Progress on the origin, function and evolutionary mechanism of RHR transcription factor family
Lv Menggang,1,2, Liu Aijia1,2, Li Qingwei,1,2, Su Peng,1,21 College of Life Science, Liaoning Normal University, Dalian 116081, China 2 Lamprey Research Center, Liaoning Normal University, Dalian 116081, China;
the National Natural Science Foundation of China.31801973 the National Natural Science Foundation of China.31772884 Dalian Supports Innovation and Entrepreneurship Projects of High Level Talents.2019RQ126 the Chinese Major State Basic Research Development Program (973).2013CB835304 the Marine Public Welfare Project of the State Oceanic Administration.201305016 the project of Department of Ocean and Fisheries of Liaoning Province.201805 and the Science and Technology Innovation Fund Research Project.2018J12SN079
NFAT(nuclear factor of activated T cells)蛋白首次在T细胞中被发现[29,30]。NFAT家族有5位成员分别为NFAT1(NFATc2或NFATp)、NFAT2(NFATc1)、NFAT3(NFATc4)、NFAT4(NFATc3)和NFAT5[31]。NFAT家族蛋白包含两个TAD结构域,一个调节结构域(NFAT homology region,NHR),一个高度保守的Rel结构域和一个羧基末端结构域[32]。NFAT家族分为两类,分别为经典家族和非经典家族。NFAT蛋白的经典的家族成员包括:NFAT1、NFAT2、NFAT3和NFAT4,具有钙调蛋白结合位点,受Ca2+和钙调蛋白的调控[33,34]。大量研究表明,钙调神经磷酸酶与经典的NFAT蛋白结合并使其脱磷酸化,从而诱导其进入细胞核并发挥转录活性,非经典家族成员仅包括NFAT5,其并不需要钙调神经磷酸酶参与其活性[35]。NFAT5可以形成同源二聚体,其同源二聚对与DNA结合和转录活性至关重要[36]。
RBP (recombination signal binding protein)家族蛋白对于神经系统的发育和造血系统的分化具有至关重要的作用[71],并且对于神经细胞分化的作用在生物进化过程中是保守的[72⇓⇓~75]。后生动物RBP蛋白有两种亚型,一种是RBP-Jκ (recombination signal binding protein for immunoglobulin kappa J region),其在Notch信号通路中发挥重要作用;另一种是RBP-L (recombination signal binding protein for immunoglobulin kappa J region like),其功能并不依赖于Notch信号通路[76]。RBP蛋白由3个结构域组成:N端结构域(N-terminal domain, NTD)、β折叠结构域(β-trefoil domain, BTD)和C端结构域(C-terminal domain, CTD)[77]。经过对其序列分析和结构研究证实,RBP的NTD与NF-κB转录因子家族的Rel同源结构域的N端具有相似性[78]。并且NTD以单体形式通过识别特异性序列(C/T)GTGGGAA与DNA结合,行使类似于Rel同源结构域的功能[79]。BTD结构域可以促进这种和DNA的结合,而且还能够参与蛋白质与蛋白质的相互作用,然而CTD并不与DNA发生相互作用[77,78]。除此之外,RBP分别通过与辅助激活蛋白和辅助抑制蛋白的不同相互作用,作为转录的激活因子和抑制因子发挥双重作用[80]。
A:EBF祖先基因中H1H2a序列的外显子-内含子的结构。0相位内含子(红色横线),1相位内含子(黑色横线)。B:H2复制的第一步是复制编码H2a的外显子。由于相位剪切规则,通过复制和选择性剪切产生了H2d的祖先基因。C:第二步是激活H2d编码区下游的隐蔽3′剪切位点(浅绿色三角)。D:产生了相同相位的i11和i12内含子,并将成熟的H2d嵌入EBF蛋白。根据文献[70]修改绘制。 Fig. 2Mechanisms of H2 replication in vertebrate EBF proteins
LambertSA, JolmaA, CampitelliLF, DasPK, YinYM, AlbuM, ChenXT, TaipaleJ, HughesTR, WeirauchMT. The human transcription factors Cell, 2018, 175(2): 598- 599. [本文引用: 1]
WingenderE, SchoepsT, HaubrockM, KrullM, DönitzJ. TFClass: expanding the classification of human transcription factors to their mammalian orthologs Nucleic Acids Res, 2018, 46( D1): D343-D347. [本文引用: 2]
