The mechanism underlying histone deacetylases regulating cardiac hypertrophy
Lian Ren, Xiushan Wu, Yongqing Li,State Key Lab of Development Biology of Freshwater Fish, Key Laboratory of the Ministry of Education, Heart Development Research Center, College of Life Sciences, Hunan Normal University, Changsha 410081, China
Supported by the National Natural Science Foundation of China No.81470377 Hunan Province Biological Development Engineering and New Product R & D Collaborative Innovation Center No.2013-448-6
作者简介 About authors 任恋,在读博士研究生,专业方向:分子遗传。E-mail:82287168@qq.com。
Abstract Cardiac hypertrophy is a compensatory response that occurs as a result of increased hemodynamic requirement in peripheral tissues. In the process of cardiac hypertrophy, the expression of different types of genes in different stages is transcriptionally regulated by multiple-level physiological and pathological signals. Histone acetylation, as the most extensive post-translational modification, is closely controlled by the antagonistic histone acetyltransferases (HAT) and histone deacetylases (HDACs). Recent studies have shown that HDACs, as a family of enzymes that inhibit transcription and contain highly conserved deacetylase domains, regulate gene expression during cardiac hypertrophy through a variety of pathways. In this review, we mainly summarize the research progress on histone deacetylase in cardiac hypertrophy. By elucidating the role and molecular mechanism of different HDACs in cardiac hypertrophy, it provides new ideas for the treatment of different types of cardiac hypertrophy and heart failure, and molecular targets for new drug design. Keywords:myocardial hypertrophy;post-translational modification;transcription process;histone deacetylases;molecular mechanism
PDF (700KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 任恋, 吴秀山, 李永青. 组蛋白去乙酰化酶在调节心肌肥大过程中的作用机制. 遗传[J], 2020, 42(6): 536-547 doi:10.16288/j.yczz.19-346 Lian Ren. The mechanism underlying histone deacetylases regulating cardiac hypertrophy. Hereditas(Beijing)[J], 2020, 42(6): 536-547 doi:10.16288/j.yczz.19-346
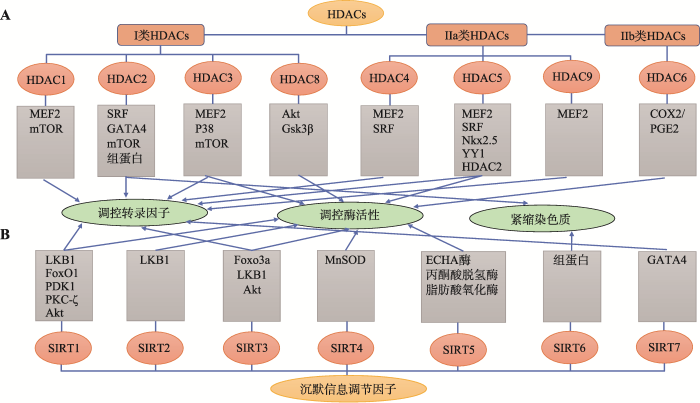
HDAC8也被证明可调节心肌肥大。在醋酸去氧皮质酮-盐诱导高血压的大鼠模型中,HDACs抑制剂丙戊酸钠处理使HDAC6和HDAC8的酶活性下降,抑制慢性高血压大鼠的心肌肥大及纤维化[24]。Yan等[25]研究显示miR-21-3p抑制TAC和血管紧张素(angiotensin II, Ang II)诱导的心肌肥大的作用靶点是沉默HDAC8;HDAC8的表达通过提升磷酸化Akt和磷酸化Gsk3β而削弱miR-21-3p介导的心肌肥大抑制作用。Li等 [20] 研究也表明HDAC8和HDAC2在肾性高血压大鼠的心脏重塑中起关键作用。有关I类HDACs调控心肌肥大的相关作用机制见图2。
KouzaridesT . Acetylation: a regulatory modification to rival phosphorylation? EMBO J, 2000,19(6):1176-1179. [本文引用: 1]
TauntonJ, HassigCA, SchreiberSL . A mammalian histone deacetylase related to the yeast transcriptional regulator Rpd3p Science, 1996,272(5260):408-411. [本文引用: 1]
