Abstract The phytohormones are important small molecules synthesized in plants, and their contents change as the external and internal conditions vary. Small volume of growth media used in the hydroculture will restrict the growth of plants, which is generally thought to be caused by the shortage of nutrients. In this study, we found that rice seedlings grew faster in the large volume of water culture system than that in the small volume of water culture system. We measured the contents of different plant hormones in culture medium and rice seedlings using liquid chromatography-mass spectrum (LC-MS). It was found that there were more stress-response related phytohormones such as ABA, SA and JA-Ile accumulated in the plants growing in the small volume water system than that in the large volume water system, and finally resulted in a low growth rate and a biomass decline. Therefore, we speculate that plants can sense the water resource through actively sensing the concentration of root-secreted phytohormones, and adjust growth and development to better adapt to the environment. The study will help to understand the physiological functions of root-secreted phytohormones and optimize the hydroculture conditions in the area of plant factory. Keywords:culture volume;phytohormones;root exudate;growth and development;rice
PDF (1384KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 俞启璐, 赵江哲, 朱晓仙, 张可伟. 水稻根分泌激素调节生长速度. 植物学报, 2021, 56(2): 175-182 doi:10.11983/CBB20133 Yu Qilu, Zhao Jiangzhe, Zhu Xiaoxian, Zhang Kewei. Regulation of Rice Growth by Root-secreted Phytohormones. Bulletin of Botany, 2021, 56(2): 175-182 doi:10.11983/CBB20133
植物激素是植物体内合成的一类重要次生代谢物质, 广泛参与植物生长发育调控, 并且可使植物感受外界环境信号进而影响其抗病和抗逆反应。例如, 当受到干旱胁迫时, 植物根部合成的脱落酸(abscisic acid, ABA)通过木质部转运至地上组织, 调控气孔关闭并影响茎秆生长, 使地上组织的生长状态适应干旱环境(Takahashi et al.2018); 同时过量的ABA使植物根部积累大量的脯氨酸来提高渗透压进而增强根部吸水能力, 起到抵抗干旱的作用(Bhaskara et al.2012; Bandurska et al.2017)。酚类植物激素水杨酸(salicylic acid, SA)可以调节植物的生长发育, 并在应对生物和非生物胁迫过程中起重要作用(Vlot et al.2009)。茉莉酸(jasmonic acid, JA)则参与植物生长发育的调控以及逆境响应和防御反应(Valenzuela et al.2016), JA-Ile为其活性形式(Ruan et al.2019)。以上3种植物激素通常会协同作用并负调控植物的生长发育以提高植物体对逆境的适应性(Verma et al.2016)。而生长素(auxin)和细胞分裂素类(cytokinins, CKs)则是促进植物生长的重要物质(Luo et al.2018; Wybouw and De Rybel, 2019), 它们在植物体内与ABA、SA和JA等胁迫相关激素通过复杂的信号网络共同调控植物的生长发育和抗逆反应(Bielach et al.2017; Cortleven et al.2019; 代宇佳等, 2019)。
干旱是常见的非生物胁迫之一, 水分缺失使植物体内积累大量活性氧并导致光合作用减弱、生长发育受抑制(任伟等, 2020)。植物的生长发育和繁殖需要充足的水分, 但气候变化经常使植物面临缺水的威胁, 因此植物需要通过多种适应机制和响应途径对干旱胁迫做出应答以维持自身的生存和生长(Huang et al.2018)。通常认为, 植物通过提高自身的抗旱性、耐旱性、避旱性或旱后恢复能力来应对干旱(Fang and Xiong, 2015)。适应干旱的方法包括关闭气孔、增厚叶蜡和卷曲叶片等方式来减少水分散失, 并通过促进根部生长、增加根部细胞渗透压和增大根冠比等来增加水分吸收以保持自身的水分平衡(Mohammadian et al.2007; Seki et al.2007; Tardieu, 2013; Hu and Xiong, 2014)。同时, 植物还通过多种分子机制感知、传导外界干旱刺激并启动适应性反应机制。例如, 通过ABA等多种信号分子调节下游抗旱相关基因的表达和功能蛋白质的合成(Xiong et al.2002; Zou et al.2010)。但目前关于植物抗旱的研究通常集中于干旱条件下的植物抗旱应答, 而对于植物预判干旱并提前为此做出准备的研究还未见报道。
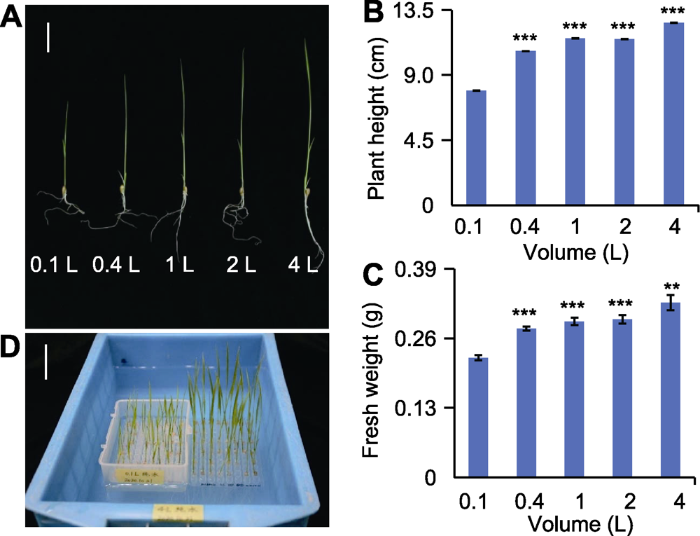
(A)不同体积纯水培养12天后水稻幼苗表型(bar=3 cm); (B) 不同体积纯水培养12天后水稻幼苗株高(n=30); (C)不同体积纯水培养12天后水稻幼苗鲜重(n=30); (D) 0.1 L与4 L培养容器及幼苗表型(bar=3 cm)。** 差异显著(P<0.01), *** 差异极显著(P<0.001)。 Figure 1The phenotype of 12-day rice seedlings grown in different water culture systems
(A) Phenotypes of rice seedlings grown in different water culture systems for 12 days (bar=3 cm); (B) Plant height of rice seedlings grown in different water culture systems for 12 days (n=30); (C) Fresh weight of rice seedlings grown in different water culture systems for 12 days (n=30); (D) 0.1 and 4 L containers used in this experiment and the phenotypes of rice seedlings grown in them, respectively (bar=3 cm). ** significant difference at P<0.01; *** significant differences at P<0.001.