WangJL, WangJ, TianCY. Evolution of KRAB- containing zinc finger proteins and their roles in species evolution Hereditas(Beijing), 2016, 38(11): 971- 978. [本文引用: 1]
LaityJH, LeeBM, WrightPE. Zinc finger proteins: new insights into structural and functional diversity Curr Opin Struct Biol, 2001, 11(1): 39- 46. [本文引用: 1]
BerisioR, CiccarelliL, SquegliaF, DeSimone A, VitaglianoL. Structural and dynamic properties of incomplete immunoglobulin-like fold domains Protein Pept Lett, 2012, 19(10): 1045- 1053. [本文引用: 1]
SenR, BaltimoreD. Multiple nuclear factors interact with the immunoglobulin enhancer sequences Cell, 1986,46(5):705- 716. [本文引用: 1]
SuP, FengSS, LiQW.Research progress of the structure and function of NF-κB and IκB in different animal groups. Hereditas(Beijing), 2016, 38(6): 523- 531. [本文引用: 1]
GhoshS, HaydenMS. New regulators of NF-kappa B in inflammation Nat Rev Immunol, 2008, 8(11): 837- 848. [本文引用: 1]
BortolottoV, CuccurazzuB, CanonicoP L, GrilliM. NF-κB mediated regulation of adult hippocampal neurogenesis: relevance to mood disorders and antidepressant activity Biomed Res Int, 2014,612798. [本文引用: 1]
Al-KhodorS, PriceCT, KaliaA, Abu Kwaik Y. Functional diversity of ankyrin repeats in microbial proteins Trends Microbiol, 2010,18(3):132- 139. [本文引用: 1]
GauthierM, DegnanBM. The transcription factor NF- kappaB in the demosponge Amphimedon queenslandica : insights on the evolutionary origin of the Rel homology domain Dev Genes Evol, 2008, 218(1): 23- 32. [本文引用: 2]
ZhangQ, LenardoMJ, BaltimoreD. 30 years of NF-κB: a blossoming of relevance to human pathobiology Cell, 2017, 168( 1- 2): 37- 57. [本文引用: 1]
GhoshS, MayMJ, KoppEB. NF-kappaB and Rel proteins: evolutionarily conserved mediators of immune responses Annu Rev Immunol, 1998,16:225- 260. [本文引用: 1]
WangTT, SunYX, JinLJ, ThackerP, LiSY, XuYP. Aj-rel and Aj-p105, two evolutionary conserved NF-κB homologues in sea cucumber (Apostichopus japonicus) and their involvement in LPS induced immunity Fish Shellfish Immunol, 2013, 34(1): 17- 22. [本文引用: 1]
SullivanJC, WolenskiFS, ReitzelAM, FrenchCE, Traylor-KnowlesN, GilmoreTD, FinnertyJR. Two alleles of NF-kappaB in the sea anemone Nematostella vectensis are widely dispersed in nature and encode proteins with distinct activities PLoS One, 2009, 4(10): e7311. [本文引用: 3]
HuguetC, CrepieuxP, LaudetV. Rel/NF-kappaB transcription factors and IkappaB inhibitors: evolution from a unique common ancestor Oncogene, 1997,15(24):2965- 2974. [本文引用: 1]
SiggersT, ChangAB, TeixeiraA, WongD, WilliamsKJ, AhmedB, RagoussisJ, UdalovaIA, SmaleST, BulykML. Principles of dimer-specific gene regulation revealed by a comprehensive characterization of NF-κB family DNA binding Nat Immunol, 2011, 13(1): 95- 102.
