aboveground dry weight, number of root tips, root length, root surface area, root-shoot ratio and root volume of A were obviously higher than those of B, C, and D at the middle and late growth stage. (2) The proportion of number, length, surface area and volume of the adventitious roots(root diameter > 0.3 mm) to the whole roots of A were higher than those of B and C, while lower than those of D at heading; the proportion of number, length, surface area and volume of the fine branches (root diameter ≤ 0.1 mm) and coarse branches (0.1 mm < root diameter ≤ 0.3 mm) to total roots of A were higher than those of D, while lower than those of B and C at heading. The proportions of root weight in 0-5 cm, 5-10 cm, and 10-15 cm soil layers to total roots weight of A were higher than those of B and C, while lower than those of D; however, the proportions of root weight in 15-25 cm, 25-35 cm, 35-45 cm, and 45-55 cm soil layers to the total root weight where lower than those of B and C, while higher than these of D at heading. (3) The total absorbing surface area, active absorbing surface area, bleeding intensity, root oxidation activity, and root reducing activity of A were higher than those of B, C, and D. Compared with B, C, and D, the indica-japonica hybrid rice of Yongyou series had significant advantage in root-shoot coordination level, root amount, branch structure, root distribution in soil and biological activity in the middle and late growth stage. This peculiarity of A provides an important guarantee to realize the super-high-yield.
Keyword:Indica-japonica hybrid rice; Yongyou series; Morphology and physiology of roots Show Figures Show Figures
表1 不同类型水稻品种产量及其构成 Table 1 Difference of grain yield and its components in different types of rice cultivars
年份 Year
类型 Type
有效穗数 No. of panicles (× 104 hm-2)
每穗粒数 Spikelets per panicle
总颖花量 Total spikelets (× 104 hm-2)
结实率 Seed-setting rate (%)
千粒重 1000-grain weight (g)
实际产量 Actual yield (kg hm-2)
2012
A
200.98 d
314.02 a
63093.94 a
84.01 c
24.19 b
12152.21 a
B
241.57 b
221.66 b
53494.86 b
86.07 b
26.33 a
11528.21 b
C
329.65 a
143.43 d
47242.65 d
91.26 a
26.56 a
10901.20 c
D
238.89 c
205.84 c
49138.74 c
82.32 d
26.57 a
10140.64 d
2013
A
203.63 c
299.36 a
60959.55 a
83.69 c
24.36 b
12026.77 a
B
246.97 b
212.55 b
52493.00 b
85.96 b
26.54 a
11385.83 b
C
336.95 a
137.53 d
46339.60 d
90.84 a
26.61 a
10742.21 c
D
243.10 b
196.23 c
47704.37 c
81.86 d
26.76 a
10043.47 d
