 ,1,*, 王台,2,*1
,1,*, 王台,2,*1 2
Advances in the Regulatory Mechanisms of Pollen Response to Heat Stress in Crops
Hao Yang1, Chen Liu1, Zhifei Wang1, Xiuli Hu,1,*, Tai Wang,2,*1 2
通讯作者:
责任编辑: 朱亚娜
收稿日期:2018-06-10接受日期:2018-10-6网络出版日期:2019-03-10
| 基金资助: |
Corresponding authors:
Received:2018-06-10Accepted:2018-10-6Online:2019-03-10

摘要
关键词:
Abstract
Keywords:
PDF (1413KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
杨浩, 刘晨, 王志飞, 胡秀丽, 王台. 作物花粉高温应答机制研究进展. 植物学报, 2019, 54(2): 157-167 doi:10.11983/CBB18133
Yang Hao, Liu Chen, Wang Zhifei, Hu Xiuli, Wang Tai.
随着全球气候逐渐变暖, 温度变化对生态环境的影响日益明显(Hansen, 2015)。IPCC (Intergovern- mental Panel on Climate Change)研究报告指出, 相比20世纪末, 目前全球气温已经增加约4°C, 而温度升高将严重影响农作物生长, 威胁粮食安全(Lobell et al., 2011)。因此, 开展作物的高温胁迫研究具有重要意义。花粉作为一种具有特殊结构和功能的微小生命有机体, 包含父本基因组信息, 是种子植物有性生殖的重要调控者。由于花粉结构简单, 因此成为研究植物细胞极性、命运分化和发育的理想体系(鲁云龙等, 2014)。在生产实践中, 雄蕊产生有活性的花粉是作物形成种子和果实的前提。花粉发育过程对高温非常敏感, 高温导致花粉活性降低甚至失活, 使作物穗粒数和坐果率显著降低, 最终导致粮食减产。因此, 花粉高温应答机制研究成为植物学家关注的重点。
通常情况下, 在高于最适温度5°C以上的环境下, 花粉就会遭受高温伤害。许多重要作物如小麦(Tri- ticum aestivum)、水稻(Oryza sativa)和玉米(Zea mays), 其开花期均集中在夏季, 极易遭遇连续或严重的高温天气, 从而导致花粉发育异常(Barnabás et al., 2008)。水稻散粉时环境温度通常在28°C以上, 温度每升高1°C, 就会造成水稻减产约8%。如果温度升高到33°C, 会导致花粉产量降低到13%; 而温度升高到39°C, 就会导致花粉完全丧失活性, 落在柱头上的花粉无法萌发(Prasad et al., 2006; Endo et al., 2009)。高温对小麦影响更为严重, 在小孢子形成期, 环境温度达30°C时就会造成小孢子发育异常; 而在小花发育阶段, 长期温和高温(25°C, 白天)也会影响小麦的花粉发育(Farooq et al., 2011)。此外, 对玉米、高粱(Sorghum bicolor)和大麦(Hordeum vulgare)等的研究表明, 白天温度高于32°C会对花粉发育造成不同程度的影响(Lyakh et al., 1991; Liu et al., 2015)。
由于花粉取材相对困难, 因此针对花粉高温应答机制的研究相对较少。但随着分离技术的发展以及多种组学技术的应用, 人们对花粉的高温应答机制已有一定认识(Qin et al., 2008; Jagadish et al., 2010; Yang et al., 2015; González-Schain et al., 2016; Keller et al., 2017, 2018)。本文针对高温对作物花粉发育过程的影响、花粉高温胁迫应答机制, 以及花粉高温胁迫研究实验设计进行综述, 以期为相关研究提供借鉴。
1 高温对花粉发育的影响
花粉作为植物高度特化的雄配子体, 其发育过程既短暂又复杂(图1) (Twell, 2011)。位于花药内的花粉母细胞(pollen mother cells, PMC)经过1次减数分裂形成4个单倍体的小孢子(uninucleate microspore, UNM)。在中央大液泡的挤压下, 小孢子细胞核发生极化, 随后进行1次不对称有丝分裂(pollen mitosis I, PMI), 产生1个大的营养细胞(vegetative cell, VC)和1个位于营养细胞内的较小的生殖细胞(generative cell, GC)。最后, 生殖细胞再进行1次有丝分裂(pollen mitosis II, PMII)产生2个精细胞(sperm cell, SC)。精细胞携带父本信息, 在营养细胞产生的花粉管帮助下被运输到胚囊中, 分别与中央细胞和卵细胞融合完成双受精。图1
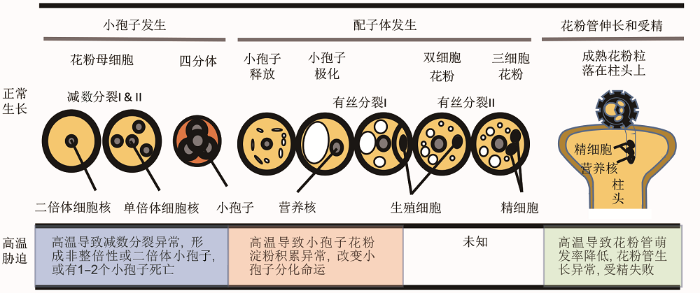 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1花粉发育过程及高温对花粉发育的影响
Figure 1Pollen development process and the influence of high temperature on pollen development
高温影响花粉发育全过程, 导致花粉结构和发育异常, 产生没有活性的花粉, 并最终导致受精失败(Porch and Jahn, 2001; Sato et al., 2006; Pecrix et al., 2011)。其中, 花粉母细胞减数分裂期到小孢子形成前期对高温最为敏感(图1)。高温影响花粉母细胞减数分裂进程, 造成同源染色体交叉和重组频率增加、染色体分离异常, 以及形成不完整的四分体和染色体倍性异常的小孢子等(Francis et al., 2007; De Storme and Geelen, 2014; Omidi et al., 2014; Draeger and Moore, 2017)。对大麦的研究表明, 高温诱导小孢子母细胞中减数分裂特异基因ASY1 (ASYNAPSIS1)上调表达, 从而提高染色体交叉和重组频率(Oshino et al., 2007)。染色体倍性异常的小孢子, 如二价体(diploid dyads)的产生则是由于高温导致维管骨架解聚以及纺锤体错误排列引起的(Francis et al., 2007; Pecrix et al., 2011; Wang et al., 2017)。已在拟南芥(Arabidopsis thaliana)中鉴定到3个调节减数分裂期纺锤丝定位的基因(FORMIN14、JASON和PARALLEL SPINDLES 1), 推测高温影响这3个基因的表达, 导致产生异常倍性的小孢子(De Storme and Geelen, 2014)。FORMIN14属于植物第二类形成素, 通过调节微管和微丝骨架的形成和交联参与花粉母细胞减数分裂过程(Li et al., 2010b)。jason和parallel spindles 1产生二倍体小孢子是由于减数分裂II期纺锤体呈异常的平行排列, 并且PARALLEL SPINDLES 1的表达受到JASON调控(d'Erfurth et al., 2008; De Storme and Geelen, 2011)。这种由于高温引起小孢子母细胞减数分裂异常, 进而导致花粉多倍体化(polyploidization)的现象对植物进化具有重要意义(Pecrix et al., 2011)。
高温还影响小孢子的发育。研究表明, 蔗糖转化酶(invertases, INVs)在高温胁迫下小孢子发育过程中发挥重要作用。高温导致小孢子和花药中INVs基因表达量下降, 从而影响蔗糖水解过程, 造成小孢子中淀粉积累异常, 产生没有活性的花粉(Jain et al., 2010)。在体外对分离的小孢子进行高温处理有助于诱导其形成胚胎, 表明高温还影响小孢子细胞的分化(Ochatt et al., 2009)。
高温通过改变花粉和柱头细胞中生长素和糖类的平衡影响花粉管在柱头内的生长, 表现出花粉萌发失败以及花粉管末端膨大、扭曲和破裂等现象, 最终导致受精过程无法完成(Kakani et al., 2005; Snider et al., 2011; Snider and Oosterhuis, 2011; Song et al., 2015; Coast et al., 2016; Parrotta et al., 2016; Shi et al., 2018; Zhang et al., 2018)。对拟南芥的研究表明, 高温下CLE45-SKM1/SKM2信号通路在确保花粉和柱头正常相互识别过程中发挥重要作用(Endo et al., 2013)。
高温还可以促进绒毡层细胞提前发生程序性死亡(programmed cell death, PCD), 进而影响花粉发育、活性以及散粉过程(Sato et al., 2002; Parish et al., 2012; De Storme and Geelen, 2014)。绒毡层细胞提前发生程序性死亡, 是由于高温导致更多的蛋白质折叠异常, 超出内质网相关降解机制(ER-associa- ted degradation machinery, ERAD)和未折叠蛋白响应机制(unfolded protein response, UPR)的修复能力(Li et al., 2006)。此外, 绒毡层分泌的β-1,3-葡聚糖酶对花粉细胞壁形成以及小孢子发育至关重要(Su- zuki et al., 2001)。在水稻小孢子形成期, YY1和YY2等绒毡层特异表达的基因在高温胁迫下显著下调, 可影响花粉在柱头上的吸附性, 并最终导致萌发率降低(Endo et al., 2009; Müller and Rieu, 2016)。
2 花粉对高温胁迫的应答机制
2.1 花粉的高温胁迫感应
植物进化出完整而复杂的胁迫响应机制来适应高温并维持自身的生长发育(Kotak et al., 2007)。已有研究表明, 花粉至少可以通过4种方式感知环境温度的变化, 并通过诱导高温胁迫相关基因的表达来适应高温环境(Mittler et al., 2012)。这4种方式分别为质膜上的钙离子通道、未折叠蛋白反应(unfolded protein response, UPR)、积累的活性氧(reactive oxygen species, ROS)以及含有组蛋白变体H2A.Z的核小体(Kumar and Wigge, 2010)。质膜上的钙离子通道是植物细胞重要的高温感受器。电化学和生化等实验证实, 温度升高可以触发质膜上钙离子通道打开, 导致胞外钙离子内流, 使胞质内钙离子增加, 引起细胞产生热激反应(heat shock response, HSR) (Saidi et al., 2009)。拟南芥基因组编码40多个钙离子通道, 且大多数位于质膜上(作为高温感受器), 可以与钙调素蛋白AtCaM3结合, 通过激活WRKY39等因子参与热激反应(Ward et al., 2009; Zhang et al., 2009; Li et al., 2010a)。其中, 一类环腺苷酸门控离子通道(cyclic nucleotide gate channels, CNGCs)被证实参与拟南芥花粉的高温感应。环境温度升高后, 花粉中的cAMP含量升高并导致CNGC16通道打开, 使得胞外的Ca2+内流, 而胞内积累的Ca2+又可以进一步激活热激转录因子(heat shock transcription factors, HSFs) HsfA2和HsfB1, 从而参与高温胁迫应答(Gao et al., 2012; Tunc- Ozdemir et al., 2013)。最近, 研究者通过分析高温胁迫下cngc16突变体与野生型花粉的基因表达情况, 鉴定到至少有27个转录因子参与花粉的高温响应; 此外, 还鉴定到cngc16和野生型在高温胁迫下存在192个差异表达基因, 其中突变体中的差异表达基因更多参与细胞生长和花粉发育过程, 并且缺少在野生型中表达的转录因子AGAMOUS-like 29, 暗示CN- GC16在调控花粉的高温胁迫响应中具有一定的特异性(Rahmati Ishka et al., 2018)。
绒毡层或者花粉细胞内的UPR反应也可以感受高温胁迫, 并通过ER-UPR (内质网中未折叠蛋白反应)提高内质网对蛋白质的折叠能力, 降低未折叠蛋白质的数量, 从而提高花粉的胁迫耐受能力。一些转录因子和RNA剪切因子也参与ER-UPR, 并在花粉发育过程中发挥重要作用, 其中erdj2a、bip1/2/3和pod1等突变体均表现出花粉发育异常表型(Fragkos- tefanakis et al., 2016a)。而ER-UPR反应的重要组分IRE1 (Inositol Requiring Enzyme 1)可以利用其核糖核酸酶活性对bZIP60的mRNA进行剪切, 进而参与花粉的高温胁迫响应(Deng et al., 2016)。IRE1还可以影响HOP3 (HSP70-HSP90 organizing protein)蛋白的表达水平, 而后者通过与BiP (binding immu- noglobulin protein)及TMS1 (THERMOSENSITIVE MALE STERILE 1)互作, 参与调控花粉在柱头上的萌发(Yang et al., 2009; Ma et al., 2015; Fernández- Bautista et al., 2017)。
ROS是一类化学性质活泼且具有较高氧化活性的分子, 高温胁迫会导致植物体内产生大量ROS。积累的ROS不仅会造成核酸和蛋白质等重要生物大分子的氧化损伤, 也可以作为信号分子激活植物体内的抗氧化系统, 从而调控植物多个器官(包括花粉)响应高温胁迫。对小麦花粉进行高温处理(42°C, 2小时)后, 发现花粉中ROS和超氧化物歧化酶(superoxide dismutase, SOD)、过氧化物酶(peroxidase, POD)和过氧化氢酶(catalase, CAT)含量显著升高, 同时伴随着热激蛋白(heat shock proteins, HSPs) HSP70、HSP90、HSP17和HSP26含量增加(Kumar et al., 2013)。番茄(Lycopersicon esculentum)和水稻花粉中也有类似机制(Frank et al., 2009; Zhao et al., 2018a)。水稻花粉中具有ROS清除能力的MT-1-4B蛋白表达量下调会导致花粉丧失育性, 进一步证明ROS参与花粉的高温胁迫应答(Hu et al., 2011)。
H2A.Z是一种组蛋白变体, 包含H2A.Z的核小体与DNA结合更加紧密。研究表明, 高温处理可以减少含有H2A.Z的核小体在基因转录起始位点的占有率, 从而调控相关基因的表达(Kumar and Wigge, 2010)。但目前关于H2A.Z参与花粉高温胁迫响应的研究非常有限。在二穗短柄草(Brachypodium distachyum)中, 含有H2A.Z的核小体参与开花和种子形成期的高温感应, 温度过高会破坏H2A.Z在染色体上的分布规律, 最终导致二穗短柄草的结实率下降(Boden et al., 2013)。
2.2 花粉的高温胁迫应答
