Tairan Zhang1, Hechen Zhang2, Ronghua Wu,1,*1College of Forestry of Henan Agriculture University, Zhengzhou 450002, China 2Horticulture Research Institute, Henan Academy of Agricultural Sciences, Zhengzhou 450002, China
Abstract Flower color is an important feature of ornamental plants. There are only a few plants containing blue flowers in nature; however, most of the ornamental plants lack blue varieties. Therefore, studying the molecular mechanism underlying blue flower formation has great significance for flower breeding in the future. Recent studies have revealed that flower pigmentation is mainly caused by the accumulation of anthocyanins. Anthocyanidins are catalyzed to the stable anthocyanin form by glycosylation, acylation or methyl modification. Furthermore, the pH environment and metal ions in the vacuole also influence the pigmentation. In this review paper, we summarized the progresses in the anthocyanin biosynthesis and motablism pathways, the function of some key regulatory genes, the effect of anthocyanin acylation, the regulation by metal ion chelation and pH environment, and molecular breeding of blue flowers. Keywords:acylation;anthocyanin biosynthetic pathway;vacuolar pH;metal ion;blue flower;genetic engineering
PDF (2190KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 张泰然, 张和臣, 武荣花. 蓝色花形成分子机理研究进展. 植物学报, 2020, 55(2): 216-227 doi:10.11983/CBB19028 Zhang Tairan, Zhang Hechen, Wu Ronghua. Recent Advances on Blue Flower Formation. Chinese Bulletin of Botany, 2020, 55(2): 216-227 doi:10.11983/CBB19028
花色是观赏植物的一个重要表现特征(Tanaka et al., 2008), 蓝色花在花卉产业中需求旺盛。然而自然界中具有蓝色花的植物数量却相对较少。以云南省为例, 全省被子植物共17 000种, 其中蓝色花植物450种, 只占全省植物资源的2.7% (杨少勇和樊国盛, 2002)。切花中, 月季(Rosa chinensis)和康乃馨(Dianthus caryophyllus)尚未拥有真正意义上的蓝色花; 通过转基因手段只获得了紫色或蓝紫色种质(Tanaka et al., 1998, 2009; Sasaki and Nakayama, 2014)。菊花(Dendranthema morifolium)通过转基因技术于2017年获得真正意义上的蓝色花(Noda et al., 2017)。因此, 研究蓝色花形成的分子机理对于花卉新品种培育具有十分重要的意义。本文从花青苷合成与代谢途径、酰化对花青苷呈色及稳定的影响、金属离子对花青苷的作用、液泡pH值相关基因以及蓝色花分子育种等方面进行综述, 以期为蓝色花种质创建提供借鉴。
DareAP, TomesS, JonesM, McGhieTK, StevensonDE, JohnsonRA, GreenwoodDR, HellensRP ( 2013). Phenotypic changes associated with RNA interference silencing of chalcone synthase in apple ( Malus × domestica) Plant J 74, 398-410. [本文引用: 1]
de VlamingP, SchramAW, WieringH ( 1983). Genes affecting flower colour and pH of flower limb homogenates in Petunia hybrida Theor Appl Genet 66, 271-278. [本文引用: 1]
DehghanS, SadeghiM, P?ppelA, FischerR, Lakes- HarlanR, KavousiHR, VilcinskasA, RahnamaeianM ( 2014). Differential inductions of phenylalanine ammonia-lyase and chalcone synthase during wounding, salicylic acid treatment, and salinity stress in safflower, Carthamus tinctorius Biosci Rep 34, e00114. [本文引用: 1]
FaracoM, LiYB, LiSJ, SpeltC, Di SansebastianoGP, RealeL, FerrantiF, VerweijW, KoesR, QuattrocchioFM ( 2017). A tonoplast P3B-ATPase mediates fusion of two types of vacuoles in petal cells Cell Rep 19, 2413-2422. [本文引用: 1]
FaracoM, SpeltC, BliekM, VerweijW, HoshinoA, EspenL, PrinsiB, JaarsmaR, TarhanE, de BoerAH, Di SansebastianoGP, KoesR, QuattrocchioFM ( 2014). Hyperacidification of vacuoles by the combined action of two different P-ATPases in the tonoplast determines flower color Cell Rep 6, 32-43. [本文引用: 1]
FedenkoVS, ShemetSA, LandiM ( 2017). UV-vis spectroscopy and colorimetric models for detecting anthocyanin-metal complexes in plants: an overview of in vitro and in vivo techniques J Plant Physiol 212, 13-28. [本文引用: 1]
Fukada-TanakaS, InagakiY, YamaguchiT, SaitoN, IidaS ( 2000). Colour-enhancing protein in blue petals Nature 407, 581. [本文引用: 2]
GotoT, KondoT ( 1991). Structure and molecular stacking of anthocyanins—flower color variation Angew Chem Int Ed 30, 17-33. [本文引用: 1]
HoltonTA ( 2000). Transgenic plants exhibiting altered flower color and methods for producing same US patent, 6080920. 2000-06-27. [本文引用: 1]
HondoT, YoshidaK, NakagawaA, KawaiT, TamuraH, GotoT ( 1992). Structural basis of blue-colour development in flower petals from Commelina communis Nature 358, 515-518. [本文引用: 1]
HuangH, HuK, HanKT, XiangQY, DaiSL ( 2013). Flower colour modification of chrysanthemum by suppression of F3'H and overexpression of the exogenous Senecio cruentus F3'5'H gene PLoS One 8, e74395. [本文引用: 1]
IshiiI, SakaguchiK, FujitaK, OzekiY, MiyaharaT ( 2017). A double knockout mutant of acyl-glucose-dependent anthocyanin glucosyltransferase genes in Delphinium grandiflorum J Plant Physiol 216, 74-78. [本文引用: 1]
KondoT, UedaM, TamuraH, YoshidaK, IsobeM, GotoT ( 1994). Composition of protocyanin, a self-assembled supramolecular pigment from the blue cornflower, Centaurea cyanus Angew Chem Int Ed 33, 978-979. [本文引用: 1]
KühlbrandtW ( 2004). Biology, structure and mechanism of P-type ATPases Nat Rev Mol Cell Biol 5, 282-295. [本文引用: 1]
MarkhamKR, MitchellKA, BoaseMR ( 1997). Malvidin- 3- O-glucoside-5-O-(6-acetylglucoside) and its colour manifestation in ‘Johnson’s Blue’ and other ‘Blue’ geraniums Phytochemistry 45, 417-423. [本文引用: 1]
MartensS, TeeriT, ForkmannG ( 2002). Heterologous expression of dihydroflavonol 4-reductases from various plants FEBS Lett 531, 453-458. [本文引用: 1]
MatsufujiH, KidoH, MisawaH, YaguchiJ, OtsukiT, ChinoM, TakedaM, YamagataK ( 2007). Stability to light, heat, and hydrogen peroxide at different pH values and DPPH radical scavenging activity of acylated anthocyanins from red radish extract J Agric Food Chem 55, 3692-3701. [本文引用: 1]
MeyerP, HeidmannI, ForkmannG, SaedlerH ( 1987). A new petunia flower colour generated by transformation of a mutant with a maize gene Nature 330, 677-678. [本文引用: 1]
MiyaharaT, SakiyamaR, OzekiY, SasakiN ( 2013). Acyl-glucose-dependent glucosyltransferase catalyzes the final step of anthocyanin formation in Arabidopsis J Plant Physiol 170, 619-624. [本文引用: 1]
MoriM, KondoT, YoshidaK ( 2008). Cyanosalvianin, a supramolecular blue metalloanthocyanin, from petals of Salvia uliginosa Phytochemistry 69, 3151-3158. [本文引用: 1]
NakatsukaT, MishibaKI, KubotaA, AbeY, YamamuraS, NakamuraN, TanakaY, NishiharaM ( 2010). Genetic engineering of novel flower colour by suppression of anthocyanin modification genes in gentian J Plant Physiol 167, 231-237. [本文引用: 1]
NegishiT, OshimaK, HattoriM, KanaiM, ManoS, NishimuraM, YoshidaK ( 2012). Tonoplast- and plasma membrane-localized aquaporin-family transporters in blue hydrangea sepals of aluminum hyperaccumulating plant PLoS One 7, e43189. [本文引用: 1]
NishizakiY, YasunagaM, OkamotoE, OkamotoM, HiroseY, YamaguchiM, OzekiY, SasakiN ( 2013). p-Hydroxybenzoyl-glucose is a Zwitter donor for the biosynthesis of 7-polyacylated anthocyanin in Delphinium Plant Cell 25, 4150-4165. [本文引用: 2]
NodaN ( 2018). Recent advances in the research and development of blue flowers Breed Sci 68, 79-87. [本文引用: 1]
NodaN, AidaR, KishimotoS, IshiguroK, Fukuchi-MizutaniM, TanakaY, OhmiyaA ( 2013). Genetic engineering of novel bluer-colored chrysanthemums produced by accumulation of delphinidin-based anthocyanins Plant Cell Physiol 54, 1684-1695. [本文引用: 1]
NodaN, YoshiokaS, KishimotoS, NakayamaM, DouzonoM, TanakaY, AidaR ( 2017). Generation of blue chrysanthemums by anthocyanin B-ring hydroxylation and glucosylation and its coloration mechanism Sci Adv 3, e1602785. [本文引用: 4]
OhnishiM, Fukada-TanakaS, HoshinoA, TakadaJ, InagakiY, IidaS ( 2005). Characterization of a novel Na+/H+ antiporter gene InNHX2 and comparison of InNHX2 with InNHX1, which is responsible for blue flower coloration by increasing the vacuolar pH in the Japanese morning glory Plant Cell Physiol 46, 259-267. [本文引用: 1]
SasakiN, NakayamaT ( 2014). Achievements and perspectives in biochemistry concerning anthocyanin modification for blue flower coloration Plant Cell Physiol 56, 28-40. [本文引用: 3]
SasakiN, NishizakiY, OzekiY, MiyaharaT ( 2014). The role of acyl-glucose in anthocyanin modifications Molecules 19, 18747-18766.
