 , 朱文博, 张静静, 朱连奇, 赵芳, 崔耀平
, 朱文博, 张静静, 朱连奇, 赵芳, 崔耀平河南大学环境与规划学院,开封 475004
Phenology of forest vegetation and its response to climate change in the Funiu Mountains
ZHANGXiaodong, ZHUWenbo, ZHANGJingjing, ZHULianqi, ZHAOFang, CUIYaoping通讯作者:
收稿日期:2017-01-9
修回日期:2017-07-19
网络出版日期:--
版权声明:2018《地理学报》编辑部本文是开放获取期刊文献,在以下情况下可以自由使用:学术研究、学术交流、科研教学等,但不允许用于商业目的.
基金资助:
作者简介:
-->
展开
摘要
关键词:
Abstract
Keywords:
-->0
PDF (1199KB)元数据多维度评价相关文章收藏文章
本文引用格式导出EndNoteRisBibtex收藏本文-->
1 引言
气候变化通过改变植物光合作用、生长期、土壤形成过程等对陆地生态系统产生重要影响,已受到科学界和国际组织的广泛关注。气候变化具有显著的区域差异性,生态脆弱带尤其是中高纬地区的地理过渡带、山地等地区是全球变暖信号的放大器[1,2,3,4,5]。其中,森林植被作为山地生态系统的重要组成部分,对气候变化较为敏感,山地森林植被物候能揭示山地植被的生长动态,直观反映山地生态系统对全球环境变化的响应和适应过程,已成为全球变化背景下物候学和植被地理学研究的热点[6,7,8]。分析森林植被物候的变化规律,研究森林植被物候对气候变化的响应,揭示山地地理环境变化的规律和区域差异,有助于深入了解植被—气候关系。基于遥感反演的NDVI、EVI等植被指数,国内外****探讨了不同地区植被物候的变化及其对气候要素的响应形式。Julien等利用GIMMS NDVI数据系统研究了全球1981-2003年的陆表植被物候变化,结果显示在全球尺度上春季物候期平均提前0.38 d/年,秋季物候平均延迟0.45 d/年,生长季长度延长0.8 d/年[9]。Liu等发现过去30年中国北方温带地区秋季植被生长季结束期平均每年推迟0.12±0.01 d,其中,季前气温(2个月)和秋季物候期在大部分地区呈现显著正相关关系。同时,降水和光照对生长季结束期变化也有较大的贡献,但是其影响随植被区域的不同而有所差异[10]。在一些特殊区域,例如高原苔原、热带雨林、干旱荒漠等,植被物候往往对气候变化响应更为敏感。因此,在这些地区开展植被物候研究具有深刻的意义。马晓芳等利用GIMMS NDVI遥感数据结合动态阈值法研究了青藏高原1982-2005年的物候期变化规律,表明青藏高原不同草地类型植被物候变化受温度的影响高于降水[11]。俎佳星等利用GIMMS AVHRR遥感数据分析我国东北地区物候对气候的响应,认为温度对春季植被生长的影响较大,而降水主要影响植被的秋季生长[12]。随着近50年来全球气候变化加剧,大部分地区都呈现出显著增温的趋势。但仍有少数地区受地形、植被、土壤等条件的限制,其气候变化趋势与全球整体趋势有所差别[13]。造成了植被物候变化与其他地区呈现较大的差异[14]。目前,国内对这些气候变化“异常”地区的植被物候研究较为薄弱。
伏牛山位于北亚热带—南暖温带的过渡区,自然地理梯度较大、植被演化过程多样,生态系统对气候变化的响应极为敏感[15,16,17,18]。同时该地区近40年的春季气温变化与其他地区不同,平均气温整体呈现降低的趋势[19],这种区域气候特征可能导致植被生长发育状况也与其他地区有较大区别。根据以往研究规律,温度降低对于春季植被生长发育有一定的抑制作用,并且会造成秋季生长季结束期的提前。那么,伏牛山地的春、秋季植被物候是否与其他地区呈现不一致的变化?这种物候的提前或推迟与温度、降水的关系是否与其他地区相同?为了揭示这些问题,本文利用Savitzky-Golay(S-G)滤波算法和阈值法,结合MODIS EVI和气温、降水数据,提取2000-2015年森林植被物候生长季始期、末期和生长季长度参数,分析伏牛山地森林植被物候的时空变化特征,深入探讨其与气温、降水的关系,识别气候要素影响物候的主要时段和范围,研究山地森林植被物候变化的趋势和规律,为全球变化背景下气候过渡带山地森林生态系统的恢复和保护提供参考。
2 数据与方法
2.1 研究区概况
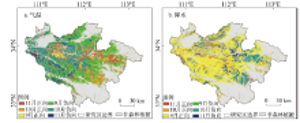