花粉感知环境中的高温信号后, 通过信号转导引起代谢、转录甚至表观遗传水平的改变, 从而实现对高温胁迫的响应。其中热激转录因子是高温胁迫应答网络中的关键组分, 可以诱导高温胁迫应答重要效应物热激蛋白的表达, 通过维持蛋白的构型稳定减少由高温导致的蛋白变性和聚集, 从而提高植物耐热性。在烟草(Nicotiana tabacum)花粉发育过程中, 高温诱导不同的HSPs表达(Volkov et al., 2005)。在番茄等植物的花粉中, 高温可以诱导HSFA2、HSFA3、HSP100和HSP17等热激转录因子和热激蛋白基因的表达(Fragkostefanakis et al., 2016b; Kumar et al., 2016)。在番茄花粉发育过程中, 抑制HSFA2的表达导致花粉小孢子发育异常(Frank et al., 2009; Giorno et al., 2010; Fragkostefanakis et al., 2016b)。在拟南芥花粉中, 高温可以诱导HSFA2、HSFB1和HSFA7a基因表达, 协同参与高温胁迫响应(Larkindale and Vierling, 2008; Gao et al., 2012)。此外, HSFA5同源基因AtREN1突变体的花粉对高温非常敏感(Reňák et al., 2014)。HSP除了受到转录因子HSF的影响, 可变剪切也可以通过调控HSPs基因的表达及多样性参与高温胁迫响应(Keller et al., 2017)。与营养器官相比, 高温诱导花粉中产生的HSFs和HSPs种类和数量仍然很少, 这也是花粉对高温更加敏感的原因之一。在拟南芥花粉中表达体细胞特异的AtHSP101可以有效提高花粉对高温的耐受能力(Burke and Chen, 2015)。活性氧清除系统、激素以及糖类物质在花粉的高温应答中也发挥重要作用(Frank et al., 2009; Min et al., 2014)。花粉和绒毡层细胞含有大量线粒体, 高出营养器官20倍以上。在高温条件下, 线粒体会产生大量的ROS, 从而给花粉带来严重的活性氧压力。这些ROS可以被SOD、CAT、抗坏血酸过氧化物酶(ascorbate peroxidase, APX)和谷胱甘肽过氧化物酶(glutathione peroxidase, GPX)等抗氧化系统清除(Kumar et al., 2014; Müller and Rieu, 2016; Zhao et al., 2018a, 2018b)。
高温胁迫下, 植物体内的激素水平也会发生显著变化(Verma et al., 2016)。番茄花粉中乙烯合成基因(SlACS3和SlACS11)及其信号通路中的相关基因(SlETR3和SlCTR2)受高温诱导表达(Jegadeesan et al., 2018)。对乙烯不敏感的番茄突变体花粉对高温非常敏感, 但在高温处理前, 外源施加乙烯可以显著提高花粉的耐热性(Firon et al., 2012), 表明乙烯参与花粉的高温胁迫响应。此外, 已有研究表明, 生长素参与调控花粉母细胞的减数分裂和小孢子的有丝分裂进程。高温处理拟南芥和大麦花粉会降低花粉和绒毡层细胞中生长素合成基因(YUC2、YUC6和TAA1/TIR2)的表达, 导致小孢子有丝分裂异常, 产生败育的花粉, 而外源施加生长素则可提高花粉的耐热性(Sakata et al., 2010; Firon et al., 2012; Higa- shitani, 2013)。研究表明, 在棉花(Gossypium hirsutum)中, microRNA (miR157和miR160等)等可以通过调节生长素信号通路影响高温胁迫下的花粉活性(Ding et al., 2017)。研究还显示, 赤霉素(GA)和脱落酸(ABA)也参与花粉热胁迫响应(Sakata et al., 2014)。ABA作为重要的植物胁迫响应激素, 参与植物的多种非生物胁迫响应(Tang et al., 2008)。越来越多的证据表明, ABA可以通过与糖信号通路互作(如在胁迫下抑制INV活性)影响植物的胁迫响应(Oliver et al., 2007)。在小孢子母细胞减数分裂时期, 用外源ABA喷施水稻叶片可以提高蔗糖的转运效率并加速糖类代谢, 从而增强花粉对高温的耐受能力(Rezaul et al., 2018)。
花粉作为有性生殖过程中光合产物重要的“库”, 其淀粉含量通常在二细胞花粉期达到最高值, 而可溶性糖的含量在散粉期达到最高值。积累的糖类物质不仅为花粉发育和花粉管形成提供能量, 也可以作为渗透物质提高花粉对胁迫的响应。通过比较不同的番茄品种可以发现, 耐高温品种的花粉中含有更多的淀粉和可溶性糖(Pressman et al., 2002; Firon et al., 2006), 而其它渗透调节物质(如甜菜碱和脯氨酸)以及次生代谢产物等也会影响花粉对高温胁迫的应答(Sangu et al., 2015; Qi et al., 2018)。
综上, 在应对高温胁迫时, 花粉至少可以通过4种机制感受高温信号并利用多种机制响应胁迫, 这些机制和途径之间也存在“交叉对话”。例如, 内流的钙离子可以参与多个途径; ROS可以影响UPR; ABA则影响糖类代谢; 而糖信号通路又可以影响ROS的平衡。这些机制之间的相互作用也增加了花粉高温胁迫研究的复杂性(图2) (Couée et al., 2006; Kotak et al., 2007)。
图2
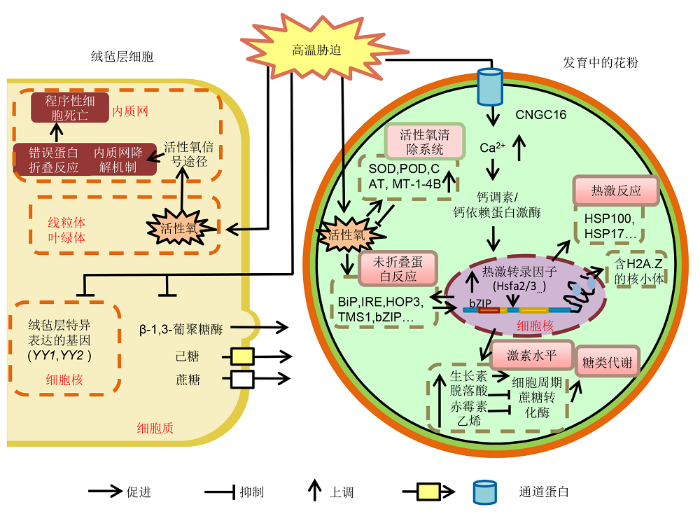 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2花粉的高温应答机制
Figure 2The mechanism of pollen response to high temperature stress
3 花粉高温胁迫研究的实验设计
在进行花粉高温胁迫研究时, 需要考虑胁迫温度、处理方式以及花粉取材时期。首先需要选择合适的温度处理范围, 不同的温度处理会产生不同的结果。温和高温可以使植物获得高温耐受能力, 严重高温则直接破坏植物细胞, 导致植株不可逆损伤甚至死亡。通过总结常见作物的花粉高温胁迫研究, 发现常见作物的高温胁迫温度通常选择30-39°C。尽管番茄花粉高温胁迫研究也有选择45°C高温的情况, 但处理时间相对较短(Zinn et al., 2010; Fragkostefanakis et al., 2015; Mesihovic et al., 2016; Arshad et al., 2017)。其次, 还需考虑胁迫施加的方式(Mesihovic et al., 2016)。对植物直接施加高温(direct applied heat stress, DAHS), 主要用来研究植物的基础高温胁迫耐受性(basal thermo tolerance, BTT)。如果在对植物施加高温处理前先预施加一个温和高温(pre-induction heat stress, PIHS), 则主要用于研究植物获得性高温胁迫耐受性(acquired thermotolerance, ATT)。例如, 在番茄花粉的ATT研究中, 可以将花粉32°C预处理1小时, 然后置于25°C恢复1小时, 最后再于50°C的高温条件下进行处理。也可以利用温和高温(30- 36°C)对植物处理几周甚至几个月, 这种处理方式被称为温和的慢性高温处理(mild chronic heat stress, MCHS), 可以引起植物产生温和高温胁迫耐受性(mild heat thermotolerance, MHTT)。由于MCHS处理方式可以更好地模拟大气环境中温度的变化并覆盖花粉整个发育阶段, 更有利于人们对花粉发育的各个时期的发育状态和生理指标进行监测, 因此已成为花粉高温处理的主要方式(Mesihovic et al., 2016)。在设计花粉高温胁迫相关实验时, 还需要考虑花粉所处的发育阶段。不同发育阶段的花粉对高温胁迫的敏感性和响应机制可能不同。在实际研究中, 通常把花粉发育过程分为4个阶段: (1) 花粉母细胞到四分体时期; (2) 小孢子形成到早期的二细胞时期; (3) 二细胞末期/三细胞期到成熟花粉期; (4) 花粉萌发期。目前, 可以通过一些形态指标对常见作物花粉发育阶段进行判断。例如, 水稻花粉的发育阶段可以通过花芽长度进行判断; 而玉米花粉的发育阶段则可以通过茎秆的叶领(leaf collar)数进行判断(Zhang et al., 2011; Begcy and Dresselhaus, 2017)。只有选择合适的花粉发育时期, 采用合适的高温处理方式, 才可以得到更加准确的实验结果。
4 总结与展望
研究作物花粉高温胁迫应答机制并提高作物花粉的高温耐受能力, 对于稳定和提高作物产量具有重要意义。目前, 人们已对高温导致的花粉败育进行了系统的形态学观察和多种组学分析, 并且已鉴定到一些参与花粉高温胁迫响应的因子, 但对于花粉高温胁迫应答调控机制的认知仍然有限(De Storme and Geelen, 2014)。已知钙离子信号通路、激素水平、糖代谢以及活性氧压力都参与花粉的高温胁迫应答, 并且它们之间存在相互作用。此外, 表观遗传机制在植物高温胁迫响应过程中也发挥重要作用(Chen et al., 2016)。研究表明, 高温胁迫可以引起花粉细胞热激蛋白编码基因甲基化水平的改变(Solís et al., 2012); 染色质构型和各种sRNA的表达均参与花粉的高温胁迫响应(Pecinka and Scheid, 2012; Bokszczanin et al., 2015; Chen et al., 2016)。但是对于表观机制具体如何调控花粉的高温胁迫响应, 以及花粉是否可以将遭受到的高温胁迫“表观印迹”传递到子代, 目前仍有争议, 需要深入研究(Lang-Mladek et al., 2010; Migi- covsky et al., 2014)。尽管目前对花粉高温胁迫响应机制认识有限, 但是随着各种组学技术、全基因组关联分析(genome- wide association study, GWAS)和CRISPR-Cas9等基因编辑技术在花粉高温胁迫研究中的应用(Paupière et al., 2014; Xing et al., 2014; Chaturvedi et al., 2015; Kim et al., 2015; Xu et al., 2017), 未来研究者们可以更好、更全面地解析高温胁迫下花粉生理生化及分子水平的变化, 并从中选择重要的调节基因进行功能研究, 最终为作物耐高温分子育种提供指导。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.3724/SP.J.1259.2014.00229URL [本文引用: 1]

开花植物精细胞的发育经历一个独特的后减数分裂过程,在此过程中每个花粉母细胞减数分裂的产物——小孢子经不对称有丝分裂产生1个大的营养细胞和1个小的生殖细胞,随后生殖细胞经过正常的有丝分裂产生2个精细胞。近几年,随着高通量组学技术的不断完善,利用组学技术比较分析生殖细胞和精细胞的分子特征、揭示决定精细胞命运与功能以及受精识别的重要分子已成为植物生殖生物学备受关注的课题。开展此项研究的关键是建立能获得大量高纯度的生殖细胞与精细胞分离纯化技术。该文综述了被子植物生殖细胞和精细胞分离方法的主要研究进展,分析了关键方法的特点和要点以及不同方法之间的差异和共性,以期为相关领域的研究人员提供借鉴。
DOI:10.1016/j.plaphy.2017.03.011URL [本文引用: 1]
DOI:10.1111/j.1365-3040.2007.01727.xURLPMID:17971069 [本文引用: 1]
Wheat plants of the drought tolerant Plainsman V and sensitive Cappelle Desprez genotypes were subjected to drought and combined drought and elevated, 34/24 潞C day/night temperature at three various phases of reproductive development, at meiosis, from the 1 st to the 5 th day after pollination and from the 5 th to the 9 th day of seed development. After seed maturation, the germination... [Show full abstract]
DOI:10.1007/s00497-017-0311-4URLPMID:29101473 [本文引用: 1]
An easy and highly reproducible nondestructive method named the Leaf Collar Method is described to identify and characterize the different stages of pollen development in maize. In plants, many cellular events such as meiosis, asymmetric cell division, cell cycle regulation, cell fate determination, nucleus movement, vacuole formation, chromatin condensation and epigenetic modifications take place during pollen development. In maize, pollen development occurs in tassels that are confined within the internal stalk of the plant. Hence, identification of the different pollen developmental stages as a tool to investigate above biological processes is impossible without dissecting the entire plant. Therefore, an efficient and reproducible method is necessary to isolate homogeneous cell populations at individual stages throughout pollen development without destroying the plant. Here, we describe a method to identify the various stages of pollen development in maize. Using theLeaf Collar Methodin the maize inbreed line B73, we have determined the duration of each stage from pollen mother cells before meiosis to mature tricellular pollen. Anther and tassel size as well as percentage of pollen stages were correlated with vegetative stages, which are easily recognized. The identification of stage-specific genes indicates the reproducibility of the method. In summary, we present an easy and highly reproducible nondestructive method to identify and characterize the different stages of pollen development in maize. This method now opens the way for many subsequent physiological, morphological and molecular analyses to study, for instance, transcriptomics, metabolomics, DNA methylation and chromatin patterns during normal and stressful conditions throughout pollen development in one of the economically most important grass species. The online version of this article (10.1007/s00497-017-0311-4) contains supplementary material, which is available to authorized users.