SatoM, KawabeT, HosokawaM, TatsuzawaF, DoiM ( 2011). Tissue culture-induced flower-color changes in Saintpaulia caused by excision of the transposon inserted in the flavonoid 3′,5′ hydroxylase (F3′5′H) promoter Plant Cell Rep 30, 929-939.
ShojiK, MikiN, NakajimaN, MomonoiK, KatoC, YoshidaK ( 2007). Perianth bottom-specific blue color development in tulip cv. Murasakizuisho requires ferric ions Plant Cell Physiol 48, 243-251. [本文引用: 1]
SigurdsonGT, GiustiMM ( 2014). Bathochromic and hyperchromic effects of aluminum salt complexation by anthocyanins from edible sources for blue color development J Agric Food Chem 62, 6955-6965. [本文引用: 1]
SigurdsonGT, RobbinsRJ, CollinsTM, GiustiMM ( 2016). Evaluating the role of metal ions in the bathochromic and hyperchromic responses of cyanidin derivatives in acidic and alkaline pH Food Chem 208, 26-34. [本文引用: 3]
SigurdsonGT, RobbinsRJ, CollinsTM, GiustiMM ( 2017a). Effects of hydroxycinnamic acids on blue color expression of cyanidin derivatives and their metal chelates Food Chem 234, 131-138. [本文引用: 1]
SigurdsonGT, RobbinsRJ, CollinsTM, GiustiMM ( 2017b). Spectral and colorimetric characteristics of metal chelates of acylated cyanidin derivatives Food Chem 221, 1088-1095. [本文引用: 1]
SigurdsonGT, RobbinsRJ, CollinsTM, GiustiMM ( 2018a). Impact of location, type, and number of glycosidic substitutions on the color expression of o-dihydroxylated anthocyanidins Food Chem 268, 416-423. [本文引用: 1]
SigurdsonGT, TangPP, GiustiMM ( 2018b). Cis-trans configuration of coumaric acid acylation affects the spectral and colorimetric properties of anthocyanins Molecules 23, 598. [本文引用: 1]
SunLL, LiY, LiSS, WuXJ, HuBZ, ChangY ( 2014). Identi?cation and characterisation of DfCHS, a chalcone synthase gene regulated by temperature and ultraviolet in Dryopteris fragrans Cell Mol Biol (Noisy-le-Grand, France) 60, 1-7. [本文引用: 1]
TaiDQ, TianJ, ZhangJ, SongTT, YaoYC ( 2014). A Malus crabapple chalcone synthase gene, McCHS, regulates red petal color and flavonoid biosynthesis PLoS One 9, e110570. [本文引用: 1]
TakamuraT, MatsuzakiT ( 2015). Characteristics of F1 progenies obtained by crosses between bluish-violet flowered cyclamen and other cyclamen cultivars Hort Res Japan 14, 412.
TakedaK, YanagisawaM, KifuneT, KinoshitaT, TimberlakeCF ( 1994). A blue pigment complex in flowers of Salvia patens Phytochemistry 35, 1167-1169. [本文引用: 1]
TamuraH, KondoT, GotoT ( 1986). The composition of commelinin, a highly associated metalloanthocyanin present in the blue flower petals of Commelina communis Tetrahedron Lett 27, 1801-1804. [本文引用: 1]
TanakaY, BruglieraF ( 2013). Flower colour and cytochromes P450 Philos Trans Roy Soc B Biol Sci 368, 20120432. [本文引用: 1]
TanakaY, BruglieraF, ChandlerS ( 2009). Recent progress of flower colour modification by biotechnology Int J Mol Sci 10, 5350-5369. [本文引用: 5]
TanakaY, SasakiN, OhmiyaA ( 2008). Biosynthesis of plant pigments: anthocyanins, betalains and carotenoids Plant J 54, 733-749. [本文引用: 1]
TanakaY, TsudaS, KusumiT ( 1998). Metabolic engineering to modify flower color Plant Cell Physiol 39, 1119-1126. [本文引用: 1]
VerweijW, SpeltC, Di SansebastianoGP, VermeerJ, RealeL, FerrantiF, KoesR, QuattrocchioF ( 2008). An H+ P-ATPase on the tonoplast determines vacuolar pH and flower colour Nat Cell Biol 10, 1456-1462. [本文引用: 1]
YoshidaK, MikiN, MomonoiK, KawachiM, KatouK, OkazakiY, UozumiN, MaeshimaM, KondoT ( 2009a). Synchrony between flower opening and petal-color change from red to blue in morning glory, Ipomoea tricolor cv ‘Heavenly Blue’. Proc Jpn Acad Ser B 85, 187-197. [本文引用: 3]
YoshidaK, MoriM, KondoT ( 2009b). Blue flower color development by anthocyanins: from chemical structure to cell physiology Nat Prod Rep 26, 884-915. [本文引用: 9]
YoshidaK, TojoK, MoriM, YamashitaK, KitaharaS, NodaM, UchiyamaS ( 2015). Chemical mechanism of petal color development of Nemophila menziesii by a metalloanthocyanin, nemophilin Tetrahedron 71, 9123-9130. [本文引用: 1]
YoshidaK, ToyamaY, KamedaK, KondoT ( 2000). Contribution of each caffeoyl residue of the pigment molecule of gentiodelphin to blue color development Phytochemistry 54, 85-92. [本文引用: 3]
YoshidaK, Toyama-KatoY, KamedaK, KondoT ( 2003). Sepal color variation of Hydrangea macrophylla and vacuolar pH measured with a proton-selective microelectrode Plant Cell Physiol 44, 262-268. [本文引用: 1]
ZeinipourM, AzadiP, MajdA, KermaniMJ, IrianS, HosseiniSM, MiiM ( 2018). Agroinfiltration: a rapid and reliable method to select suitable rose cultivars for blue flower production Physiol Mol Biol Plant 24, 503-511. [本文引用: 1]
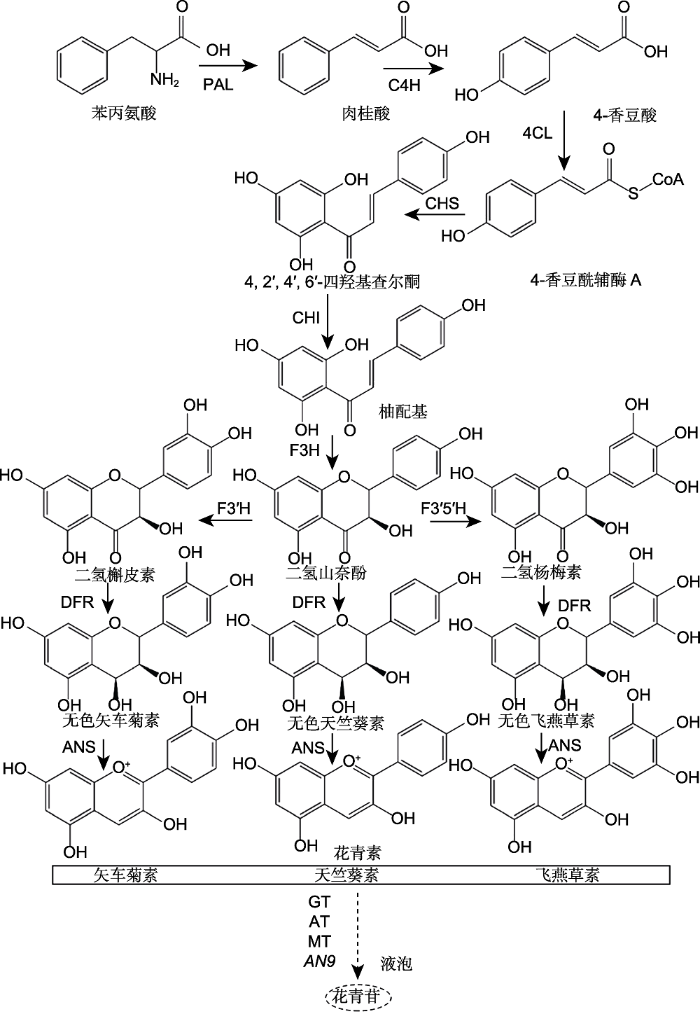
... 作为植物色素, 花青苷在植物的花瓣呈色中起重要作用, 控制着从粉色到蓝紫色一系列花色(Zhao and Tao, 2015).花青素分子在单体的情况下极不稳定, 易在中性或弱酸性介质中被水解失色.因此, 花青素生成的蓝色往往还需要通过糖基化形成花青苷, 之后再通过酰化、甲基化、金属离子络合、共染色和液泡pH值等条件来加强蓝色的稳定性(杨少勇等, 2003; Tanaka et al., 2009; Yoshida et al., 2009b). ...