伏牛山位于河南省西部,属秦岭东部余脉,位于110°30′~113°30′E,32°45′~34°20′N之间,包括栾川、嵩县、鲁山、西峡、内乡、镇平、卢氏和南召8个县(图1),海拔在44~2173 m。年平均气温13.6~15.1 ℃,年降水量为700~1100 mm,降水主要集中在5-9月,雨热同期有利于植被的生长发育。伏牛山地也是我国地形第二阶梯向第三阶梯的过渡地区,自然地理景观差异较大,复杂的气候和地貌条件形成了从南暖温带落叶阔叶林向北亚热带常绿落叶混交林过渡的多样化植被类型。土壤以棕壤、黄棕壤和褐土为主,土层较薄,地形起伏度较大,立地条件较差,生态环境脆弱[20]。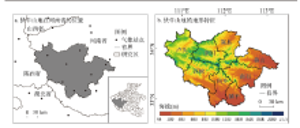 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1伏牛山地在河南省的位置及其地形特征
-->Fig. 1The location of Funiu Mountains in Henan Province and its terrain
-->
2.2 数据准备和处理
2.2.1 EVI数据 MODIS EVI数据来源于美国国家航空航天局(NASA)提供的MOD13Q1数据集,时间范围为2000-2015年,空间分辨率250 m,时间分辨率为16 d。首先用MRT工具从MOD13Q1原始数据集中提取出EVI数据,并进行重投影。最后利用研究区边界对数据进行裁剪,获得2000-2015年长时间序列的EVI影像。2.2.2 气象数据 气象数据为研究区及其周边的19个气象站点2000-2015年的月平均气温和月降水数据,其中14个站点的气象数据来源于中国气象数据网(http://data.cma.cn),为了更精确分析伏牛山地的水热变化情况,又从河南省气象局获取了5个站点的气象数据。考虑到森林植被的生长周期特点,气象数据插值的时间范围为2-11月。
2.2.3 其他数据 本文中的地形特征属性采用了空间分辨率为30 m的DEM数据,该数据为V2版本的ASTER GDEM,数据来源于中国科学院计算机网络信息中心地理空间数据云平台(http://www.gscloud.cn)。首先用ENVI 5.1将DEM数据进行镶嵌拼接,然后将处理好的DEM数据进行重投影并重采样至250 m,最后将研究区边界与DEM数据在ArcGIS软件里进行裁剪,得到伏牛山地的地形数据。
森林植被提取选取无云且清晰的国产环境一号30 m多光谱数据,日期为2015年10月,进行大气校正、几何校正、镶嵌以及研究区影像裁剪等预处理。
2.3 研究方法
2.3.1 森林植被物候的遥感提取方法 本文利用TIMESAT软件中的动态阈值法对伏牛山地森林植被物候期进行提取[21,22]。首先,选用软件包中的S-G滤波法对2000-2015年EVI影像进行平滑处理。然后将EVI增加或降低到当年EVI振幅一定比例的时间定义为生长季始期或生长季末期,本文将这一比例设置为50%[23]。根据这一方法逐像元提取研究区2000-2015年的森林植被物候参数,即生长季始期(Start of the growing season, SOS)和生长季末期(End of the growing season, EOS),EOS和SOS的差即为生长季长度(Growing season length, GSL)。采用儒略日对森林植被物候期进行换算,即将物候期表示为距1月1日的实际日数。2.3.2 森林植被识别方法 环境一号卫星在提取植被覆盖信息的方面得到广泛应用。通过对HJ-CCD遥感数据中反演的NDVI进行差值速率构建森林植被识别模型,并结合野外数据进行分析评价,结果表明HJ-CCD可以应用在植被的识别方面[24]。
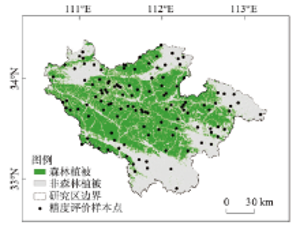
采用面向对象分类方法进行森林植被提取,可以避免由遥感影像中“同物异谱”、“异物同谱”和“椒盐现象”等影响因素对分类的影响,研究区10月份的农田植被覆盖度较低及草丛枯萎,可以较容易与森林植被区分开来(图2)。选120个样本点来验证分类精度,并依据伏牛山地植被相关数据进行精度评价,结果表明分类精度高于85%,山区森林植被更新演替的时间周期较长且空间变化范围有限,因此可用该数据进行研究。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2伏牛山地森林植被空间分布图