A: indica-japonica hybrid rice; B: japonica hybrid rice; C: conventional japonica rice; D: indicahybrid rice. Values followed by different letters are significantly different at the 0.05 probability level. A: 籼粳杂交稻; B: 杂交粳稻; C: 常规粳稻; D: 杂交籼稻。数字后跟不同小写字母表示在0.05水平上差异显著。
表1 不同类型水稻品种产量及其构成 Table 1 Difference of grain yield and its components in different types of rice cultivars
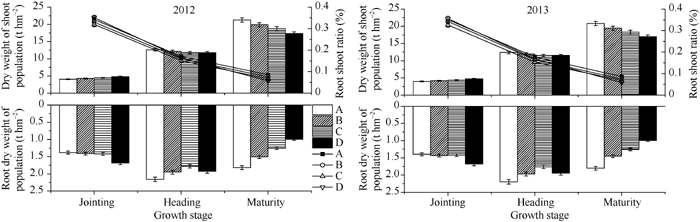
表2 不同类型水稻品种群体根系的形态特征 Table 2 Root system morphological characteristics of populations in different types of rice cultivars
年份 Year
类型 Type
根尖数 Number of tips (× 106 tips hm-2)
长度 Length (× 103 m hm-2)
表面积 Surface area (× 102 m2 hm-2)
体积 Volume (× 102 m3 hm-2)
拔节期Jointing
2012
A
26927.53 d
9507.00 d
820.99 a
17.71 d
B
27353.82 c
9603.94 c
808.65 b
18.23 c
C
27675.06 b
9661.07 b
791.82 c
18.69 b
D
31756.82 a
11107.10 a
812.86 b
22.35 a
2013
A
26773.24 d
9419.91 d
694.37 a
18.58 d
B
28208.20 c
10175.11 c
779.50 b
19.31 c
C
28907.54 b
10091.31 b
846.60 c
19.52 b
D
32943.02 a
11521.97 a
843.22 b
23.19 a
抽穗期Heading
2012
A
40882.08 a
17253.67 a
1846.64 a
26.71 a
B
36281.56 c
15366.48 c
1700.33 b
23.25 c
C
32605.26 d
13856.47 d
1581.22 d
20.50 d
D
37259.80 b
15642.03 b
1617.34 c
24.83 b
2013
A
43261.31 a
18257.79 a
1954.11 a
28.27 a
B
38197.05 c
16177.75 c
1790.10 b
24.48 c
C
33694.91 d
14319.54 d
1634.06 d
21.19 d
D
39121.31 b
16423.51 b
1698.14 c
26.07 b
成熟期Maturity
2012
A
33162.28 a
12389.51 a
1219.55 a
23.51 a
B
26888.34 b
10209.68 b
1154.91 b
18.67 b
C
22053.76 c
8503.08 c
1077.91 c
15.00 c
D
18472.15 d
6833.09 d
564.35 d
13.48 d
2013
A
34299.63 a
12814.43 a
1261.38 a
24.32 a
B
27046.61 b
10269.78 b
1161.71 b
18.78 b
C
23122.18 c
8915.02 c
1130.14 c
15.73 c
D
19254.60 d
7122.53 d
588.25 d
14.05 d
Values followed by different letters are significantly different at the 0.05 probability level. Abbreviations are the same as those given in Table 1. 数字后不同小写字母表示在0.05水平上差异显著。缩写同表1。
表2 不同类型水稻品种群体根系的形态特征 Table 2 Root system morphological characteristics of populations in different types of rice cultivars
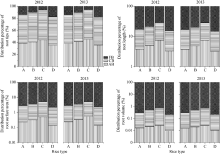
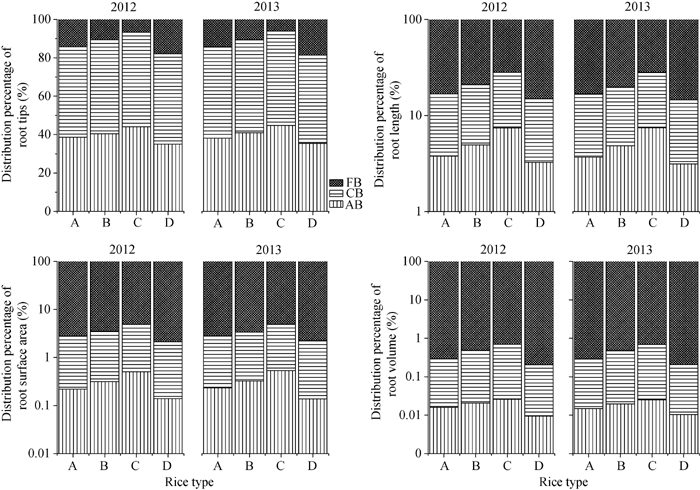
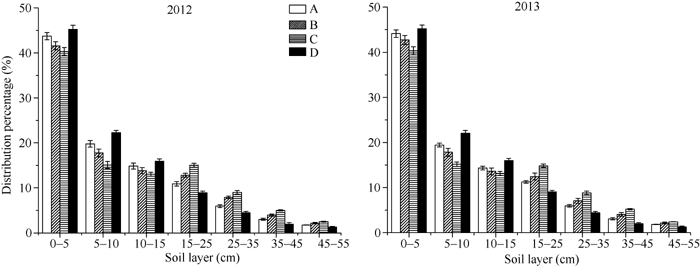
图2 不同类型水稻品种抽穗期根数分布比例其他缩写同图1。Fig. 2 Distribution of different percentage of different pants of roots in different types of rice cultivarsFB: fine branch; CB: course branch; AB: adventitious branch.Other abbreviations are the same as those given in Figure 1.
图3 不同类型水稻品种抽穗期根系干重分布比例缩写同图1。Fig. 3 Distribution percentage of roots in different types of rice cultivarsAbbreviations are the same as those given in Figure 1.