... 花青苷可通过酰基转移酶以物种特异方式进行酰基化修饰.花青苷根据酰化的数量可分为单酰或多酰花青苷(杨少勇等, 2003).由于蓝色花花青苷大多存在酰化现象, 所以花青苷的酰化一直被认为是蓝色花着色的重要步骤之一(Yoshida et al., 2009b).花青苷的酰化由BAHD和SCPL两个不同的蛋白家族控制.BAHD家族使用酰基-CoA作为酰基供体, 在细胞质中对花青苷进行酰化修饰; SCPL家族则以酰基-葡萄糖作为酰基供体, 在液泡中进行酰化修饰(Miyahara et al., 2013; Nishizaki et al., 2013; Sasaki et al., 2014).龙胆翠雀花素(gentiodelphin)来自蓝色龙胆(Gentiana makinoi), 是第1个被发表的完整多酰花青苷, 龙胆翠雀花素是观赏龙胆植物蓝色花瓣的主要决定因素.龙胆翠雀花素在3位、5位和3'位3个位点上糖基化, 之后通过5位与3'位的糖苷酰化形成飞燕草素3-O-葡糖基-5-O-咖啡酰-葡萄糖基-3'-O-咖啡酰-葡萄糖苷的结构(图2) (Yoshida et al., 2009b).Nakatsuka等(2010)通过RNAi技术沉默龙胆(Gentiana cv. ‘Albireo’)中负责3′与5位酰化的基因, 使植物只能产生飞燕草素3,5,3'-O-三葡萄糖苷的花青苷.酰化相关基因沉默植株的花色明显淡于野生型.这为酰化影响蓝色呈色提供了直接的体内证据.Ishii等(2017)通过对飞燕草品种的筛选得到AA7BG-GT1与AA7BG-GT2双敲除的突变体, 该突变体与野生型相比无法在7位的酰基上继续添加葡萄糖, 因此无法生成飞燕草苷(violdelphin), 其花色最终表现为淡粉色. ...
云南省蓝色花植物研究初探 1 2002
... 花色是观赏植物的一个重要表现特征(Tanaka et al., 2008), 蓝色花在花卉产业中需求旺盛.然而自然界中具有蓝色花的植物数量却相对较少.以云南省为例, 全省被子植物共17 000种, 其中蓝色花植物450种, 只占全省植物资源的2.7% (杨少勇和樊国盛, 2002).切花中, 月季(Rosa chinensis)和康乃馨(Dianthus caryophyllus)尚未拥有真正意义上的蓝色花; 通过转基因手段只获得了紫色或蓝紫色种质(Tanaka et al., 1998, 2009; Sasaki and Nakayama, 2014).菊花(Dendranthema morifolium)通过转基因技术于2017年获得真正意义上的蓝色花(Noda et al., 2017).因此, 研究蓝色花形成的分子机理对于花卉新品种培育具有十分重要的意义.本文从花青苷合成与代谢途径、酰化对花青苷呈色及稳定的影响、金属离子对花青苷的作用、液泡pH值相关基因以及蓝色花分子育种等方面进行综述, 以期为蓝色花种质创建提供借鉴. ...
Genes affecting flower colour and pH of flower limb homogenates in Petunia hybrida 1 1983
... 目前已知植物液泡的pH环境由PH1-PH7和InNHX1- InNHX2两组基因控制.PH1-7与H+的转运相关, 而InNHX1-2负责K+/Na+的转运(Vlaming et al., 1983; Fukadatanaka et al., 2000).Verweij等(2008)发现PH5编码的蛋白为定位于液泡膜上的H+P3A-ATPase质子泵, 负责向液泡运输氢离子, 使液泡pH值下降.PH1最早被认为是一种编码Mg2+转运蛋白的基因, 仅存在于细菌中(Kühlbrandt, 2004).Faraco等(2014)通过对矮牵牛pH突变体的研究, 发现PH1是一种定位于液泡膜上的P3B-ATPase类型蛋白, 该蛋白与PH5相互作用, 可加强PH5的转运能力从而使液泡达到超酸化.PH3和PH4基因编码与转录因子AN1和AN11相互作用的WRKY和MYB蛋白, 可调控PH5基因的表达(Faraco et al., 2017).Fukadatanaka等(2000)研究表明, InNHX1和InNHX2基因编码K+/ Na+的转运蛋白, 可以提升液泡pH值.Ohnishi等(2005)研究表明, InNHX1在根、茎和叶中几乎不表达, 开花前12个小时在花梗中开始表达; 而InNHX2在根、茎和叶中表达, 在花瓣中的表达逐渐增强. ...
Differential inductions of phenylalanine ammonia-lyase and chalcone synthase during wounding, salicylic acid treatment, and salinity stress in safflower, Carthamus tinctorius 1 2014
Structure and molecular stacking of anthocyanins—flower color variation 1 1991
... 金属花青苷是一种自组装的超分子金属络合物, 由化学计量的花青苷、黄酮和金属离子以6:6:2的比例组合而成(图3) (Tanaka et al., 2009).目前已经发现鸭跖草苷(commelina)、原矢车菊苷(protocyanin)、原飞燕草苷(protodelphin)、含氰鼠尾草苷(cyanosalvianin)和粉蝶花苷(nemophilin)共5种金属花青苷(表1; 图4) (Hondo et al., 1992; Kondo et al., 1994; Takeda et al., 1994; Mori et al., 2008; Yoshida et al., 2015).鸭跖草苷和原矢车菊苷是最先被研究的2种金属花青苷, 通过解析成分, 得到2种金属花青苷的金属离子成分分别为Mg2+和Fe3+、Mg2+ (Tamura et al., 1986; Goto and Kondo, 1991; Yoshida et al., 2009b).Yoshida等(2009b)通过总结金属花青苷的构成, 报道了在不同发色团下, 形成蓝色花所需要的金属粒子类型, 在飞燕草素的发色团中Mg2+就可以满足蓝色的生成; 在以矢车菊素或牵牛花素(petunidin)为发色团的情况下, 必须有1/6Fe3+; 天竺葵素(pelargonidin)或锦葵素(malvidin)的发色团则无法形成蓝色的金属混合物. ...
Transgenic plants exhibiting altered flower color and methods for producing same 1 2000
Structural basis of blue-colour development in flower petals from Commelina communis 1 1992
... 金属花青苷是一种自组装的超分子金属络合物, 由化学计量的花青苷、黄酮和金属离子以6:6:2的比例组合而成(图3) (Tanaka et al., 2009).目前已经发现鸭跖草苷(commelina)、原矢车菊苷(protocyanin)、原飞燕草苷(protodelphin)、含氰鼠尾草苷(cyanosalvianin)和粉蝶花苷(nemophilin)共5种金属花青苷(表1; 图4) (Hondo et al., 1992; Kondo et al., 1994; Takeda et al., 1994; Mori et al., 2008; Yoshida et al., 2015).鸭跖草苷和原矢车菊苷是最先被研究的2种金属花青苷, 通过解析成分, 得到2种金属花青苷的金属离子成分分别为Mg2+和Fe3+、Mg2+ (Tamura et al., 1986; Goto and Kondo, 1991; Yoshida et al., 2009b).Yoshida等(2009b)通过总结金属花青苷的构成, 报道了在不同发色团下, 形成蓝色花所需要的金属粒子类型, 在飞燕草素的发色团中Mg2+就可以满足蓝色的生成; 在以矢车菊素或牵牛花素(petunidin)为发色团的情况下, 必须有1/6Fe3+; 天竺葵素(pelargonidin)或锦葵素(malvidin)的发色团则无法形成蓝色的金属混合物. ...
Flower colour modification of chrysanthemum by suppression of F3'H and overexpression of the exogenous Senecio cruentus F3'5'H gene 1 2013
Composition of protocyanin, a self-assembled supramolecular pigment from the blue cornflower, Centaurea cyanus 1 1994
... 金属花青苷是一种自组装的超分子金属络合物, 由化学计量的花青苷、黄酮和金属离子以6:6:2的比例组合而成(图3) (Tanaka et al., 2009).目前已经发现鸭跖草苷(commelina)、原矢车菊苷(protocyanin)、原飞燕草苷(protodelphin)、含氰鼠尾草苷(cyanosalvianin)和粉蝶花苷(nemophilin)共5种金属花青苷(表1; 图4) (Hondo et al., 1992; Kondo et al., 1994; Takeda et al., 1994; Mori et al., 2008; Yoshida et al., 2015).鸭跖草苷和原矢车菊苷是最先被研究的2种金属花青苷, 通过解析成分, 得到2种金属花青苷的金属离子成分分别为Mg2+和Fe3+、Mg2+ (Tamura et al., 1986; Goto and Kondo, 1991; Yoshida et al., 2009b).Yoshida等(2009b)通过总结金属花青苷的构成, 报道了在不同发色团下, 形成蓝色花所需要的金属粒子类型, 在飞燕草素的发色团中Mg2+就可以满足蓝色的生成; 在以矢车菊素或牵牛花素(petunidin)为发色团的情况下, 必须有1/6Fe3+; 天竺葵素(pelargonidin)或锦葵素(malvidin)的发色团则无法形成蓝色的金属混合物. ...