(A)-(J)0.1和4 L纯水培养12天后水稻幼苗地上部组织ABA、SA、JA-Ile、IAA、tZ、tZR、cZ、cZR、iP和iPR的含量。* 差异显著(P<0.05); *** 差异极显著(P<0.001)。 Figure 2Hormone profiling of shoot of rice seedlings grown in different water culture sytems
(A)-(J) The ABA, SA, JA-Ile, IAA, tZ, tZR, cZ, cZR, iP, and iPR contents in shoot of rice seedlings after planting in 0.1 and 4 L water culture systems for 12 days, respectively. * significant differences at P<0.05; *** significant differences at P<0.001.
(A)-(J)0.1和4 L纯水培养12天后水稻幼苗根部ABA、SA、JA-Ile、IAA、tZ、tZR、cZ、cZR、iP和iPR的含量。* 差异显著(P<0.05); *** 差异极显著(P<0.001)。 Figure 3Profiling of phytohormones in the root of rice after planting in different water culture systems
(A)-(J)The ABA, SA, JA-Ile, IAA, tZ, tZR, cZ, cZR, iP, and iPR contents in the root of rice after planting in 0.1 and 4 L water culture systems, respectively. * significant differences at P<0.05; *** significant differences at P<0.001.
(A)-(H)0.1和4 L纯水培养水稻幼苗12天后水中ABA、SA、JA-Ile、IAA、cZ、cZR、iP和iPR的含量。* 差异显著(P<0.05); *** 差异极显著(P<0.001)。 Figure 4Phytohormone profiling of the different hydroculture systems
(A)-(H) The ABA, SA, JA-Ile, IAA, cZ, cZR, iP, and iPR contents in 0.1 and 4 L water culture systems after being planted with rice seedlings for 12 days, respectively. * significant differences at P<0.05; *** significant differences at P<0.001.
AlbrechtT, ArguesoCT (2017). Should I fight or should I grow now? The role of cytokinins in plant growth and immunity and in the growth-defence trade-off Ann Bot119,725-735. [本文引用: 1]
BandurskaH, NiedzielaJ, Pietrowska-BorekM, NucK, ChadzinikolauT, RadzikowskaD (2017). Regulation of proline biosynthesis and resistance to drought stress in two barley ( Hordeum vulgare L.) genotypes of different origin Plant Physiol Biochem118,427-437. DOI:10.1016/j.plaphy.2017.07.006URL [本文引用: 1]
BhaskaraGB, NguyenTT, VersluesPE (2012). Unique drought resistance functions of the highly ABA-induced clade A protein phosphatase 2Cs Plant Physiol160,379-395. [本文引用: 2]
BielachA, HrtyanM, TognettiVB (2017). Plants under stress: involvement of auxin and cytokinin Int J Mol Sci18,1427. DOI:10.3390/ijms18071427URL [本文引用: 1]
CortlevenA, LeuendorfJE, FrankM, PezzettaD, BoltS, SchmüllingT (2019). Cytokinin action in response to abiotic and biotic stresses in plants Plant Cell Environ42,998-1018. DOI:10.1111/pce.13494PMID:WOS:000459014800019 [本文引用: 1] The phytohormone cytokinin was originally discovered as a regulator of cell division. Later, it was described to be involved in regulating numerous processes in plant growth and development including meristem activity, tissue patterning, and organ size. More recently, diverse functions for cytokinin in the response to abiotic and biotic stresses have been reported. Cytokinin is required for the defence against high light stress and to protect plants from a novel type of abiotic stress caused by an altered photoperiod. Additionally, cytokinin has a role in the response to temperature, drought, osmotic, salt, and nutrient stress. Similarly, the full response to certain plant pathogens and herbivores requires a functional cytokinin signalling pathway. Conversely, different types of stress impact cytokinin homeostasis. The diverse functions of cytokinin in responses to stress and crosstalk with other hormones are described. Its emerging roles as a priming agent and as a regulator of growth-defence trade-offs are discussed.
FangYJ, XiongLZ (2015). General mechanisms of drought response and their application in drought resistance improvement in plants Cell Mol Life Sci72,673-689. DOI:10.1007/s00018-014-1767-0URL [本文引用: 2]
Gargallo-GarrigaA, PreeceC, SardansJ, OravecM, UrbanO, Pe?uelasJ (2018). Root exudate metabolomes change under drought and show limited capacity for recovery Sci Rep8,12696. DOI:10.1038/s41598-018-30150-0URL [本文引用: 3]
HuHH, XiongLZ (2014). Genetic engineering and breeding of drought-resistant crops Annu Rev Plant Biol65,715- 741. DOI:10.1146/annurev-arplant-050213-040000URL [本文引用: 1] Drought is one of the most important environmental stresses affecting the productivity of most field crops. Elucidation of the complex mechanisms underlying drought resistance in crops will accelerate the development of new varieties with enhanced drought resistance. Here, we provide a brief review on the progress in genetic, genomic, and molecular studies of drought resistance in major crops. Drought resistance is regulated by numerous small-effect loci and hundreds of genes that control various morphological and physiological responses to drought. This review focuses on recent studies of genes that have been well characterized as affecting drought resistance and genes that have been successfully engineered in staple crops. We propose that one significant challenge will be to unravel the complex mechanisms of drought resistance in crops through more intensive and integrative studies in order to find key functional components or machineries that can be used as tools for engineering and breeding drought-resistant crops.