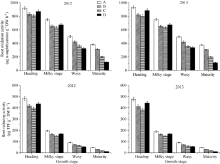
表3 不同类型品种群体根系总吸收面积与活跃吸收面积 Table 3 Total absorbing surface area, active absorbing surface area and ratio of active absorbing surface area to total absorbing surface area in different types of rice cultivars
类型Type
群体根系总吸收面积 Total absorbing surface area of population (× 104 m2 hm-2)
群体根系活跃吸收面积 Active absorbing surface area of population (× 104 m2 hm-2)
拔节期 Jointing
抽穗期 Heading
成熟期 Maturity
拔节期 Jointing
抽穗期 Heading
成熟期 Maturity
2012
A
896.95 b
1643.14 a
899.32 a
495.12 b
941.52 a
327.17 a
B
862.70 c
1518.31 c
816.06 b
484.68 c
882.14 b
286.85 b
C
813.54 d
1405.22 d
757.75 c
466.62 d
835.30 c
260.98 c
D
942.27 a
1562.47 b
510.97 d
508.05 a
884.40 b
171.08 d
2013
A
880.99 b
1631.25 a
885.85 a
487.69 b
938.12 a
323.60 a
B
857.79 c
1504.82 c
789.91 b
478.65 c
874.69 b
278.82 b
C
801.42 d
1400.11 d
753.00 c
457.64 d
826.71 c
260.78 c
D
936.09 a
1562.49 b
491.67 d
501.51 a
882.47 b
161.37 d
Values followed by different letters are significantly different at the 0.05 probability level. Abbreviations are the same as those given in Table 1. 数字后跟不同小写字母表示在0.05水平上差异显著。缩写同表1。
表3 不同类型品种群体根系总吸收面积与活跃吸收面积 Table 3 Total absorbing surface area, active absorbing surface area and ratio of active absorbing surface area to total absorbing surface area in different types of rice cultivars
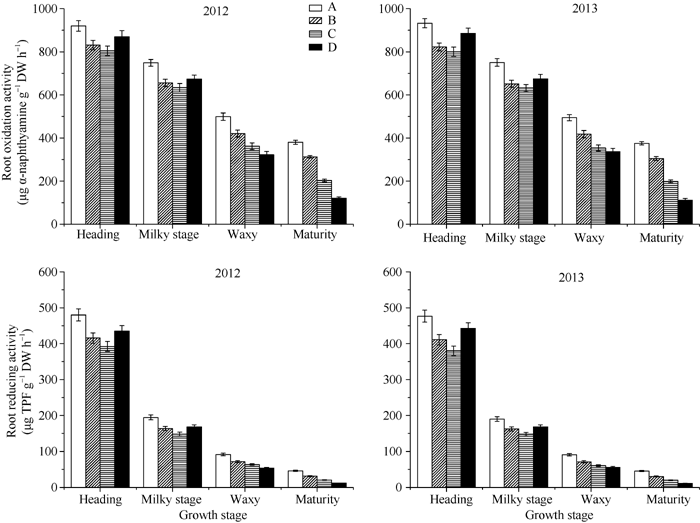
图4 不同类型品种抽穗后根系α -萘胺氧化力和TTC还原力缩写同图1。Fig. 4 Oxidation ability and reducing capacity in different types of rice cultivars after headingAbbreviations are the same as those given in Figure 1.
图5 不同类型品种抽穗后群体根系伤流强度缩写同图1。Fig. 5 Root bleeding intensity in different types of rice cultivars after headingAbbreviations are the same as those given in Figure 1.
4 结论甬优系列籼粳杂交稻根系融合了籼、粳亚种根系的优点, 具有较高的根冠协调水平, 在生育中后期能在群体水平上形成比籼稻更为庞大的根量和形态, 在根系构型及分布上吸取了粳稻分支多、深扎性好的特点, 同时克服籼稻生育后期早衰的弊端, 在整个灌浆期保持较强劲的根系生理活性, 一定程度上满足了超高产群体对肥水的需求, 增强了群体安全支撑性能, 为灌浆期地上部光合系统持续产出、养分水分高效利用、籽粒灌浆有效充分及产量优势的形成提供了重要保障。 The authors have declared that no competing interests exist.
