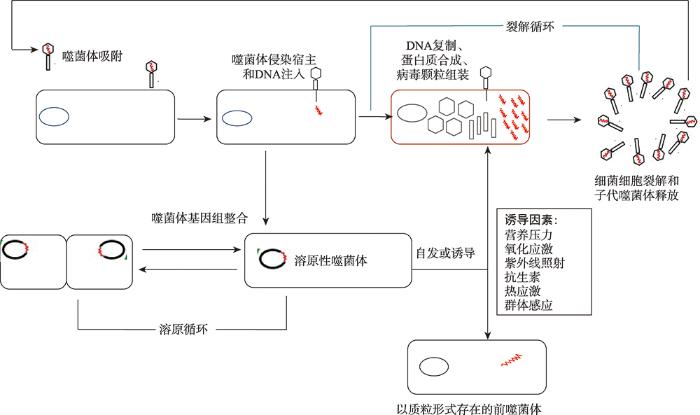
噬菌体采用不同的生命周期:烈性噬菌体遵循其复制的裂解周期,并在周期结束时导致宿主细胞的裂解;温和噬菌将其基因组整合到宿主细菌染色体中。前噬菌体DNA与细菌染色体一起复制,并传给子细胞。在某些条件下,包括造成DNA损伤的各种压力,前噬体会被切除并进入裂解循环,而一些噬菌体也会以质粒的形式存在。根据参考文献[2]修改绘制。 Fig. 1Different life cycles of phages
宿主细胞密度影响噬菌体裂解-溶原的转换,表明温和噬菌体可能利用特定的机制来感知宿主细胞密度。早期细胞生理学和最适感染复数(multiplicity of infection, MOI)有利于溶原性,这表明可能在共感染噬菌体之间发生一些细胞内通讯,以做出有利于两种入侵噬菌体的选择。但是,共同感染的噬菌体如何在裂解和溶原这两个相反的生存途径上折中仍然是个难题。Ghosh等[62]首次证明依赖细胞密度的前噬菌体诱导与群体感应系统有关。宿主细胞内的噬菌体具有进化的机制以感知其宿主密度,并在溶原-裂解之间做出协调决定,以便噬菌体可以根据宿主细胞密度估计后代噬菌体的成功感染率,从而最大限度地繁殖和避免耗尽敏感宿主。
FernándezL, RodríguezA, GarcíaP. Phage or foe: an insight into the impact of viral predation on microbial communities ISME J , 2018,12(5):1171- 1179. [本文引用: 1]
AkhterS, AzizRK, EdwardsRA. PhiSpy: a novel algorithm for finding prophages in bacterial genomes that combines similarity- and composition-based strategies Nucleic Acids Res , 2012,40( 16): e126. [本文引用: 1]
RouxS, EnaultF, HurwitzBL, SullivanMB. VirSorter: mining viral signal from microbial genomic data PeerJ , 2015,3: e985. [本文引用: 1]
RenJ, AhlgrenNA, LuYY, FuhrmanJA, SunFZ. VirFinder: a novel k-mer based tool for identifying viral sequences from assembled metagenomic data Microbiome . 2017, 65( 1): 69. [本文引用: 1]
JurtzVI, VillarroelJ, LundO, VoldbyLarsen M, NielsenM. MetaPhinder-Identifying bacteriophage sequences in metagenomic data sets PLoS One , 2016,11( 9): e0163111. [本文引用: 1]
ArndtD, MarcuA, LiangYJ, WishartDS. Phast, Phaster and Phastest: Tools for finding prophage in bacterial genomes Brief Bioinform , 2019,20(4):1560- 1567. [本文引用: 1]
AmgartenD, BragaLPP, daSilva AM, SetubalJC. Marvel, a tool for prediction of bacteriophage sequences in metagenomic bins Front Genet , 2018,9: 304. [本文引用: 1]
SongWC, SunHX, ZhangC, ChengL, PengY, DengZQ, WangD, WangY, HuM, LiuW, YangHM, ShenY, LiJH, YouLC, XiaoMF. Prophage Hunter: an integrative hunting tool for active prophages Nucleic Acids Res , 2019; 47( W1): W74-W80. [本文引用: 1]
StevensRH, ZhangHM, SedgleyC, BergmanA, MandaAR. The prevalence and impact of lysogeny among oral isolates of Enterococcus faecalis J Oral Microbiol , 2019, 11( 1): 1643207. [本文引用: 2]
MaRJ. Citromicrobium bathyomarinum: diversity, activity and interaction with hosts Xiamen Univ , 2018. [本文引用: 1]
SchmiegerH, SchicklmaierP. Transduction of multiple drug resistance of Salmonella enterica serovar typhimurium DT104 FEMS Microbiol Lett , 1999, 170(1): 251- 256. [本文引用: 1]
SchicklmaierP, MoserE, WielandT, RabschW, SchmiegerH. A comparative study on the frequency of prophages among natural isolates of Salmonella and Escherichia coli with emphasis on generalized transducers Antonie Van Leeuwenhoek , 1998, 73(1): 49- 54. [本文引用: 2]
OsawaRO, IyodaS, NakayamaSI, WadaA, YamaiS, WatanabeH. Genotypic variations of Shiga toxin-converting phages from enterohaemorrhagic Escherichia coli O157: H7 isolates J Med Microbiol , 2000, 49(6): 565- 574. [本文引用: 1]