HuangJL, ZhaiJQ, JiangT, WangYJ, LiXC, WangR, XiongM, SuBD, ThomasF (2018). Analysis of future drought characteristics in China using the regional climate model CCLM Climate Dyn50,507-525. DOI:10.1007/s00382-017-3623-zURL [本文引用: 1]
KramerPJ (1983). Water Relation of Plant. New York: Academic Press. pp.168-191. [本文引用: 1]
LuoJ, ZhouJJ, ZhangJZ (2018). Aux/ IAA gene family in plants: molecular structure, regulation, and function Int J Mol Sci19,259. DOI:10.3390/ijms19010259URL [本文引用: 1]
MohammadianMA, WatlingJR, HillRS (2007). The impact of epicuticular wax on gas-exchange and photoinhibition in Leucadendron lanigerum (Proteaceae) Acta Oecol31,93-101. [本文引用: 1]
NorthGB, NobelPS (1992). Drought-induced changes in hydraulic conductivity and structure in roots of Ferocactus acanthodes and Opuntia ficus-indica New Phytol120,9-19. [本文引用: 1]
ShinozakiK, Yamaguchi-ShinozakiK (1997). Gene expression and signal transduction in water-stress response Plant Physiol115,327-334. [本文引用: 1]
TakahashiF, KuromoriT, SatoH, ShinozakiK (2018). Regulatory gene networks in drought stress responses and resistance in plants Adv Exp Med Biol1081,189-214. [本文引用: 1]
TardieuF (2013). Plant response to environmental conditions: assessing potential production, water demand, and negative effects of water deficit Front Physiol4,17. [本文引用: 1]
ValenzuelaCE, Acevedo-AcevedoO, MirandaGS, Vergara-BarrosP, HoluigueL, FigueroaCR, FigueroaPM (2016). Salt stress response triggers activation of the jasmonate signaling pathway leading to inhibition of cell elongation in Arabidopsis primary root J Exp Bot67,4209- 4220. [本文引用: 1]
... 植物激素是植物体内合成的一类重要次生代谢物质, 广泛参与植物生长发育调控, 并且可使植物感受外界环境信号进而影响其抗病和抗逆反应.例如, 当受到干旱胁迫时, 植物根部合成的脱落酸(abscisic acid, ABA)通过木质部转运至地上组织, 调控气孔关闭并影响茎秆生长, 使地上组织的生长状态适应干旱环境(Takahashi et al.2018); 同时过量的ABA使植物根部积累大量的脯氨酸来提高渗透压进而增强根部吸水能力, 起到抵抗干旱的作用(Bhaskara et al.2012; Bandurska et al.2017).酚类植物激素水杨酸(salicylic acid, SA)可以调节植物的生长发育, 并在应对生物和非生物胁迫过程中起重要作用(Vlot et al.2009).茉莉酸(jasmonic acid, JA)则参与植物生长发育的调控以及逆境响应和防御反应(Valenzuela et al.2016), JA-Ile为其活性形式(Ruan et al.2019).以上3种植物激素通常会协同作用并负调控植物的生长发育以提高植物体对逆境的适应性(Verma et al.2016).而生长素(auxin)和细胞分裂素类(cytokinins, CKs)则是促进植物生长的重要物质(Luo et al.2018; Wybouw and De Rybel, 2019), 它们在植物体内与ABA、SA和JA等胁迫相关激素通过复杂的信号网络共同调控植物的生长发育和抗逆反应(Bielach et al.2017; Cortleven et al.2019; 代宇佳等, 2019). ...
... 干旱是常见的非生物胁迫之一, 水分缺失使植物体内积累大量活性氧并导致光合作用减弱、生长发育受抑制(任伟等, 2020).植物的生长发育和繁殖需要充足的水分, 但气候变化经常使植物面临缺水的威胁, 因此植物需要通过多种适应机制和响应途径对干旱胁迫做出应答以维持自身的生存和生长(Huang et al.2018).通常认为, 植物通过提高自身的抗旱性、耐旱性、避旱性或旱后恢复能力来应对干旱(Fang and Xiong, 2015).适应干旱的方法包括关闭气孔、增厚叶蜡和卷曲叶片等方式来减少水分散失, 并通过促进根部生长、增加根部细胞渗透压和增大根冠比等来增加水分吸收以保持自身的水分平衡(Mohammadian et al.2007; Seki et al.2007; Tardieu, 2013; Hu and Xiong, 2014).同时, 植物还通过多种分子机制感知、传导外界干旱刺激并启动适应性反应机制.例如, 通过ABA等多种信号分子调节下游抗旱相关基因的表达和功能蛋白质的合成(Xiong et al.2002; Zou et al.2010).但目前关于植物抗旱的研究通常集中于干旱条件下的植物抗旱应答, 而对于植物预判干旱并提前为此做出准备的研究还未见报道. ...
Regulation of proline biosynthesis and resistance to drought stress in two barley ( Hordeum vulgare L.) genotypes of different origin 1 2017
... 植物激素是植物体内合成的一类重要次生代谢物质, 广泛参与植物生长发育调控, 并且可使植物感受外界环境信号进而影响其抗病和抗逆反应.例如, 当受到干旱胁迫时, 植物根部合成的脱落酸(abscisic acid, ABA)通过木质部转运至地上组织, 调控气孔关闭并影响茎秆生长, 使地上组织的生长状态适应干旱环境(Takahashi et al.2018); 同时过量的ABA使植物根部积累大量的脯氨酸来提高渗透压进而增强根部吸水能力, 起到抵抗干旱的作用(Bhaskara et al.2012; Bandurska et al.2017).酚类植物激素水杨酸(salicylic acid, SA)可以调节植物的生长发育, 并在应对生物和非生物胁迫过程中起重要作用(Vlot et al.2009).茉莉酸(jasmonic acid, JA)则参与植物生长发育的调控以及逆境响应和防御反应(Valenzuela et al.2016), JA-Ile为其活性形式(Ruan et al.2019).以上3种植物激素通常会协同作用并负调控植物的生长发育以提高植物体对逆境的适应性(Verma et al.2016).而生长素(auxin)和细胞分裂素类(cytokinins, CKs)则是促进植物生长的重要物质(Luo et al.2018; Wybouw and De Rybel, 2019), 它们在植物体内与ABA、SA和JA等胁迫相关激素通过复杂的信号网络共同调控植物的生长发育和抗逆反应(Bielach et al.2017; Cortleven et al.2019; 代宇佳等, 2019). ...
Unique drought resistance functions of the highly ABA-induced clade A protein phosphatase 2Cs 2 2012
... 植物激素是植物体内合成的一类重要次生代谢物质, 广泛参与植物生长发育调控, 并且可使植物感受外界环境信号进而影响其抗病和抗逆反应.例如, 当受到干旱胁迫时, 植物根部合成的脱落酸(abscisic acid, ABA)通过木质部转运至地上组织, 调控气孔关闭并影响茎秆生长, 使地上组织的生长状态适应干旱环境(Takahashi et al.2018); 同时过量的ABA使植物根部积累大量的脯氨酸来提高渗透压进而增强根部吸水能力, 起到抵抗干旱的作用(Bhaskara et al.2012; Bandurska et al.2017).酚类植物激素水杨酸(salicylic acid, SA)可以调节植物的生长发育, 并在应对生物和非生物胁迫过程中起重要作用(Vlot et al.2009).茉莉酸(jasmonic acid, JA)则参与植物生长发育的调控以及逆境响应和防御反应(Valenzuela et al.2016), JA-Ile为其活性形式(Ruan et al.2019).以上3种植物激素通常会协同作用并负调控植物的生长发育以提高植物体对逆境的适应性(Verma et al.2016).而生长素(auxin)和细胞分裂素类(cytokinins, CKs)则是促进植物生长的重要物质(Luo et al.2018; Wybouw and De Rybel, 2019), 它们在植物体内与ABA、SA和JA等胁迫相关激素通过复杂的信号网络共同调控植物的生长发育和抗逆反应(Bielach et al.2017; Cortleven et al.2019; 代宇佳等, 2019). ...