FitterA. Characteristics and functions of root systems. In: Waisel Y, Eshel A, Kafkafi U, eds. Plant Roots, the Hidden Half. New York: Marcel Dekker Inc. , 2002. pp 15-32[本文引用:1]
[2]
InukaiY, AshikariM, KitanoH. Function of the root system and molecular mechanism of crown root formation in rice. Plant Cell Physiol, 2004, 45(suppl): 17[本文引用:]
[3]
ArimaS, SaishoK, HaradaJ. Morphological analysis of the rice root system based on root diameter. Jpn J Crop Sci, 2001, 70: 408-417[本文引用:1]
[4]
ShimizuH, TanabataT, Xie XZ, InagakiN, TakanoM, ShinomuraT. Physiological function of phytochromes in seminal root growth of rice seedlings. Plant Cell Physiol, 2006, 47(suppl): 203-206[本文引用:1][JCR: 4.134]
[5]
朱德峰, 林贤青, 曹卫星. 水稻深层根系对生长和产量的影响. 中国农业科学, 2001, 34: 429-432Zhu DF, Lin XQ, Cao WX. Effects of deep roots on growth and yield in two rice varieties. Sci Agric Sin, 2001, 34: 429-432 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[6]
何强, 邓华凤, 舒服, 杨益善, 刘国华, 刘建丰, 陈立云. 杂交水稻苗期发根性状与生育后期根系活力及穗部性状的关系. 杂交水稻, 2006, 21(3): 75-77HeQ, Deng HF, ShuF, Yang YS, Liu GH, Liu JF, Chen LY. Correlation of rooting traits in seedling stage to activity of root system in late growth stage and panicle traits in hybrid rice. Hybrid Rice, 2006, 21(3): 75-77 (in Chinese with English abstract)[本文引用:1][CJCR: 0.605]
[7]
石庆华, 李木英, 许锦彪, 谭雪明. 高温胁迫对早稻根系质膜ATPase活性及NH4+吸收的影响. 作物学报, 2006, 32: 1044-1048Shi QH, Li MY, Xu JB, Tan XM. Effects of high temperature stress on ATPase activity of plasma membrane and NH4+ absorption rate in roots of early rice. Acta Agron Sin, 2006, 32: 1044-1048 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[8]
YangL, WangY, KobayashiK, ZhuJ, HuangJ, YangH, WangY, DongG, LiuG, HanY, ShanY, HuJ, ZhouJ. Seasonal changes in the effects of free-air CO2 enrichment (FACE) on growth, morphology and physiology of rice root at three levels of nitrogen fertilization. Global Change Biol, 2008, 14: 1-10[本文引用:1][JCR: 6.91]
曾翔, 李阳生, 谢小立, 肖国樱, 廖江林. 不同灌溉模式对杂交水稻生育后期根系生理特性和剑叶光合特性的影响. 中国水稻科学, 2003, 17: 355-359ZengX, Li YS, Xie XL, Xiao GY, Liao JL. Effects of different irrigation patterns on physiological characteristics of root and photosynthetic traits of flag leaf after flowering stage in hybrid rice. Chin J Rice Sci, 2003, 17: 355-359 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[11]
张洪程, 张军, 龚金龙, 常勇, 李敏, 高辉, 戴其根, 霍中洋, 许轲, 魏海燕. “籼改粳”的生产优势及其形成机理. 中国农业科学, 2013, 46: 686-704Zhang HC, ZhangJ, Gong JL, ChangY, LiM, GaoH, Dai QG, Huo ZY, XuK, Wei HY. The productive advantages and formation mechanisms of “indica rice to japonica rice”. Sci Agric Sin, 2013, 46: 686-704 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[12]
李敏, 张洪程, 杨雄, 葛梦婕, 马群, 魏海燕, 戴其根, 霍中洋, 许轲, 曹利强, 吴浩. 水稻高产氮高效型品种的根系形态生理特征. 作物学报, 2012, 38: 648-656LiM, Zhang HC, YangX, Ge MJ, MaQ, Wei HY, Dai QG, Huo ZY, XuK, Cao LiQ, WuH. Root morphological and physiological characteristics of rice cultivars with high yield and high nitrogen use efficiency. Acta Agron Sin, 2012, 38: 648-656 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[13]
