关键词:陆地棉; 枯萎病; Solexa测序; 转录因子 Expression Changes of Transcription Factors in Susceptible and Resistant Upland Cotton ( Gossypium hirsutumL.) Cultivars in Response to Fusarium wilt HAN Ze-Gang1, ZHAO Zeng-Qiang1, HE Lan-Lan1, CHAI Meng-Liang2, LI Hui-Hui1, ZHANG Wei1,* 1Agricultural College, Shihezi University, Shihezi 832000, China
2 College of Life Sciences, Shihezi University, Shihezi 832000, China
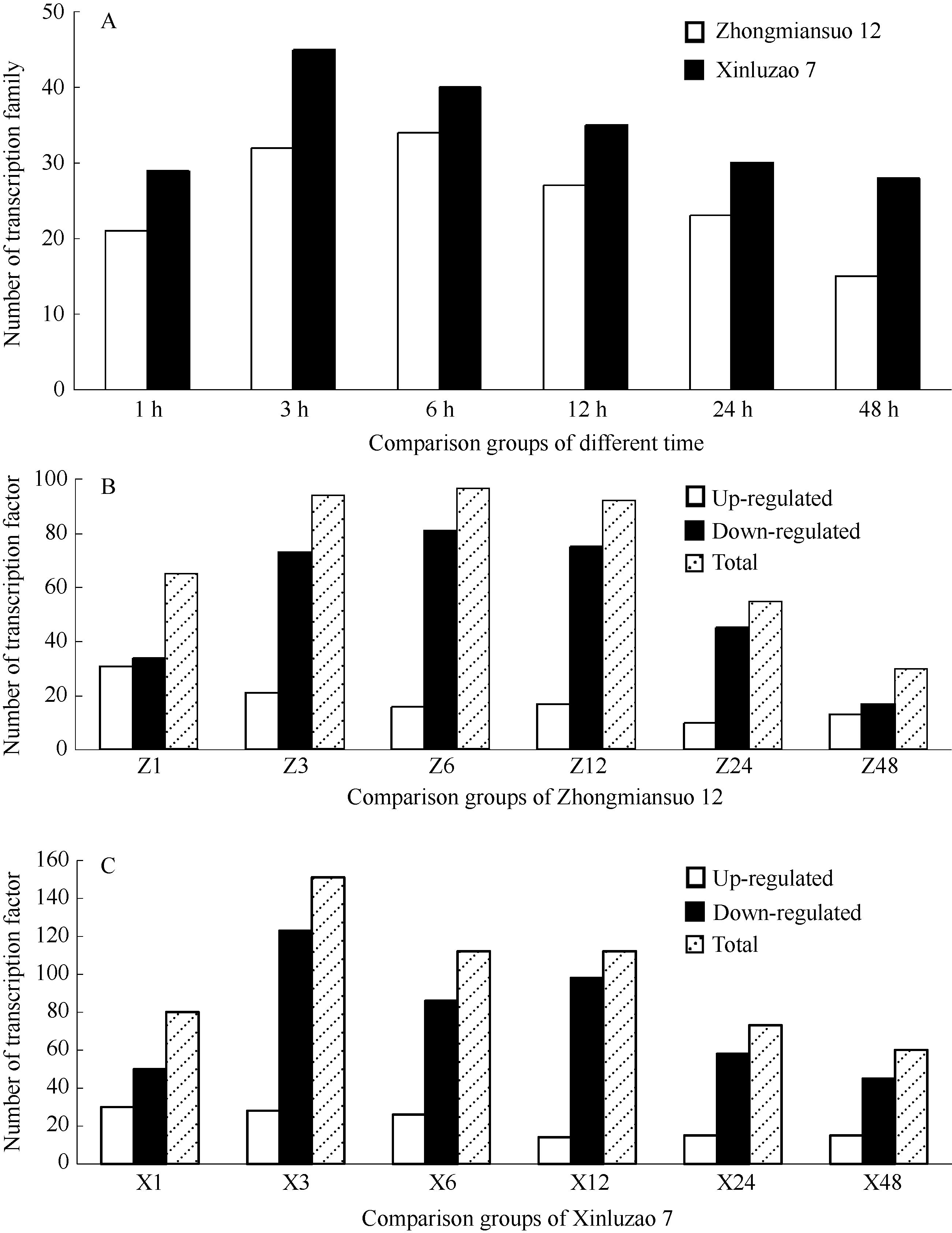
AbstractThe Solexa sequencing technique was used to study the expression changes of transcription factors and their families in upland cotton cultivars susceptible and resistant to Fusarium wilt. A total of 39 transcription factor families and 433 transcription factors in Zhongmiansuo 12 and 52 transcription factor families and 588 transcription factors in Xinluzao 7 were changed in expression in at least one comparison group induced by Fusarium wilt. The number of transcription factor families responsed to Fusarium wilt was more in Xinluzao 7 than in Zhongmiansuo 12, and the number of down-regulated genes was more than that of up-regulated genes in two varieties. After the induction by Fusarium wilt, in the process of time, the number of transcription factor families and transcription factors responsed to the Fusarium wilt in two cultivars showed the same changing trend of increase first and decrease then. The maximum number was reached after six hours in Zhongmiansuo 12, while at three hours after the inducing in Xinluzao 7. Among six comparison groups, there were nine overlapping transcription factors in six transcription factor families of Zhongmiansuo 12 and 31 overlapping transcription factors in 17 transcription factor families of Xinluzao 7. The expression of transcription factor showed strong cultivar specificity among varieties tolerant to Fusarium wilt. Except for 37 transcription factor families shared between two cultivars, two transcription factor families were unique to Zhongmiansuo 12 and Zhongmiansuo 15 transcription factor families were unique to Xinluzao 7.
Keyword:Upland cotton; Fusarium wilt; Solexa sequencing; Transcription factor Show Figures Show Figures
吴征彬, 杨业华, 刘小丰, 王强. 枯萎病对棉花产量和纤维品质的影响. 棉花学报, 2004, 16: 236-239Wu ZB, Yang YH, Liu XF, WangQ. Effect of Fusarium wilt on the cotton yield and fiber quality. Cotton Sci, 2004, 16: 236-239 (in Chinese with English abstract)[本文引用:1][CJCR: 1.462]