Plants under stress: involvement of auxin and cytokinin 1 2017
... 植物激素是植物体内合成的一类重要次生代谢物质, 广泛参与植物生长发育调控, 并且可使植物感受外界环境信号进而影响其抗病和抗逆反应.例如, 当受到干旱胁迫时, 植物根部合成的脱落酸(abscisic acid, ABA)通过木质部转运至地上组织, 调控气孔关闭并影响茎秆生长, 使地上组织的生长状态适应干旱环境(Takahashi et al.2018); 同时过量的ABA使植物根部积累大量的脯氨酸来提高渗透压进而增强根部吸水能力, 起到抵抗干旱的作用(Bhaskara et al.2012; Bandurska et al.2017).酚类植物激素水杨酸(salicylic acid, SA)可以调节植物的生长发育, 并在应对生物和非生物胁迫过程中起重要作用(Vlot et al.2009).茉莉酸(jasmonic acid, JA)则参与植物生长发育的调控以及逆境响应和防御反应(Valenzuela et al.2016), JA-Ile为其活性形式(Ruan et al.2019).以上3种植物激素通常会协同作用并负调控植物的生长发育以提高植物体对逆境的适应性(Verma et al.2016).而生长素(auxin)和细胞分裂素类(cytokinins, CKs)则是促进植物生长的重要物质(Luo et al.2018; Wybouw and De Rybel, 2019), 它们在植物体内与ABA、SA和JA等胁迫相关激素通过复杂的信号网络共同调控植物的生长发育和抗逆反应(Bielach et al.2017; Cortleven et al.2019; 代宇佳等, 2019). ...
Cytokinin action in response to abiotic and biotic stresses in plants 1 2019
... 植物激素是植物体内合成的一类重要次生代谢物质, 广泛参与植物生长发育调控, 并且可使植物感受外界环境信号进而影响其抗病和抗逆反应.例如, 当受到干旱胁迫时, 植物根部合成的脱落酸(abscisic acid, ABA)通过木质部转运至地上组织, 调控气孔关闭并影响茎秆生长, 使地上组织的生长状态适应干旱环境(Takahashi et al.2018); 同时过量的ABA使植物根部积累大量的脯氨酸来提高渗透压进而增强根部吸水能力, 起到抵抗干旱的作用(Bhaskara et al.2012; Bandurska et al.2017).酚类植物激素水杨酸(salicylic acid, SA)可以调节植物的生长发育, 并在应对生物和非生物胁迫过程中起重要作用(Vlot et al.2009).茉莉酸(jasmonic acid, JA)则参与植物生长发育的调控以及逆境响应和防御反应(Valenzuela et al.2016), JA-Ile为其活性形式(Ruan et al.2019).以上3种植物激素通常会协同作用并负调控植物的生长发育以提高植物体对逆境的适应性(Verma et al.2016).而生长素(auxin)和细胞分裂素类(cytokinins, CKs)则是促进植物生长的重要物质(Luo et al.2018; Wybouw and De Rybel, 2019), 它们在植物体内与ABA、SA和JA等胁迫相关激素通过复杂的信号网络共同调控植物的生长发育和抗逆反应(Bielach et al.2017; Cortleven et al.2019; 代宇佳等, 2019). ...
General mechanisms of drought response and their application in drought resistance improvement in plants 2 2015
... 干旱是常见的非生物胁迫之一, 水分缺失使植物体内积累大量活性氧并导致光合作用减弱、生长发育受抑制(任伟等, 2020).植物的生长发育和繁殖需要充足的水分, 但气候变化经常使植物面临缺水的威胁, 因此植物需要通过多种适应机制和响应途径对干旱胁迫做出应答以维持自身的生存和生长(Huang et al.2018).通常认为, 植物通过提高自身的抗旱性、耐旱性、避旱性或旱后恢复能力来应对干旱(Fang and Xiong, 2015).适应干旱的方法包括关闭气孔、增厚叶蜡和卷曲叶片等方式来减少水分散失, 并通过促进根部生长、增加根部细胞渗透压和增大根冠比等来增加水分吸收以保持自身的水分平衡(Mohammadian et al.2007; Seki et al.2007; Tardieu, 2013; Hu and Xiong, 2014).同时, 植物还通过多种分子机制感知、传导外界干旱刺激并启动适应性反应机制.例如, 通过ABA等多种信号分子调节下游抗旱相关基因的表达和功能蛋白质的合成(Xiong et al.2002; Zou et al.2010).但目前关于植物抗旱的研究通常集中于干旱条件下的植物抗旱应答, 而对于植物预判干旱并提前为此做出准备的研究还未见报道. ...
... 植物在生长过程中通过根部向生长介质中释放多种无机离子和有机小分子物质, 以改变植物根系周围物理、化学或生物条件进而利于自身生长(Gargallo- Garriga et al.2018).关于根系分泌物的研究通常集中在其对营养物质吸收的竞争与调节、促进或抑制根系菌群生长以及根系间交互作用等方面(洪常青和聂艳丽, 2003), 较少关注根部向外分泌激素对植物生长的影响.Vives-Peris等(2017)发现根部分泌物中包括SA、JA和ABA等多种植物激素在内的次生代谢物在植物耐盐和耐热中发挥重要作用.本研究表明植物亦可通过感知根际激素的浓度来预判外界供水量是否能满足其长期生长的需要, 进而调节自身的生长发育和提高抗逆性以适应生存环境. ...
Genetic engineering and breeding of drought-resistant crops 1 2014
... 干旱是常见的非生物胁迫之一, 水分缺失使植物体内积累大量活性氧并导致光合作用减弱、生长发育受抑制(任伟等, 2020).植物的生长发育和繁殖需要充足的水分, 但气候变化经常使植物面临缺水的威胁, 因此植物需要通过多种适应机制和响应途径对干旱胁迫做出应答以维持自身的生存和生长(Huang et al.2018).通常认为, 植物通过提高自身的抗旱性、耐旱性、避旱性或旱后恢复能力来应对干旱(Fang and Xiong, 2015).适应干旱的方法包括关闭气孔、增厚叶蜡和卷曲叶片等方式来减少水分散失, 并通过促进根部生长、增加根部细胞渗透压和增大根冠比等来增加水分吸收以保持自身的水分平衡(Mohammadian et al.2007; Seki et al.2007; Tardieu, 2013; Hu and Xiong, 2014).同时, 植物还通过多种分子机制感知、传导外界干旱刺激并启动适应性反应机制.例如, 通过ABA等多种信号分子调节下游抗旱相关基因的表达和功能蛋白质的合成(Xiong et al.2002; Zou et al.2010).但目前关于植物抗旱的研究通常集中于干旱条件下的植物抗旱应答, 而对于植物预判干旱并提前为此做出准备的研究还未见报道. ...