.
[本文引用: 1]
DOI:10.1186/s12864-015-1901-xURLPMID:4575465 [本文引用: 1]
The unprecedented role of sncRNAs in the regulation of pollen biogenesis on both transcriptional and epigenetic levels has been experimentally proven. However, little is known about their global regulation, especially under stress conditions. We used tomato pollen in order to identify pollen stage-specific sncRNAs and their target mRNAs. We further deployed elevated temperatures to discern stress responsive sncRNAs. For this purpose high throughput sncRNA-sequencing as well as Massive Analysis of cDNA Ends (MACE) were performed for three-replicated sncRNAs libraries derived from tomato tetrad, post-meiotic, and mature pollen under control and heat stress conditions. Using the omiRas analysis pipeline we identified known and predicted novel miRNAs as well as sncRNAs from other classes, responsive or not to heat. Differential expression analysis revealed that post-meiotic and mature pollen react most strongly by regulation of the expression of coding and non-coding genomic regions in response to heat. To gain insight to the function of these miRNAs, we predicted targets and annotated them to Gene Ontology terms. This approach revealed that most of them belong to protein binding, transcription, and Serine/Threonine kinase activity GO categories. Beside miRNAs, we observed differential expression of both tRNAs and snoRNAs in tetrad, post-meiotic, and mature pollen when comparing normal and heat stress conditions. Thus, we describe a global spectrum of sncRNAs expressed in pollen as well as unveiled those which are regulated at specific time-points during pollen biogenesis. We integrated the small RNAs into the regulatory network of tomato heat stress response in pollen. The online version of this article (doi:10.1186/s12864-015-1901-x) contains supplementary material, which is available to authorized users.
DOI:10.1371/journal.pone.0122933URLPMID:4388472 [本文引用: 1]
Comparison of average crop yields with reported record yields has shown that major crops exhibit annual average yields three- to seven-fold lower than record yields because of unfavorable environments. The current study investigated the enhancement of pollen heat tolerance through expressing an Arabidopsis thaliana heat shock protein 101 (AtHSP101) that is not normally expressed in pollen but reported to play a crucial role in vegetative thermotolerance. The AtHSP101 construct under the control of the constitutive ocs/mas 'superpromoter' was transformed into cotton Coker 312 and tobacco SRI lines via Agrobacterium mediated transformation. Thermotolerance of pollen was evaluated by in vitro pollen germination studies. Comparing with those of wild type and transgenic null lines, pollen from AtHSP101 transgenic tobacco and cotton lines exhibited significantly higher germination rate and much greater pollen tube elongation under elevated temperatures or after a heat exposure. In addition, significant increases in boll set and seed numbers were also observed in transgenic cotton lines exposed to elevated day and night temperatures in both greenhouse and field studies. The results of this study suggest that enhancing heat tolerance of reproductive tissues in plant holds promise in the development of crops with improved yield production and yield sustainability in unfavorable environments.
DOI:10.1021/pr501240nURLPMID:26419256 [本文引用: 1]
Recently, we have developed a quantitative shotgun proteomics strategy called mass accuracy precursor alignment (MAPA). The MAPA algorithm uses high mass accuracy to bin mass-to-charge (m/z) ratios of precursor ions from LC-MS analyses, determines their intensities, and extracts a quantitative sample versus m/z ratio data alignment matrix from a multitude of samples. Here, we introduce a novel feature of this algorithm that allows the extraction and alignment of proteotypic peptide precursor ions or any other target peptide from complex shotgun proteomics data for accurate quantification of unique proteins. This strategy circumvents the problem of confusing the quantification of proteins due to indistinguishable protein isoforms by a typical shotgun proteomics approach. We applied this strategy to a comparison of control and heat-treated tomato pollen grains at two developmental stages, post-meiotic and mature. Pollen is a temperature-sensitive tissue involved in the reproductive cycle of plants and plays a major role in fruit setting and yield. By LC-MS-based shotgun proteomics, we identified more than 2000 proteins in total for all different tissues. By applying the targeted MAPA data-processing strategy, 51 unique proteins were identified as heat-treatment-responsive protein candidates. The potential function of the identified candidates in a specific developmental stage is discussed.
DOI:10.1007/s00497-015-0271-5URL [本文引用: 2]
DOI:10.1111/pce.12475URL [本文引用: 1]
Resilience of rice cropping systems to potential global climate change will partly depend on temperature tolerance of pollen germination (PG) and tube growth (PTG). Germination of pollen of high temperature susceptible Oryza glaberrima Steud. (cv. CG14) and O. sativa L. ssp. indica (cv. IR64) and high temperature tolerant O. sativa ssp. aus (cv. N22), was assessed on a 5.6‐45.4°C temperature gradient system. Mean maximum PG was 85% at 27°C with 1488 μm PTG at 25°C. The hypothesis that in each pollen grain, minimum temperature requirements (T) and maximum temperature limits (T) for germination operate independently was accepted by comparing multiplicative and subtractive probability models. The maximum temperature limit for PG in 50% of grains (T) was lowest (29.8°C) in IR64 compared with CG14 (34.3°C) and N22 (35.6°C). Standard deviation (s) of Twas also low in IR64 (2.3°C) suggesting that the mechanism of IR64's susceptibility to high temperatures may relate to PG. Optimum germination temperatures and thermal times for 1mm PTG were not linked to tolerating high temperatures at anthesis. However, the parameters Tand sin the germination model define new pragmatic criteria for successful and resilient PG, preferable to the more traditional cardinal (maximum and minimum) temperatures.
DOI:10.1093/jxb/erj027URLPMID:16397003 [本文引用: 1]
Soluble sugars, especially sucrose, glucose, and fructose, play an obviously central role in plant structure and metabolism at the cellular and whole-organism levels. They are involved in the responses to a number of stresses, and they act as nutrient and metabolite signalling molecules that activate specific or hormone-crosstalk transduction pathways, thus resulting in important modifications of gene expression and proteomic patterns. Various metabolic reactions and regulations directly link soluble sugars with the production rates of reactive oxygen species, such as mitochondrial respiration or photosynthesis regulation, and, conversely, with anti-oxidative processes, such as the oxidative pentose-phosphate pathway and carotenoid biosynthesis. Moreover, stress situations where soluble sugars are involved, such as chilling, herbicide injury, or pathogen attack, are related to important changes in reactive oxygen species balance. These converging or antagonistic relationships between soluble sugars, reactive oxygen species production, and anti-oxidant processes are generally confirmed by current transcriptome analyses, and suggest that sugar signalling and sugar-modulated gene expression are related to the control of oxidative stress. All these links place soluble carbohydrates in a pivotal role in the pro-oxidant and antioxidant balance, and must have constrained the selection of adaptive mechanisms involving soluble sugars and preventing de-regulation of reactive oxygen species production. Finally, in line with the specific role of sucrose in oxygenic photosynthetic organisms, this role of soluble sugars in oxidative stress regulation seems to entail differential effects of glucose and sucrose, which emphasizes the unresolved issue of characterizing sucrose-specific signalling pathways.
DOI:10.1104/pp.110.170415URLPMID:21257792 [本文引用: 1]
Abstract In plants, whole-genome doubling (polyploidization) is a widely occurring process largely contributing to plant evolution and diversification. The generation and fusion of diploid gametes is now considered the major route of plant polyploidization. The parallel arrangement or fusion of meiosis II MII spindles (ps) is one of the most frequently reported mechanisms generating triploid offspring. Through a forward genetics screen of an Arabidopsis (Arabidopsis thaliana) ethyl methanesulfonate population, we identified Arabidopsis thaliana Parallel Spindles1 (AtPS1), which was recently reported as a major gene implicated in the control of the ps meiotic defect. In addition, we describe the isolation and characterization of a novel allele of JASON, involved in male gametophytic ploidy regulation in plants. Similar to atps1 mutants, jason produces more than 25% 2n pollen grains and spontaneously forms triploid offspring. By combining both cytological and genetic approaches, we demonstrate that loss of JASON causes the formation of parallel arranged and fused spindles in male MII, resulting in the production of unreduced first division restitution 2n spores. Although JASON encodes a protein of unknown function, we additionally show that the meiotic ps defect in jason is caused by a reduction in AtPS1 transcript levels, indicating that JASON positively regulates AtPS1 expression, allowing the proper organization and orientation of metaphase II spindle plates in MII.
DOI:10.1111/pce.12142URLPMID:23731015 [本文引用: 4]
In plants, male reproductive development is extremely sensitive to adverse climatic environments and (a)biotic stress. Upon exposure to stress, male gametophytic organs often show morphological, structural and metabolic alterations that typically lead to meiotic defects or premature spore abortion and male reproductive sterility. Depending on the type of stress involved (e.g. heat, cold, drought) and the duration of stress exposure, the underlying cellular defect is highly variable and either involves cytoskeletal alterations, tapetal irregularities, altered sugar utilization, aberrations in auxin metabolism, accumulation of reactive oxygen species (ROS; oxidative stress) or the ectopic induction of programmed cell death (PCD). In this review, we present the critically stress-sensitive stages of male sporogenesis (meiosis) and male gametogenesis (microspore development), and discuss the corresponding biological processes involved and the resulting alterations in male reproduction. In addition, this review also provides insights into the molecular and/or hormonal regulation of the environmental stress sensitivity of male reproduction and outlines putative interaction(s) between the different processes involved.