DeJuan D, PazosF, ValenciaA.Emerging methods in protein co-evolution. Nat Rev Genet, 2013,14(4):249- 261. [本文引用: 1]
Sebé-PedrósA, deMendoza A, LangBF, DegnanBM, Ruiz-TrilloI.Unexpected repertoire of metazoan transcription factors in the unicellular holozoan Capsaspora owczarzaki. Mol Biol Evol, 2011,28(3):1241- 1254. [本文引用: 1]
WolenskiFS, GarbatiMR, LubinskiTJ, Traylor-KnowlesN, DresselhausE, StefanikDJ, GoucherH, FinnertyJR, GilmoreTD.Characterization of the core elements of the NF-κB signaling pathway of the sea anemone Nematostella vectensis. Mol Cell Biol, 2011,31(5):1076- 1087. [本文引用: 1]
ShawJP, UtzPJ, DurandDB, TooleJJ, EmmelEA, CrabtreeGR. Identification of a putative regulator of early T cell activation genes Science, 1988, 241(4862): 202- 205. [本文引用: 1]
HoganPG. Calcium-NFAT transcriptional signalling in T cell activation and T cell exhaustion Cell Calcium, 2017,63:66- 69. [本文引用: 1]
MüllerMR, RaoA. NFAT, immunity and cancer: a transcription factor comes of age. Nat Rev Immunol, 2010,10(9):645- 656. [本文引用: 1]
LeeJU, KimLK, ChoiJM. Revisiting the concept of targeting NFAT to control T cell immunity and autoimmune diseases Front Immunol, 2018,9:2747. [本文引用: 1]
MacianF. NFAT proteins: key regulators of T-cell development and function Nat Rev Immunol, 2005,5(6):472- 484. [本文引用: 1]
HoganPG, ChenL, NardoneJ, RaoA. Transcriptional regulation by calcium, calcineurin, and NFAT Genes Dev, 2003,17(18):2205- 2232. [本文引用: 1]
FeskeS, OkamuraH, HoganPG, RaoA. Ca 2+/calcineurin signalling in cells of the immune system Biochem Biophys Res Commun, 2003,311(4):1117- 1132. [本文引用: 1]
AramburuJ, López-RodríguezC. Regulation of inflammatory functions of macrophages and T lymphocytes by NFAT5 Front Immunol, 2019,10:535. [本文引用: 1]
SullivanJC, KalaitzidisD, GilmoreTD, FinnertyJR.Rel homology domain-containing transcription factors in the cnidarian Nematostella vectensis. Dev Genes Evol, 2007, 217(1): 63- 72. [本文引用: 1]
LiW, ZhengNZ, YuanQ, XuK, YangF, GuL, ZhengGY, LuoGJ, FanC, JiGJ, ZhangB, CaoH, TianXL. Erratum to: NFAT5-mediated CACNA1C expression is critical for cardiac electrophysiological development and maturation J Mol Med (Berl), 2016,94(9):1003- 1004. [本文引用: 1]
HuangXD, WeiGJ, ZhangH, HeMX. Nuclear factor of activated T cells (NFAT) in pearl oyster pinctada fucata : molecular cloning and functional characterization Fish Shellfish Immunol, 2015, 42(1): 108- 113. [本文引用: 1]
SongX, HuJ, JinP, ChenL, MaF. Identification and evolution of an NFAT gene involving Branchiostoma belcheri innate immunity Genomics, 2013,102(4):355- 362. [本文引用: 3]