-->Fig. 2Spatial distribution of forest vegetation in Funiu Mountains
-->
2.3.3 气象插值方法 澳大利亚****Hutchinson基于样条函数理论开发了ANUSPLIN软件[25],该软件可引入多个影响因子进行气象要素空间插值,尤其适合时间序列的气象数据处理[26]。在复杂的山地环境,通过ANUSPLIN方法对气温、降水的插值精度误差较小[27]。谭剑波等利用ANUSPLIN在地表复杂的青藏高原对气温进行插值,结果显示,均方差仅为0.82 ℃[28]。因此,本文选用ANUSPLIN对2000-2015年的2-11月的月平均气温和月降水进行插值,样条次数为2,协变量为经度、纬度和高程,插值数据空间分辨率为250 m。
2.3.4 趋势和相关性分析 Theil-Sen斜率(T-sen)是一种非参数估计方法,可以用来计算长时间序列数据的变化情况[29],Mann-Kandall检验是一种非参数统计方法,具有受异常值干扰较少且无需对数据序列进行分布检验等特点,被广泛应用于长时间序列数据的趋势检验[30]。T-sen斜率和M-K检验在IDRISI软件里进行运算。本文采用Theil-Sen斜率方法,对伏牛山地森林植被生长季始期、末期和生长季长度的变化进行判断(T-sen)。若T-sen > 0,则表明物候期趋势是推迟或延长;T-sen < 0,则表明物候期趋势是提前或缩短。同时,进行Mann-Kendall统计检验(置信度为95%),结合T-sen和M-K检验结果,将伏牛山地森林植被物候期年际变化趋势等级分为:显著推迟或延长(T-sen > 0,p < 0.05);显著提前或缩短(T-sen <0,p < 0.05);不显著推迟或延长(T-sen > 0,p > 0.05);不显著提前或缩短(T-sen < 0,p > 0.05)。
利用R语言程序对伏牛山地森林植被物候期与2-11月平均气温、月降水进行Pearson相关和偏相关分析及显著性检验,得到伏牛山地森林植被物候动态对气温、降水的响应特征。其中,生长季始期分别与2月、3月和4月气温和降水进行相关性计算,生长季末期分别与9月、10月和11月气温和降水进行相关性计算,生长季长度分别与5月、6月、7月、8月和9月气温和降水进行相关性计算。同时,为深入分析伏牛山地森林植被物候期主要受哪个月份的气温和降水影响,运用ENVI/IDL程序分别对不同月份气温、降水和森林植被物候期的相关系数影像进行多波段合成,识别相关系数绝对值最大的月份,并标记出像元点是正向影响还是负向影响。
3 结果分析
3.1 森林植被多年平均物候期
为研究伏牛山地森林植被物候期的空间格局,计算了2000-2015年森林植被SOS、EOS和GSL的多年平均值(图3)。结果表明,森林植被生长季始期呈现出从边缘低海拔地区向中心高海拔区域推迟的规律,边缘地区生长季始期较早,时间为105~120 d(4月下旬),中心区域生长季始期较晚,多数在120~135 d(5月上旬)(图3a)。其中,在东部和南部的部分地区森林植被生长季始期最早,约为90~105 d。从图3b中可以看出南部和东部低海拔区域森林植被生长季末期最晚,主要集中在第300~315 d,其他区域森林植被生长季末期集中在第285~300 d。就森林植被生长季长度而言,其呈现出从边缘至中心区域逐渐缩短的规律(图3c)。东部和南部低海拔区域植被生长季长度多数为195~210 d,西北部地区的大部分植被生长季长度为165~180 d,中心区域的植被生长季长度多数仅为150~165 d。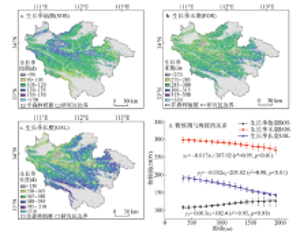 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图32000-2015年伏牛山地森林植被物候期多年均值空间分布及其与海拔的关系
-->Fig. 3Spatial distribution of multi-annual mean forest phenology in Funiu Mountains and its relationship with altitude from 2000 to 2015
-->
森林植被物候期平均值和标准差随海拔的变化特征如图3d所示。随海拔上升,森林植被生长季始期呈显著推迟趋势,推迟幅度为1.3 d/100m(r2 = 0.95,p < 0.01),生长季末期显著提前,提高幅度为1.7 d/100m(r2 = 0.95,p < 0.01),生长季长度以3.2 d/100m的趋势显著缩短(r2 = 0.98,p < 0.01)。