2
徐秋华, 张献龙, 聂以春, 冯纯大. 我国棉花抗枯萎病品种的遗传多样性分析. 中国农业科学, 2002, 35: 272-276Xu QH, Zhang XL, Nie YC, Feng CD. Genetic diversity evaluation of cultivars (G. hirsumtum L. ) resistant to Fusarium wilt by RAPD markers. Sci Agric Sin, 2002, 35: 272-276 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
3
金慧, 栾雨时. 转录因子在植物抗病基因工程中的研究进展. 中国生物工程杂志, 2010, 30(10): 94-99JinH, Luan YS. Progress of transcription factor in gene engineering of diseases resistances in plant. Chin Biotechnol, 2010, 30(10): 94-99 (in Chinese with English abstract)[本文引用:3][CJCR: 0.6505]
4
赵明辉, 马殿荣, 王嘉宇, 徐海, 唐亮, 陈温福. 低氮胁迫下水稻剑叶转录因子表达变化. 中国水稻科学, 2012, 26: 275-282Zhao MH, Ma DR, Wang JY, XuH, TangL, Chen WF. Expression of transcription factors of rice flag leaf under low nitrogen stress. Chin J Rice Sci, 2012, 26: 275-282 (in Chinese with English abstract)[本文引用:1][CJCR: 1.494]
5
马廷臣, 余蓉蓉, 陈荣军, 曾汉来, 张端品. 全基因组表达分析不同强度干旱胁迫下常规籼稻根系转录因子表达变化. 核农学报, 2013, 27: 1258-1269Ma TC, Yu RR, Chen RJ, Zeng HL, Zhang DP. Global genome expression change of transcription factors in conventional indica rice roots under different drought stress. Acta Agric Nucl Sin, 2013, 27: 1258-1269 (in Chinese with English abstract)[本文引用:3]
6
马廷臣, 陈荣军, 余蓉蓉, 曾汉来, 张端品. 全基因组分析PEG胁迫下水稻根系转录因子表达变化. 作物学报, 2009, 35: 1030-1037Ma TC, Chen RJ, Yu RR, Zeng HL, Zhang DP. Global genome expression analysis of transcription factors under PEG osmotic stress in rice root system. Acta Agron Sin, 2009, 35: 1030-1037 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
7
王曦, 汪小我, 王立坤, 冯智星, 张学工. 新一代高通量RNA测序数据的处理与分析. 生物化学与生物物理进展, 2010, 37: 834-846WangX, Wang XW, Wang LK, Feng ZX, Zhang XG. Processing and analysis of a new generation of high throughput RNA sequencing data. Prog Biochem Biophysics, 2010, 37: 834-846 (in Chinese with English abstract)[本文引用:1]
8
谢为博. 基于表达谱芯片和新一代测序技术的高通量基因分型方法的开发. 华中农业大学博士学位论文, 湖北武汉, 2010. pp 17-18Xie WB. Development of High Throughput Genotyping Methods Based on DNA Microarray and New Generation Sequencing Technologies. PhD Dissertation of Huazhong Agricultural University, Wuhan, China, 2010. pp 17-18 (in Chinese with English abstract)[本文引用:1]
9
魏利斌, 苗红梅, 张海洋. 芝麻发育转录组分析. 中国农业科学, 2012, 45: 1246-1256Wei LB, Miao HM, Zhang HY. Transcriptomic analysis of sesame development. Sci Agric Sin, 2012, 45: 1246-1256 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