KaiW, KikuchiK, TohariS, ChewAK, TayA, FujiwaraA, HosoyaS, SuetakeH, NaruseK, BrennerS, SuzukiY, VenkateshB. Integration of the genetic map and genome assembly of fugu facilitates insights into distinct features of genome evolution in teleosts and mammals Genome Biol Evol, 2011,3:424- 442. [本文引用: 1]
VihmaH, PruunsildP, TimmuskT. Alternative splicing and expression of human and mouse NFAT genes Genomics, 2008,92(5):279- 291. [本文引用: 1]
VechettiIJ Jr, AguiarAF, deSouza RW, AlmeidaFL, deAlmeida Dias HB, deAguiar Silva MA, CaraniFR, FerraressoRL, CarvalhoRF, Dal-Pai-SilvaM. NFAT isoforms regulate muscle fiber type transition without altering CaN during aerobic training Int J Sports Med, 2013, 34(10): 861- 867. [本文引用: 1]
UenoM, ShenWJ, PatelS, GreenbergAS, AzharS, KraemerFB. Fat-specific protein 27 modulates nuclear factor of activated T cells 5 and the cellular response to stress J Lipid Res, 2013, 54(3):734- 743. [本文引用: 1]
GraefIA, GastierJM, FranckeU, CrabtreeGR. Evolutionary relationships among Rel domains indicate functional diversification by recombination Proc Natl Acad Sci USA, 2001, 98(10): 5740- 5745. [本文引用: 1]
International Chicken Genome Sequencing Consortium. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution Nature, 2004,432(7018):695- 716. [本文引用: 1]
LeeN, KimD, KimWU. Role of NFAT5 in the immune system and pathogenesis of autoimmune diseases Front Immunol, 2019,10: 270. [本文引用: 1]
MeyerA, Van de Peer Y. From 2R to 3R: evidence for a fish-specific genome duplication (FSGD) Bioessays, 2005,27(9):937- 945. [本文引用: 1]
HoeggS, MeyerA. Hox clusters as models for vertebrate genome evolution Trends Genet, 2005,21(8):421- 424. [本文引用: 1]
HagmanJ, BelangerC, TravisA, TurckCW, GrosschedlR. Cloning and functional characterization of early B-cell factor, a regulator of lymphocyte-specific gene expression Genes Dev, 1993,7(5):760- 773. [本文引用: 1]
deTaffin M, CarrierY, DuboisL, BatailléL, PainsetA, LeGras S, JostB, CrozatierM, VincentA. Genome-wide mapping of collier in vivo binding sites highlights its hierarchical position in different transcription regulatory networks PLoS One, 2015, 10( 7): e0133387. [本文引用: 2]
WangMM, ReedRR. Molecular cloning of the olfactory neuronal transcription factor Olf-1 by genetic selection in yeast Nature, 1993, 364(6433):121- 126. [本文引用: 1]
LiaoD. Emerging roles of the EBF family of transcription factors in tumor suppression Mol Cancer Res, 2009, 7(12): 1893- 1901. [本文引用: 5]
