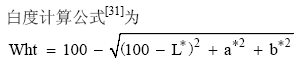摘要利用已开发的面粉色泽相关性状基因的分子标记, 检测了182份新疆小麦品种(系)的 Psy-A1、 Ppo-A1、 Ppo-D1、 TaLox-B1等位变异, 并结合表型鉴定结果, 分析不同等位变异对面粉色泽的影响。结果表明, 单基因等位变异对面粉色泽的影响不同, 其中4个基因等位变异L*值的差异均不显著; 对a*值的效应, Psy-A1a高于 Psy-A1b( P < 0.01), Ppo-A1b高于 Ppo-A1a( P < 0.05), TaLox-B1a高于 TaLox-B1b( P < 0.05); Psy-A1a基因型的b*值高于 Psy-A1b基因型( P < 0.01); Psy-A1a基因型的Wht值低于 Psy-A1b基因型( P < 0.01)。说明 Psy-A1是影响面粉色泽(红度、黄度和白度)的重要基因, 而 Ppo-A1和 TaLox-B1基因的等位变异只对面粉红度有显著影响。4个检测基因共有12种等位变异组合, 对a*、b*和Wht值有显著效应( P < 0.01), 但对L*值影响不显著( P > 0.05); Psy-A1b/Ppo-A1a/Ppo-D1b/ TaLox-B1b组合具有最高的Wht值和最低的a*、b*值, Psy-A1a/Ppo-A1a/Ppo-D1b/TaLox-B1a组合具有最低的Wht值和最高的a*、b*值。182份品种的Wht值平均为86.86, 其中审定品种平均为86.87, 冬小麦品种为86.42, 春小麦品种为87.32。青春5号具有优良的基因型和较高的面粉白度, 应加强利用。总体来看, 改良新疆小麦面粉的白度, 应重点加强对 Psy-A1基因的选择, 提高 Psy-A1b比例。
关键词:新疆; 面粉色泽基因; 等位基因; 功能标记 Effects of Psy-A1, Ppo-A1, Ppo-D1, and TaLox-B1 Alleles on Flour Color of Xinjiang Wheats XIANG Ji-Shan1,2, MU Pei-Yuan1,2,*, SANG Wei1,2, XU Hong-Jun1,2,*, NIE Ying-Bin1,2, CUI Feng-Juan1,2, ZHUANG Li2,3, HAN Xin-Nian1,2, ZOU Bo1 1Crop Research Institute, Xinjiang Academy of Agricultural and Reclamation Science, Shihezi 832000, China
2 Key Laboratory of Xinjiang Production and Construction Corps for Cereal Quality Research and Genetic Improvement, Shihezi 832000, China
3 College of Life Science, Shihezi University, Shihezi 832003, China
AbstractThe allelic variations on Psy-A1, Ppo-A1, Ppo-D1, and TaLox-B1 loci in 182 Xinjiang wheat varieties (lines) were tested using developed molecular markers that are associated with flour color. The effects of these alleles were analyzed in combination with phenotypes with the L*, a*, and b* values as well as the Wht data. On the view of a single gene, the allelic variation showed diverse effects on flour color. For L* value, the allelic effect was not significant. For a* value, the effect was Psy-A1a> Psy-A1b( P < 0.01), Ppo-A1b > Ppo-A1a ( P < 0.05), and TaLox-B1a > TaLox-B1b ( P < 0.05). For b* value, the effect of Psy-A1awas higher than that of Psy-A1b( P < 0.01). As a result, the Wht value of Psy-A1awas signlificantly lower than that of Psy-A1b( P < 0.01). These results suggest that Psy-A1 significantly affects flour redness, yellowness and whiteness, whereas Ppo-A1 and TaLox-B1 only have significant effects on flour redness. A total of 12 allelic combinations were found on the four gene loci, with significant effect on a*, b* and Wht values ( P < 0.01) and slight effect on L* value ( P > 0.05). The combination Psy-A1b/PPO- A1a/PPO-D1b/TaLox-B1b had the highest Wht value and the lowest a* and b* values, and combination Psy-A1a/PPO-A1a/PPO- D1b/TaLox-B1ahad the lowest Wht value and the highest a* and b* values. The average Wht value of total varieties (lines) tested was 86.86, and that of the released, winter, and spring varieties (lines) was 86.87, 86.42, and 87.32, respectively. Qingchun 5 should be used frequently as a promising parent in breeding programs for its excellent genotype and high flour whiteness. Our results indicate that Psy-A1is a key gene for improving flour whiteness, and the frequency of Psy-A1b should be promoted in Xinjiang wheats.