DOI:10.1111/tpj.13239URLPMID:27304577 [本文引用: 1]
Summary The unfolded protein response (UPR) is activated by various stresses during vegetative development in Arabidopsis, but is constitutively active in anthers of unstressed plants. To understand the role of the UPR during reproductive development, we analyzed a double mutant, ire1a ire1b . The double mutant knocks out the RNA-splicing arm of the UPR signaling pathway. It is fertile at room temperature but male sterile at modestly elevated temperature (ET). The conditional male sterility in the mutant is a sporophytic trait, and when the double mutant was grown at ET, defects appeared in the structure of the tapetum. As a result, the tapetum in the double mutant failed to properly deposit the pollen coat at ET, which made pollen grains clump and prevented their normal dispersal. IRE1 is a dual protein kinase/ribonuclease involved in the splicing of bZIP60 mRNA, and through complementation analysis of various mutant forms of IRE1b it was demonstrated that the ribonuclease activity of IRE1 was required for protecting male fertility from ET. It was also found that overexpression of SEC31A rescued the conditional male sterility in the double mutant. SEC31A is involved in trafficking from the endoplasmic reticulum to Golgi and a major target of the IRE1-mediated UPR signaling in stressed seedlings. Thus, IRE1, a major component of the UPR, plays an important role in protecting pollen development from ET.
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00122-017-2925-1URLPMID:28550436 [本文引用: 1]
Exposure of wheat to high temperatures during male meiosis prevents normal meiotic progression and reduces grain number. We define a temperature-sensitive period and link heat tolerance to chromosome
DOI:10.1093/pcp/pcp135URLPMID:19808807 [本文引用: 2]
Abstract Plant male reproductive development is highly organized and sensitive to various environmental stressors, including high temperature. We have established an experimental procedure to evaluate high temperature injury in japonica rice plants. High temperature treatment (39 degrees C/30 degrees C) starting at the microspore stage repeatedly reduced spikelet fertility in our system. Morphological observations revealed that pollen viability in plants exposed to high temperatures was lower than that in control plants. Most pollen grains in high temperature-treated plants displayed a normal round shape and stained reddish purple with Alexander's reagent; however, the pollen grains were very poorly attached and displayed limited germination on the stigma. To investigate gene regulatory mechanisms in the anther in high temperature environments, DNA microarray analysis was performed by comparing non-treated samples with samples treated with 2-4 d of high heat. Genes responsive to high temperatures were identified from clustering of microarray data. Among these, at least 13 were designated as high temperature-repressed genes in the anther. Expression analyses revealed that these genes were expressed specifically in the immature anther mainly in the tapetum at the microspore stage and down-regulated after 1 d of high temperature. The expression levels of Osc6, OsRAFTIN and TDR, which are tapetum-specific genes, were unaffected by high temperatures. These results suggest that not all tapetal genes are inhibited by increased temperatures and the tapetum itself is not degraded in such an environment. However, high temperatures may disrupt some of the tapetum functions required for pollen adhesion and germination on the stigma.
DOI:10.1016/j.cub.2013.06.060URLPMID:23910659 [本文引用: 1]
Flowering plants in the reproductive stage are particularly vulnerable to ambient temperature fluctuations [1-6]. Nevertheless, they maintain seed production under certain levels of exposure to temperature change. The mechanisms underlying this temperature tolerance are largely unknown. Using an in vitro Arabidopsis pollen tube culture, we found that a synthetic CLV3/ESR-related peptide, CLE45, prolonged pollen tube growth. A subsequent screen of Arabidopsis mutants of leucine-rich repeat receptor-like kinase genes identified two candidate receptors for CLE45 peptide, STERILITY-REGULATING KINASE MEMBER1 (SKM1) and SKM2. The double loss-of-function mutant was insensitive to CLE45 peptide in terms of pollen tube growth in vitro. The SKM1 protein actually interacted with CLE45 peptide. CLE45was preferentially expressed in the stigma in the pistil at 22 degrees C, but upon temperature shift to 30 degrees C, its expression expanded to the transmitting tract, along which pollen tubes elongated. In contrast, both SKM1 and SKM2 were expressed in pollen. Disturbance of CLE45-SKM1/SKM2 signaling transduction by either RNAi suppression of CLE45 expression or introduction of a kinase-dead version of SKM1 into skm1 plants reduced seed production at 30 degrees C, but not at 22 degrees C. Taken together with the finding that CLE45 peptide application alleviated mitochondrial decay during the in vitro pollen tube culture, these results strongly suggest that the pollen-pistil interaction via the CLE45-SKM1/SKM2 signaling pathway sustains pollen performance under higher temperatures, leading to successful seed production.
DOI:10.1080/07352689.2011.615687URL [本文引用: 1]
Ambient temperatures have increased since the beginning of the century and are predicted to continue rising under climate change. Such increases in temperature can cause heat stress: a severe threat to wheat production in many countries, particularly when it occurs during reproductive and grain-filling phases. Heat stress reduces plant photosynthetic capacity through metabolic limitations and oxidative damage to chloroplasts, with concomitant reductions in dry matter accumulation and grain yield. Genotypes expressing heat shock proteins are better able to withstand heat stress as they protect proteins from heat-induced damage. Heat tolerance can be improved by selecting and developing wheat genotypes with heat resistance. Wheat pre-breeding and breeding may be based on secondary traits like membrane stability, photosynthetic rate and grain weight under heat stress. Nonetheless, improvement in grain yield under heat stress implies selecting genotypes for grain size and rate of grain filling. Integrating physiological and biotechnological tools with conventional breeding techniques will help to develop wheat varieties with better grain yield under heat stress during reproductive and grain-filling phases. This review discusses the impact of heat stress during reproductive and grain-filling stages of wheat on grain yield and suggests strategies to improve heat stress tolerance in wheat.
DOI:10.1111/pce.12927URLPMID:28155228 [本文引用: 1]
Abstract HSP70-HSP90 organizing protein (HOP) is a well-studied family of cytosolic cochaperones. However, the possible role of HOP during the endoplasmic reticulum (ER) stress response and the identity of its interactors within the ER were not previously addressed in any eukaryote. We have demonstrated that Arabidopsis HOP3, whose function was not studied before, interacts in vivo with cytosolic HSP90 and HSP70, and, unexpectedly, with binding immunoglobulin protein (BiP), a HSP70 ER-resident protein. Although BiP lacks the domain described in other eukaryotes for HOP-HSP70 binding, it interacts with HOP3 through a non-canonical association to its nucleotide binding domain. Consistent with this interaction with BiP, HOP3 is partially localized at the ER. Moreover, HOP3 is induced both at transcript and protein levels by unfolded protein response (UPR) inducer agents by a mechanism dependent on inositol-requiring enzyme 1 (IRE1). Importantly, hop3 loss-of-function mutants show a reduction in pollen germination and a hypersensitive phenotype in the presence of ER stress inducer agents, a phenotype that is reverted by the addition of the chemical chaperone tauroursodeoxycholic acid (TUDCA). All these data demonstrate, for the first time in any eukaryote, a main role of HOP as an important regulator of the ER stress response, a process intimately linked in plants to important specific developmental programs and to environmental stress sensing and response. 2017 John Wiley & Sons Ltd.
[本文引用: 2]
DOI:10.1016/j.scienta.2006.03.007URL [本文引用: 1]
Exposure to high temperatures (heat stress) causes reduced yield in tomatoes ( Lycopersicon esculentum), mainly by affecting male gametophyte development. Two experiments were conducted where several tomato cultivars were grown under heat stress, in growth chambers (day/night temperatures of 31/25 C) or in greenhouses (day/night temperatures of 32/26 C), or under control (day/night temperatures of 28/22 C) conditions. In heat-sensitive cultivars, heat stress caused a reduction in the number of pollen grains, impaired their viability and germinability, caused reduced fruit set and markedly reduced the numbers of seeds per fruit. In the heat-tolerant cultivars, however, the number and quality of pollen grains, the number of fruits and the number of seeds per fruit were less affected by high temperatures. In all the heat-sensitive cultivars, the heat-stress conditions caused a marked reduction in starch concentration in the developing pollen grains at 3 days before anthesis, and a parallel decrease in the total soluble sugar concentration in the mature pollen, whereas in the four heat-tolerant cultivars tested, starch accumulation at 3 days before anthesis and soluble sugar concentration at anthesis were not affected by heat stress. These results indicate that the carbohydrate content of developing and mature tomato pollen grains may be an important factor in determining pollen quality, and suggest that heat-tolerant cultivars have a mechanism for maintaining the appropriate carbohydrate content under heat stress.
DOI:10.1007/s00497-016-0276-8URLPMID:27022919 [本文引用: 1]
Abstract KEY MESSAGE: Importance of the UPR for pollen. Pollen is particularly sensitive to environmental conditions that disturb protein homeostasis, such as higher temperatures. Their survival is dependent on subcellular stress response systems, one of which maintains protein homeostasis in the endoplasmic reticulum (ER). Disturbance of ER proteostasis due to stress leads to the activation of the unfolded protein response (UPR) that mitigates stress damage mainly by increasing ER-folding capacity and reducing folding demands. The UPR is controlled by ER membrane-associated transcription factors and an RNA splicing factor. They are important components of abiotic stress responses including general heat stress response and thermotolerance. In addition to responding to environmental stresses, the UPR is implicated in developmental processes required for successful male gametophyte development and fertilization. Consequently, defects in the UPR can lead to pollen abortion and male sterility. Several UPR components are involved in the elaboration of the ER network, which is required for pollen germination and polar tube growth. Transcriptome and proteome analyses have shown that components of the ER-folding machinery and the UPR are upregulated at specific stages of pollen development supporting elevated demands for secretion. Furthermore, genetic studies have revealed that knockout mutants of UPR genes are defective in producing viable or competitive pollen. In this review, we discuss recent findings regarding the importance of the UPR for both pollen development and stress response.
DOI:10.1104/pp.15.01913URLPMID:26917685 [本文引用: 2]
Male reproductive tissues are more sensitive to heat stress (HS) compared to vegetative tissues, but the basis of this phenomenon is poorly understood. Heat stress transcription factors (Hsfs) regulate the transcriptional changes required for protection from HS. In tomato (Solanum lycopersicum), HsfA2 acts as coactivator of HsfA1a and is one of the major Hsfs accumulating in response to elevated temperatures. The contribution of HsfA2 in heat stress response (HSR) and thermotolerance was investigated in different tissues of transgenic tomato plants with suppressed HsfA2 levels (A2AS). Global transcriptome analysis and immunodetection of two major Hsps in vegetative and reproductive tissues showed that HsfA2 regulates subsets of HS-induced genes in a tissue-specific manner. Accumulation of HsfA2 by a moderate HS treatment enhances the capacity of seedlings to cope with a subsequent severe HS, suggesting an important role for HsfA2 in regulating acquired thermotolerance. In pollen, HsfA2 is an important coactivator of HsfA1a during HSR. HsfA2 suppression reduces the viability and germination rate of pollen that received the stress during the stages of meiosis and microspore formation but had no effect onmore advanced stages. In general, pollenmeiocytes andmicrospores are characterized by increased susceptibility to HS due to their lower capacity to induce a strong HSR. This sensitivity is partially mitigated by the developmentally regulated expression of HsfA2 and several HS-responsive genes mediated by HsfA1a under nonstress conditions. Thereby, HsfA2 is an important factor for the priming process that sustains pollen thermotolerance during microsporogenesis.
DOI:10.1111/pce.12396URLPMID:24995670 [本文引用: 1]
Abstract Cell survival under high temperature conditions involves the activation of heat stress response (HSR), which in principle is highly conserved among different organisms, but shows remarkable complexity and unique features in plant systems. The transcriptional reprogramming at higher temperatures is controlled by the activity of the heat stress transcription factors (Hsfs). Hsfs allow the transcriptional activation of HSR genes, among which heat shock proteins (Hsps) are best characterized. Hsps belong to multigene families encoding for molecular chaperones involved in various processes including maintenance of protein homeostasis as a requisite for optimal development and survival under stress conditions. Hsfs form complex networks to activate downstream responses, but are concomitantly subjected to cell-type-dependent feedback regulation through factor-specific physical and functional interactions with chaperones belonging to Hsp90, Hsp70 and small Hsp families. There is increasing evidence that the originally assumed specialized function of Hsf/chaperone networks in the HSR turns out to be a complex central stress response system that is involved in the regulation of a broad variety of other stress responses and may also have substantial impact on various developmental processes. Understanding in detail the function of such regulatory networks is prerequisite for sustained improvement of thermotolerance in important agricultural crops.