FieldsS, TernyakK, GaoH, OstraatR, AkerlundJ, HagmanJ. The 'zinc knuckle' motif of early B cell factor is required for transcriptional activation of B cell-specific genes Mol Immunol, 2008,45(14):3786- 3796. [本文引用: 1]
SiponenMI, WisniewskaM, LehtiöL, JohanssonI, SvenssonL, RaszewskiG, NilssonL, SigvardssonM, BerglundH. Structural determination of functional domains in early B-cell factor (EBF) family of transcription factors reveals similarities to Rel DNA-binding proteins and a novel dimerization motif J Biol Chem, 2010,285(34):25875- 25879. [本文引用: 2]
PangK, MatusDQ, MartindaleMQ. The ancestral role of COE genes may have been in chemoreception: evidence from the development of the sea anemone Nematostella vectensis (Phylum Cnidaria; Class Anthozoa). Dev Genes Evol, 2004, 214(3): 134- 138. [本文引用: 1]
StolfiA, GainousTB, YoungJJ, MoriA, LevineM, ChristiaenL. Early chordate origins of the vertebrate second heart field Science, 2010, 329(5991): 565- 568. [本文引用: 1]
KimK, ColosimoME, YeungH, SenguptaP. The UNC-3 Olf/EBF protein represses alternate neuronal programs to specify chemosensory neuron identity Dev Biol, 2005, 286( 1): 136- 148. [本文引用: 1]
MazetF, MasoodS, LukeGN, HollandND, ShimeldSM. Expression of AmphiCoe, an amphioxus COE/EBF gene, in the developing central nervous system and epidermal sensory neurons Genesis, 2004, 38(2): 58- 65. [本文引用: 1]
Lara-RamírezR, PonceletG, PattheyC, ShimeldSM.splicingsplicing, The structure, synteny and expression of lamprey COE genes and the evolution of the COE gene family in chordates Dev Genes Evol, 2017,227(5):319- 338. [本文引用: 2]
PozzoliO, BosettiA, CrociL, ConsalezGG, VetterML.Xebf3 is a regulator of neuronal differentiation during primary neurogenesis in Xenopus. Dev Biol, 2001, 233( 2): 495- 512. [本文引用: 1]
CatelaC, CorreaE, WenK, AburasJ, CrociL, ConsalezGG, KratsiosP. An ancient role for collier/Olf/Ebf (COE)-type transcription factors in axial motor neuron development Neural Dev, 2019, 14( 1): 2. [本文引用: 1]
TreiberN, TreiberT, ZocherG, GrosschedlR. Structure of an EBF1:DNA complex reveals unusual DNA recognition and structural homology with Rel proteins Genes Dev, 2010, 24(20): 2270- 2275. [本文引用: 2]
BollerS, GrosschedlR. The regulatory network of B-cell differentiation: a focused view of early B-cell factor 1 function Immunol Rev, 2014, 261( 1): 102- 115. [本文引用: 1]
DaburonV, MellaS, PlouhinecJL, MazanS, CrozatierM, VincentA. The metazoan history of the COE transcription factors. Selection of a variant HLH motif by mandatory inclusion of a duplicated exon in vertebrates BMC Evol Biol, 2008,8:131. [本文引用: 2]