Analysis of future drought characteristics in China using the regional climate model CCLM 1 2018
... 干旱是常见的非生物胁迫之一, 水分缺失使植物体内积累大量活性氧并导致光合作用减弱、生长发育受抑制(任伟等, 2020).植物的生长发育和繁殖需要充足的水分, 但气候变化经常使植物面临缺水的威胁, 因此植物需要通过多种适应机制和响应途径对干旱胁迫做出应答以维持自身的生存和生长(Huang et al.2018).通常认为, 植物通过提高自身的抗旱性、耐旱性、避旱性或旱后恢复能力来应对干旱(Fang and Xiong, 2015).适应干旱的方法包括关闭气孔、增厚叶蜡和卷曲叶片等方式来减少水分散失, 并通过促进根部生长、增加根部细胞渗透压和增大根冠比等来增加水分吸收以保持自身的水分平衡(Mohammadian et al.2007; Seki et al.2007; Tardieu, 2013; Hu and Xiong, 2014).同时, 植物还通过多种分子机制感知、传导外界干旱刺激并启动适应性反应机制.例如, 通过ABA等多种信号分子调节下游抗旱相关基因的表达和功能蛋白质的合成(Xiong et al.2002; Zou et al.2010).但目前关于植物抗旱的研究通常集中于干旱条件下的植物抗旱应答, 而对于植物预判干旱并提前为此做出准备的研究还未见报道. ...
Aux/ IAA gene family in plants: molecular structure, regulation, and function 1 2018
... 植物激素是植物体内合成的一类重要次生代谢物质, 广泛参与植物生长发育调控, 并且可使植物感受外界环境信号进而影响其抗病和抗逆反应.例如, 当受到干旱胁迫时, 植物根部合成的脱落酸(abscisic acid, ABA)通过木质部转运至地上组织, 调控气孔关闭并影响茎秆生长, 使地上组织的生长状态适应干旱环境(Takahashi et al.2018); 同时过量的ABA使植物根部积累大量的脯氨酸来提高渗透压进而增强根部吸水能力, 起到抵抗干旱的作用(Bhaskara et al.2012; Bandurska et al.2017).酚类植物激素水杨酸(salicylic acid, SA)可以调节植物的生长发育, 并在应对生物和非生物胁迫过程中起重要作用(Vlot et al.2009).茉莉酸(jasmonic acid, JA)则参与植物生长发育的调控以及逆境响应和防御反应(Valenzuela et al.2016), JA-Ile为其活性形式(Ruan et al.2019).以上3种植物激素通常会协同作用并负调控植物的生长发育以提高植物体对逆境的适应性(Verma et al.2016).而生长素(auxin)和细胞分裂素类(cytokinins, CKs)则是促进植物生长的重要物质(Luo et al.2018; Wybouw and De Rybel, 2019), 它们在植物体内与ABA、SA和JA等胁迫相关激素通过复杂的信号网络共同调控植物的生长发育和抗逆反应(Bielach et al.2017; Cortleven et al.2019; 代宇佳等, 2019). ...
The impact of epicuticular wax on gas-exchange and photoinhibition in Leucadendron lanigerum (Proteaceae) 1 2007
... 干旱是常见的非生物胁迫之一, 水分缺失使植物体内积累大量活性氧并导致光合作用减弱、生长发育受抑制(任伟等, 2020).植物的生长发育和繁殖需要充足的水分, 但气候变化经常使植物面临缺水的威胁, 因此植物需要通过多种适应机制和响应途径对干旱胁迫做出应答以维持自身的生存和生长(Huang et al.2018).通常认为, 植物通过提高自身的抗旱性、耐旱性、避旱性或旱后恢复能力来应对干旱(Fang and Xiong, 2015).适应干旱的方法包括关闭气孔、增厚叶蜡和卷曲叶片等方式来减少水分散失, 并通过促进根部生长、增加根部细胞渗透压和增大根冠比等来增加水分吸收以保持自身的水分平衡(Mohammadian et al.2007; Seki et al.2007; Tardieu, 2013; Hu and Xiong, 2014).同时, 植物还通过多种分子机制感知、传导外界干旱刺激并启动适应性反应机制.例如, 通过ABA等多种信号分子调节下游抗旱相关基因的表达和功能蛋白质的合成(Xiong et al.2002; Zou et al.2010).但目前关于植物抗旱的研究通常集中于干旱条件下的植物抗旱应答, 而对于植物预判干旱并提前为此做出准备的研究还未见报道. ...
Drought-induced changes in hydraulic conductivity and structure in roots of Ferocactus acanthodes and Opuntia ficus-indica 1 1992
... 植物激素是植物体内合成的一类重要次生代谢物质, 广泛参与植物生长发育调控, 并且可使植物感受外界环境信号进而影响其抗病和抗逆反应.例如, 当受到干旱胁迫时, 植物根部合成的脱落酸(abscisic acid, ABA)通过木质部转运至地上组织, 调控气孔关闭并影响茎秆生长, 使地上组织的生长状态适应干旱环境(Takahashi et al.2018); 同时过量的ABA使植物根部积累大量的脯氨酸来提高渗透压进而增强根部吸水能力, 起到抵抗干旱的作用(Bhaskara et al.2012; Bandurska et al.2017).酚类植物激素水杨酸(salicylic acid, SA)可以调节植物的生长发育, 并在应对生物和非生物胁迫过程中起重要作用(Vlot et al.2009).茉莉酸(jasmonic acid, JA)则参与植物生长发育的调控以及逆境响应和防御反应(Valenzuela et al.2016), JA-Ile为其活性形式(Ruan et al.2019).以上3种植物激素通常会协同作用并负调控植物的生长发育以提高植物体对逆境的适应性(Verma et al.2016).而生长素(auxin)和细胞分裂素类(cytokinins, CKs)则是促进植物生长的重要物质(Luo et al.2018; Wybouw and De Rybel, 2019), 它们在植物体内与ABA、SA和JA等胁迫相关激素通过复杂的信号网络共同调控植物的生长发育和抗逆反应(Bielach et al.2017; Cortleven et al.2019; 代宇佳等, 2019). ...