DOI:10.1073/pnas.0608936104URL [本文引用: 2]
DOI:10.1093/jxb/erp234URLPMID:2736902 [本文引用: 3]
Abstract Above-optimal temperatures reduce yield in tomato largely because of the high heat stress (HS) sensitivity of the developing pollen grains. The high temperature response, especially at this most HS-sensitive stage of the plant, is poorly understood. To obtain an overview of molecular mechanisms underlying the HS response (HSR) of microspores, a detailed transcriptomic analysis of heat-stressed maturing tomato microspores was carried out using a combination of Affymetrix Tomato Genome Array and cDNA-amplified fragment length polymorphism (AFLP) techniques. The results were corroborated by reverse transcription-PCR (RT-PCR) and immunoblot analyses. The data obtained reveal the involvement of specific members of the small heat shock protein (HSP) gene family, HSP70 and HSP90, in addition to the HS transcription factors A2 (HSFA2) and HSFA3, as well as factors other than the classical HS-responsive genes. The results also indicate HS regulation of reactive oxygen species (ROS) scavengers, sugars, plant hormones, and regulatory genes that were previously implicated in other types of stress. The use of cDNA-AFLP enabled the detection of genes representing pollen-specific functions that are missing from the tomato Affymetrix chip, such as those involved in vesicle-mediated transport and a pollen-specific, calcium-dependent protein kinase (CDPK2). For several genes, including LeHSFA2, LeHSP17.4-CII, as well as homologues of LeHSP90 and AtVAMP725, higher basal expression levels were detected in microspores of cv. Hazera 3042 (a heat-tolerant cultivar) compared with microspores of cv. Hazera 3017 (a heat-sensitive cultivar), marking these genes as candidates for taking part in microspore thermotolerance. This work provides a comprehensive analysis of the molecular events underlying the HSR of maturing microspores of a crop plant, tomato.
DOI:10.1111/j.1365-313X.2012.04969.xURLPMID:22372427 [本文引用: 2]
Summary An increased concentration of cytosolic calcium ions (Ca 2+ ) is an early response by plant cells to heat shock. However, the molecular mechanism underlying the heat-induced initial Ca 2+ response in plants is unclear. In this study, we identified and characterized a heat-activated Ca 2+ -permeable channel in the plasma membrane of Arabidopsis thaliana root protoplasts using reverse genetic analysis and the whole-cell patch-clamp technique. The results indicated that A.thaliana cyclic nucleotide-gated ion channel6 (CNGC6) mediates heat-induced Ca 2+ influx and facilitates expression of heat shock protein (HSP) genes and the acquisition of thermotolerance. GUS and GFP reporter assays showed that CNGC6 expression is ubiquitous in A.thaliana , and the protein is localized to the plasma membrane of cells. Furthermore, it was found that the level of cytosolic cAMP was increased by a mild heat shock, that CNGC6 was activated by cytosolic cAMP, and that exogenous cAMP promoted the expression of HSP genes. The results reveal the role of cAMP in transduction of heat shock signals in plants. The correlation of an increased level of cytosolic cAMP in a heat-shocked plant with activation of the Ca 2+ channels and downstream expression of HSP genes sheds some light on how plants transduce a heat stimulus into a signal cascade that leads to a heat shock response.
DOI:10.1093/jxb/erp316URLPMID:2803211 [本文引用: 1]
The high sensitivity of male reproductive cells to high temperatures may be due to an inadequate heat stress response. The results of a comprehensive expression analysis of HsfA2 and Hsp17-CII, two important members of the heat stress system, in the developing anthers of a heat-tolerant tomato genotype are reported here. A transcriptional analysis at different developmental anther/pollen stages was performed using semi-quantitative and real-time PCR. The messengers were localized usingin situRNA hybridization, and protein accumulation was monitored using immunoblot analysis. Based on the analysis of the gene and protein expression profiles, HsfA2 and Hsp17-CII are finely regulated during anther development and are further induced under both short and prolonged heat stress conditions. These data suggest that HsfA2 may be directly involved in the activation of protection mechanisms in the tomato anther during heat stress and, thereby, may contribute to tomato fruit set under adverse temperatures.
DOI:10.1093/pcp/pcv174URLPMID:26561535 [本文引用: 1]
Abstract Rice is one of the main food crops in the world. In the near future, yield is expected to be under pressure due to unfavorable climatic conditions, such as increasing temperatures. Therefore, improving rice germplasm in order to guarantee rice production under harsh environmental conditions is of top priority. Although many physiological studies have contributed to understanding heat responses during anthesis, the most heat-sensitive stage, molecular data are still largely lacking. In this study, an RNA-sequencing approach of heat- and control-treated reproductive tissues during anthesis was carried out using N22, one of the most heat-tolerant rice cultivars known to date. This analysis revealed that expression of genes encoding a number of transcription factor families, together with signal transduction and metabolic pathway genes, is repressed. On the other hand, expression of genes encoding heat shock factors and heat shock proteins was highly activated. Many of these genes are predominantly expressed at late stages of anther development. Further physiological experiments using heat-tolerant N22 and two sensitive cultivars suggest that reduced yield in heat-sensitive plants may be associated with poor pollen development or production in anthers prior to anthesis. In parallel, induction levels of a set of heat-responsive genes in these tissues correlated well with heat tolerance. Altogether, these findings suggest that proper expression of protective chaperones in anthers is needed before anthesis to overcome stress damage and to ensure fertilization. Genes putatively controlling this process were identified and are valuable candidates to consider for molecular breeding of highly productive heat-tolerant cultivars. The Author 2015. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists. All rights reserved. For permissions, please email: journals.permissions@oup.com.
DOI:10.1016/j.cosust.2015.05.005URL [本文引用: 1]
Natural and human systems are increasingly affected by climate change. A synopsis of the documentation of scientific evidence for the observed effects of climate change in the third, fourth and fifth assessment reports of the Intergovernmental Panel on Climate Change shows that the amount of evidence available, the range of impacts observed and their geographical scope has expanded rapidly. Fifteen years ago, robust evidence for observed climate change impacts was almost exclusively available for the cryosphere and terrestrial ecosystems in mid to high northern latitudes or mountain regions. By contrast, the effects of climate change are now documented for all land areas and oceans, for both natural and human systems. Over the last decade, evidence has increased especially for impacts on marine ecosystems, food production and wildfire regimes. No recent progress has been found in the documentation of impacts of climate change related sea level rise. Though the evidence base has improved substantially for regions in the Southern hemisphere and developing countries, the global distribution of observed impacts remains uneven.
DOI:10.3389/fpls.2013.00047URLPMID:3593198 [本文引用: 1]
Plant reproductive development is more sensitive than vegetative growth to many environmental stresses. High temperature (HT) injury is becoming an increasingly serious problem due to recent global warming. In wheat, barley, and other crops, the early phase of anther development is most susceptible to HT. I and my colleagues recently demonstrated that HT causes cell proliferation arrest and represses auxin signaling in a tissue-specific manner in the anther cells of barley andArabidopsis. HT also caused comprehensive alterations in transcription. The application of auxin at the same time blocked the transcriptional alterations, led to the production of normal pollen grains, and restored the normal seed setting rate under increasing temperatures. Although synthetic auxins have been used widely as potent and selective herbicides, these recent results indicate that auxin is useful for the promotion of fertility and maintenance of crop yields under the threat of global warming.
DOI:10.1105/tpc.110.074369URLPMID:21297036 [本文引用: 1]
The rice (Oryza sativa) floral homeotic C-class gene, MADS3, was previously shown to be required for stamen identity determination during early flower development. Here, we describe a role for MADS3 in regulating late anther development and pollen formation. Consistent with this role, MADS3 is highly expressed in the tapetum and microspores during late anther development, and a newly identified MADS3 mutant al ele, mads3-4, displays defective anther walls, aborted microspores, and complete male sterility. During late anther development, mads3-4 exhibits oxidative stress-related phenotypes. Microarray analysis revealed expression level changes in many genes in mads3-4 anthers. Some of these genes encode proteins involved in reactive oxygen species (ROS) homeostasis; among them is MT-1-4b, which encodes a type 1 small Cys-rich and metal binding protein. In vivo and in vitro assays showed that MADS3 is associated with the promoter of MT-1-4b, and recombinant MT-1-4b has Superoxide ani n and hydroxyl radical scavenging activity. Reducing the expression of MT-1-4b causes decreased pollen fertility and an increased level of superoxide anion in transgenic plants. Our findings suggest that MADS3 is a key transcriptional regulator that functions in rice male reproductive development, at least in part, by modulating ROS levels through MT-1-4b.
DOI:10.1093/jxb/erp289URLPMID:19858118 [本文引用: 1]
Episodes of high temperature at anthesis, which in rice is the most sensitive stage to temperature, are expected to occur more frequently in future climates. The morphology of the reproductive organs and pollen number, and changes in anther protein expression, were studied in response to high temperature at anthesis in three rice (Oryza sativa L.) genotypes. Plants were exposed to 6 h of high (38 degrees C) and control (29 degrees C) temperature at anthesis and spikelets collected for morphological and proteomic analysis. Moroberekan was the most heat-sensitive genotype (18% spikelet fertility at 38 degrees C), while IR64 (48%) and N22 (71%) were moderately and highly heat tolerant, respectively. There were significant differences among the genotypes in anther length and width, apical and basal pore lengths, apical pore area, and stigma and pistil length. Temperature also affected some of these traits, increasing anther pore size and reducing stigma length. Nonetheless, variation in the number of pollen on the stigma could not be related to measured morphological traits. Variation in spikelet fertility was highly correlated (r=0.97, n=6) with the proportion of spikelets with >= 20 germinated pollen grains on the stigma. A 2D-gel electrophoresis showed 46 protein spots changing in abundance, of which 13 differentially expressed protein spots were analysed by MS/MALDI-TOF. A cold and a heat shock protein were found significantly up-regulated in N22, and this may have contributed to the greater heat tolerance of N22. The role of differentially expressed proteins and morphology during anther dehiscence and pollination in shaping heat tolerance and susceptibility is discussed.
DOI:10.1016/j.jplph.2009.11.007URLPMID:20044168 [本文引用: 1]
Grain sorghum ( Sorghum bicolor) crop yield is significantly compromised by high temperature stress-induced male sterility, and is attributed to reduced cell wall invertase (CWI)-mediated sucrose hydrolysis in microspores and anthers leading to altered carbohydrate metabolism and starch deficiency in pollen ( Jain et al., 2007). Sorghum plants were grown under season-long ambient (30/20 °C day-time maximum/night-time minimum) or high temperature stress (HS, 36/26 °C) environments, or reciprocally transferred for 5–10 days between either temperature regimens through panicle and microspore developmental stages. Quantitative RT-PCR analyses for CWI gene SbIncw1, plasma membrane H +-ATPase ( Mha1) and sugar transporter proteins ( OsSUT3 and OsMST7 homologs in sorghum), starch deficiency and pollen sterility data are presented to confirm HS-sensitivity of pre- and post-meiotic stages of sorghum microsporogenesis. Heat stress-induced reduction in Incw transcriptional activity during microspore meiosis was irreversible despite return of optimal growth temperature conditions through further reproductive development.
DOI:10.1007/s00497-018-0339-0URL [本文引用: 1]
Key message Tomato pollen grains have the capacity for ethylene production, possessing specific components of the ethylene-biosynthesis and -signaling pathways, being affected/responsive to...
DOI:10.1093/aob/mci149URLPMID:15851397 [本文引用: 1]
61 Background and Aims High-temperature environments with >30 °C during flowering reduce boll retention and yield in cotton. Therefore, identification of cotton cultivars with high-temperature tolerance would be beneficial in both current and future climates. 61 Methods Response to temperature (10-45 °C at 5 °C intervals) of pollen germination and pollen tube growth was quantified, and their relationship to cell membrane thermostability was studied in 12 cultivars. A principal component analysis was carried out to classify the genotypes for temperature tolerance. 61 Key Results Pollen germination and pollen tube length of the cultivars ranged from 20 to 60 % and 411 to 903 08m, respectively. A modified bilinear model best described the response to temperature of pollen germination and pollen tube length. Cultivar variation existed for cardinal temperatures (Tmin, Topt and Tmax) of pollen germination percentage and pollen tube growth. Mean cardinal temperatures calculated from the bilinear model for the 12 cultivars were 15·0, 31·8 and 43·3 °C for pollen germination and 11·9, 28·6 and 42·9 °C for pollen tube length. No significant correlations were found between pollen parameters and leaf membrane thermostability. Cultivars were classified into four groups based on principal component analysis. 61 Conclusions Based on principal component analysis, it is concluded that higher pollen germination percentages and longer pollen tubes under optimum conditions and with optimum temperatures above 32 °C for pollen germination would indicate tolerance to high temperature.