Biology, structure and mechanism of P-type ATPases 1 2004
... 目前已知植物液泡的pH环境由PH1-PH7和InNHX1- InNHX2两组基因控制.PH1-7与H+的转运相关, 而InNHX1-2负责K+/Na+的转运(Vlaming et al., 1983; Fukadatanaka et al., 2000).Verweij等(2008)发现PH5编码的蛋白为定位于液泡膜上的H+P3A-ATPase质子泵, 负责向液泡运输氢离子, 使液泡pH值下降.PH1最早被认为是一种编码Mg2+转运蛋白的基因, 仅存在于细菌中(Kühlbrandt, 2004).Faraco等(2014)通过对矮牵牛pH突变体的研究, 发现PH1是一种定位于液泡膜上的P3B-ATPase类型蛋白, 该蛋白与PH5相互作用, 可加强PH5的转运能力从而使液泡达到超酸化.PH3和PH4基因编码与转录因子AN1和AN11相互作用的WRKY和MYB蛋白, 可调控PH5基因的表达(Faraco et al., 2017).Fukadatanaka等(2000)研究表明, InNHX1和InNHX2基因编码K+/ Na+的转运蛋白, 可以提升液泡pH值.Ohnishi等(2005)研究表明, InNHX1在根、茎和叶中几乎不表达, 开花前12个小时在花梗中开始表达; 而InNHX2在根、茎和叶中表达, 在花瓣中的表达逐渐增强. ...
Malvidin- 3- O-glucoside-5-O-(6-acetylglucoside) and its colour manifestation in ‘Johnson’s Blue’ and other ‘Blue’ geraniums 1 1997
Heterologous expression of dihydroflavonol 4-reductases from various plants 1 2002
... DFR是具有底物特异性的酶, 不同植物中DFR的特异性表现不同(Martens et al., 2002).康乃馨中DFR可以还原DHK、DHQ和DHM; 在矮牵牛(Petunia hybrida)中DFR只能还原DHQ和DHM (Zhao and Tao, 2015).Zhao等(2012)发现DFR的大量表达与花青苷的积累呈正相关. ...
Stability to light, heat, and hydrogen peroxide at different pH values and DPPH radical scavenging activity of acylated anthocyanins from red radish extract 1 2007
... 花青素本身很不稳定、易分解, 通过糖基化形成花青苷可以提升稳定性, 而花青苷的酰化可以大大增强稳定性.酰化的花青苷比非酰化的花青苷在高温、pH环境变化和强光等影响中稳定性更高.研究表明, 花青苷酰化的数量、位置以及种类对于花青苷的稳定及颜色有重要影响(Matsufuji et al., 2007; 李云等, 2010; Zhao et al., 2017).Yoshida等(2000)通过碱性水解的方式从纯化的龙胆翠雀花素中分别得到3′位与5位的单酰花青苷与完全去酰基的花青苷.通过记录几种物质在水溶液中的UV-VIS光谱, 发现3′位的单酰基花青苷更蓝, 基本接近龙胆翠雀素本身.类似的结果也在蓝色新西兰蓝莓(Dianella nigra)浆果的花青苷分析中得到验证. ...
A new petunia flower colour generated by transformation of a mutant with a maize gene 1 1987
Acyl-glucose-dependent glucosyltransferase catalyzes the final step of anthocyanin formation in Arabidopsis 1 2013
... 花青苷可通过酰基转移酶以物种特异方式进行酰基化修饰.花青苷根据酰化的数量可分为单酰或多酰花青苷(杨少勇等, 2003).由于蓝色花花青苷大多存在酰化现象, 所以花青苷的酰化一直被认为是蓝色花着色的重要步骤之一(Yoshida et al., 2009b).花青苷的酰化由BAHD和SCPL两个不同的蛋白家族控制.BAHD家族使用酰基-CoA作为酰基供体, 在细胞质中对花青苷进行酰化修饰; SCPL家族则以酰基-葡萄糖作为酰基供体, 在液泡中进行酰化修饰(Miyahara et al., 2013; Nishizaki et al., 2013; Sasaki et al., 2014).龙胆翠雀花素(gentiodelphin)来自蓝色龙胆(Gentiana makinoi), 是第1个被发表的完整多酰花青苷, 龙胆翠雀花素是观赏龙胆植物蓝色花瓣的主要决定因素.龙胆翠雀花素在3位、5位和3'位3个位点上糖基化, 之后通过5位与3'位的糖苷酰化形成飞燕草素3-O-葡糖基-5-O-咖啡酰-葡萄糖基-3'-O-咖啡酰-葡萄糖苷的结构(图2) (Yoshida et al., 2009b).Nakatsuka等(2010)通过RNAi技术沉默龙胆(Gentiana cv. ‘Albireo’)中负责3′与5位酰化的基因, 使植物只能产生飞燕草素3,5,3'-O-三葡萄糖苷的花青苷.酰化相关基因沉默植株的花色明显淡于野生型.这为酰化影响蓝色呈色提供了直接的体内证据.Ishii等(2017)通过对飞燕草品种的筛选得到AA7BG-GT1与AA7BG-GT2双敲除的突变体, 该突变体与野生型相比无法在7位的酰基上继续添加葡萄糖, 因此无法生成飞燕草苷(violdelphin), 其花色最终表现为淡粉色. ...
Cyanosalvianin, a supramolecular blue metalloanthocyanin, from petals of Salvia uliginosa 1 2008
... 金属花青苷是一种自组装的超分子金属络合物, 由化学计量的花青苷、黄酮和金属离子以6:6:2的比例组合而成(图3) (Tanaka et al., 2009).目前已经发现鸭跖草苷(commelina)、原矢车菊苷(protocyanin)、原飞燕草苷(protodelphin)、含氰鼠尾草苷(cyanosalvianin)和粉蝶花苷(nemophilin)共5种金属花青苷(表1; 图4) (Hondo et al., 1992; Kondo et al., 1994; Takeda et al., 1994; Mori et al., 2008; Yoshida et al., 2015).鸭跖草苷和原矢车菊苷是最先被研究的2种金属花青苷, 通过解析成分, 得到2种金属花青苷的金属离子成分分别为Mg2+和Fe3+、Mg2+ (Tamura et al., 1986; Goto and Kondo, 1991; Yoshida et al., 2009b).Yoshida等(2009b)通过总结金属花青苷的构成, 报道了在不同发色团下, 形成蓝色花所需要的金属粒子类型, 在飞燕草素的发色团中Mg2+就可以满足蓝色的生成; 在以矢车菊素或牵牛花素(petunidin)为发色团的情况下, 必须有1/6Fe3+; 天竺葵素(pelargonidin)或锦葵素(malvidin)的发色团则无法形成蓝色的金属混合物. ...
Genetic engineering of novel flower colour by suppression of anthocyanin modification genes in gentian 1 2010
Characterization of a novel Na+/H+ antiporter gene InNHX2 and comparison of InNHX2 with InNHX1, which is responsible for blue flower coloration by increasing the vacuolar pH in the Japanese morning glory 1 2005
... 目前已知植物液泡的pH环境由PH1-PH7和InNHX1- InNHX2两组基因控制.PH1-7与H+的转运相关, 而InNHX1-2负责K+/Na+的转运(Vlaming et al., 1983; Fukadatanaka et al., 2000).Verweij等(2008)发现PH5编码的蛋白为定位于液泡膜上的H+P3A-ATPase质子泵, 负责向液泡运输氢离子, 使液泡pH值下降.PH1最早被认为是一种编码Mg2+转运蛋白的基因, 仅存在于细菌中(Kühlbrandt, 2004).Faraco等(2014)通过对矮牵牛pH突变体的研究, 发现PH1是一种定位于液泡膜上的P3B-ATPase类型蛋白, 该蛋白与PH5相互作用, 可加强PH5的转运能力从而使液泡达到超酸化.PH3和PH4基因编码与转录因子AN1和AN11相互作用的WRKY和MYB蛋白, 可调控PH5基因的表达(Faraco et al., 2017).Fukadatanaka等(2000)研究表明, InNHX1和InNHX2基因编码K+/ Na+的转运蛋白, 可以提升液泡pH值.Ohnishi等(2005)研究表明, InNHX1在根、茎和叶中几乎不表达, 开花前12个小时在花梗中开始表达; 而InNHX2在根、茎和叶中表达, 在花瓣中的表达逐渐增强. ...
Achievements and perspectives in biochemistry concerning anthocyanin modification for blue flower coloration 3 2014
... 花色是观赏植物的一个重要表现特征(Tanaka et al., 2008), 蓝色花在花卉产业中需求旺盛.然而自然界中具有蓝色花的植物数量却相对较少.以云南省为例, 全省被子植物共17 000种, 其中蓝色花植物450种, 只占全省植物资源的2.7% (杨少勇和樊国盛, 2002).切花中, 月季(Rosa chinensis)和康乃馨(Dianthus caryophyllus)尚未拥有真正意义上的蓝色花; 通过转基因手段只获得了紫色或蓝紫色种质(Tanaka et al., 1998, 2009; Sasaki and Nakayama, 2014).菊花(Dendranthema morifolium)通过转基因技术于2017年获得真正意义上的蓝色花(Noda et al., 2017).因此, 研究蓝色花形成的分子机理对于花卉新品种培育具有十分重要的意义.本文从花青苷合成与代谢途径、酰化对花青苷呈色及稳定的影响、金属离子对花青苷的作用、液泡pH值相关基因以及蓝色花分子育种等方面进行综述, 以期为蓝色花种质创建提供借鉴. ...