10
宋雯雯, 李文滨, 韩雪, 高慕娟, 王继安. 干旱胁迫下大豆幼苗根系基因的表达谱分析. 中国农业科学, 2010, 43: 4579-4586Song WW, Li WB, HanX, Gao MJ, Wang JA. Analysis of gene expression profiles in soybean roots under drought stress. Sci Agric Sin, 2010, 43: 4579-4586 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
11
孙爱清, 张杰道, 万勇善, 刘风珍, 张昆, 孙利. 花生干旱胁迫响应基因的数字表达谱分析. 作物学报, 2013, 39: 1045-1053Sun AQ, Zhang JD, Wan YS, Liu FZ, ZhangK, SunL. In silico expression profile of genes in response to drought in peanut. Acta Agron Sin, 2013, 39: 1045-1053 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
12
WangG, Zhu QG, Meng QW, Wu CG. Transcript profiling during salt stress of young cotton (Gossypium hirsutum) seedlings via Solexa sequencing. Acta Physiol Plant, 2012, 34: 107-115[本文引用:1][JCR: 1.305]
13
WuJ, Zhang YL, Zhang HQ, HuangH, Folta KM, LuJ. Whole genome wide expression profiles of Vitis amurensis grape responding to downy mildew by using Solexa sequencing technology. BMC Plant Biol, 2010, 10: 234[本文引用:1][JCR: 4.354]
14
Yu SC, Zhang FL, Yu YJ, Zhang DS, Zhao XY, Wang WH. Transcriptome profiling of dehydration stress in the Chinese cabbage (Brassica rapa L. ssp. pekinensis) by tag sequencing. Plant Mol Biol Rep, 2012, 30: 17-28[本文引用:1][JCR: 5.319]
15
PangT, Ye CY, Xia XL, Yin, WL. De novo sequencing and transcriptome analysis of the desert shrub, Ammopiptanthus mongolicus, during cold acclimation using Illumina/Solexa. BMC Gene, 2013, 14: 488-503[本文引用:1]
16
Shan XH, Li YD, JiangY, Jiang ZL, Hao WY, Yuan YP. Transcriptome profile analysis of maize seedlings in response to high-salinity, drought and cold stresses by deep sequencing. Plant Mol Biol Rep, 2013, 31: 1485-1491[本文引用:1][JCR: 5.319]
17
Andrew JC, Liu DC, RamilM, YueI C H, RachidS. Transcriptome profiling of leaf elongation zone under drought in contrasting rice cultivars. PloS One, 2013, 8: e54537[本文引用:1][JCR: 3.73]
18
Chen JH, Song YP, ZhangH, Zhang DQ. Genome-wide analysis of gene expression in response to drought stress in Populus simonii. Plant Mol Biol Rep, 2013, 31: 946-962[本文引用:1][JCR: 5.319]
19
WangY, XuL, Chen YL, ShenH, Gong YQ, CeciliaL, LiuL W. Transcriptome profiling of radish (Raphanus sativus L. ) root and identification of genes involved in response to lead (Pb) stress with next generation sequencing. PloS One, 2013, 8: e66539[本文引用:1][JCR: 3.73]