3.2 森林植被物候期年际变化
森林植被生长季始期在大部分像元点(76.57%)呈现推迟趋势,但是推迟的程度较小并且显著推迟的区域仅占2.16%,主要在中部地区有零星分布(图4a)。对于其余像元点,不显著提前区域主要集中在南部、北部和东部部分地区,显著提前区域分布于东部少数地区,且分布较为分散。森林植被生长季末期也主要呈现推迟的趋势(83.81%),其中,显著推迟的像元点占6.38%,主要分布于东南部低海拔地区。呈不显著推迟趋势的区域主要集中在中部和偏北部。提前的像元集中于南部和东部,显著提前的区域仅有0.04%(图4b)。森林植被生长季长度的变化不显著,60.85%和36.25%的像元点分别呈现不显著延长和缩短的趋势(图4c)。不显著缩短区域集中在中部、南部和偏东部分地区,不显著延长区域分散分布于中部地区。对于变化显著的像元点而言,显著延长区域(0.36%)主要集中在偏东南部地区以及偏北部少部分地区。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图42000-2015年伏牛山地森林植被物候期年际变化空间分布
-->Fig. 4Spatial distribution of interannual variation of forest phenology in Funiu Mountains from 2000 to 2015
-->
3.3 森林植被物候动态对气温、降水的响应
3.3.1 生长季始期对气温、降水的响应 从森林植被生长季始期与2-4月平均气温和月降水量偏相关空间分布可以看出(图5),在气温方面,森林植被生长季始期与2月和3月平均气温的偏相关系数主要为负值,表明气温降低,森林植被生长季始期推迟,而与4月份平均气温的偏相关系数多为正值。在降水方面,2月降水量在较高海拔区域对森林植被生长季始期的影响多为负向,在较低海拔区域对森林植被生长季始期的影响多为正向。3月降水量与森林植被生长季始期的偏相关系数正负值在空间上的分布较为均匀。4月降水量与森林植被生长季始期的偏相关系数多为正值。从显著性水平来看,森林植被生长季始期与3月平均气温的偏相关系数显著的像元点最多(13.94%),这表明森林植被生长季始期在更多的地区受3月平均气温的影响。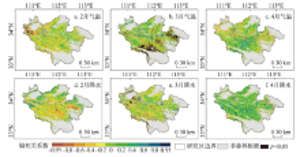 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5伏牛山地森林植被生长季始期与2-4月气温降水的偏相关系数空间分布
-->Fig. 5Spatial distribution of partial correlation coefficients between SOS and February-April temperature/precipitation in Funiu Mountains
-->
图6描述了森林植被生长季始期与对其影响最大的月份的气温和降水的相关关系。就气温对生长季始期的影响来看,在37.24%和37.25%的像元点,生长季始期主要受2月和3月气温的影响,并且月平均气温降低,导致生长季始期推迟。主要受2月气温负向影响的点集中分布于西北部,而受3月气温负向影响的点主要分布在东部和南部。4月气温对生长季始期起影响作用的像元点占22.90%,其中,正向和负向影响的像元点分别占2.93%和19.97%。就降水而言,4月降水对生长季始期的影响最为明显。在40.33%的像元点,4月降水增加引起生长季始期推迟。在另外4.60%的像元点,4月降水增加使生长季始期提前。此外,2月和3月降水对生长季始期影响最大的像元分别占28.96%和26.11%,并且影响作用以负向为主。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6伏牛山地森林植被生长季始期对气温和降水响应的空间分布
-->Fig. 6Spatial distribution of response of SOS to temperature and precipitation in Funiu Mountains
-->