YamamotoN. Genetic evolution of bacteriophage. I. Hybrids between unrelated bacteriophages P22 and Fels 2 Proc Natl Acad Sci USA , 1969, 62(1): 63- 69. [本文引用: 1]
McClellandM, FloreaL, SandersonK, CliftonSW, ParkhillJ, ChurcherC, DouganG, WilsonRK, MillerW. Comparison of the Escherichia coli K-12 genome with sampled genomes of a Klebsiella pneumoniae and three salmonella enterica serovars, Typhimurium, Typhi and Paratyphi Nucleic Acids Res , 2000, 28(24): 4974- 4986. [本文引用: 1]
Figueroa-BossiN, BossiL. Inducible prophages contribute to Salmonella virulence in mice Mol Microbiol , 1999, 33(1): 167- 176. [本文引用: 1]
PoppA, HertwigS, LurzR, AppelB. Comparative study of temperate bacteriophages isolated from Yersinia Syst Appl Microbiol , 2000,23(4):469- 478. [本文引用: 1]
HugginsAR, SandineWE.Incidence and properties of temperate bacteriophages induced from lactic streptococci. Appl Environ Microbiol , 1977, 33(1): 184- 191. [本文引用: 1]
Dominguez-MirazoM, JinR.WeitzJS. Functional and Comparative Genomic Analysis of Integrated Prophage-Like Sequences in “ Candidatus Liberibacter asiaticus” mSphere , 2019,4(6): e00409-e00419. [本文引用: 2]
CastilloD, MiddelboeM.Genomic diversity of bacteriophages infecting the fish pathogen Flavobacterium psychrophilum. FEMS Microbiol Lett , 2016,363(24): fnw272. [本文引用: 1]
PetrovVM, RatnayakaS, NolanJM, MillerES, KaramJD. Genomes of the T4-related bacteriophages as windows on microbial genome evolution Virol J , 2010,7: 292. [本文引用: 1]
BlaisdellBE, CampbellAM, KarlinS. Similarities and dissimilarities of phage genomes Proc Natl Acad Sci USA , 1996, 93(12): 5854- 5859. [本文引用: 1]
AgronPG, WalkerRL, KindeH, SawyerSJ, HayesDC, WollardJ, AndersenGL. Identification by subtractive hybridization of sequences specific for Salmonella enterica serovar enteritidis Appl Environ Microbiol , 2001, 67(11): 4984- 4991. [本文引用: 1]
ChangCC, GilsdorfJR, DiRitaVJ, MarrsCF. Identification and genetic characterization of Haemophilus influenzae genetic island 1 Infect Immun , 2000,68(5):2630- 2637. [本文引用: 1]
DepMS, MendzGL, TrendMA, ColoePJ, FryBN, KorolikV. Differentiation between Campylobacter hyoilei and Campylobater coli using genotypic and phenotypic analyses Int J Syst Evol Microbiol , 2001,51(Pt 3):819- 826. [本文引用: 1]
KleeSR, NassifX, KusecekB, MerkerP, BerettiJL, AchtmanM, TinsleyCR. Molecular and biological analysis of eight genetic islands that distinguish Neisseria meningitidis from the closely related pathogen Neisseria gonorrhoeae Infect Immun , 2000,68(4):2082- 2095. [本文引用: 1]
BrandtK, Tilsala-TimisjärviA, AlatossavaT.Phage-related DNA polymorphism in dairy and probiotic Lactobacillus. Micron , 2001, 32(1): 59- 65. [本文引用: 1]
KrishnamurthiR, GhoshS, KhedkarS, SeshasayeeASN. Repression of YdaS toxin is mediated by transcriptional repressor RacR in the cryptic rac prophage of Escherichia coli K-12 mSphere , 2017,2(6):e00392- 17. [本文引用: 1]