20
彭姗, 吕学莲, 高峰, 李国英, 李晖. 一种新的棉花黄, 枯萎病快速接种方法的研究. 棉花学报, 2008, 20: 174-178PengS, Lü XL, GaoF, Li GY, LiH. Study on a new rapid inoculation method for Verticillium wilt and Fusarium wilt of cotton. Cotton Sci, 2008, 20: 174-178 (in Chinese with English abstract)[本文引用:1][CJCR: 1.462]
21
MortazaviA, Williams BA, McCueK, SchaefferL, WoldB. Mapping and quantifying mammalian transcriptomes by RNA-seq. Natl Meth, 2008, 5: 621-628[本文引用:1]
22
AudicS, Claverie JM. The significance of digital gene expression profiles. Genome Res, 1997, 7: 986-995[本文引用:1][JCR: 14.397]
罗红丽, 陈银华. 植物抗病反应相关转录因子的研究进展. 热带生物学报, 2011, 2: 83-88Luo HL, Chen YH. Advance on transcription factors involved in plant disease resistance response. Chin J Trop Crops, 2011, 2: 83-88 (in Chinese with English abstract)[本文引用:2][CJCR: 0.562]
25
ZhangY, ZhangG, XiaN, Wang XJ, Huang LL, Kang ZS. Cloning and characterization of a bZIP transcription factor gene in wheat and its expression in response to stripe rust pathogen infection and abiotic stresses. Physiol Mol Plant Pathol, 2008, 73: 88-94[本文引用:1]
26
Lee SC, Choi HW, Hwang IS, Choi DS, Hwang BK. Functional roles of the pepper pathogen-induced bZIP transcription factor, CAbZIP1, in enhanced resistance to pathogen infection and environmental stresses. Planta, 2006, 224: 1209-1225[本文引用:1][JCR: 3.347]
27
秦捷, 王武, 左开井, 唐克轩. AP2基因家族的起源和棉花AP2转录因子在抗病中的作用. 棉花学报, 2006, 17: 366-370QinJ, WangW, Zuo KJ, Tang KX. Origin of AP2 gene family and cotton AP2 transcription factors roles in plant resistance. Cotton Sci, 2006, 17: 366-370 (in Chinese with English abstract)[本文引用:1][CJCR: 1.462]
28
何兰兰, 柴蒙亮, 韩泽刚, 赵曾强, 张薇. 棉花抗枯萎病相关ERF-B3亚组转录因子的克隆与表达. 西北植物学报, 2013, 33: 2375-2381He LL, Chai ML, Han ZG, Zhao ZQ, ZhangW. Cloning and expression of ERF-B3 subgroup transcription factor related to resistance Fusarium wilt in cotton. Acta Bot Boreali-Occident Sin, 2013, 33: 2375-2381 (in Chinese with English abstract)[本文引用:1][CJCR: 1.321]
29
王磊, 高晓清, 朱苓华, 周永力, 黎志康. 植物WRKY转录因子家族基因抗病相关功能的研究进展. 植物遗传资源学报, 2011, 12: 80-85WangL, Gao XQ, Zhu LH, Zhou YL, Li ZK. Advances in research on function of WRKY transcription factor genes in plant resistance. J Plant Genet Res, 2011, 12: 80-85 (in Chinese with English abstract)[本文引用:1]
30