董桂春, 陈琛, 王熠, 仲军, 袁秋梅, 羊彬, 于小风, 李进前, 田昊, 张燕, 蒋亚明, 孟令响, 王余龙. 生育期与粳稻品种根系性状的关系分析. 中国水稻科学, 2013, 27: 398-404Dong GC, ChenC, WangY, ZhongJ, Yuan QM, YangB, Yu XF, Li JQ, TianH, ZhangY, Jiang YM, Meng LX, Wang YL. Chin J Rice Sci, 2013, 27: 398-404 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[14]
Li RH, Jiang TB, Xu CG, Li XH, Wang XK. Relationship between morphological and genetic differentiation in rice (Oryza sativa L. ). Euphytica, 2000, 114: 1-8[本文引用:1][JCR: 1.643]
[15]
马荣荣, 许德海, 王晓燕, 禹盛苗, 金千瑜, 欧阳由男, 朱练峰. 籼粳亚种间杂交稻甬优6号超高产株形特征与竞争优势分析. 中国水稻科学, 2007, 21: 281-286Ma RR, Xu DH, Wang XY, Yu SM, Jin QY, Ou-YangY N, ZhuL F. Heterosis on plant morphology of Yongyou6, an indica japonica inter subspecific super high yielding hybrid rice. Chin J Rice Sci, 2007, 21: 281-286 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[16]
许德海, 王晓燕, 马荣荣, 禹盛苗, 朱练峰, 欧阳由男, 金千瑜. 重穗型籼粳杂交稻甬优6号超高产生理特性. 中国农业科学, 2007, 21: 281-286Xu DH, Wang XY, Ma RR, Yu SM, Zhu LF, Ou-Yang Y N, Jin Q Y. Analysis on physiological properties of the heavy panicle type of indica-japonica inter-subspecific hybrid rice Yongyou 6. Sci Agric Sin, 2007, 21: 281-286(in Chinese with English abstract)[本文引用:3][CJCR: 1.889]
[17]
SamejimaH, KondoM, ItoO, NozoeT, ShinanoT, OsakiM. Root-shoot interaction as a limiting factor of biomass productivity in new tropical rice lines. Soil Sci Plant Nutr, 2004, 50: 545-554[本文引用:1][JCR: 0.889]
[18]
SamejimaH, KondoM, ItoO, NozoeT, ShinanoT, OsakiM. Characterization of root systems with respect to morphological traits and nitrogen-absorbing ability in the new plant type of tropical rice lines. J Plant Nutr, 2005, 28: 835-850[本文引用:1][JCR: 0.526]
[19]
ZhangH, Huang ZH, Wang JC, Wang ZQ, Yang JC. Changes in morphological and physiological traits of roots and their relationships with grain yield during the evolution of mid-season indica rice cultivars in Jiangsu Province. Acta Agron Sin, 2011, 37: 1020-1030[本文引用:1][CJCR: 1.667]
[20]
ZhangH, Xue YG, Wang ZQ. Morphological and physiological traits of roots and their relationships with shoot growth in super rice. Field Crops Res, 2009, 113: 31-40[本文引用:1][JCR: 2.474]
[21]
潘晓华, 王永锐, 傅家瑞. 水稻根系生长生理的研究进展. 植物学通报, 1996, 13(2): 13-20Pan XH, Wang YR, Fu JR. Advance in the study on the growth-physiology in rice of root system (Oryza sativa). Chin Bull Bot, 1996, 13(2): 13-20 (in Chinese with English abstract)[本文引用:1]
[22]
刘文兆, 李秧秧. 断伤作物根系对籽粒产量与水分利用效率的影响研究现状及问题. 西北植物学报, 2003, 23: 1320-1324Liu WZ, Li YY. Effect of crop root-cutting on grain yield and water use efficiency: a review. Acta Bot Boreali-Occident Sin, 2003, 23(8): 1320-1324 (in Chinese with English abstract)[本文引用:1][CJCR: 1.321]
[23]
汪强, 樊小林, 刘芳, 李方敏, KlausD, SattemacherB. 断根和覆草旱作条件下水稻的产量效应. 中国水稻科学, 2004, 18: 437-442WangQ, Fan XL, LiuF, Li FM, KlausD, SattemacherB. Effect of root cutting on rice yield by shifting normal paddy to upland cultivation. Chin J Rice Sci, 2004, 18: 437-442 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[24]