JonesP, AltamuraS, De FrancescoR, GallinariP, LahmA, NeddermannP, RowleyM, SerafiniS, SteinkühlerC . Probing the elusive catalytic activity of vertebrate class IIa histone deacetylases Bioorg Med Chem Lett, 2008,18(6):1814-1819. [本文引用: 1]
DuanBY, YeD, ZhuSC, JiaWW, LuCQ, WangGY, GuoXD, YuYY, WuCY, KangJH . HDAC10 promotes angiogenesis in endothelial cells through the PTPN22/ ERK axis Oncotarget, 2017,8(37):61338-61349. [本文引用: 1]
HoutkooperRH, PirinenE, AuwerxJ . Sirtuins as regulators of metabolism and healthspan Nat Rev Mol Cell Biol, 2012,13(4):225-238. [本文引用: 1]
CaoJ, SunL, AramsangtienchaiP, SpiegelmanNA, ZhangXY, SetoE, LinHN . DAC11 regulates type I interferon signaling through defatty-acylation of SHMT2 Proc Natl Acad Sci USA, 2019,116(12):5487-5492. [本文引用: 1]
MoralesCR, LiDL, PedrozoZ, MayHI, JiangN, KyrychenkoV, ChoGW, KimSY, WangZV, RotterD, RothermelBA, SchneiderJW, LavanderoS, GilletteTG, Hill JA. Inhibition of class I histone deacetylases blunts cardiac hypertrophy through TSC2-dependent mTOR repression Sci Signal, 2016, 9(422): ra34. [本文引用: 4]
LaggerG, O'Carroll D, RemboldM, KhierH, TischlerJ, WeitzerG, SchuettengruberB, HauserC, BrunmeirR, JenuweinT, SeiserC . Essential function of histone deacetylase 1 in proliferation control and CDK inhibitor repression EMBO J, 2002,21(11):2672-2681. [本文引用: 3]
YangXJ, SetoE . Collaborative spirit of histone deacetylases in regulating chromatin structure and gene expression Curr Opin Genet Dev, 2003,13(2):143-153. [本文引用: 1]
EomGH, KookH . Role of histone deacetylase 2 and its posttranslational modifications in cardiac hypertrophy BMB Rep, 2015,48(3):131-138. [本文引用: 2]
RaghunathanS, GoyalRK, PatelBM . Selective inhibition of HDAC2 by magnesium valproate attenuates cardiac hypertrophy Can J Physiol Pharmacol, 2017,95(3):260-267. [本文引用: 1]
KookH, LeporeJJ, GitlerAD, LuMM, Wing-Man Yung W, MackayJ, ZhouR, FerrariV, GruberP, EpsteinJA . Cardiac hypertrophy and histone deacetylase-dependent transcriptional repression mediated by the atypical homeodomain protein Hop J Clin Invest, 2003,112(6):863-871. [本文引用: 1]
TrivediCM, ZhuW, WangQ, JiaC, KeeHJ, LiL, HannenhalliS, EpsteinJA . Hopx and Hdac2 interact to modulate Gata4 acetylation and embryonic cardiac myocyte proliferation Dev Cell, 2010,19(3):450-459. [本文引用: 1]
LiRF, CaoSS, FangWJ, SongY, LuoXT, WangHY, WangJG . Roles of HDAC2 and HDAC8 in cardiac remodeling in renovascular hypertensive rats and the effects of valproic acid sodium Pharmacology, 2017,99(1-2):27-39. [本文引用: 3]
PillaiVB, SundaresanNR, SamantSA, WolfgeherD, TrivediCM, GuptaMP . Acetylation of a conserved lysine residue in the ATP binding pocket of p38 augments its kinase activity during hypertrophy of cardiomyocytes Mol Cell Biol, 2011,31(11):2349-2363. [本文引用: 2]