表2 4个基因不同等位变异对面粉色泽的影响 Table 2 Effect of alleles of four genes on flour color
基因 Gene
等位变异 Genetype
品种数 No. of varieties
频率 Frequency (%)
L* 值 L* value
a* 值 a* value
b* 值 b* value
白度 Wht value
Psy-A1
Psy-A1a
168
92.3
90.88
-1.02* *
9.50* *
86.76
Psy-A1b
14
7.7
91.20
-0.51
7.57
88.36* *
Ppo-A1
Ppo-A1a
128
70.3
90.90
-0.95
9.27
86.93
Ppo-A1b
54
29.7
90.96
-1.08*
9.58
86.76
Ppo-D1
Ppo-D1a
158
86.8
90.87
-1.00
9.47
86.79
Ppo-D1b
24
13.2
91.22
-0.94
8.86
87.41
TaLox-B1
TaLox-B1a
46
25.3
90.86
-1.11*
9.55
86.70
TaLox-B1b
136
74.7
90.92
-1.00
9.44
86.91
* and * * indicate significant difference between two alleles on one gene locus at P < 0.05 and P < 0.01 according to t-test, respectively. * 和* * 分别表示经t检验同一基因位点上2个等位变异之间差异显著(P < 0.05)和差异极显著(P < 0.01)。
表2 4个基因不同等位变异对面粉色泽的影响 Table 2 Effect of alleles of four genes on flour color
表3 4个基因不同等位变异组合及其色泽差异 Table 3 Flour colors of different allelic combination on Psy-A1, Ppo-A1, Ppo-D1, and TaLox-B1loci
等位变异组合 Allelic combination
品种数 No. of varieties
频率 Frequency (%)
L* 值 L* value
a* 值 a* value
b* 值 b* value
白度 Wht value
Psy-A1a/Ppo-A1a/Ppo-D1a/TaLox-B1a
25
13.7
90.73
-1.10 ABC
9.66 A
86.53 AB
Psy-A1a/Ppo-A1a/Ppo-D1a/TaLox-B1b
75
41.2
90.80
-0.93 ABC
9.34 AB
86.82 AB
Psy-A1a/Ppo-A1a/Ppo-D1b/TaLox-B1a
2
1.1
91.04
-1.38 A
10.36 A
86.23 B
Psy-A1a/Ppo-A1a/Ppo-D1b/TaLox-B1b
16
8.8
91.38
-1.05 ABC
9.16 AB
87.30 AB
Psy-A1a/Ppo-A1b/Ppo-D1a/TaLox-B1a
16
8.8
91.12
-1.21 AB
9.56 A
86.88 AB
Psy-A1a/Ppo-A1b/Ppo-D1a/TaLox-B1b
31
17.0
90.92
-1.11 ABC
9.76 A
86.60 AB
Psy-A1a/Ppo-A1b/Ppo-D1b/TaLox-B1b
3
1.7
90.39
-0.57 ABC
8.71 AB
87.02 AB
Psy-A1b/Ppo-A1a/Ppo-D1a/TaLox-B1a
3
1.7
90.39
-0.51 BC
8.02 AB
87.44 AB
Psy-A1b/Ppo-A1a/Ppo-D1a/TaLox-B1b
5
2.8
91.37
-0.53 BC
7.90 AB
88.28 AB
Psy-A1b/Ppo-A1a/Ppo-D1b/TaLox-B1b
2
1.1
91.63
-0.35 C
6.01 B
89.67 A
Psy-A1b/Ppo-A1b/Ppo-D1a/TaLox-B1b
3
1.7
91.63
-0.57 ABC
7.72 AB
88.58 AB
Only one variety is Psy-A1b/Ppo-A1b/Ppo-D1b/TaLox-B1bgenotype, which is not included in the table. Capital letters indicate significant difference between flour colors on different allelic combinations at P < 0.01 according to LSD multiple-comparison. 基因型Psy-A1b/Ppo-A1b/Ppo-D1b/TaLox-B1b只有1个品种, 未列入表; 大写字母表示经LSD法多重比较色泽在不同等位变异组合之间差异极显著(P < 0.01)。
表3 4个基因不同等位变异组合及其色泽差异 Table 3 Flour colors of different allelic combination on Psy-A1, Ppo-A1, Ppo-D1, and TaLox-B1loci