... 花青苷可通过酰基转移酶以物种特异方式进行酰基化修饰.花青苷根据酰化的数量可分为单酰或多酰花青苷(杨少勇等, 2003).由于蓝色花花青苷大多存在酰化现象, 所以花青苷的酰化一直被认为是蓝色花着色的重要步骤之一(Yoshida et al., 2009b).花青苷的酰化由BAHD和SCPL两个不同的蛋白家族控制.BAHD家族使用酰基-CoA作为酰基供体, 在细胞质中对花青苷进行酰化修饰; SCPL家族则以酰基-葡萄糖作为酰基供体, 在液泡中进行酰化修饰(Miyahara et al., 2013; Nishizaki et al., 2013; Sasaki et al., 2014).龙胆翠雀花素(gentiodelphin)来自蓝色龙胆(Gentiana makinoi), 是第1个被发表的完整多酰花青苷, 龙胆翠雀花素是观赏龙胆植物蓝色花瓣的主要决定因素.龙胆翠雀花素在3位、5位和3'位3个位点上糖基化, 之后通过5位与3'位的糖苷酰化形成飞燕草素3-O-葡糖基-5-O-咖啡酰-葡萄糖基-3'-O-咖啡酰-葡萄糖苷的结构(图2) (Yoshida et al., 2009b).Nakatsuka等(2010)通过RNAi技术沉默龙胆(Gentiana cv. ‘Albireo’)中负责3′与5位酰化的基因, 使植物只能产生飞燕草素3,5,3'-O-三葡萄糖苷的花青苷.酰化相关基因沉默植株的花色明显淡于野生型.这为酰化影响蓝色呈色提供了直接的体内证据.Ishii等(2017)通过对飞燕草品种的筛选得到AA7BG-GT1与AA7BG-GT2双敲除的突变体, 该突变体与野生型相比无法在7位的酰基上继续添加葡萄糖, 因此无法生成飞燕草苷(violdelphin), 其花色最终表现为淡粉色. ...
The role of acyl-glucose in anthocyanin modifications 0 2014
Tissue culture-induced flower-color changes in Saintpaulia caused by excision of the transposon inserted in the flavonoid 3′,5′ hydroxylase (F3′5′H) promoter 0 2011
Perianth bottom-specific blue color development in tulip cv. Murasakizuisho requires ferric ions 1 2007
... 与金属花青苷相比, 非化学计量的金属络合物在蓝色花瓣中更为常见, 但与金属花青苷相比对花青苷稳定性的提升要小得多(Tanaka et al., 2009).金属离子一般与B环上含有2-3羟基的花青素络合, 而单羟基花青素基本不发生这种络合反应(Sigurdson et al., 2016; Fedenko et al., 2017).Shoji等(2007)发现郁金香(Tulipa gesneriana)花被的蓝紫差异是由于蓝色部分比紫色部分积累更多的Fe3+.Sigurdson和Giusti (2014)通过向不同植物花青苷提取物中添加AlCl3, 发现所有的花青苷都与铝离子发生络合并表现出红移; 随着AlCl3浓度的增加, 吸光值也随之增加.Sigurdson等(2016)通过对酰化和非酰化的矢车菊素衍生物进行多种不同的金属络合, 发现所有金属离子都可使花青苷产生一定的红移与增色, 但不同金属离子产生的效果不同, 产生红移的金属离子作用大小为Fe3+? Ga3+>Al3+>Cr3+>Mg2+, 非酰化矢车菊素产生的红移要大于酰化的矢车菊素, 但最大吸光值小于酰化矢车菊素.Negishi等(2012)在绣球花(Hydrangea macrophylla)中鉴定了VALT和PALT1转运蛋白, 并确定这2个蛋白分别作用于液泡膜与胞质膜进行Al3+转运.这表明金属离子转运蛋白在花青苷金属离子络合过程中起重要作用. ...
Bathochromic and hyperchromic effects of aluminum salt complexation by anthocyanins from edible sources for blue color development 1 2014
... 与金属花青苷相比, 非化学计量的金属络合物在蓝色花瓣中更为常见, 但与金属花青苷相比对花青苷稳定性的提升要小得多(Tanaka et al., 2009).金属离子一般与B环上含有2-3羟基的花青素络合, 而单羟基花青素基本不发生这种络合反应(Sigurdson et al., 2016; Fedenko et al., 2017).Shoji等(2007)发现郁金香(Tulipa gesneriana)花被的蓝紫差异是由于蓝色部分比紫色部分积累更多的Fe3+.Sigurdson和Giusti (2014)通过向不同植物花青苷提取物中添加AlCl3, 发现所有的花青苷都与铝离子发生络合并表现出红移; 随着AlCl3浓度的增加, 吸光值也随之增加.Sigurdson等(2016)通过对酰化和非酰化的矢车菊素衍生物进行多种不同的金属络合, 发现所有金属离子都可使花青苷产生一定的红移与增色, 但不同金属离子产生的效果不同, 产生红移的金属离子作用大小为Fe3+? Ga3+>Al3+>Cr3+>Mg2+, 非酰化矢车菊素产生的红移要大于酰化的矢车菊素, 但最大吸光值小于酰化矢车菊素.Negishi等(2012)在绣球花(Hydrangea macrophylla)中鉴定了VALT和PALT1转运蛋白, 并确定这2个蛋白分别作用于液泡膜与胞质膜进行Al3+转运.这表明金属离子转运蛋白在花青苷金属离子络合过程中起重要作用. ...
Evaluating the role of metal ions in the bathochromic and hyperchromic responses of cyanidin derivatives in acidic and alkaline pH 3 2016
Characteristics of F1 progenies obtained by crosses between bluish-violet flowered cyclamen and other cyclamen cultivars 0 2015
A blue pigment complex in flowers of Salvia patens 1 1994
... 金属花青苷是一种自组装的超分子金属络合物, 由化学计量的花青苷、黄酮和金属离子以6:6:2的比例组合而成(图3) (Tanaka et al., 2009).目前已经发现鸭跖草苷(commelina)、原矢车菊苷(protocyanin)、原飞燕草苷(protodelphin)、含氰鼠尾草苷(cyanosalvianin)和粉蝶花苷(nemophilin)共5种金属花青苷(表1; 图4) (Hondo et al., 1992; Kondo et al., 1994; Takeda et al., 1994; Mori et al., 2008; Yoshida et al., 2015).鸭跖草苷和原矢车菊苷是最先被研究的2种金属花青苷, 通过解析成分, 得到2种金属花青苷的金属离子成分分别为Mg2+和Fe3+、Mg2+ (Tamura et al., 1986; Goto and Kondo, 1991; Yoshida et al., 2009b).Yoshida等(2009b)通过总结金属花青苷的构成, 报道了在不同发色团下, 形成蓝色花所需要的金属粒子类型, 在飞燕草素的发色团中Mg2+就可以满足蓝色的生成; 在以矢车菊素或牵牛花素(petunidin)为发色团的情况下, 必须有1/6Fe3+; 天竺葵素(pelargonidin)或锦葵素(malvidin)的发色团则无法形成蓝色的金属混合物. ...
The composition of commelinin, a highly associated metalloanthocyanin present in the blue flower petals of Commelina communis 1 1986
... 金属花青苷是一种自组装的超分子金属络合物, 由化学计量的花青苷、黄酮和金属离子以6:6:2的比例组合而成(图3) (Tanaka et al., 2009).目前已经发现鸭跖草苷(commelina)、原矢车菊苷(protocyanin)、原飞燕草苷(protodelphin)、含氰鼠尾草苷(cyanosalvianin)和粉蝶花苷(nemophilin)共5种金属花青苷(表1; 图4) (Hondo et al., 1992; Kondo et al., 1994; Takeda et al., 1994; Mori et al., 2008; Yoshida et al., 2015).鸭跖草苷和原矢车菊苷是最先被研究的2种金属花青苷, 通过解析成分, 得到2种金属花青苷的金属离子成分分别为Mg2+和Fe3+、Mg2+ (Tamura et al., 1986; Goto and Kondo, 1991; Yoshida et al., 2009b).Yoshida等(2009b)通过总结金属花青苷的构成, 报道了在不同发色团下, 形成蓝色花所需要的金属粒子类型, 在飞燕草素的发色团中Mg2+就可以满足蓝色的生成; 在以矢车菊素或牵牛花素(petunidin)为发色团的情况下, 必须有1/6Fe3+; 天竺葵素(pelargonidin)或锦葵素(malvidin)的发色团则无法形成蓝色的金属混合物. ...