TrivediCM, LuMM, WangQH, EpsteinJA . Transgenic over-expression of Hdac3 in the heart produces increased postnatal cardiac myocyte proliferation but does not induce hypertrophy J Biol Chem, 2008,283(39):26484-26489. [本文引用: 2]
KeeHJ, BaeEH, ParkS, LeeKE, SuhSH, KimSW, JeongMH. HDAC inhibition suppresses cardiac hypertrophy and fibrosis in DOCA-salt hypertensive rats via regulation of HDAC6/HDAC8 enzyme activity Kidney Blood Press Res, 2013,37(4-5):229-239. [本文引用: 4]
ZhangCL, McKinsey TA, ChangS, AntosCL, HillJA, OlsonEN, . Class II histone deacetylases act as signal- responsive repressors of cardiac hypertrophy Cell, 2002,110(4):479-488. [本文引用: 2]
ZhangCL, McKinsey TA, Olson EN. The transcriptional corepressor MITR is a signal-responsive inhibitor of myogenesis Proc Natl Acad Sci USA, 2001,98(13):7354-7359. [本文引用: 2]
KeeHJ, BaeEH, ParkS, LeeKE, SuhSH, KimSW, JeongMH. HDAC inhibition suppresses cardiac hypertrophy and fibrosis in DOCA-salt hypertensive rats via regulation of HDAC6/HDAC8 enzyme activity Kidney Blood Press Res, 2013,37(4-5):229-239. [本文引用: 2]
Demos-DaviesKM, FergusonBS, CavasinMA, MahaffeyJH, WilliamsSM, SpiltoirJI, SchuetzeKB, HornTR, ChenB, FerraraC, ScelliniB, PiroddiN, TesiC, PoggesiC, JeongMY, McKinsey TA. HDAC6 contributes to pathological responses of heart and skeletal muscle to chronic angiotensin-II signaling Am J Physiol Heart Circ Physiol, 2014,307(2):H252-258. [本文引用: 4]
MaS, FengJ, ZhangR, ChenJW, HanD, LiX, YangB, LiXJ, FanMM, LiCY, TianZH, WangYB, CaoF . SIRT1 activation by resveratrol alleviates cardiac dysfunction via mitochondrial regulation in diabetic cardiomyopathy mice Oxid Med Cell Longev, 2017,2017:4602715. [本文引用: 2]
SundaresanNR, PillaiVB, WolfgeherD, SamantS, VasudevanP, ParekhV, RaghuramanH, CunninghamJM, GuptaM, GuptaMP. The deacetylase sirt1 promotes membrane localization and activation of akt and pdk1 during tumorigenesis and cardiac hypertrophy Sci Signal, 2011, 4(182): ra46. [本文引用: 2]
LiJY, HuangJY, LuJ, GuoZ, LiZM, GaoH, WangPX, LuoWW, CaiSD, HuYH, GuoKT, WangLP, LiZZ, WangMH, ZhangXL, LiuPQ . Sirtuin 1 represses PKC-ζ activity through regulating interplay of acetylation and phosphorylation in cardiac hypertrophy Br J Pharmacol, 2019,176(3):416-435. [本文引用: 1]
TangXQ, ChenXF, WangNY, WangXM, LiangST, ZhengW, LuYB, ZhaoX, HaoDL, ZhangZQ, ZouMH, LiuDP, ChenHZ . SIRT2 acts as a cardioprotective deacetylase in pathological cardiac hypertrophy Circulation, 2017,136(21):2051-2067. [本文引用: 2]
PillaiVB, BinduS, SharpW, FangYH, KimG, GuptaM, SamantS, GuptaMP . Sirt3 protects mitochondrial DNA damage and blocks the development of doxorubicin- induced cardiomyopathy in mice Am J Physiol Heart Circ Physiol, 2016,310(8):H962-972. [本文引用: 2]