杨朝柱, 张磊, 司红起, 马传喜. 小麦面粉白度研究进展. 麦类作物学报, 2002, 3: 74-77Yang CZ, ZhangL, Si HQ, Ma CX. Review of wheat flour whiteness. J Triticeae Crops, 2002, 3: 74-77 (in Chinese with English abstract)[本文引用:3][CJCR: 1.007]
[2]
He ZH, YangJ, ZhangY, Quail KJ, Pena RJ. Pan bread and dry white Chinese noodle quality in Chinese winter wheats. Euphytica, 2004, 139: 257-267[本文引用:1][JCR: 1.643]
[3]
杨金, 张艳, 何中虎, 阎俊, 王德森, 刘建军, 王美芳. 小麦品质性状与面包和面条品质关系分析. 作物学报, 2004, 30: 739-744YangJ, ZhangY, He ZH, YanJ, Wang DS, Liu JJ, Wang MF. Association between wheat quality traits and performance of pan bread and dry white Chinese noodle. Acta Agron Sin, 2004, 30: 739-744 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[4]
Asenstorfer RE, WangY, Mares DJ. Chemical structure of flavonoid compounds in wheat (Triticum aestivum L. ) flour that contribute to the yellow colour of Asian alkaline noodles. J Cereal Sci, 2006, 43: 108-119[本文引用:1][JCR: 2.088]
[5]
Fu BX. Asian noodles: history, classification, raw materials, and processing. Food Res Int, 2008, 41: 888-902[本文引用:1][JCR: 3.005]
[6]
LintigJ, WelschR, BonkM, GiulianoG, BatschauerA, KleinigH. Light-dependent regulation of carotenoid biosynthesis occurs at the level of phytoene synthase expression and is mediated by phytochrome in Sinapis alba and Arabidopsis thaliana seedlings. Plant J, 1997, 12: 625-634[本文引用:2][JCR: 6.582]
[7]
Irvine GN, Anderson JA. Variation in principal quality factors of durum wheat with a quality prediction test for wheat or semolina. Cereal Chem, 1953, 30: 334-342[本文引用:1][JCR: 1.251]
[8]
ŽilićS, DodigD, ŠukalovićV H T, MaksimovićM, SaratlićG, ŠkrbićB. Bread and durum wheat compared for antioxidants contents, and lipoxygenase and peroxidase activities. Int J Food Sci Technol, 2010, 45: 1360-1367[本文引用:1]
[9]
MannaF, Borrelli GM, Massardo DR, WolfK, AlifanoP, Del GiudiceL, Di FonzoN. Expression of lipoxygenase genes among durum wheat cultivars. Cereal Res Commun, 1998, 26: 23-30[本文引用:1][JCR: 0.549]
[10]
Lindgren LO, Stalberg KG, Hoglund AS. Seed-specific over expression of an endogenous Arabidopsis phytoene synthase gene results in delayed germination and increased levels of carotenoids, chlorophyll, and abscisic acid. Plant Physiol, 2003, 132: 779-785[本文引用:1][JCR: 6.555]
[11]
朱长甫, 陈星, 王英典. 植物类胡萝卜素生物合成及其相关基因在基因工程中的应用. 植物生理与分子生物学学报, 2005, 30: 609-618Zhu CF, ChenX, Wang YD. Carotenoid biosynthesis in plants and application of its relative gene in gene engineering. J Plant Physiol Mol Biol, 2005, 30: 609-618 (in Chinese with English abstract)[本文引用:1][CJCR: 1.344]