Flower colour and cytochromes P450 1 2013
... F3H、F3′H和F3′5′H是该途径合成中的另一组关键酶基因.自然界中大多数蓝色花的花青苷都由飞燕草素(delphinidin)组成, 而DHM是飞燕草素合成的必要底物.因此F3′5′H也被称为蓝色花的核心基因(Tanaka and Brugliera, 2013).Holton等(1993)研究表明, F3′5′H属于细胞色素P450家族.Sato等(2011)发现F3′5′H启动子区域转座子的插入与缺失引起花色变化.目前, 已从多种植物中分离出F3′5′H基因并对其进行深入研究.在蓝色花育种中, 为避免底物竞争, 多数在导入F3′5′H基因的同时会对F3′H基因进行RNAi处理(Brugliera et al., 2013). ...
Recent progress of flower colour modification by biotechnology 5 2009
... 花色是观赏植物的一个重要表现特征(Tanaka et al., 2008), 蓝色花在花卉产业中需求旺盛.然而自然界中具有蓝色花的植物数量却相对较少.以云南省为例, 全省被子植物共17 000种, 其中蓝色花植物450种, 只占全省植物资源的2.7% (杨少勇和樊国盛, 2002).切花中, 月季(Rosa chinensis)和康乃馨(Dianthus caryophyllus)尚未拥有真正意义上的蓝色花; 通过转基因手段只获得了紫色或蓝紫色种质(Tanaka et al., 1998, 2009; Sasaki and Nakayama, 2014).菊花(Dendranthema morifolium)通过转基因技术于2017年获得真正意义上的蓝色花(Noda et al., 2017).因此, 研究蓝色花形成的分子机理对于花卉新品种培育具有十分重要的意义.本文从花青苷合成与代谢途径、酰化对花青苷呈色及稳定的影响、金属离子对花青苷的作用、液泡pH值相关基因以及蓝色花分子育种等方面进行综述, 以期为蓝色花种质创建提供借鉴. ...
... 作为植物色素, 花青苷在植物的花瓣呈色中起重要作用, 控制着从粉色到蓝紫色一系列花色(Zhao and Tao, 2015).花青素分子在单体的情况下极不稳定, 易在中性或弱酸性介质中被水解失色.因此, 花青素生成的蓝色往往还需要通过糖基化形成花青苷, 之后再通过酰化、甲基化、金属离子络合、共染色和液泡pH值等条件来加强蓝色的稳定性(杨少勇等, 2003; Tanaka et al., 2009; Yoshida et al., 2009b). ...
... 花瓣液泡中金属离子对蓝色花的形成有重要作用.目前发现的大多数蓝色花瓣中都存在金属离子, 金属离子被认为是蓝色花形成的一个重要组件(Tanaka et al., 2009).研究金属离子对花青苷显色与稳定的作用对蓝色花育种十分重要. ...
... 金属花青苷是一种自组装的超分子金属络合物, 由化学计量的花青苷、黄酮和金属离子以6:6:2的比例组合而成(图3) (Tanaka et al., 2009).目前已经发现鸭跖草苷(commelina)、原矢车菊苷(protocyanin)、原飞燕草苷(protodelphin)、含氰鼠尾草苷(cyanosalvianin)和粉蝶花苷(nemophilin)共5种金属花青苷(表1; 图4) (Hondo et al., 1992; Kondo et al., 1994; Takeda et al., 1994; Mori et al., 2008; Yoshida et al., 2015).鸭跖草苷和原矢车菊苷是最先被研究的2种金属花青苷, 通过解析成分, 得到2种金属花青苷的金属离子成分分别为Mg2+和Fe3+、Mg2+ (Tamura et al., 1986; Goto and Kondo, 1991; Yoshida et al., 2009b).Yoshida等(2009b)通过总结金属花青苷的构成, 报道了在不同发色团下, 形成蓝色花所需要的金属粒子类型, 在飞燕草素的发色团中Mg2+就可以满足蓝色的生成; 在以矢车菊素或牵牛花素(petunidin)为发色团的情况下, 必须有1/6Fe3+; 天竺葵素(pelargonidin)或锦葵素(malvidin)的发色团则无法形成蓝色的金属混合物. ...
... 与金属花青苷相比, 非化学计量的金属络合物在蓝色花瓣中更为常见, 但与金属花青苷相比对花青苷稳定性的提升要小得多(Tanaka et al., 2009).金属离子一般与B环上含有2-3羟基的花青素络合, 而单羟基花青素基本不发生这种络合反应(Sigurdson et al., 2016; Fedenko et al., 2017).Shoji等(2007)发现郁金香(Tulipa gesneriana)花被的蓝紫差异是由于蓝色部分比紫色部分积累更多的Fe3+.Sigurdson和Giusti (2014)通过向不同植物花青苷提取物中添加AlCl3, 发现所有的花青苷都与铝离子发生络合并表现出红移; 随着AlCl3浓度的增加, 吸光值也随之增加.Sigurdson等(2016)通过对酰化和非酰化的矢车菊素衍生物进行多种不同的金属络合, 发现所有金属离子都可使花青苷产生一定的红移与增色, 但不同金属离子产生的效果不同, 产生红移的金属离子作用大小为Fe3+? Ga3+>Al3+>Cr3+>Mg2+, 非酰化矢车菊素产生的红移要大于酰化的矢车菊素, 但最大吸光值小于酰化矢车菊素.Negishi等(2012)在绣球花(Hydrangea macrophylla)中鉴定了VALT和PALT1转运蛋白, 并确定这2个蛋白分别作用于液泡膜与胞质膜进行Al3+转运.这表明金属离子转运蛋白在花青苷金属离子络合过程中起重要作用. ...
Biosynthesis of plant pigments: anthocyanins, betalains and carotenoids 1 2008
... 花色是观赏植物的一个重要表现特征(Tanaka et al., 2008), 蓝色花在花卉产业中需求旺盛.然而自然界中具有蓝色花的植物数量却相对较少.以云南省为例, 全省被子植物共17 000种, 其中蓝色花植物450种, 只占全省植物资源的2.7% (杨少勇和樊国盛, 2002).切花中, 月季(Rosa chinensis)和康乃馨(Dianthus caryophyllus)尚未拥有真正意义上的蓝色花; 通过转基因手段只获得了紫色或蓝紫色种质(Tanaka et al., 1998, 2009; Sasaki and Nakayama, 2014).菊花(Dendranthema morifolium)通过转基因技术于2017年获得真正意义上的蓝色花(Noda et al., 2017).因此, 研究蓝色花形成的分子机理对于花卉新品种培育具有十分重要的意义.本文从花青苷合成与代谢途径、酰化对花青苷呈色及稳定的影响、金属离子对花青苷的作用、液泡pH值相关基因以及蓝色花分子育种等方面进行综述, 以期为蓝色花种质创建提供借鉴. ...
Metabolic engineering to modify flower color 1 1998
... 花色是观赏植物的一个重要表现特征(Tanaka et al., 2008), 蓝色花在花卉产业中需求旺盛.然而自然界中具有蓝色花的植物数量却相对较少.以云南省为例, 全省被子植物共17 000种, 其中蓝色花植物450种, 只占全省植物资源的2.7% (杨少勇和樊国盛, 2002).切花中, 月季(Rosa chinensis)和康乃馨(Dianthus caryophyllus)尚未拥有真正意义上的蓝色花; 通过转基因手段只获得了紫色或蓝紫色种质(Tanaka et al., 1998, 2009; Sasaki and Nakayama, 2014).菊花(Dendranthema morifolium)通过转基因技术于2017年获得真正意义上的蓝色花(Noda et al., 2017).因此, 研究蓝色花形成的分子机理对于花卉新品种培育具有十分重要的意义.本文从花青苷合成与代谢途径、酰化对花青苷呈色及稳定的影响、金属离子对花青苷的作用、液泡pH值相关基因以及蓝色花分子育种等方面进行综述, 以期为蓝色花种质创建提供借鉴. ...
An H+ P-ATPase on the tonoplast determines vacuolar pH and flower colour 1 2008
... 目前已知植物液泡的pH环境由PH1-PH7和InNHX1- InNHX2两组基因控制.PH1-7与H+的转运相关, 而InNHX1-2负责K+/Na+的转运(Vlaming et al., 1983; Fukadatanaka et al., 2000).Verweij等(2008)发现PH5编码的蛋白为定位于液泡膜上的H+P3A-ATPase质子泵, 负责向液泡运输氢离子, 使液泡pH值下降.PH1最早被认为是一种编码Mg2+转运蛋白的基因, 仅存在于细菌中(Kühlbrandt, 2004).Faraco等(2014)通过对矮牵牛pH突变体的研究, 发现PH1是一种定位于液泡膜上的P3B-ATPase类型蛋白, 该蛋白与PH5相互作用, 可加强PH5的转运能力从而使液泡达到超酸化.PH3和PH4基因编码与转录因子AN1和AN11相互作用的WRKY和MYB蛋白, 可调控PH5基因的表达(Faraco et al., 2017).Fukadatanaka等(2000)研究表明, InNHX1和InNHX2基因编码K+/ Na+的转运蛋白, 可以提升液泡pH值.Ohnishi等(2005)研究表明, InNHX1在根、茎和叶中几乎不表达, 开花前12个小时在花梗中开始表达; 而InNHX2在根、茎和叶中表达, 在花瓣中的表达逐渐增强. ...