DOI:10.1093/dnares/dsw051URLPMID:28025318 [本文引用: 2]
Abstract Alternative splicing (AS) is a key control mechanism influencing signal response cascades in different developmental stages and under stress conditions. In this study, we examined heat stress (HS)-induced AS in the heat sensitive pollen tissue of two tomato cultivars. To obtain the entire spectrum of HS-related AS, samples taken directly after HS and after recovery were combined and analysed by RNA-seq. For nearly 9,200 genes per cultivar, we observed at least one AS event under HS. In comparison to control, for one cultivar we observed 76% more genes with intron retention (IR) or exon skipping (ES) under HS. Furthermore, 2,343 genes had at least one transcript with IR or ES accumulated under HS in both cultivars. These genes are involved in biological processes like protein folding, gene expression and heat response. Transcriptome assembly of these genes revealed that most of the alternative spliced transcripts possess truncated coding sequences resulting in partial or total loss of functional domains. Moreover, 141 HS specific and 22 HS repressed transcripts were identified. Further on, we propose AS as layer of stress response regulating constitutively expressed genes under HS by isoform abundance. The Author 2016. Published by Oxford University Press on behalf of Kazusa DNA Research Institute.
DOI:10.1186/s12864-018-4824-5URLPMID:29884134 [本文引用: 1]
Background Pollen development is central for plant reproduction and is assisted by changes of the transcriptome and proteome. At the same time, pollen development and viability is largely sensitive to...
DOI:10.1155/2015/238704URLPMID:26618163 [本文引用: 1]
In rice, the stage of development most sensitive to high temperature stress is flowering, and exposure at this stage can result in spikelet sterility, thereby leading to significant yield losses. In this study, protein expression patterns of rice anthers from Dianxi4, a high temperature tolerant Japonica rice variety, were compared between samples exposed to high temperature and those grown in natural field conditions in Korea. Shotgun proteomics analysis of three replicate control and high-temperature-treated samples identified 3,266 nonredundant rice anther proteins (false discovery rate < 0.01). We found that high levels of ATP synthase, cupin domain-containing proteins, and pollen allergen proteins were present in rice anthers. Comparative analyses of 1,944 reproducibly expressed proteins identified 139 differentially expressed proteins, with 95 increased and 44 decreased in response to high temperature conditions. Heat shock, DnaK family, and chaperone proteins showed highly increased expression, suggesting that the high temperature tolerance of Dianxi4 is achieved by stabilization of proteins in pollen cells. Trehalose synthase was also highly increased after heat treatment, suggesting a possible role for trehalose in preventing protein denaturation through desiccation.
DOI:10.1016/j.pbi.2007.04.011URLPMID:17482504 [本文引用: 2]
Plants have evolved a variety of responses to elevated temperatures that minimize damage and ensure protection of cellular homeostasis. New information about the structure and function of heat stress proteins and molecular chaperones has become available. At the same time, transcriptome analysis of Arabidopsis has revealed the involvement of factors other than classical heat stress responsive genes in thermotolerance. Recent reports suggest that both plant hormones and reactive oxygen species also contribute to heat stress signaling. Additionally, an increasing number of mutants that have altered thermotolerance have extended our understanding of the complexity of the heat stress response in plants.
DOI:10.1016/j.sjbs.2015.03.006URLPMID:4778538 [本文引用: 1]
Heat stress adversely affects the growth and yield of faba bean crop. Accumulation of ClpB/Hsp100 class of proteins is a critical parameter in induction of acquired heat stress tolerance in plants. Heat-induced expression of ClpB/Hsp100 genes has been noted in diverse plant species. Using primers complementary to soybean ClpB/Hsp100 gene, we analyzed the transcript expression profile of faba bean ClpB/Hsp100 gene in leaves of seedlings and flowering plants and in pollen grains. ClpB/Hsp100 protein accumulation profile was analyzed in leaves of faba bean seedlings using Arabidopsis thaliana cytoplasmic Hsp101 antibodies. The transcript and protein levels of faba bean ClpB/Hsp100 were significantly induced in response to heat stress.
DOI:10.1007/s13562-013-0214-xURL [本文引用: 1]
The reproductive stage of wheat is highly sensitive to the heat stress, especially pollen viability. Here, we report the alterations in the expression of heat shock proteins and antioxidant enzymes in wheat pollen of thermotolerant (C-306) and susceptible (HD2329) cultivars under the heat shock (4202°C, 202h) with or without ascorbic acid (40002mM) treatment. A significant fold change in expression of catalase ( CAT ) and ascorbate peroxidase ( APX ) was observed in the pollens of HD2329 in response to 40002mM ascorbic acid (AsA) before heat stress (T 1 ). The SOD transcript was observed significantly high in C-306 pollens compared to HD2329 in response to T 1 . The transcripts of high and low molecular weight HSPs ( HSP70 , HSP90 , HSP17 and HSP26 ) were very high in pollens of C-306 as compared to HD2329. Semi-quantitative RT-PCR showed marked variations in the transcript of HSPs and antioxidant enzymes in pollens of C-306 and HD2329 under T 0 and T 1 treatments. 1D and 2D polyacrylamide gel electrophoresis of pollens showed more differentially expressed proteins in C-306 than in HD2329 in response to T 1 . A significant increase in the accumulation of osmolyte (proline), H 2 O 2 and endogenous AsA levels were observed in the pollens of C-306 than in HD2329, in response to T 0 and T 1 treatments. Immunoblot analysis using monoclonal antibodies of HSP70 and HSP90 showed abundance of HSP70 proteins in the pollens of C-306 and HSP90 proteins in pollens of HD2329. Scanning electron microscopy of pollen under the heat stress showed disintegrated and dehydrated exine layer and alteration in pollen structure from spheroid to ovoid in both the cultivars. The pollen viability was found to be less in HD2329 than in C-306 in response to T 1 treatment. A significant increase in the total antioxidant capacity was observed in pollens of C-306 as compared to HD2329 in response to the treatments. Findings indicate that pre-anthesis treatment of 40002mM AsA before HS (4202°C, 202h) enhances the thermotolerance capacity of wheat pollens, as assessed through biochemical markers; further investigations on similar elicitors may yield useful information on mitigating the effect of the terminal heat on reproductive system of wheat.
DOI:10.1016/j.cell.2009.11.006URLPMID:20079334 [本文引用: 2]
Abstract Plants are highly sensitive to temperature and can perceive a difference of as little as 1 degrees C. How temperature is sensed and integrated in development is unknown. In a forward genetic screen in Arabidopsis, we have found that nucleosomes containing the alternative histone H2A.Z are essential to perceiving ambient temperature correctly. Genotypes deficient in incorporating H2A.Z into nucleosomes phenocopy warm grown plants, and show a striking constitutive warm temperature transcriptome. We show that nucleosomes containing H2A.Z display distinct responses to temperature in vivo, independently of transcription. Using purified nucleosomes, we are able to show that H2A.Z confers distinct DNA-unwrapping properties on nucleosomes, indicating a direct mechanism for the perception of temperature through DNA-nucleosome fluctuations. Our results show that H2A.Z-containing nucleosomes provide thermosensory information that is used to coordinate the ambient temperature transcriptome. We observe the same effect in budding yeast, indicating that this is an evolutionarily conserved mechanism.
DOI:10.1093/mp/ssq014URLPMID:20410255 [本文引用: 1]
Plants, as sessile organisms, need to sense and adapt to heterogeneous environments and have developed sophisticated responses by changing their cellular physiology, gene regulation, and genome stability. Recent work demonstrated heritable stress effects on the control of genome stability in plants phenomenon that was suggested to be of epigenetic nature. Here, we show that temperature and UV-B stress cause immediate and heritable changes in the epigenetic control of a silent reporter gene in Arabidopsis. This stress-mediated release of gene silencing correlated with pronounced alterations in histone occupancy and in histone H3 acetylation but did not involve adjustments in DNA methylation. We observed transmission of stress effects on reporter gene silencing to non-stressed progeny, but this effect was restricted to areas consisting of a small number of cells and limited to a few non-stressed progeny generations. Furthermore, stress-induced release of gene silencing was antagonized and reset during seed aging. The transient nature of this phenomenon highlights the ability of plants to restrict stress-induced relaxation of epigenetic control mechanisms, which likely contributes to safeguarding genome integrity.
DOI:10.1104/pp.107.112060URL [本文引用: 1]
Plants can acclimate rapidly to environmental conditions, including high temperatures. To identify molecular events important for acquired thermotolerance, we compared viability and transcript profiles of Arabidopsis thaliana treated to severe heat stress (45°C) without acclimation or following two different acclimation treatments. Notably, a gradual increase to 45°C (22°C to 45°C over 6 h) led to higher survival and to more and higher-fold transcript changes than a step-wise acclimation (90 min at 38°C plus 120 min at 22°C before 45°C). There were significant differences in the total spectrum of transcript changes in the two treatments, but core components of heat acclimation were apparent in the overlap between treatments, emphasizing the importance of performing transcriptome analysis in the context of physiological response. In addition to documenting increases in transcripts of specific genes involved in processes predicted to be required for thermotolerance (i.e. protection of proteins and of translation, limiting oxidative stress), we also found decreases in transcripts (i.e. for programmed cell death, basic metabolism, and biotic stress responses), which are likely equally important for acclimation. Similar protective effects may also be achieved differently, such as prevention of proline accumulation, which is toxic at elevated temperatures and which was reduced by both acclimation treatments but was associated with transcript changes predicted to either reduce proline synthesis or increase degradation in the two acclimation treatments. Finally, phenotypic analysis of T-DNA insertion mutants of genes identified in this analysis defined eight new genes involved in heat acclimation, including cytosolic ascorbate peroxidase and the transcription factors HsfA7a (heat shock transcription factor A7a) and NF-X1.
[本文引用: 1]
[本文引用: 1]
DOI:10.4161/psb.6.5.14979URLPMID:20709814 [本文引用: 1]
The final shape of shoot lateral organs, namely, leaves and flowers, is determined by coordinated growth after the initiation of primordia from shoot meristems in seed plants. This coordination is achieved by the complex action of many transcription factors, which include the TEOSINTE BRANCHED1, CYCLOIDEA, and PCF (TCP) family. We have recently reported that CINCINNATA-like (CIN-like) TCP genes act dose-dependently to regulate the flat and smooth morphology of leaves in Arabidopsis thaliana. In contrast, the roles of CIN-like TCP genes in flower development are poorly understood. In this report, using multiple tcp mutants and transgenic plants in which the activity of CIN-like TCP transcription factors is dominantly inhibited, we found that these TCPs regulate the smooth and flat morphology of petals. Based on these findings, we discuss a possible strategy to generate a fringed morphology in floricultural plants.
DOI:10.3389/fpls.2015.00267URLPMID:4408840 [本文引用: 1]
Plants have evolved sophisticated genetic and epigenetic regulatory systems to respond quickly to unfavorable environmental conditions such as heat, cold, drought, and pathogen infections. In particular, heat greatly affects plant growth and development, immunity and circadian rhythm, and poses a serious threat to the global food supply. According to temperatures exposing, heat can be usually classified as warm ambient temperature (about 22–27°C), high temperature (27–30°C) and extremely high temperature (37–42°C, also known as heat stress) for the model plant Arabidopsis thaliana. The genetic mechanisms of plant responses to heat have been well studied, mainly focusing on elevated ambient temperature-mediated morphological acclimation and acceleration of flowering, modulation of circadian clock and plant immunity by high temperatures, and thermotolerance to heat stress. Recently, great progress has been achieved on epigenetic regulation of heat responses, including DNA methylation, histone modifications, histone variants, ATP-dependent chromatin remodeling, histone chaperones, small RNAs, long non-coding RNAs and other undefined epigenetic mechanisms. These epigenetic modifications regulate the expression of heat-responsive genes and function to prevent heat-related damages. This review focuses on recent progresses regarding the genetic and epigenetic control of heat responses in plants, and pays more attention to the role of the major epigenetic mechanisms in plant heat responses. Further research perspectives are also discussed.