[12]
He XY, Zhang YL, He ZH, Wu YP, Xiao YG, Ma CX, Xia XC. Characterization of a phytoene synthase 1 gene (Psy1) located on common wheat chromosome 7A and development of a functional marker. Theor Appl Genet, 2008, 116: 213-221[本文引用:2][JCR: 3.658]
[13]
He XY, He ZH, MaW, AppelsR, Xia XC. Allelic variants of phytoene synthase 1 (Psy1) genes in Chinese and CIMMYT wheat cultivars and development of functional markers for flour color. Mol Breed, 2009, 23: 553-563[本文引用:4][JCR: 3.251]
Kruger JE, Hatcher DW, DePauwR. A whole seed assay for polyphenol oxidase in Canadian prairie spring wheats and its usefulness as a measure of noodle darkening. Cereal Chem, 1994, 7: 324-326[本文引用:3][JCR: 1.251]
[16]
张立平, 葛秀秀, 何中虎, 王德森, 闫俊, 夏先春, Sutherland MW. 普通小麦多酚氧化酶活性的QTL分析. 作物学报, 2005, 31: 7-10Zhang LP, Ge XX, He ZH, Wang DS, YanJ, Xia XC, Sutherland MW. Mapping QTLs for polyphenol oxidase activity in a DH population from common wheat. Acta Agron Sin, 2005, 31: 7-10 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[17]
Sun DJ, He ZH, Xia XC, Zhang LP, Morris CF, AppelsR, Ma WJ, WangH. A novel STS marker for polyphenol oxidase activity in bread wheat. Mol Breed, 2005, 16: 209-218[本文引用:3][JCR: 3.251]
[18]
He XY, He ZH, Zhang LP, Sun DJ, Morris CF, Fuerst EP, Xia XC. Allelic variation of polyphenol oxidase (PPO) genes located on chromosomes 2A and 2D and development of functional markers for the PPO genes in common wheat. Theor Appl Genet, 2007, 115: 47-58[本文引用:3][JCR: 3.658]
[19]
Borrelli GM, TroccoliA, Di FonzoN, FaresC. Durum wheat lipoxygenase activity and other quality parameters that affect pasta color. Cereal Chem, 1999, 76: 335-340[本文引用:2][JCR: 1.251]
[20]
ArdaS, VuralG. Effects of β-carotene on soybean lipoxygenase activity: kinetic studies. Eur Food Res Technol, 2007, 224: 743-748[本文引用:2][JCR: 1.436]
[21]
CarreraA, EeheniqueV, ZhangW, HelgueraM, MantheyF, SchragerA, PiccaA, CervigniG, DubcovskyJ. A deletion at the Lpx-B1 locus is associated with low lipoxygenase activity and improved pasta color in durum wheat. J Cereal Sci, 2007, 45: 67-77[本文引用:1][JCR: 2.088]
[22]
Parker GD, Chalmers KJ, Rathjen AJ, LangridgeP. Mapping loci associated with flour color in wheat (Triticum aestivum L. ). Theor Appl Genet, 1998, 97: 238-245[本文引用:1][JCR: 3.658]
[23]
Hessler TG, Thomson MJ, BenseherD, Nachit MM, Sorrells ME. Association of a lipoxygenase locus, Lpx-Bl, with variation in lipoxygenase activity in durum wheat seeds. Crop Sci, 2002, 42: 1695-1700[本文引用:1][JCR: 1.513]