PursgloveSE, MackayJP. CSL: a notch above the rest Int J Biochem Cell Biol, 2005, 37(12): 2472- 2477. [本文引用: 1]
MaierD. The evolution of transcriptional repressors in the notch signaling pathway: a computational analysis Hereditas, 2019, 156( 1): 5. [本文引用: 1]
SiegerD, TautzD, GajewskiM. The role of suppressor of hairless in notch mediated signalling during zebrafish somitogenesis Mech Dev, 2003,120(9):1083- 1094. [本文引用: 1]
ItoM, KatadaT, MiyataniS, KinoshitaT. XSu(H)2 is an essential factor for gene expression and morphogenesis of the Xenopus gastrula embryo Int J Dev Biol, 2007, 51( 1): 27- 36. [本文引用: 1]
WilsonJJ, KovallRA. Crystal structure of the CSL-notch- mastermind ternary complex bound to DNA. Cell, 2006,124(5):985- 996. [本文引用: 2]
HallDP, KovallRA. Structurally conserved binding motifs of transcriptional regulators to notch nuclear effector CSL. Exp Biol Med (Maywood ), 2019, 244(17): 1520- 1529. [本文引用: 2]
OakleyF, MannJ, RuddellRG, PickfordJ, WeinmasterG, MannDA. Basal expression of IkappaBalpha is controlled by the mammalian transcriptional repressor RBP-J (CBF1) and its activator notch1 J Biol Chem, 2003, 278(27): 24359- 24370. [本文引用: 1]
CollinsKJ, YuanZ, KovallRA. Structure and function of the CSL-KyoT2 corepressor complex: a negative regulator of notch signaling Structure, 2014, 22( 1): 70- 81. [本文引用: 1]
HedgesSB. The origin and evolution of model organisms Nat Rev Genet, 2002, 3(11): 838- 849. [本文引用: 1]
CiccarelliFD, DoerksT, vonMering C, CreeveyCJ, SnelB, BorkP. Toward automatic reconstruction of a highly resolved tree of life Science, 2006,311(5765):1283- 1287. [本文引用: 1]
BeresTM, MasuiT, SwiftGH, ShiL, HenkeRM, MacDonaldRJ. PTF1 is an organ-specific and notch- independent basic helix-loop-helix complex containing the mammalian suppressor of hairless (RBP-J) or its paralogue, RBP-L Mol Cell Biol, 2006, 26( 1): 117- 130. [本文引用: 1]
MinoguchiS, TaniguchiY, KatoH, OkazakiT, StroblLJ, Zimber-StroblU, BornkammGW, HonjoT. RBP-L, a transcription factor related to RBP-Jkappa Mol Cell Biol, 1997,17(5):2679- 2687. [本文引用: 1]
 ,1,2, 刘艾嘉1,2, 李庆伟
,1,2, 刘艾嘉1,2, 李庆伟
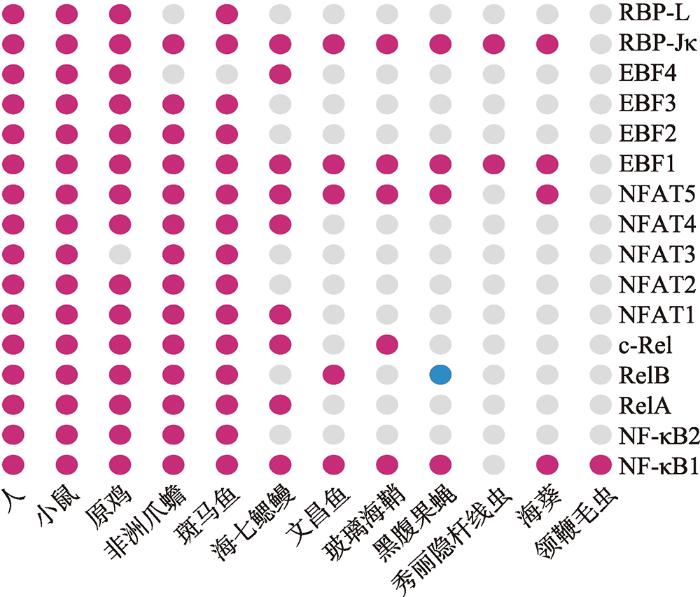 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT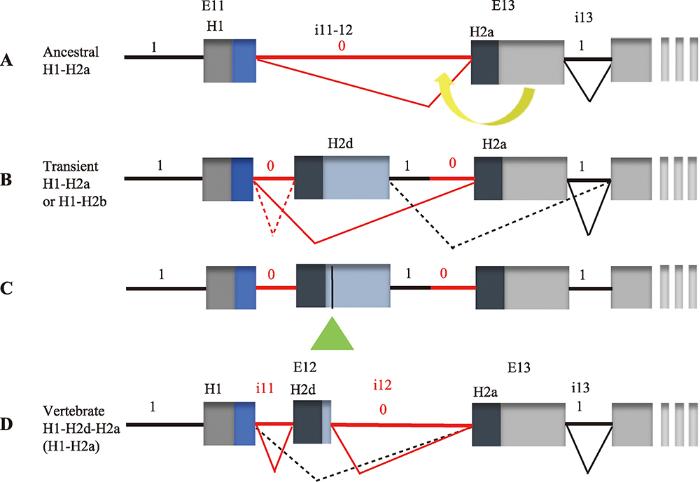 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT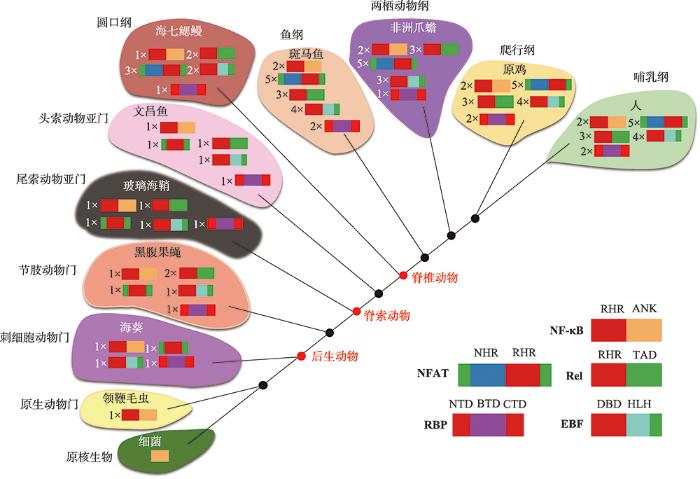 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}