DOI:10.1126/science.1204531URLPMID:21551030 [本文引用: 1]
Abstract Efforts to anticipate how climate change will affect future food availability can benefit from understanding the impacts of changes to date. We found that in the cropping regions and growing seasons of most countries, with the important exception of the United States, temperature trends from 1980 to 2008 exceeded one standard deviation of historic year-to-year variability. Models that link yields of the four largest commodity crops to weather indicate that global maize and wheat production declined by 3.8 and 5.5%, respectively, relative to a counterfactual without climate trends. For soybeans and rice, winners and losers largely balanced out. Climate trends were large enough in some countries to offset a significant portion of the increases in average yields that arose from technology, carbon dioxide fertilization, and other factors.
DOI:10.1007/BF00021240URL [本文引用: 1]
The effects of high temperature on mature pollen of various maize lines were investigated. Genotypic differences in pollen reaction to high temperature were revealed. Pollen grains resistant to high temperature (35°C, 26°C) were characterized by higher germination capacity and better ability to develop normal pollen tubes. The studies are of interest to evaluate reproductive system tolerance and conduct gamete selection at the mature pollen grain stage in maize.
DOI:10.1371/journal.pone.0132500URLPMID:4505944 [本文引用: 1]
The Arabidopsis TMS1 encodes a heat shock protein identical to the Hsp40 protein AtERdj3A and plays important roles in the thermotolerance of pollen tubes and other plant tissues. Despite its importance to plant growth and reproduction, little has been known about its mechanisms underlying thermotolerance of plants. In this study, the relationship between TMS1 and the Hsp70 proteins, Binding Immunoglobulin Proteins (BiPs) was explored to understand the molecular mechanisms of TMS1 in thermotolerance of plants. The expression of TMS1 was induced not only by heat shock, but also by dithiothreitol (DTT) and L-azetidine-2-carboxylic acid (AZC), similarly to the three BiP genes, indicating that TMS1 may be involved in unfolded protein response (UPR). The firefly luciferase complementary imaging (LCI), GST pull-down and ATPase enzyme activity assays demonstrated that the DnaJ domain of TMS1 could interact with BiP1 and BiP3, and could stimulate their ATPase enzyme activities. In addition, the expression level of TMS1 was reduced in the bzip28 bzip60 double mutant. These results suggest that TMS1 may function at the downstream of bZIP28 and bZIP60 and be involved in termotolerance of plants, possibly by participating in refolding or degradation of unfolded and misfolded proteins through interaction with the BiPs.
DOI:10.1007/s00497-016-0281-yURLPMID:27016360 [本文引用: 3]
Key message Pollen thermotolerance.
DOI:10.4161/psb.27971URLPMID:4091214 [本文引用: 1]
Exposure to heat stress causes physiological and epigenetic changes in plants, which may also be altered in the progeny. We compared the progeny of stressed and control Arabidopsis thaliana wild type and Dicer-like mutant dcl2, dcl3, and dcl4 plants for variations in physiology and molecular profile, including global genome methylation, mRNA levels, and histone modifications in the subset of differentially expressed genes at normal conditions and in response to heat stress. We found that the immediate progeny of heat-stressed plants had fewer, but larger leaves, and tended to bolt earlier. Transposon expression was elevated in the progeny of heat-stressed plants, and heat stress in the same generation tended to decrease global genome methylation. Progeny of stressed plants had increased expression of HSFA2, and reduction in MSH2, ROS1, and several SUVH genes. Gene expression positively correlated with permissive histone marks and negatively correlated with repressive marks. Overall, the progeny of heat stressed plants varied in both their physiology and epigenome and dcl2 and dcl3 mutants were partially deficient for these changes.
DOI:10.1104/pp.113.232314URLPMID:24481135 [本文引用: 1]
Male reproduction in flowering plants is highly sensitive to high temperature (HT). To investigate molecular mechanisms of the response of cotton (Gossypium hirsutum) anthers to HT, a relatively complete comparative transcriptome analysis was performed during anther development of cotton lines 84021 and H05 under normal temperature and HT conditions. In total, 4,599 differentially expressed genes were screened; the differentially expressed genes were mainly related to epigenetic modifications, carbohydrate metabolism, and plant hormone signaling. Detailed studies showed that the deficiency in S-adenosyl-L-homocysteine hydrolase1 and the inhibition of methyltransferases contributed to genome-wide hypomethylation in H05, and the increased expression of histone constitution genes contributed to DNA stability in 84021. Furthermore, HT induced the expression of casein kinasei (GhCKI) in H05, coupled with the suppression of starch synthase activity, decreases in glucose level during anther development, and increases in indole-3-acetic acid (IAA) level in late-stage anthers. The same changes also were observed in Arabidopsis (Arabidopsis thaliana) GhCKI overexpression lines. These results suggest that GhCKI, sugar, and auxin may be key regulators of the anther response to HT stress. Moreover, phytochrome-interacting factor genes (PIFs), which are involved in linking sugar and auxin and are regulated by sugar, might positively regulate IAA biosynthesis in the cotton anther response to HT. Additionally, exogenous IAA application revealed that high background IAA may be a disadvantage for late-stage cotton anthers during HT stress. Overall, the linking of HT, sugar, PIFs, and IAA, together with our previously reported data on GhCKI, may provide dynamic coordination of plant anther responses to HT stress.
DOI:10.1016/j.tibs.2011.11.007URLPMID:22236506 [本文引用: 1]
In plants, the heat stress response (HSR) is highly conserved and involves multiple pathways, regulatory networks and cellular compartments. At least four putative sensors have recently been proposed to trigger the HSR. They include a plasma membrane channel that initiates an inward calcium flux, a histone sensor in the nucleus, and two unfolded protein sensors in the endoplasmic reticulum and the cytosol. Each of these putative sensors is thought to activate a similar set of HSR genes leading to enhanced thermotolerance, but the relationship between the different pathways and their hierarchical order is unclear. In this review, we explore the possible involvement of different thermosensors in the plant response to warming and heat stress.
DOI:10.1007/s00497-016-0282-xURLPMID:4909792 [本文引用: 2]
Pollen heat acclimation. As a consequence of global warming, plants have to face more severe and more frequently occurring periods of high temperature stress. While this affects the whole plant, development of the male gametophyte, the pollen, seems to be the most sensitive process. Given the great importance of functioning pollen for the plant life cycle and for agricultural production, it is necessary to understand this sensitivity. While changes in temperature affect different components of all cells and require a cellular response and acclimation, high temperature effects and responses in developing pollen are distinct from vegetative tissues at several points. This could be related to specific physiological characteristics of developing pollen and supporting tissues which make them vulnerable to high temperature, or its derived effects such as ROS accumulation and carbohydrate starvation. But also expression of heat stress-responsive genes shows unique patterns in developing pollen when compared to vegetative tissues that might explain the failure to withstand high temperatures. As an alternative to viewing pollen failure under high temperature as a result of inherent sensitivity of a specific developmental process, we end by discussing whether it might actually be an adaptation.
DOI:10.1016/j.jplph.2009.01.011URLPMID:19324457 [本文引用: 1]
To induce androgenesis in field pea, grass pea and the model legume species Medicago truncatula, isolated microspores of various genotypes of these three species were submitted to a range of abiotic stresses prior to and during their initial culture, in order to stimulate them to divide and form embryos. Some stress agents had a positive effect on androgenesis from the treated microspores. Submission of flower buds to a cold period prior to anther excision or microspore isolation, modifying the osmotic pressure of the medium during initial culture and electroporation of isolated microspores were the three major individual stress agents to have an impact on the efficiency of androgenetic proliferation and subsequent differentiation from the microspores of pea, grass pea and M. truncatula genotypes. A combination of osmotic and electric shocks significantly improved responses from isolated microspores and yielded microcalluses and then calluses, but only few underwent morphogenesis. Further work is under way to improve responses and extend them to other genotypes. The results reported here are, to the best of our knowledge, the first successful results from isolated microspores of these species.
DOI:10.1093/pcp/pcm100URLPMID:17693452 [本文引用: 1]
Abstract Cold temperatures cause pollen sterility and large reductions in grain yield in temperate rice growing regions of the world. Induction of pollen sterility by cold involves a disruption of sugar transport in anthers, caused by the cold-induced repression of the apoplastic sugar transport pathway in the tapetum. Here we demonstrate that the phytohormone ABA is a potential signal for cold-induced pollen sterility (CIPS). Cold treatment of the cold-sensitive cultivar Doongara resulted in increased anther ABA levels. Exogenous ABA treatment at the young microspore stage induced pollen sterility and affected cell wall invertase and monosaccharide transporter gene expression in a way similar to cold treatment. In the cold-tolerant cultivar R31, ABA levels were significantly lower under normal circumstances and remained low after cold treatment. The differences in endogenous ABA levels in Doongara and R31 correlated with differences in expression of the ABA biosynthetic genes encoding zeaxanthin epoxidase (OSZEP1) and 9-cis-epoxycarotenoid dioxygenase (OSNCED2, OSNCED3) in anthers. The expression of three ABA-8-hydroxylase genes (ABA8OX1, 2 and 3) in R31 anthers was higher under control conditions and was regulated differently by cold compared with Doongara. Our results indicate that the cold tolerance phenotype of R31 is correlated with lower endogenous ABA levels and a different regulation of ABA metabolism.
DOI:10.1508/cytologia.79.49URL [本文引用: 1]
DOI:10.1007/s00438-007-0229-xURLPMID:17429694 [本文引用: 1]
High-temperature stress causes abortive male reproductive development in many plant species. Here, we report a putative mechanism of high-temperature injury during anther early development in barley plants ( Hordeum vulgare L). Under high-temperature conditions (30 C day/25 C night), cell-proliferation arrest, increased vacuolization, over-development of chloroplasts, and certain abnormalities of the mitochondria, nuclear membrane, and rough endoplasmic reticulum (RER) were observed in developing anther cells, but not in developing ovule cells. Moreover, premature degradation of tapetum cells and premature progression to meiotic prophase in pollen mother cells (PMCs) were also observed. To monitor transcriptional alterations during high-temperature injury, we performed DNA microarray analysis using the 22K Barley1 GeneChip. Expression profiles were captured at four time points during the early development of panicles, and during vegetative growth of seedlings as a control, with or without high-temperature treatment. Abiotic or biotic stress related genes were equally or more dominantly up-regulated in the seedlings exposed to high temperatures compared with the panicles. In contrast, certain genes associated with histones, DNA replication initiation, mitochondria, and ribosomes were specifically repressed in the exposed panicles. In situ hybridization studies indicated that repression locally occurred on the developing anther cells exposed to high temperatures. Microarray analysis also indicated that a series of genes, including a meiosis-specific gene Asy1 and anther-specific lipid transfer protein genes, was prematurely up-regulated at an earlier stage under high-temperature conditions. Real-time quantitative RT-PCR analyses well confirmed the expression differences of certain key genes predicted by the DNA microarrays. These results suggest that high-temperature causes premature progression of anther early development program and fate, such as progression to meiosis of PMCs, cell-proliferation arrest and degradation in anther wall cells, accompanied by comprehensive alterations in transcription.
DOI:10.1071/FP12090URL [本文引用: 1]
Many self-fertilising crops are particularly sensitive to abiotic stress at the reproductive stage. In rice (Oryza sativa L.) and wheat (Triticum aestivum L.), for example, abiotic stress during meiosis and the young microspore stage indicates the tapetum is highly vulnerable and that the developmental program appears to be compromised. Tapetal hypertrophy can occur as a consequence of cold and drought stress, and programmed cell death (PCD) is delayed or inhibited. Since the correct timing of tapetal PCD is essential for pollen reproduction, substantial losses in grain yield occur. In wheat and rice, a decrease in tapetal cell wall invertase levels is correlated with pollen abortion and results in the amount of hexose sugars reaching the tapetum, and subsequently the developing microspores, being severely reduced ( tarvation hypothesis ). ABA and gibberellin levels may be modified by cold and drought, influencing levels of cell wall invertase(s) and the tapetal developmental program, respectively. Many genes regulating tapetal and microspore development have been identified in Arabidopsis thaliana (L.) Heynh. and rice and the specific effects of abiotic stresses on the program and pathways can now begin to be assessed.