MehtaP, CasjensS, KrishnaswamyS. Analysis of the lambdoid prophage element e14 in the E.coli K-12 genome BMC Microbiol , 2004,4: 4. [本文引用: 1]
JingTT, FuZW, ShenP, ChenXD. General investigation of PBSX-like defective prophages resident in Bacillus subtilis strains Microbiology , 2016, 43(6): 1244- 1252. [本文引用: 1]
RuzinA, LindsayJ, NovickRP. Molecular genetics of SaPI1--a mobile pathogenicity island in Staphylococcus aureus. Mol Microbiol 2001, 41(2): 365- 377. [本文引用: 1]
RezaieN, BakhshiB, Najar-PeerayehS. The role of CTX and RS1 satellite phages genomic arrangement in Vibrio cholera toxin production in two recent cholera outbreaks (2012 and 2013) in IR Iran Microb Pathog , 2017,112: 89- 94. [本文引用: 1]
DziewitL, RadlinskaM. Two inducible prophages of an antarctic Pseudomonas sp . ANT_H14 use the same capsid for packaging their genomes - characterization of a novel phage helper-satellite system PLoS One , 2016, 11(7): e0158889. [本文引用: 1]
DieterleME, FinaMartin J, DuránR, NemirovskySI, RivasCS, BowmanC, RussellD, HatfullGF, CambillauC, PiuriM. Characterization of prophages containing “evolved” Dit/Tal modules in the genome of Lactobacillus casei BL23 Appl Microbiol Biotechnol , 2016, 100(21): 9201- 9215. [本文引用: 1]
CrestaniC, FordeTL, ZadoksRN. Development and application of a prophage integrase typing scheme for group B Streptococcus. Front Microbiol , 2020,11: 1993. [本文引用: 1]
ChenJ, ZhuY, YinM, XuY, LiangX, HuangYP. Characterization of maltocin S16, a phage tail-like bacteriocin with antibacterial activity against Stenotrophomonas maltophilia and Escherichia coli J Appl Microbiol , 2019, 127(1): 78- 87. [本文引用: 1]
NakayamaK, TakashimaK, IshiharaH, ShinomiyaT, KageyamaM, KanayaS, OhnishiM, MurataT, MoriH, HayashiT. The R-type pyocin of Pseudomonas aeruginosa is related to P2 phage, and the F-type is related to lambda phage Mol Microbiol , 2000, 38( 2): 213- 231. [本文引用: 1]
LotzW, MayerF. Isolation and characterization of a bacteriophage tail-like bacteriocin from a strain of Rhizobium J Virol , 1972, 9(1): 160- 173. [本文引用: 1]
LangAS, ZhaxybayevaO, BeattyJT. Gene transfer agents: phage-like elements of genetic exchange Nat Rev Microbiol , 2012, 10(7): 472- 482. [本文引用: 1]
MarrsB. Genetic recombination in Rhodopseudomonas capsulata Proc Natl Acad Sci USA , 1974,71(3):971- 973. [本文引用: 1]
WahlLM.PattendenT. Prophage provide a safe haven for adaptive exploration in temperate viruses Genetics , 2017, 206(1): 407- 416. [本文引用: 1]
GentileGM, WetzelKS, DedrickRM, MontgomeryMT, GarlenaRA, Jacobs-SeraD, HatfullGF. More evidence of collusion: a new prophage-mediated viral defense system encoded by mycobacteriophage Sbash mBio , 2019, 10(2): e00196- 19. [本文引用: 1]