3.3.2 生长季末期对气温、降水的响应 从森林植被生长季末期与9-11月平均气温和月降水量偏相关空间分布可以看出(图7),在气温方面,森林植被生长季末期与9月平均气温偏相关系数多为正值,与10月和11月平均气温偏相关系数多为负值。在降水方面,伏牛山地多数区域森林植被生长季末期与9月降水量呈正相关关系,而与10月和11月呈正相关关系的区域面积较少。从显著性水平上看,森林植被生长季末期与9-11月份气温的偏相关关系显著的区域均少于5%(分别为4.09%、3.03%和2.17%)。森林植被生长季末期与9月降水的偏相关显著的像元占10.47%,主要分布于西南部,与10月和11月降水偏相关显著的像元数量明显较少,分别为2.88%和1.30%。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图7伏牛山地森林植被生长季末期与9-11月气温降水的偏相关系数空间分布
-->Fig. 7Spatial distribution of partial correlation coefficient between EOS and September-November temperature/precipitation in Funiu Mountains
-->
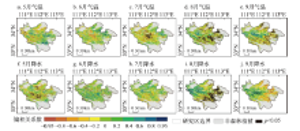
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图8伏牛山地森林植被生长季末期对气温和降水响应的空间分布
-->Fig. 8Spatial distribution of response of EOS to temperature and precipitation in Funiu Mountains
-->
从森林植被生长季末期与对其影响最大的月份的气温和降水的相关关系可以看出(图8),在气温方面,生长季末期主要受9月和10月平均气温负向影响的像元点最多(42.87%和23.27%)。在降水方面,9月降水对植被生长季末期起主要影响作用的点最多,影响为正向的点占64.99%,表明对于大部分地区而言,9月降水增多会使植被生长季末期推迟。10月降水起主要影响作用的地区位于东部低海拔区域,且在这些地区,10月降水与生长季末期多为负相关关系(16.94%)。
3.3.3 生长季长度对气温、降水的响应 在气温方面,5-7月平均气温和生长季长度在较高海拔的中心区域多为负相关关系,在边缘地区多为正相关关系(图9);8月份和9月平均气温对东部地区的生长季长度具有负面影响,对西部地区大部分为正面影响。在降水方面,5-7月份的降水增多使得大部分地区的生长季长度延长。而8月份和9月份的月降水对生长季长度的影响具有明显的空间异质性。其中,降水与北部、南部和东部的生长季长度主要呈负相关关系,和西北部多数地区的生长季长度呈正相关关系。从显著性水平来看,8月、9月平均气温和生长季长度关系显著的像元点较多(16.07%和11.61%),主要分布于北部、东部和南部区域,同时,8月份降水与生长季长度在18.14%的区域呈现显著关系,这些像元点基本都分布于东南部低海拔地区。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图9伏牛山地植被生长季长度与5-9月气温降水的偏相关系数空间分布
-->Fig. 9Spatial distribution of partial correlation coefficient between GSL and May-September temperature/precipitation in Funiu Mountains
-->
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图10伏牛山地森林植被生长季长度对气温和降水响应的空间分布
-->Fig. 10Spatial distribution of response of GSL to temperature and precipitation in Funiu Mountains
-->
森林植被生长季长度与对其影响最大的月份的气温和降水的相关关系较为复杂(图10)。在气温方面,9月份平均气温对生长季长度影响较大的像元点最多(40.46%),且大部分呈现负相关关系。6月和7月平均气温对生长季长度影响较大的像元点数量相近,分别为11.61%和18.68%。从分布上来看,低海拔的边缘地区受气温的正向影响大,说明气温的升高使低海拔区的森林植被生长季长度延长。在降水方面,西北部主要受8月和9月降水的正向影响,而东部主要受5月和6月降水的负向影响,表明降水对森林植被生长季长度的影响同时存在时间和空间的差异。
4 讨论