[24]
HidalgoA, Brand oliniA, PompeiC. Carotenoids evolution during pasta, bread and water biscuit preparation from wheat flours. Food Chem, 2010, 121: 746-751[本文引用:2][JCR: 3.334]
[25]
Geng HW, Xia XC, Zhang LP, Qu YY, He ZH. Development of functional markers for a lipoxygenase gene TaLox-B1 on chromosome 4BS in common wheat. Crop Sci, 2012, 52: 568-576[本文引用:5][JCR: 1.513]
[26]
芦静, 张新忠, 吴新元, 黄天荣. 小麦品质性状与面制食品加工特性相关性研究. 新疆农业科学, 2002, 39: 290-292LuJ, Zhang XZ, Wu XY, Huang TR. Investigation on correlation between quality characters of wheat and processing quality of food. Xinjiang Agric Sci, 2002, 39: 290-292 (in Chinese with English abstract)[本文引用:1][CJCR: 0.778]
[27]
穆培源, 桑伟, 王亮, 庄丽, 王祥军, 芦静, 徐红军, 韩新年, 于芳祥. 新疆市售小麦面粉制作新疆拉面的加工品质特性及其专用粉品质评价指标的研究. 麦类作物学报, 2008, 27: 1034-1041Mu PY, SangW, WangL, ZhuangL, Wang XJ, LuJ, Xu HJ, Han XN, Yu FX. Study on processing quality of Xinjiang hand -stretched noodle and quality evaluation parameters of specific flour for the commercial wheat flours in Xinjiang. J Triticeae Crops, 2008, 27: 1034-1041 (in Chinese with English abstract)[本文引用:1][CJCR: 1.007]
[28]
王亮, 穆培源, 徐红军, 庄丽, 桑伟, 聂迎彬, 韩新年, 邹波. 新疆小麦品种黄色素含量基因(Psy-A1)等位变异的分子检测. 麦类作物学报, 2009, 29: 782-786WangL, Mu PY, Xu HJ, ZhaungL, SangW, Nie YB, Han XN, ZouB. Allelic variations of Psy-A1 gene for yellow pigment content in Xinjiang wheat cultivars. J Triticeae Crops, 2009, 29: 782-786 (in Chinese with English abstract)[本文引用:2][CJCR: 1.007]
[29]
王亮, 穆培源, 徐红军, 庄丽, 桑伟, 韩新年, 聂迎彬, 夏先春, 何中虎. 新疆小麦品种多酚氧化酶(PPO)活性基因等位变异检测. 麦类作物学报, 2008, 28: 766-771WangL, Mu PY, Xu HJ, ZhaungL, SangW, Han XN, Nie YB, Xia XC, He ZH. Distribution of allelic variations of polyphenol oxidase genes in Xinjiang wheat cultivars. J Triticeae Crops, 2008, 28: 766-771 (in Chinese with English abstract)[本文引用:2][CJCR: 1.007]
[30]
相吉山, 穆培源, 桑伟, 聂迎彬, 徐红军, 崔凤娟, 韩新年, 邹波. 新疆小麦品种资源脂肪氧化酶活性基因TaLox-B1的分布特征. 麦类作物学报, 2013, 33: 279-285Xiang JS, Mu PY, SangW, Nie YB, Xu HJ, Cui FJ, Han XN, ZouB. Distribution characteristics of lipoxygenase gene TaLox-B1 in wheat varieties from Xinjiang. J Triticeae Crops, 2013, 33: 279-285 (in Chinese with English abstract)[本文引用:2][CJCR: 1.007]
[31]
汪帆, 郑文寅, 黄建华, 王冠球, 崔文礼, 张文明, 姚大年. 20个小麦品种(系)籽粒LOX活性和类胡萝卜素含量及全麦粉色泽的研究. 麦类作物学报, 2012, 32: 68-73WangF, Zheng WY, Huang JH, Wang GQ, Cui WL, Zhang WM, Yao DN. Lipoxygenase acticity, carotenoids content and wholemill color in grains of twenty wheat varieties. J Triticeae Crops, 2012, 32: 68-73 (in Chinese with English abstract)[本文引用:1][CJCR: 1.007]