Synchrony between flower opening and petal-color change from red to blue in morning glory, Ipomoea tricolor cv 3 2009
... 5种金属花青苷的成分(Yoshida et al., 2009a) ...
... Composition of five metalloanthocyanin (Yoshida et al., 2009a) ...
Blue flower color development by anthocyanins: from chemical structure to cell physiology 9 2009
... 作为植物色素, 花青苷在植物的花瓣呈色中起重要作用, 控制着从粉色到蓝紫色一系列花色(Zhao and Tao, 2015).花青素分子在单体的情况下极不稳定, 易在中性或弱酸性介质中被水解失色.因此, 花青素生成的蓝色往往还需要通过糖基化形成花青苷, 之后再通过酰化、甲基化、金属离子络合、共染色和液泡pH值等条件来加强蓝色的稳定性(杨少勇等, 2003; Tanaka et al., 2009; Yoshida et al., 2009b). ...
... 花青苷可通过酰基转移酶以物种特异方式进行酰基化修饰.花青苷根据酰化的数量可分为单酰或多酰花青苷(杨少勇等, 2003).由于蓝色花花青苷大多存在酰化现象, 所以花青苷的酰化一直被认为是蓝色花着色的重要步骤之一(Yoshida et al., 2009b).花青苷的酰化由BAHD和SCPL两个不同的蛋白家族控制.BAHD家族使用酰基-CoA作为酰基供体, 在细胞质中对花青苷进行酰化修饰; SCPL家族则以酰基-葡萄糖作为酰基供体, 在液泡中进行酰化修饰(Miyahara et al., 2013; Nishizaki et al., 2013; Sasaki et al., 2014).龙胆翠雀花素(gentiodelphin)来自蓝色龙胆(Gentiana makinoi), 是第1个被发表的完整多酰花青苷, 龙胆翠雀花素是观赏龙胆植物蓝色花瓣的主要决定因素.龙胆翠雀花素在3位、5位和3'位3个位点上糖基化, 之后通过5位与3'位的糖苷酰化形成飞燕草素3-O-葡糖基-5-O-咖啡酰-葡萄糖基-3'-O-咖啡酰-葡萄糖苷的结构(图2) (Yoshida et al., 2009b).Nakatsuka等(2010)通过RNAi技术沉默龙胆(Gentiana cv. ‘Albireo’)中负责3′与5位酰化的基因, 使植物只能产生飞燕草素3,5,3'-O-三葡萄糖苷的花青苷.酰化相关基因沉默植株的花色明显淡于野生型.这为酰化影响蓝色呈色提供了直接的体内证据.Ishii等(2017)通过对飞燕草品种的筛选得到AA7BG-GT1与AA7BG-GT2双敲除的突变体, 该突变体与野生型相比无法在7位的酰基上继续添加葡萄糖, 因此无法生成飞燕草苷(violdelphin), 其花色最终表现为淡粉色. ...
... 金属花青苷是一种自组装的超分子金属络合物, 由化学计量的花青苷、黄酮和金属离子以6:6:2的比例组合而成(图3) (Tanaka et al., 2009).目前已经发现鸭跖草苷(commelina)、原矢车菊苷(protocyanin)、原飞燕草苷(protodelphin)、含氰鼠尾草苷(cyanosalvianin)和粉蝶花苷(nemophilin)共5种金属花青苷(表1; 图4) (Hondo et al., 1992; Kondo et al., 1994; Takeda et al., 1994; Mori et al., 2008; Yoshida et al., 2015).鸭跖草苷和原矢车菊苷是最先被研究的2种金属花青苷, 通过解析成分, 得到2种金属花青苷的金属离子成分分别为Mg2+和Fe3+、Mg2+ (Tamura et al., 1986; Goto and Kondo, 1991; Yoshida et al., 2009b).Yoshida等(2009b)通过总结金属花青苷的构成, 报道了在不同发色团下, 形成蓝色花所需要的金属粒子类型, 在飞燕草素的发色团中Mg2+就可以满足蓝色的生成; 在以矢车菊素或牵牛花素(petunidin)为发色团的情况下, 必须有1/6Fe3+; 天竺葵素(pelargonidin)或锦葵素(malvidin)的发色团则无法形成蓝色的金属混合物. ...
... 金属花青苷组成(改自Yoshida et al., 2009b) Formation of metalloanthocyanin (modified from <xref ref-type="bibr" rid="b66">Yoshida et al., 2009b</xref>)Figure 3
金属花青苷组分结构(<xref ref-type="bibr" rid="b66">Yoshida et al., 2009b</xref>) A1-A4为花青苷类物质; F1-F4为黄酮类物质. ...
... Formation of metalloanthocyanin (modified from Yoshida et al., 2009b) Figure 3
金属花青苷组分结构(<xref ref-type="bibr" rid="b66">Yoshida et al., 2009b</xref>) A1-A4为花青苷类物质; F1-F4为黄酮类物质. ...
... 金属花青苷组分结构(Yoshida et al., 2009b) A1-A4为花青苷类物质; F1-F4为黄酮类物质. ...
... A1-A4为花青苷类物质; F1-F4为黄酮类物质.
Structure of metalloanthocyanin component (<xref ref-type="bibr" rid="b66">Yoshida et al., 2009b</xref>) A1-A4 are anthocyanins; F1-F4 are flavonoids. ...
Chemical mechanism of petal color development of Nemophila menziesii by a metalloanthocyanin, nemophilin 1 2015
... 金属花青苷是一种自组装的超分子金属络合物, 由化学计量的花青苷、黄酮和金属离子以6:6:2的比例组合而成(图3) (Tanaka et al., 2009).目前已经发现鸭跖草苷(commelina)、原矢车菊苷(protocyanin)、原飞燕草苷(protodelphin)、含氰鼠尾草苷(cyanosalvianin)和粉蝶花苷(nemophilin)共5种金属花青苷(表1; 图4) (Hondo et al., 1992; Kondo et al., 1994; Takeda et al., 1994; Mori et al., 2008; Yoshida et al., 2015).鸭跖草苷和原矢车菊苷是最先被研究的2种金属花青苷, 通过解析成分, 得到2种金属花青苷的金属离子成分分别为Mg2+和Fe3+、Mg2+ (Tamura et al., 1986; Goto and Kondo, 1991; Yoshida et al., 2009b).Yoshida等(2009b)通过总结金属花青苷的构成, 报道了在不同发色团下, 形成蓝色花所需要的金属粒子类型, 在飞燕草素的发色团中Mg2+就可以满足蓝色的生成; 在以矢车菊素或牵牛花素(petunidin)为发色团的情况下, 必须有1/6Fe3+; 天竺葵素(pelargonidin)或锦葵素(malvidin)的发色团则无法形成蓝色的金属混合物. ...
Contribution of each caffeoyl residue of the pigment molecule of gentiodelphin to blue color development 3 2000
... 花青苷可通过酰基转移酶以物种特异方式进行酰基化修饰.花青苷根据酰化的数量可分为单酰或多酰花青苷(杨少勇等, 2003).由于蓝色花花青苷大多存在酰化现象, 所以花青苷的酰化一直被认为是蓝色花着色的重要步骤之一(Yoshida et al., 2009b).花青苷的酰化由BAHD和SCPL两个不同的蛋白家族控制.BAHD家族使用酰基-CoA作为酰基供体, 在细胞质中对花青苷进行酰化修饰; SCPL家族则以酰基-葡萄糖作为酰基供体, 在液泡中进行酰化修饰(Miyahara et al., 2013; Nishizaki et al., 2013; Sasaki et al., 2014).龙胆翠雀花素(gentiodelphin)来自蓝色龙胆(Gentiana makinoi), 是第1个被发表的完整多酰花青苷, 龙胆翠雀花素是观赏龙胆植物蓝色花瓣的主要决定因素.龙胆翠雀花素在3位、5位和3'位3个位点上糖基化, 之后通过5位与3'位的糖苷酰化形成飞燕草素3-O-葡糖基-5-O-咖啡酰-葡萄糖基-3'-O-咖啡酰-葡萄糖苷的结构(图2) (Yoshida et al., 2009b).Nakatsuka等(2010)通过RNAi技术沉默龙胆(Gentiana cv. ‘Albireo’)中负责3′与5位酰化的基因, 使植物只能产生飞燕草素3,5,3'-O-三葡萄糖苷的花青苷.酰化相关基因沉默植株的花色明显淡于野生型.这为酰化影响蓝色呈色提供了直接的体内证据.Ishii等(2017)通过对飞燕草品种的筛选得到AA7BG-GT1与AA7BG-GT2双敲除的突变体, 该突变体与野生型相比无法在7位的酰基上继续添加葡萄糖, 因此无法生成飞燕草苷(violdelphin), 其花色最终表现为淡粉色.
花青苷酰化结构(<xref ref-type="bibr" rid="b68">Yoshida et al., 2000</xref>; <xref ref-type="bibr" rid="b12">Boor, 2001</xref>)
Recent advances on the development and regulation of flower color in ornamental plants 3 2015
... 作为植物色素, 花青苷在植物的花瓣呈色中起重要作用, 控制着从粉色到蓝紫色一系列花色(Zhao and Tao, 2015).花青素分子在单体的情况下极不稳定, 易在中性或弱酸性介质中被水解失色.因此, 花青素生成的蓝色往往还需要通过糖基化形成花青苷, 之后再通过酰化、甲基化、金属离子络合、共染色和液泡pH值等条件来加强蓝色的稳定性(杨少勇等, 2003; Tanaka et al., 2009; Yoshida et al., 2009b). ...