郎有忠, 杨建昌, 朱庆森. 亚种间杂交稻根系形态生理特征及其与籽粒充实度关系的研究. 作物学报, 2003, 29: 230-235Lang YZ, Yang JC, Zhu QS. Studies on the morphological and physiological characteristics of the root system of interspecific hybrid rice and their relationship with grain plumpness. Acta Agron Sin, 2003, 29: 230-235 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[25]
徐其军, 汤亮, 顾东祥, 朱艳. 基于形态参数的水稻根系三维建模及可视化. 农业工程学报, 2010, 26(10): 188-194Xu QJ, TangL, Gu DX, ZhuY. Architectural parameter-based three dimensional modeling and visualization of rice roots. Trans CSAE, 2010, 26(10): 188-194 (in Chinese with English abstract)[本文引用:1][CJCR: 1.299]
[26]
顾东祥, 汤亮, 徐其军, 雷晓俊, 曹卫星, 朱艳. 水氮处理下不同品种水稻根系生长分布特征. 植物生态学报, 2011, 35: 558-566Gu DX, TangL, Xu QJ, Lei XJ, Cao WX, ZhuY. Root growth and distribution in rice cultivars as affected by nitrogen and water supply. Chin J Plant Ecol, 2011, 35: 558-566 (in Chinese with English abstract)[本文引用:5][CJCR: 1.989]
[27]
戢林, 李廷轩, 张锡洲, 余海英. 氮高效利用基因型水稻根系形态和活力特征. 中国农业科学, 2012, 45: 4770-4781JiL, Li TX, Zhang XZ, Yu HY. Root morphological and activity characteristics of rice genotype with high nitrogen utilization efficiency. Sci Agric Sin, 2012, 45: 4770-4781 (in Chinese with English abstract)[本文引用:2][CJCR: 1.889]
[28]
MoritaS, IwabuchiA, YamazakiK. Relationships between the growth direction of primary roots and yield in rice plants. Jpn J Crop Sci, 1996, 55: 520-525[本文引用:3]
[29]
李杰, 张洪程, 常勇, 龚金龙, 胡雅杰, 龙厚元, 戴其根, 霍中洋, 许轲, 魏海燕, 高辉. 高产栽培条件下种植方式对超级稻根系形态生理特征的影响. 作物学报, 2011, 37: 2208-2220LiJ, Zhang HC, ChangY, Gong JL, Hu YJ, Long HY, Dai QG, Huo ZY, XuK, Wei HY, GaoH. Influence of planting methods on root system morphological and physiological characteristics of super rice under high-yielding cultivation condition. Acta Agron Sin, 2011, 37: 2208-2220 (in Chinese with English abstract)[本文引用:2][CJCR: 1.667]
[30]
YoshidaS. Relation between rice plant type and root growth. Soil Sci Plant Nutr, 1982, 28: 473-482[本文引用:3][JCR: 0.889]
[31]
吴伟明, 宋祥甫, 孙宗修, 于永红, 邹国燕. 不同类型水稻的根系分布特征比较. 中国水稻科学, 2001, 15: 276-280Wu WM, Song XF, Sun ZX, Yu YH, Zou GY. Comparison of root distribution between different type rice. Chin J Rice Sci, 2001, 15: 276-280 (in Chinese with English abstract)[本文引用:4][CJCR: 1.494]
[32]
许明, 贾德涛, 陈温福. 北方粳型超级稻根系生长与地上部分生物量的变化. 华北农学报, 2010, 25(3): 140-144XuM, Jia DT, Chen WF. Change on root growth and acrial part biomass in north China japonica super rice. Acta Agric Boreali- Sin, 2010, 25(3): 140-144 (in Chinese with English abstract)[本文引用:1][CJCR: 0.951]
[33]
沈波, 王熹. 两个亚种间杂交稻组合的根系生理活性. 中国水稻科学, 2002, 16: 146-150ShenB, WangX. Physiological activities of root system in two iner-subspecific hybrid rice combinations. Chin J Rice Sci, 2002, 16: 146-150 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
[34]
付景, 陈露, 黄钻华, 王志琴, 杨建昌. 超级稻叶片光合特性和根系生理性状与产量的关系. 作物学报, 2012, 38: 1264-1276FuJ, ChenL, Huang ZH, Wang ZQ, Yang JC. Relationship of leaf photosynthetic characteristics and root physiological traits with grain yield in super rice. Acta Agron Sin, 2012, 38: 1264-1276 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
 , 许俊伟, 赵可, 韦还和, 孙建军, 张洪程
, 许俊伟, 赵可, 韦还和, 孙建军, 张洪程




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}