ChenYF, LiuSY, LiangZB, LuMF, ZhouJL, ZhangLH. Quorum sensing and microbial drug resistance Hereditas (Beijing) , 201638 (10): 881- 893. [本文引用: 1]
YuMK, KimMA, RosaV, HwangYC, DelFabbro M, SohnWJ, MinKS. Role of extracellular DNA in Enterococcus faecalis biofilm formation and its susceptibility to sodium hypochlorite J Appl Oral Sci , 2019,27: e20180699. [本文引用: 1]
ShenMY, YangYH, ShenW, CenLJ, McLeanJS, ShiWY, LeS, HeXS. A linear plasmid-like prophage of actinomyces odontolyticus promotes biofilm assembly Appl Environ Microbiol , 2018, 84(17): e01263- 18. [本文引用: 2]
RiceSA, TanCH, MikkelsenPJ, KungV, WooJ, TayM, HauserA, McDougaldD, WebbJS, KjellebergS. The biofilm life cycle and virulence of Pseudomonas aeruginosa are dependent on a filamentous prophage ISME J , 2009,3(3):271- 282. [本文引用: 1]
RossmannFS, RacekT, WobserD, PuchalkaJ, RabenerEM, ReigerM, HendrickxAA, DiederichAK, JungK, KleinC, HuebnerJ. Phage-mediated dispersal of biofilm and distribution of bacterial virulence genes is induced by quorum sensing PLoS Pathog , 2015, 11( 2): e1004653. [本文引用: 1]
UnterholznerSJ, PoppenbergerB, RozhonW. Toxin- antitoxin systems: biology, identification, and application Mob Genet Elements , 2013,3(5): e26219. [本文引用: 1]
ZanderI, ShmidovE, RothS, Ben-DavidY, ShovalI, ShoshaniS, DanielliA, BaninE. Characterization of PfiT/PfiA toxin-antitoxin system of Pseudomonas aeruginosa that affects cell elongation and prophage induction Environ Microbiol , 2020, 22(12): 5048- 5057. [本文引用: 1]
LiYM, LiuXX, TangKH, WangWQ, WangYX, WangXX. Prophage encoding toxin/antitoxin system PfiT/PfiA inhibits Pf4 production in Pseudomonas aeruginosa. Microb Biotechnol , 2020. 13(4): 1132- 1144. [本文引用: 1]
GuoYX, QuirogaC, ChenQ, McAnultyMJ, BenedikMJ, WoodTK, WangXX.RalR (a DNase) and RalA (a small RNA) form a type I toxin-antitoxin system in Escherichia coli. Nucleic Acids Res , 2014, 42(10): 6448- 6462. [本文引用: 1]
ChenYY, WangJT, LinTL Gong YN, LiTH, HuangYY, HsiehYC. Prophage excision in Streptococcus pneumoniae serotype 19A ST320 promote colonization: insight into its evolution from the ancestral clone taiwan 19F-14 (ST236 ). Front Microbiol , 2019,10: 205. [本文引用: 1]
GhoshD, RoyK, WilliamsonKE, SrinivasiahS, WommackKE, RadosevichM. Acyl-homoserine lactones can induce virus production in lysogenic bacteria: an alternative paradigm for prophage induction Appl Environ Microbiol , 2009, 75(22): 7142- 7152. [本文引用: 1]
PawlukA, StaalsRHJ, TaylorC, WatsonBNJ, SahaS, FineranPC, MaxwellKL, DavidsonAR. Inactivation of CRISPR-Cas systems by anti-CRISPR proteins in diverse bacterial species Nat Microbiol , 2016,1(8): 16085. [本文引用: 2]
Bondy-DenomyJ, PawlukA, MaxwellKL, DavidsonAR. Bacteriophage genes that inactivate the CRISPR/Cas bacterial immune system Nature , 2013, 493(7432): 429- 432. [本文引用: 3]
 ,1, 魏云林1, 季秀玲
,1, 魏云林1, 季秀玲
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}