... DFR是具有底物特异性的酶, 不同植物中DFR的特异性表现不同(Martens et al., 2002).康乃馨中DFR可以还原DHK、DHQ和DHM; 在矮牵牛(Petunia hybrida)中DFR只能还原DHQ和DHM (Zhao and Tao, 2015).Zhao等(2012)发现DFR的大量表达与花青苷的积累呈正相关. ...
Flower color diversity revealed by differential expression of flavonoid biosynthetic genes and flavonoid accumulation in herbaceous peony ( Paeonia lactiflora Pall.) 1 2012
... DFR是具有底物特异性的酶, 不同植物中DFR的特异性表现不同(Martens et al., 2002).康乃馨中DFR可以还原DHK、DHQ和DHM; 在矮牵牛(Petunia hybrida)中DFR只能还原DHQ和DHM (Zhao and Tao, 2015).Zhao等(2012)发现DFR的大量表达与花青苷的积累呈正相关. ...
 ,1,*
,1,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT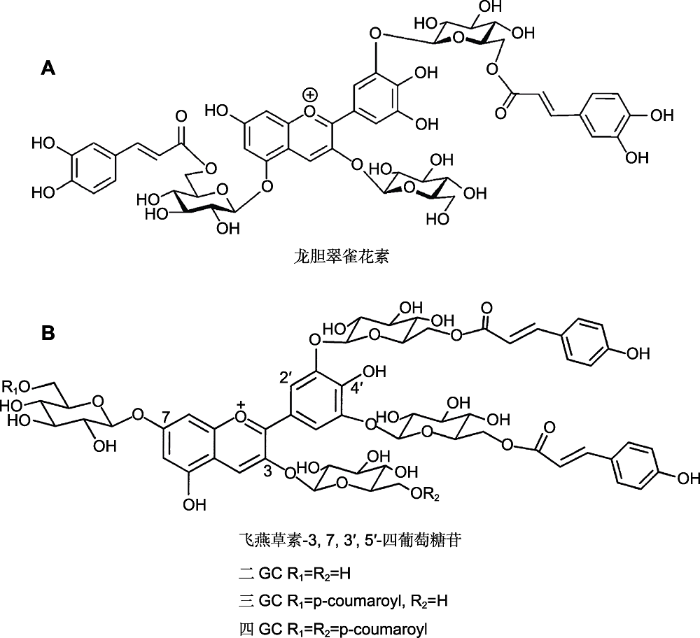 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT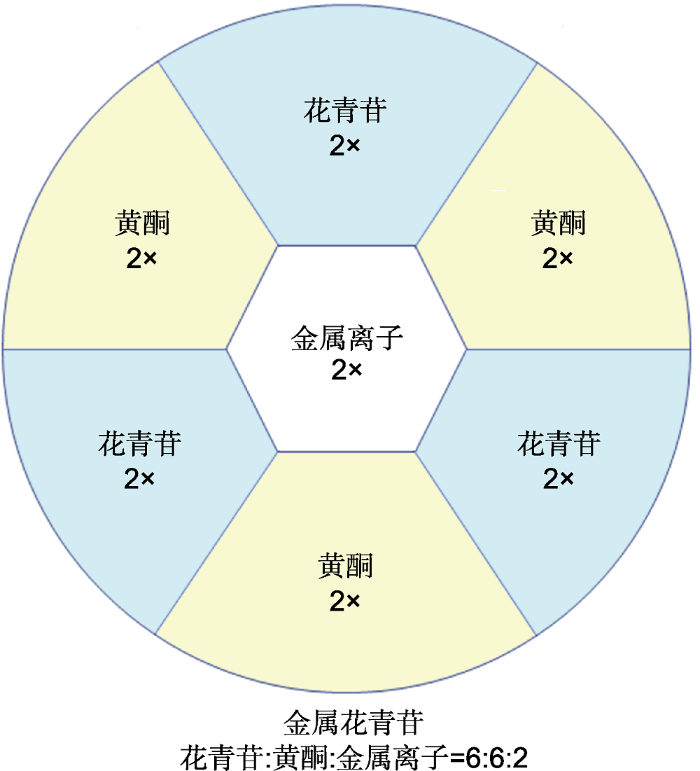 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT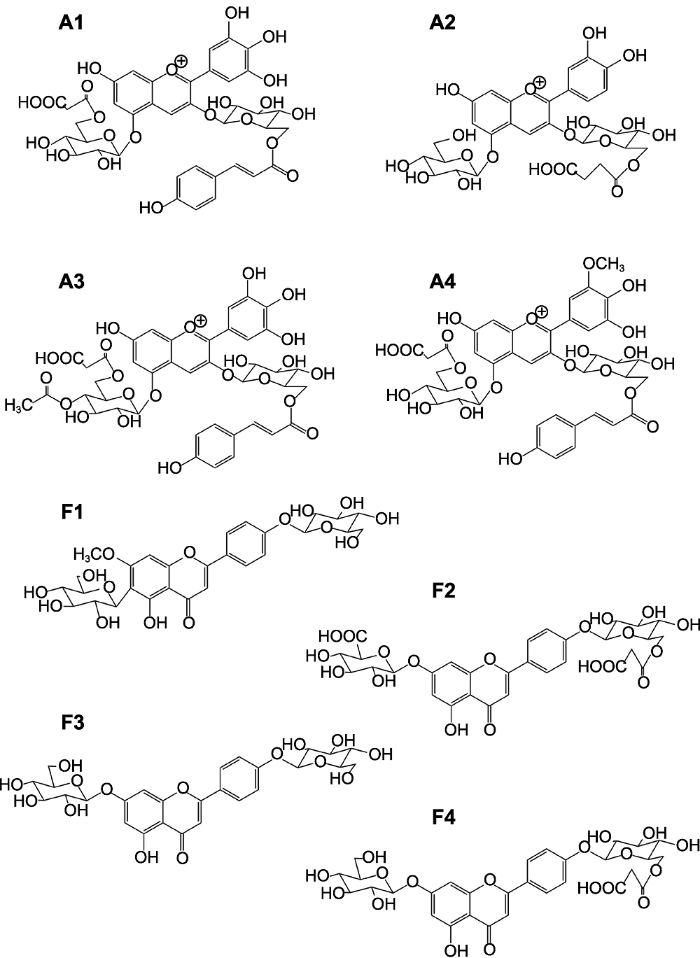 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT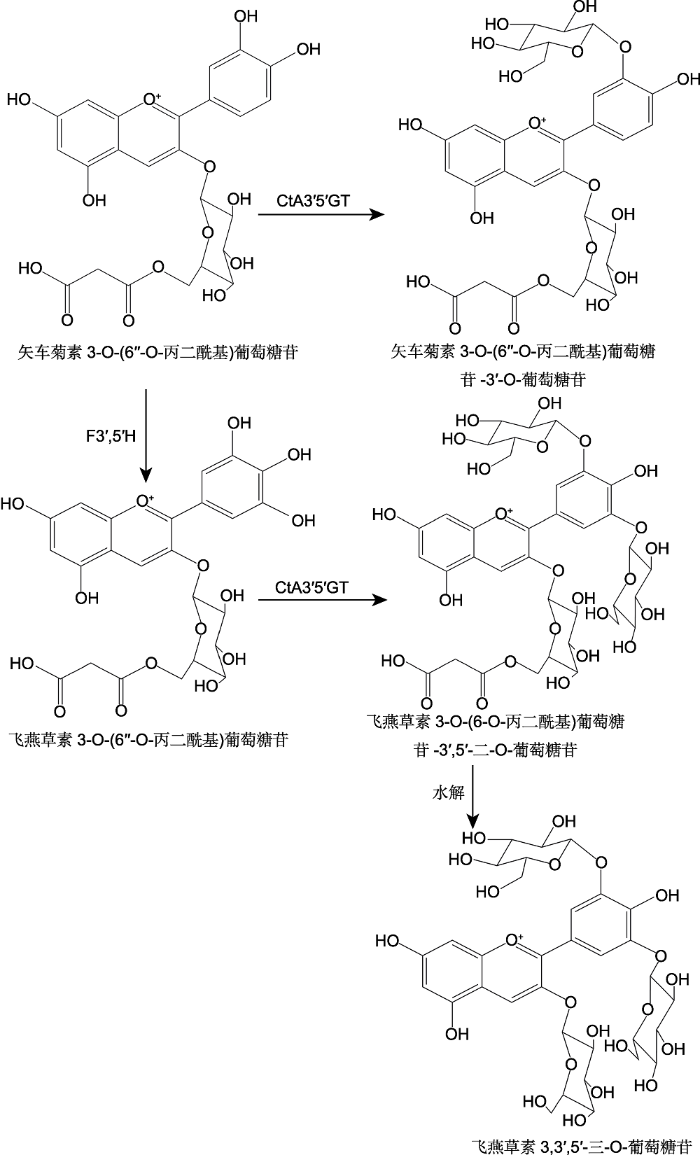 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}