Regulatory metabolic networks in drought stress responses 1 2007
... 干旱是常见的非生物胁迫之一, 水分缺失使植物体内积累大量活性氧并导致光合作用减弱、生长发育受抑制(任伟等, 2020).植物的生长发育和繁殖需要充足的水分, 但气候变化经常使植物面临缺水的威胁, 因此植物需要通过多种适应机制和响应途径对干旱胁迫做出应答以维持自身的生存和生长(Huang et al.2018).通常认为, 植物通过提高自身的抗旱性、耐旱性、避旱性或旱后恢复能力来应对干旱(Fang and Xiong, 2015).适应干旱的方法包括关闭气孔、增厚叶蜡和卷曲叶片等方式来减少水分散失, 并通过促进根部生长、增加根部细胞渗透压和增大根冠比等来增加水分吸收以保持自身的水分平衡(Mohammadian et al.2007; Seki et al.2007; Tardieu, 2013; Hu and Xiong, 2014).同时, 植物还通过多种分子机制感知、传导外界干旱刺激并启动适应性反应机制.例如, 通过ABA等多种信号分子调节下游抗旱相关基因的表达和功能蛋白质的合成(Xiong et al.2002; Zou et al.2010).但目前关于植物抗旱的研究通常集中于干旱条件下的植物抗旱应答, 而对于植物预判干旱并提前为此做出准备的研究还未见报道. ...
Gene expression and signal transduction in water-stress response 1 1997
Regulatory gene networks in drought stress responses and resistance in plants 1 2018
... 植物激素是植物体内合成的一类重要次生代谢物质, 广泛参与植物生长发育调控, 并且可使植物感受外界环境信号进而影响其抗病和抗逆反应.例如, 当受到干旱胁迫时, 植物根部合成的脱落酸(abscisic acid, ABA)通过木质部转运至地上组织, 调控气孔关闭并影响茎秆生长, 使地上组织的生长状态适应干旱环境(Takahashi et al.2018); 同时过量的ABA使植物根部积累大量的脯氨酸来提高渗透压进而增强根部吸水能力, 起到抵抗干旱的作用(Bhaskara et al.2012; Bandurska et al.2017).酚类植物激素水杨酸(salicylic acid, SA)可以调节植物的生长发育, 并在应对生物和非生物胁迫过程中起重要作用(Vlot et al.2009).茉莉酸(jasmonic acid, JA)则参与植物生长发育的调控以及逆境响应和防御反应(Valenzuela et al.2016), JA-Ile为其活性形式(Ruan et al.2019).以上3种植物激素通常会协同作用并负调控植物的生长发育以提高植物体对逆境的适应性(Verma et al.2016).而生长素(auxin)和细胞分裂素类(cytokinins, CKs)则是促进植物生长的重要物质(Luo et al.2018; Wybouw and De Rybel, 2019), 它们在植物体内与ABA、SA和JA等胁迫相关激素通过复杂的信号网络共同调控植物的生长发育和抗逆反应(Bielach et al.2017; Cortleven et al.2019; 代宇佳等, 2019). ...
Plant response to environmental conditions: assessing potential production, water demand, and negative effects of water deficit 1 2013
... 干旱是常见的非生物胁迫之一, 水分缺失使植物体内积累大量活性氧并导致光合作用减弱、生长发育受抑制(任伟等, 2020).植物的生长发育和繁殖需要充足的水分, 但气候变化经常使植物面临缺水的威胁, 因此植物需要通过多种适应机制和响应途径对干旱胁迫做出应答以维持自身的生存和生长(Huang et al.2018).通常认为, 植物通过提高自身的抗旱性、耐旱性、避旱性或旱后恢复能力来应对干旱(Fang and Xiong, 2015).适应干旱的方法包括关闭气孔、增厚叶蜡和卷曲叶片等方式来减少水分散失, 并通过促进根部生长、增加根部细胞渗透压和增大根冠比等来增加水分吸收以保持自身的水分平衡(Mohammadian et al.2007; Seki et al.2007; Tardieu, 2013; Hu and Xiong, 2014).同时, 植物还通过多种分子机制感知、传导外界干旱刺激并启动适应性反应机制.例如, 通过ABA等多种信号分子调节下游抗旱相关基因的表达和功能蛋白质的合成(Xiong et al.2002; Zou et al.2010).但目前关于植物抗旱的研究通常集中于干旱条件下的植物抗旱应答, 而对于植物预判干旱并提前为此做出准备的研究还未见报道. ...
Salt stress response triggers activation of the jasmonate signaling pathway leading to inhibition of cell elongation in Arabidopsis primary root 1 2016
... 植物激素是植物体内合成的一类重要次生代谢物质, 广泛参与植物生长发育调控, 并且可使植物感受外界环境信号进而影响其抗病和抗逆反应.例如, 当受到干旱胁迫时, 植物根部合成的脱落酸(abscisic acid, ABA)通过木质部转运至地上组织, 调控气孔关闭并影响茎秆生长, 使地上组织的生长状态适应干旱环境(Takahashi et al.2018); 同时过量的ABA使植物根部积累大量的脯氨酸来提高渗透压进而增强根部吸水能力, 起到抵抗干旱的作用(Bhaskara et al.2012; Bandurska et al.2017).酚类植物激素水杨酸(salicylic acid, SA)可以调节植物的生长发育, 并在应对生物和非生物胁迫过程中起重要作用(Vlot et al.2009).茉莉酸(jasmonic acid, JA)则参与植物生长发育的调控以及逆境响应和防御反应(Valenzuela et al.2016), JA-Ile为其活性形式(Ruan et al.2019).以上3种植物激素通常会协同作用并负调控植物的生长发育以提高植物体对逆境的适应性(Verma et al.2016).而生长素(auxin)和细胞分裂素类(cytokinins, CKs)则是促进植物生长的重要物质(Luo et al.2018; Wybouw and De Rybel, 2019), 它们在植物体内与ABA、SA和JA等胁迫相关激素通过复杂的信号网络共同调控植物的生长发育和抗逆反应(Bielach et al.2017; Cortleven et al.2019; 代宇佳等, 2019). ...
Plant hormone-mediated regulation of stress responses 1 2016
... 植物激素是植物体内合成的一类重要次生代谢物质, 广泛参与植物生长发育调控, 并且可使植物感受外界环境信号进而影响其抗病和抗逆反应.例如, 当受到干旱胁迫时, 植物根部合成的脱落酸(abscisic acid, ABA)通过木质部转运至地上组织, 调控气孔关闭并影响茎秆生长, 使地上组织的生长状态适应干旱环境(Takahashi et al.2018); 同时过量的ABA使植物根部积累大量的脯氨酸来提高渗透压进而增强根部吸水能力, 起到抵抗干旱的作用(Bhaskara et al.2012; Bandurska et al.2017).酚类植物激素水杨酸(salicylic acid, SA)可以调节植物的生长发育, 并在应对生物和非生物胁迫过程中起重要作用(Vlot et al.2009).茉莉酸(jasmonic acid, JA)则参与植物生长发育的调控以及逆境响应和防御反应(Valenzuela et al.2016), JA-Ile为其活性形式(Ruan et al.2019).以上3种植物激素通常会协同作用并负调控植物的生长发育以提高植物体对逆境的适应性(Verma et al.2016).而生长素(auxin)和细胞分裂素类(cytokinins, CKs)则是促进植物生长的重要物质(Luo et al.2018; Wybouw and De Rybel, 2019), 它们在植物体内与ABA、SA和JA等胁迫相关激素通过复杂的信号网络共同调控植物的生长发育和抗逆反应(Bielach et al.2017; Cortleven et al.2019; 代宇佳等, 2019). ...