SundaresanNR, GuptaM, KimG, RajamohanSB, IsbatanA, GuptaMP . Sirt3 blocks the cardiac hypertrophic response by augmenting foxo3a-dependent antioxidant defense mechanisms in mice J Clin Invest, 2009,119(9):2758-2771. [本文引用: 2]
PillaiVB, SundaresanNR, KimG, GuptaM, RajamohanSB, PillaiJB, SamantS, RavindraPV, IsbatanA, GuptaMP . Exogenous nad blocks cardiac hypertrophic response via activation of the sirt3-lkb1-amp-activated kinase pathway J Biol Chem, 2010,285(5):3133-3144. [本文引用: 2]
SchuelerM, ZhangQ, SchlesingerJ, T?njesM, SperlingSR . Dynamics of Srf, p300 and histone modifications during cardiac maturation in mouse Mol Biosyst, 2012,8(2):495-503. [本文引用: 1]
SchlesingerJ, SchuelerM, GrunertM, FischerJJ, ZhangQ, KruegerT, LangeM, T?njesM, DunkelI, SperlingSR . The cardiac transcription network modulated by Gata4, Mef2a, Nkx2.5, Srf, histone modifications, and microRNAs PLoS Genet, 2011,7(2):e1001313. [本文引用: 1]
MathiasRA, GuiseAJ, CristeaIM . Post-translational modifications regulate class IIa histone deacetylase (HDAC) function in health and disease Mol Cell Proteomics, 2015,14(3):456-470. [本文引用: 1]
VegaRB, HarrisonBC, MeadowsE, RobertsCR, PapstPJ, OlsonEN , McKinsey TA. Protein kinases C and D mediate agonist-dependent cardiac hypertrophy through nuclear export of histone deacetylase 5 Mol Cell Biol, 2004,24(19):8374-8385. [本文引用: 1]
McKinseyTA, ZhangCL, LuJ, OlsonEN . Signal-dependent nuclear export of a histone deacetylase regulates muscle differentiation Nature, 2000,408(6808):106-111. [本文引用: 1]
SparrowDB, MiskaEA, LangleyE, Reynaud-DeonauthS, KotechaS, TowersN, SpohrG, KouzaridesT, MohunTJ . MEF-2 function is modified by a novel co-repressor, MITR EMBO J, 1999,18(18):5085-5098. [本文引用: 1]
Morin S1, CharronF, RobitailleL, NemerM . GATA- dependent recruitment of MEF2 proteins to target promoters EMBO J, 2000,19(9):2046-2055. [本文引用: 1]
YounHD, ChatilaTA, LiuJO . Integration of calcineurin and MEF2 signals by the coactivator p300 during T-cell apoptosis EMBO J, 2000,19(16):4323-4331. [本文引用: 1]
ZhangM, HagenmuellerM, RiffelJH, KreusserMM, BernholdE, FanJ, KatusHA, BacksJ, HardtSE . Calcium/Calmodulin-dependent protein kinase II couples Wnt signaling with histone deacetylase 4 and mediates dishevelled-induced cardiomyopathy Hypertension, 2015,65(2):335-344. [本文引用: 1]
CalalbMB, McKinsey TA, NewkirkS, HuynhK, SucharovCC, BristowMR. Increased phosphorylation- dependent nuclear export of class II histone deacetylases in failing human heart Clin Transl Sci, 2009,2(5):325-332. [本文引用: 1]