王瑞, 吴华玲, 王会芳, 黄珂, 霍春艳, 倪中福, 孙其信. 小麦TaWRKY44基因的克隆、表达分析及功能鉴定. 作物学报, 2013, 39: 1944-1951WangR, Wu HL, Wang HF, HuangK, Huo CY, Ni ZF, Sun QX. Cloning, characterization, and functional analysis of TaWRKY44 gene from wheat. Acta Agron Sin, 2013, 39: 1944-1951 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
31
ChenC, ChenZ. Potentiation of developmentally regulated plant defense response by AtWRKY18, a pathogen-induced Arabidopsis transcription factor. Plant Physiol, 2002, 129: 706-716[本文引用:1][JCR: 6.555]
32
Zhang ZL, XieZ, Zou XL, CasarettoJ, HoT H D, ShenQ X J. A rice WRKY gene encodes a transcriptional repressor of the gibberellin signaling pathway in aleurone cells. Plant Physiol, 2004, 134: 1500-1513[本文引用:1][JCR: 6.555]
33
Xu YH, Wang JW, WangS, Wang JY, Chen XY. Characterization of GaWRKY1, a cotton transcription factor that regulates the sesquiterpene synthase gene-delta-cadinene sythase-A. Plant Physiol, 2004, 135: 507-515[本文引用:]
34
LuoH, SongF, Goodman RM, ZhengZ. Up-regulation of OsBIHD1, a rice gene encoding BELL homeodomain transcriptional factor, in disease resistance responses. Plant Biol, 2005, 7: 459-468[本文引用:1][JCR: 2.32]
35
LuoH, SongF, ZhengZ. Overexpression in transgenic tobacco reveals different roles for the rice homeodomain gene OsBIHD1 in biotic and abiotic stress responses. J Exp Bot, 2005, 56: 2673-2682[本文引用:1][JCR: 5.242]
36
CoegoA, RamirezV, Gil MJ, FlorsbV, Manic BM, VeraaP. An Arabidopsis homeodomain transcription factor, OVEREXPRESSOR OF CATIONIC PEROXIDASE 3, mediates resistance to infection by necrotrophic pathogens. Plant Cell, 2005, 17: 2123-2137[本文引用:1][JCR: 9.251]
37
邢国芳, 张雁明, 张魏斌, 马新耀, 韩渊怀. 植物NAC转录因子的研究进展. 山西农业科学, 2012, 40: 409-411Xing GF, Zhang YM, Zhang WB, Ma XY, Han YH. Research progress of NAC transcription factor in plant. J Shanxi Agric Sci, 2012, 40: 409-411 (in Chinese with English abstract)[本文引用:1][CJCR: 0.8386]
38
李方正, 杨素欣, 吴春霞, 魏海超, 曲瑞莲, 冯献忠. 大豆KNOX基因家族的结构和表达分析. 植物学报, 2012, 47: 236-247Li FZ, Yang SX, Wu QX, Wei HC, Qu RL, Feng XZ. Structure and expression analysis of KNOX gene family in soybean. Chin Bull Bot, 2012, 47: 236-247 (in Chinese with English abstract)[本文引用:1]
39
刘晓月, 王文生, 傅彬英. 植物bHLH转录因子家族的功能研究进展. 生物技术进展, 2012, (1): 391-397Liu XY, Wang WS, Fu BY. Research progress of plant bHLH transcription factor family. Curr Biotechnol, 2012, (1): 391-397 (in Chinese with English abstract)[本文引用:1]
 , 赵曾强
, 赵曾强

{kind=link}