研究山地森林植被物候变化可以了解森林植被生长动态,识别山地森林植被对气候变化的响应。本文结果表明,伏牛山地过去16年春季物候整体呈现推迟的趋势,这一研究结果与夏浩铭等在秦岭东部低海拔地区的结果相一致[31]。物候期的推迟可能影响植被的生长发育[32],导致初级生产力减少[33],森林生态系统服务退化,同时也会产生正面的作用,例如降低了植被遭受早春霜冻的风险[34]。但和中国温带大部分地区植被春季物候期提前的趋势相比,伏牛山地森林植被春季物候变化具有明显的区域特殊性,这种特殊性可以用区域的气候特征来解释。根据本文的研究结果,森林植被生长季始期、末期和生长季长度受到了多个月份温度和降水变化的共同影响。植被物候的变化一方面取决于植被物候和与之相关月份的温度、降水的相关关系,另一方面取决于温度和降水变化。从相关关系的角度来看,伏牛山地春季物候期,即植被生长季始期受2月和3月份平均气温的影响最大,主要呈现负相关关系,即春季温度降低导致春季物候期的推迟。相比之下,降水对生长季始期的作用具有较大的空间差异。春季降水与生长季始期呈现负相关关系的像元占54.33%,其余像元呈现正相关关系。降水的增加在某些地区能够使植被获取充足的水分,提高其生长速率,物候期提前。但在一些针叶林地区,降水增加会导致气温降低,影响春季温度积累,反而推迟物候期[12]。总体而言,伏牛山地区春季物候期与温度、降水的关系与全国其他地区基本一致。但从伏牛山地的月平均气温的变化情况来看,2月和3月的气温均呈明显的降低趋势,降低幅度分别达到了-0.68 ℃/10a和-0.25 ℃/10a。这与全国大部分地区的春季升温情况不同[13]。由此可见,春季温度的“异常”变化是导致伏牛山地春季物候期推迟的主要因素。
相比于春季物候,温度和降水对生长季末期的影响在不同月份具有明显差异。其中,9月份的温度和降水都与生长季末期正相关,表明温度升高或者降水增加导致结束期推迟。这也与中国北方地区的研究结果相吻合[35,36]。但是本文也发现10月和11月的降水增多会导致部分区域生长季末期提前,可能原因是降水增多加大了土壤湿度,进而影响植被羟化作用和光合作用速率,使植被加速完成生长,最终导致生长季末期提前[37]。同时,春、秋季的物候变化除了受温度、降水的影响外,也可能受到其他环境因素的影响,例如光周期、辐射强度、碳浓度等[38,39,40,41]。虽然这些环境因素对植被物候的影响与温度降水相比较小,但仍不容忽视。
本文提取的植被物候期还与地面观测的物候资料进行了对比验证。本文收集了该地区2个观测站点内乡榆树和卢氏小叶杨2000-2015年的展叶和落叶数据。其中榆树多年平均展叶始期为3月下旬,落叶始期为9月上旬;小叶杨多年平均展叶始期为4月上旬,落叶始期为9月上旬,叶变色全变为9月中旬。这与本文得出的森林植被生长季始期集中在第90~120 d,生长季末期集中在第270~300 d基本一致。虽然遥感物候反映的是大尺度的植被物候变化,与野外实地观测的植物物候有所差异,但两者在物候的发生时间也能很好的对应。遥感物候数据和地面物候观测数据的尺度转换,是下一步研究的重点。
5 结论
本文基于MODIS EVI和气温、降水数据,利用S-G滤波算法和阈值法提取了2000-2015年森林植被物候生长季始期、末期和生长季长度参数,并系统分析了伏牛山地森林植被物候的时空变化特征,深入探讨了其与气温、降水的关系。本文的主要结论如下:(1)伏牛山地森林植被生长季始期主要集中在第105~120 d,森林植被的生长季始期呈现出从边缘低海拔地区向中心高海拔区域推迟的规律。生长季末期主要集中在第285~315 d。生长季长度主要集中在165~195 d,呈现出从边缘至中心区域逐渐缩短的规律。随着海拔的升高,森林植被生长季始期、末期和长度整体上分别呈显著推迟、提前及缩短趋势。
(2)从森林植被物候近16年的年际变化来看,森林植被生长季始期和生长季末期呈推迟的面积分别占森林植被总面积的76.57%和83.81%,森林植被生长季长度呈延长的面积比为61.21%,但是显著的比例仅有0.36%。
(3)伏牛山地森林植被生长季始期与2-4月平均气温主要呈负相关关系,其中3月平均气温降低对生长季始期推迟的影响最大,但降水与生长季始期的关系存在较大空间差异。植被生长季末期主要受9月降水增多的正向影响。就生长季长度而言,9月平均气温对生长季长度负向影响较大的像元点最多(40.46%),西北部主要受8月和9月降水的正向影响,东部主要受5月和6月降水的负向影响。
The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}