CaoDS, WangZG, ZhangCL, OhJ, XingW, LiS, RichardsonJA, WangDZ, OlsonEN . Modulation of smooth muscle gene expression by association of histone acetyltransferases and deacetylases with myocardin Mol Cell Biol, 2005,25(1):364-376. [本文引用: 1]
McDonaldOG, WamhoffBR, HoofnagleMH, OwensGK . Control of SRF binding to CArG box chromatin regulates smooth muscle gene expression in vivo J Clin Invest, 2006,116(1):36-48. [本文引用: 1]
FischleW, DequiedtF, HendzelMJ, GuentherMG, LazarMA, VoelterW, VerdinE . Enzymatic activity associated with class II HDACs is dependent on a multiprotein complex containing HDAC3 and SMRT/N-CoR Mol Cell, 2002,9(1):45-57. [本文引用: 1]
KookH, LeporeJJ, GitlerAD, LuMM, Wing-Man Yung W, MackayJ, ZhouR, FerrariV, GruberP, EpsteinJA. Cardiac hypertrophy and histone deacetylase-dependent transcriptional repression mediated by the atypical homeodomain protein Hop J Clin Invest, 2003,112(6):863-871. [本文引用: 2]
KongYQ, TannousP, LuGR, BerenjiK, RothermelBA, OlsonEN, HillJA . Suppression of class I and II histone deacetylases blunts pressure-overload cardiac hypertrophy Circulation, 2006,113(22):2579-2588. [本文引用: 2]
IyerA, FenningA, LimJ, LeGT, ReidRC, HaliliMA, FairlieDP, BrownL . Antifibrotic activity of an inhibitor of histone deacetylases in DOCA-salt hypertensive rats Br J Pharmacol, 2010,159(7):1408-1417. [本文引用: 1]
KangSH, SeokYM, SongMJ, LeeHA, KurzT, KimI . Histone deacetylase inhibition attenuates cardiac hypertrophy and fibrosis through acetylation of mineralocorticoid receptor in spontaneously hypertensive rats Mol Pharmacol, 2015,87(5):782-791. [本文引用: 1]
ChoYK, EomGH, KeeHJ, KimHS, ChoiWY, NamKI, MaJS, KookH . Sodium valproate, a histone deacetylase inhibitor, but not captopril, prevents right ventricular hypertrophy in rats Circ J, 2010,74(4):760-770. [本文引用: 1]
GalloP, LatronicoMV, GalloP, GrimaldiS, BorgiaF, TodaroM, JonesP, GallinariP, De FrancescoR, CilibertoG, SteinkühlerC, EspositoG, CondorelliG . Inhibition of class I histone deacetylase with an apicidin derivative prevents cardiac hypertrophy and failure Cardiovasc Res, 2008,80(3):416-424. [本文引用: 1]
OoiJY, TuanoNK, RafehiH, GaoXM, ZiemannM, DuXJ, El-OstaA . HDAC inhibition attenuates cardiac hypertrophy by acetylation and deacetylation of target genes Epigenetics, 2015,10(5):418-430. [本文引用: 1]
CaoJH, LiaoWT, WoC, XuGR, XuHX, LiPL, TaoY, WangP, LinJR, DengLR . Transcriptome screening and verification of genes related to metabolism affected by histone deacetylase inhibitors Hereditas(Beijing), 2015,37(9):918-925. [本文引用: 1]
 ,湖南师范大学省部共建淡水鱼类发育生物学国家重点实验室,教育部重点实验室,生命科学学院心脏发育研究中心,长沙 410081
,湖南师范大学省部共建淡水鱼类发育生物学国家重点实验室,教育部重点实验室,生命科学学院心脏发育研究中心,长沙 410081
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT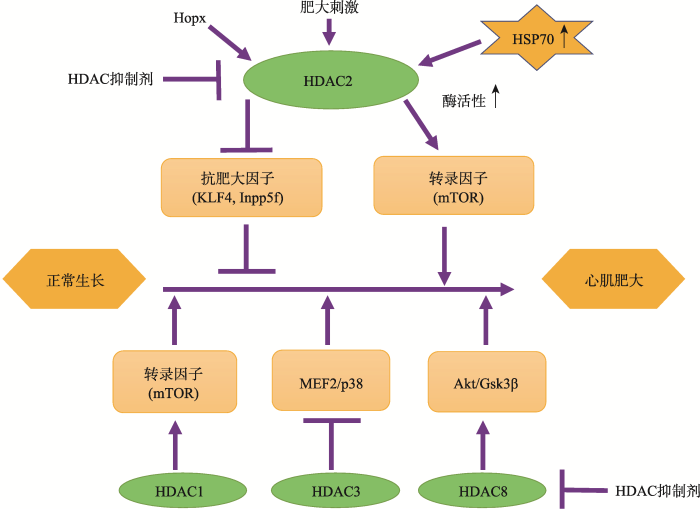 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}