Citrus plants exude proline and phytohormones under abiotic stress conditions 2 2017
... 植物在生长过程中通过根部向生长介质中释放多种无机离子和有机小分子物质, 以改变植物根系周围物理、化学或生物条件进而利于自身生长(Gargallo- Garriga et al.2018).关于根系分泌物的研究通常集中在其对营养物质吸收的竞争与调节、促进或抑制根系菌群生长以及根系间交互作用等方面(洪常青和聂艳丽, 2003), 较少关注根部向外分泌激素对植物生长的影响.Vives-Peris等(2017)发现根部分泌物中包括SA、JA和ABA等多种植物激素在内的次生代谢物在植物耐盐和耐热中发挥重要作用.本研究表明植物亦可通过感知根际激素的浓度来预判外界供水量是否能满足其长期生长的需要, 进而调节自身的生长发育和提高抗逆性以适应生存环境. ...
Salicylic acid, a multifaceted hormone to combat disease 1 2009
... 植物激素是植物体内合成的一类重要次生代谢物质, 广泛参与植物生长发育调控, 并且可使植物感受外界环境信号进而影响其抗病和抗逆反应.例如, 当受到干旱胁迫时, 植物根部合成的脱落酸(abscisic acid, ABA)通过木质部转运至地上组织, 调控气孔关闭并影响茎秆生长, 使地上组织的生长状态适应干旱环境(Takahashi et al.2018); 同时过量的ABA使植物根部积累大量的脯氨酸来提高渗透压进而增强根部吸水能力, 起到抵抗干旱的作用(Bhaskara et al.2012; Bandurska et al.2017).酚类植物激素水杨酸(salicylic acid, SA)可以调节植物的生长发育, 并在应对生物和非生物胁迫过程中起重要作用(Vlot et al.2009).茉莉酸(jasmonic acid, JA)则参与植物生长发育的调控以及逆境响应和防御反应(Valenzuela et al.2016), JA-Ile为其活性形式(Ruan et al.2019).以上3种植物激素通常会协同作用并负调控植物的生长发育以提高植物体对逆境的适应性(Verma et al.2016).而生长素(auxin)和细胞分裂素类(cytokinins, CKs)则是促进植物生长的重要物质(Luo et al.2018; Wybouw and De Rybel, 2019), 它们在植物体内与ABA、SA和JA等胁迫相关激素通过复杂的信号网络共同调控植物的生长发育和抗逆反应(Bielach et al.2017; Cortleven et al.2019; 代宇佳等, 2019). ...
Cytokinin—a developing story 1 2019
... 植物激素是植物体内合成的一类重要次生代谢物质, 广泛参与植物生长发育调控, 并且可使植物感受外界环境信号进而影响其抗病和抗逆反应.例如, 当受到干旱胁迫时, 植物根部合成的脱落酸(abscisic acid, ABA)通过木质部转运至地上组织, 调控气孔关闭并影响茎秆生长, 使地上组织的生长状态适应干旱环境(Takahashi et al.2018); 同时过量的ABA使植物根部积累大量的脯氨酸来提高渗透压进而增强根部吸水能力, 起到抵抗干旱的作用(Bhaskara et al.2012; Bandurska et al.2017).酚类植物激素水杨酸(salicylic acid, SA)可以调节植物的生长发育, 并在应对生物和非生物胁迫过程中起重要作用(Vlot et al.2009).茉莉酸(jasmonic acid, JA)则参与植物生长发育的调控以及逆境响应和防御反应(Valenzuela et al.2016), JA-Ile为其活性形式(Ruan et al.2019).以上3种植物激素通常会协同作用并负调控植物的生长发育以提高植物体对逆境的适应性(Verma et al.2016).而生长素(auxin)和细胞分裂素类(cytokinins, CKs)则是促进植物生长的重要物质(Luo et al.2018; Wybouw and De Rybel, 2019), 它们在植物体内与ABA、SA和JA等胁迫相关激素通过复杂的信号网络共同调控植物的生长发育和抗逆反应(Bielach et al.2017; Cortleven et al.2019; 代宇佳等, 2019). ...
Cell signaling during cold, drought, and salt stress 1 2002
... 干旱是常见的非生物胁迫之一, 水分缺失使植物体内积累大量活性氧并导致光合作用减弱、生长发育受抑制(任伟等, 2020).植物的生长发育和繁殖需要充足的水分, 但气候变化经常使植物面临缺水的威胁, 因此植物需要通过多种适应机制和响应途径对干旱胁迫做出应答以维持自身的生存和生长(Huang et al.2018).通常认为, 植物通过提高自身的抗旱性、耐旱性、避旱性或旱后恢复能力来应对干旱(Fang and Xiong, 2015).适应干旱的方法包括关闭气孔、增厚叶蜡和卷曲叶片等方式来减少水分散失, 并通过促进根部生长、增加根部细胞渗透压和增大根冠比等来增加水分吸收以保持自身的水分平衡(Mohammadian et al.2007; Seki et al.2007; Tardieu, 2013; Hu and Xiong, 2014).同时, 植物还通过多种分子机制感知、传导外界干旱刺激并启动适应性反应机制.例如, 通过ABA等多种信号分子调节下游抗旱相关基因的表达和功能蛋白质的合成(Xiong et al.2002; Zou et al.2010).但目前关于植物抗旱的研究通常集中于干旱条件下的植物抗旱应答, 而对于植物预判干旱并提前为此做出准备的研究还未见报道. ...
ABC transporter OsABCG18 controls the shootward transport of cytokinins and grain yield in rice 1 2019
... 植物激素含量测定参照Zhao等(2019)的方法. ...