[32]
Lagudah ES, ApplesR, McNeilD. The Nor-D3 locus of Triticum tauschii: naturiation variation and genetic linkage to markers in chromosome 5. Genome, 1991, 34: 387-395[本文引用:4][JCR: 1.668]
[33]
Symons SJ. Computer analysis of fluoreseence for the measurement of flour refinement as determined by flour ash content, flour grade colour, and tristimulus colour measurements. Cereal Chem, 1991, 68: 454-460[本文引用:1][JCR: 1.251]
[34]
Oh NH, Seib PA, Chung DS. Noodles: III. Effects of processing variables on quality characteristics of dry noodles. Cereal Chem, 1985, 62: 437-440[本文引用:1][JCR: 1.251]
[35]
Moss HJ, Miskelly DM, MossR. The effect of alkaline conditions on the properties of wheat flour dough and Cantonese-style noodles. J Cereal Sci, 1986, 4: 261-268[本文引用:1][JCR: 2.088]
Kumagai MH, DonsonJ, Della-CioppaG, HarveyD, HanleyK, Grill LK. Cytoplasmic inhibition of carotenoid biosynthesis with virus-derived RNA. Proc Natl Acad Sci USA, 1995, 92: 1679-1683[本文引用:1][JCR: 9.737]
[38]
孙道杰, 何中虎, 王辉. 小麦面粉黄色素相关基因研究. 西北植物学报, 2006, 26: 655-660Sun DJ, He ZH, WangH. Genes related with yellow pigments in wheat flour. Acta Bot Boreali-Occident Sin, 2006, 26: 655-660 (in Chinese with English abstract)[本文引用:1][CJCR: 1.321]
[39]
LeenhardtF, LyanB, RockE, BoussardA, PotusJ, ChanliaudE, RemesyC. Genetic variability of carotenoid concentration, and lipoxygenase and peroxidase activities among cultivated wheat species and bread wheat varieties. Eur J Agron, 2006, 25: 170-176[本文引用:1][JCR: 2.8]
[40]
Irvine GN, Winkler CA. Factors affecting the color of macaroni: II. Kinetic studies of pigment destruction during mixing. Cereal Chem, 1950, 27: 205-218[本文引用:3][JCR: 1.251]
[41]
桑伟, 穆培源, 徐红军, 庄丽, 聂迎彬, 崔凤娟, 邹波. 新疆春小麦品种磨粉品质与新疆拉面加工品质的关系. 西北农业学报, 2012, 21: 54-59SangW, Mu PY, Xu HJ, ZhuangL, Nie YB, Cui FJ, ZouB. Correlations between milling qualities of spring wheats and hand -stretched noodle in Xinjiang. Acta Agric Boreali-Occident Sin, 2012, 21: 54-59 (in Chinese with English abstract)[本文引用:2]
[42]
桑伟, 穆培源, 徐红军, 庄丽, 王亮, 邹波. 新疆小麦品种籽粒性状、磨粉品质及其关系的研究. 麦类作物学报, 2010, 30: 50-55SangW, Mu PY, Xu HJ, ZhuangL, WangL, ZouB. Study on the grain traits, milling quality and their relationships in Xinjiang winter and spring wheat cultivars. J Triticeae Crops, 2010, 30: 50-55 (in Chinese with English abstract)[本文引用:1][CJCR: 1.007]
 , 穆培源
, 穆培源