Arabidopsis calcium-dependent protein kinase CPK10 functions in abscisic acid- and Ca 2+-mediated stomatal regulation in response to drought stress 1 2010
... 干旱是常见的非生物胁迫之一, 水分缺失使植物体内积累大量活性氧并导致光合作用减弱、生长发育受抑制(任伟等, 2020).植物的生长发育和繁殖需要充足的水分, 但气候变化经常使植物面临缺水的威胁, 因此植物需要通过多种适应机制和响应途径对干旱胁迫做出应答以维持自身的生存和生长(Huang et al.2018).通常认为, 植物通过提高自身的抗旱性、耐旱性、避旱性或旱后恢复能力来应对干旱(Fang and Xiong, 2015).适应干旱的方法包括关闭气孔、增厚叶蜡和卷曲叶片等方式来减少水分散失, 并通过促进根部生长、增加根部细胞渗透压和增大根冠比等来增加水分吸收以保持自身的水分平衡(Mohammadian et al.2007; Seki et al.2007; Tardieu, 2013; Hu and Xiong, 2014).同时, 植物还通过多种分子机制感知、传导外界干旱刺激并启动适应性反应机制.例如, 通过ABA等多种信号分子调节下游抗旱相关基因的表达和功能蛋白质的合成(Xiong et al.2002; Zou et al.2010).但目前关于植物抗旱的研究通常集中于干旱条件下的植物抗旱应答, 而对于植物预判干旱并提前为此做出准备的研究还未见报道. ...
 ,*
,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT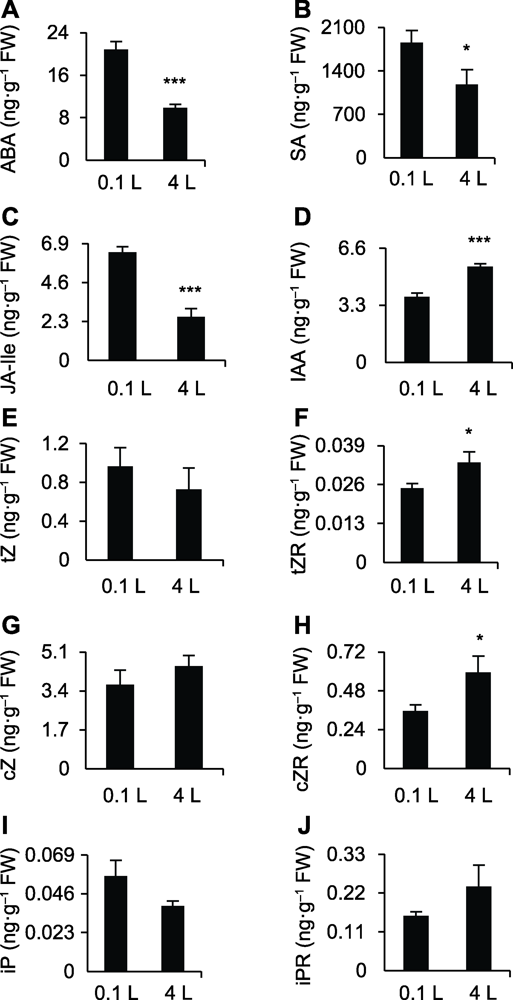 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT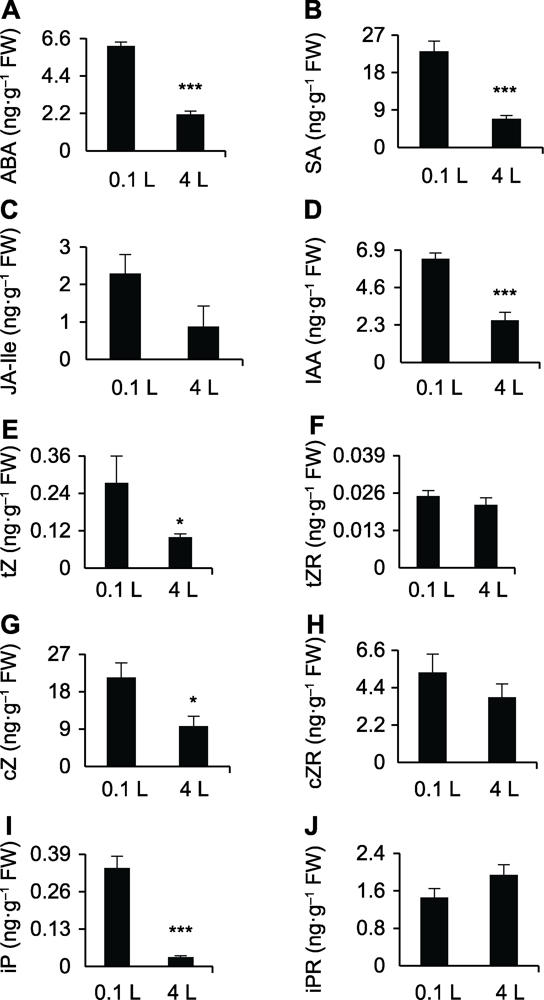 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT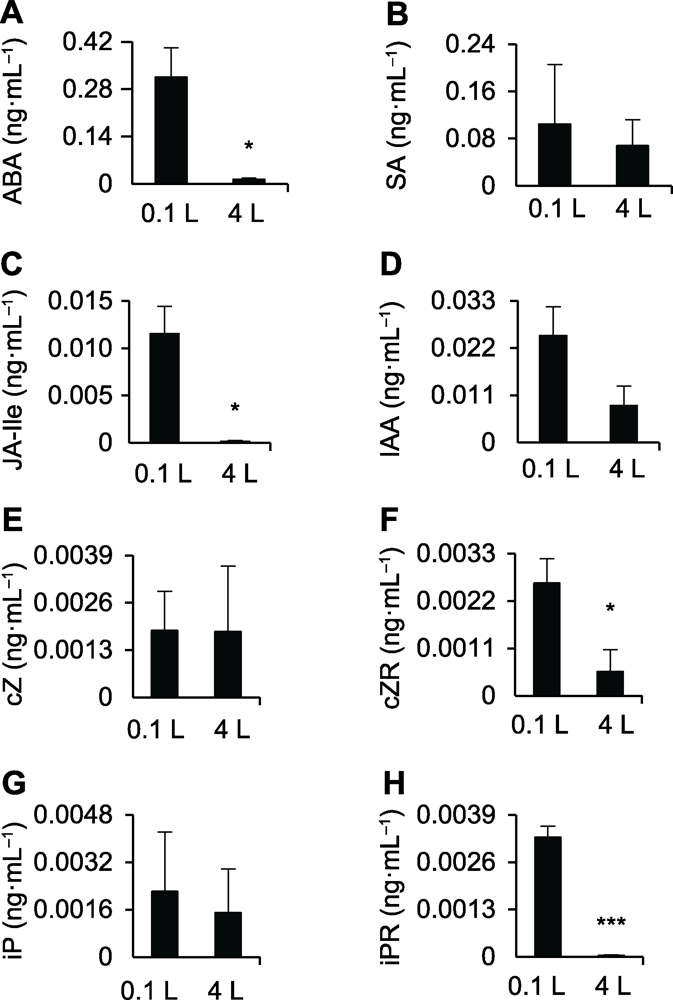 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}