DOI:10.1007/s00425-015-2394-1URLPMID:26335855 [本文引用: 1]
http://link.springer.com/article/10.1007/s00425-015-2394-1
DOI:10.3390/metabo4040889URLPMID:4279151 [本文引用: 1]
Crop production is highly sensitive to elevated temperatures. A rise of a few degrees above the optimum growing temperature can lead to a dramatic yield loss. A predicted increase of 1 3 degrees in the twenty first century urges breeders to develop thermo-tolerant crops which are tolerant to high temperatures. Breeding for thermo-tolerance is a challenge due to the low heritability of this trait. A better understanding of heat stress tolerance and the development of reliable methods to phenotype thermo-tolerance are key factors for a successful breeding approach. Plant reproduction is the most temperature-sensitive process in the plant life cycle. More precisely, pollen quality is strongly affected by heat stress conditions. High temperature leads to a decrease of pollen viability which is directly correlated with a loss of fruit production. The reduction in pollen viability is associated with changes in the level and composition of several (groups of) metabolites, which play an important role in pollen development, for example by contributing to pollen nutrition or by providing protection to environmental stresses. This review aims to underline the importance of maintaining metabolite homeostasis during pollen development, in order to produce mature and fertile pollen under high temperature. The review will give an overview of the current state of the art on the role of various pollen metabolites in pollen homeostasis and thermo-tolerance. Their possible use as metabolic markers to assist breeding programs for plant thermo-tolerance will be discussed.
DOI:10.1093/pcp/pcs044URL [本文引用: 1]
DOI:10.1093/jxb/err052URLPMID:21398431 [本文引用: 3]
Polyploidy is an important evolutionary phenomenon but the mechanisms by which polyploidy arises still remain underexplored. There may be an environmental component to polyploidization. This study aimed to clarify how temperature may promote diploid gamete formation considered an essential element for sexual polyploidization. First of all, a detailed cytological analysis of microsporogenesis and microgametogenesis was performed to target precisely the key developmental stages which are the most sensitive to temperature. Then, heat-induced modifications in sporad and pollen characteristics were analysed through an exposition of high temperature gradient. Rosa plants are sensitive to high temperatures with a developmental sensitivity window limited to meiosis. Moreover, the range of efficient temperatures is actually narrow. 36 C at early meiosis led to a decrease in pollen viability, pollen ectexine defects but especially the appearance of numerous diploid pollen grains. They resulted from dyads or triads mainly formed following heat-induced spindle misorientations in telophase II. A high temperature environment has the potential to increase gamete ploidy level. The high frequencies of diplogametes obtained at some extreme temperatures support the hypothesis that polyploidization events could have occurred in adverse conditions and suggest polyploidization facilitating in a global change context.
[本文引用: 1]
DOI:10.1016/j.fcr.2005.04.008URL [本文引用: 1]
Spikelet fertility (seed-set) is an important component of yield that is sensitive to high temperature. The objectives of this research were (a) to quantify the effects of high temperature on spikelet fertility and harvest index of rice; (b) to determine if there were species, ecotype, and/or cultivar differences in response to high temperature; and (c) to understand the reasons for lower and/or differential spikelet fertility and harvest index of rice cultivars at high temperatures. Fourteen rice cultivars of different species ( Oryza sativa and Oryza glaberrima), ecotypes ( indica and japonica) and origin (temperate and tropical) were exposed to ambient and high temperature (ambient + 5 C) at Gainesville, Florida. High temperature significantly decreased spikelet fertility across all cultivars, but effects varied among cultivars. Based on decreases in spikelet fertility at high temperature, cultivar N-22 was most tolerant, while cultivars L-204, M-202, Labelle, Italica Livorna, WAB-12, CG-14 and CG-17 were highly susceptible and cultivars M-103, S-102, Koshihikari, IR-8 and IR-72 were moderately susceptible to high temperature. There were no clear species or ecotype differences, as some cultivars in each species or within ecotypes of tropical and temperature origin were equally susceptible to high temperature (for example M-202 temperate japonica, Labelle tropical japonica, CG-14 O. glaberrima, and WAB-12 interspecific). Decreased spikelet fertility and cultivar difference at high temperature were due mainly to decreased pollen production and pollen reception (pollen numbers on stigma). Lower spikelet fertility at elevated temperature resulted in fewer filled grains, lower grain weight per panicle, and decreased harvest index. There is a potential for genetic improvement for heat tolerance, thus it is important to screen and identify heat-tolerant cultivars. Spikelet fertility at high temperature can be used as a screening tool for heat tolerance during the reproductive phase.
DOI:10.1093/aob/mcf240URLPMID:4240456 [本文引用: 1]
Continuous exposure of tomato 'Trust' to high temperatures (day/night temperatures of 32/26 C) markedly reduced the number of pollen grains per flower and decreased viability. The effect of heat stress on pollen viability was associated with alterations in carbohydrate metabolism in various parts of the anther during its development. Under control, favourable temperature conditions (28/22 C), starch accumulated in the pollen grains, where it reached a maximum value 3 d before anthesis; it then diminished towards anthesis. During anther development, the concentration of total soluble sugars gradually increased in the anther walls and in the pollen grains (but not in the locular fluid), reaching a maximum at anthesis. Continuous exposure of the plants to high temperatures (32/26 C) prevented the transient increase in starch concentration and led to decreases in the concentrations of soluble sugars in the anther walls and the pollen grains. In the locular fluid, however, a higher soluble sugar concentration was detected under the high-temperature regime throughout anther development. These results suggest that a major effect of heat stress on pollen development is a decrease in starch concentration 3 d before anthesis, which results in a decreased sugar concentration in the mature pollen grains. These events possibly contribute to the decreased pollen viability in tomato.
[本文引用: 1]
DOI:10.1186/1471-2164-9-432URLPMID:18808683 [本文引用: 1]
pAbstract/p pBackground/p pWheat is a major crop in the world, and the high temperature stress can reduce the yield of wheat by as much as 15%. The molecular changes in response to heat stress are poorly understood. Using GeneChipsup /supWheat Genome Array, we analyzed genome-wide gene expression profiles in the leaves of two wheat genotypes, namely, heat susceptible Chinese Spring (CS) and heat tolerant TAM107 (TAM)./p pResults/p pA total of 6560 (~10.7%) probe sets displayed 2-fold or more changes in expression in at least one heat treatment (false discovery rate, FDR, = 0.001). Except for heat shock protein (HSP) and heat shock factor (HSF) genes, these putative heat responsive genes encode transcription factors and proteins involved in phytohormone biosynthesis/signaling, calcium and sugar signal pathways, RNA metabolism, ribosomal proteins, primary and secondary metabolisms, as well as proteins related to other stresses. A total of 313 probe sets were differentially expressed between the two genotypes, which could be responsible for the difference in heat tolerance of the two genotypes. Moreover, 1314 were differentially expressed between the heat treatments with and without pre-acclimation, and 4533 were differentially expressed between short and prolonged heat treatments./p pConclusion/p pThe differences in heat tolerance in different wheat genotypes may be associated with multiple processes and mechanisms involving HSPs, transcription factors, and other stress related genes. Heat acclimation has little effects on gene expression under prolonged treatments but affects gene expression in wheat under short-term heat stress. The heat stress responsive genes identified in this study will facilitate our understanding of molecular basis for heat tolerance in different wheat genotypes and future improvement of heat tolerance in wheat and other cereals./p
DOI:10.1186/s12864-018-4930-4URLPMID:30041596 [本文引用: 1]
In flowering plants, the male gametophyte (pollen) is one of the most vulnerable cells to temperature stress. InArabidopsis thaliana, a pollen-specificCyclicNucleotide-Gated cationChannel 16(cngc16), is required for plant reproduction under temperature-stress conditions. Plants harboring acncg16knockout are nearly sterile under conditions of hot days and cold nights. To understand the underlying cause, RNA-Seq was used to compare the pollen transcriptomes of wild type (WT) andcngc16under normal and heat stress (HS) conditions. Here we show that a heat-stress response (HSR) in WT pollen resulted in 2102 statistically significant transcriptome changes ( 2-fold changes with adjustedp-value 0.01), representing approximately 15% of 14,226 quantified transcripts. Of these changes, 89 corresponded to transcription factors, with 27 showing a preferential expression in pollen over seedling tissues. In contrast to WT,cngc16pollen showed 1.9-fold more HS-dependent changes (3936 total, with 2776 differences between WT andcngc16). In a quantitative direct comparison between WT andcngc16transcriptomes, the number of statistically significant differences increased from 21 pre-existing differences under normal conditions to 192 differences under HS. Of the 20 HS-dependent changes in WT that were most different incngc16, half corresponded to genes encoding proteins predicted to impact cell wall features or membrane dynamics. Results here define an extensive HS-dependent reprogramming of approximately 15% of the WT pollen transcriptome, and identify at least 27 transcription factor changes that could provide unique contributions to a pollen HSR. The number of statistically significant transcriptome differences between WT andcngc16increased by more than 9-fold under HS, with most of the largest magnitude changes having the potential to specifically impact cell walls or membrane dynamics, and thereby potentiatecngc16pollen to be hypersensitive to HS. However, HS-hypersensitivity could also be caused by the extensive number of differences throughout the transcriptome having a cumulative effect on multiple cellular pathways required for tip growth and fertilization. Regardless, results here support a model in which a functional HS-dependent reprogramming of the pollen transcriptome requires a specific calcium-permeableCyclic Nucleotide-Gated cation Channel, CNGC16. The online version of this article (10.1186/s12864-018-4930-4) contains supplementary material, which is available to authorized users.
DOI:10.1111/pce.12186URLPMID:23961845 [本文引用: 1]
Heat shock transcription factors (Hsfs) are involved in multiple aspects of stress response and plant growth. However, their role during male gametophyte development is largely unknown, although the generative phase is the most sensitive and critical period in the plant life cycle. Based on a wide screen of T-DNA mutant lines, we identified the atren1 mutation (restricted to nucleolus1) in early male gametophytic gene At1g77570, which has the closest homology to HSFA5 gene, the member of a heat shock transcription factor (HSF) gene family. The mutation causes multiple defects in male gametophyte development in both structure and function. Because the mutation disrupts an early acting (AtREN1) gene, these pollen phenotype abnormalities appear from bicellular pollen stage to pollen maturation. Moreover, the consequent progamic phase is compromised as well as documented by pollen germination defects and limited transmission via male gametophyte. In addition, atren1/ plants are defective in heat stress (HS) response and produce notably higher proportion of aberrant pollen grains. AtREN1 protein is targeted specifically to the nucleolus that, together with the increased size of the nucleolus in atren1 pollen, suggests that it is likely to be involved in ribosomal RNA biogenesis or other nucleolar functions.
DOI:10.1111/ppl.12759URLPMID:29766507 [本文引用: 1]
Abstract Heat stress at the pollen mother cell (PMC) meiotic stage leads to pollen sterility in rice, in which the reactive oxygen species (ROS) and sugar homeostasis are always adversely affected. This damage is reversed by abscisic acid (ABA), but the mechanisms underlying the interactions among the ABA, sugar metabolism, ROS and heat shock proteins (HSPs) in rice spikelets under heat stress are unclear. Two rice genotypes, Zhefu802 (a recurrent parent) and fgl (its near-isogenic line) were subjected to heat stress of 40 C after pre-foliage sprayed with ABA and its biosynthetic inhibitor fluridone at the meiotic stage of PMC. The results revealed that exogenous application of ABA reduced pollen sterility caused by heat stress. This was achieved through various means, including: increased levels of soluble sugars, starch and non-structural carbohydrates, markedly higher relative expression levels of heat shock proteins (HSP24.1 and HSP71.1) and genes related to sugar metabolism and transport, such as sucrose transporters (SUT) genes, sucrose synthase (SUS) genes, and invertase (INV) genes as well as increased antioxidant activities and increased content of ATP and endogenous ABA in spikelets. In short, exogenous application of ABA prior to heat stress enhanced sucrose transport and accelerated sucrose metabolism to maintain the carbon balance and energy homeostasis, thus ABA contributed to heat tolerance in rice. This article is protected by copyright. All rights reserved.
DOI:10.1105/tpc.108.065318URLPMID:19773386 [本文引用: 1]
Land plants are prone to strong thermal variations and must therefore sense early moderate temperature increments to induce appropriate cellular defenses, such as molecular chaperones, in anticipation of upcoming noxious temperatures. To investigate how plants perceive mild changes in ambient temperature, we monitored in recombinant lines of the moss Physcomitrella patens the activation of a heat-inducible promoter, the integrity of a thermolabile enzyme, and the fluctuations of cytoplasmic calcium. Mild temperature increments, or isothermal treatments with membrane fluidizers or Hsp90 inhibitors, induced a heat shock response (HSR) that critically depended on a preceding Ca0562 transient through the plasma membrane. Electrophysiological experiments revealed the presence of a Ca0562 -permeable channel in the plasma membrane that is transiently activated by mild temperature increments or chemical perturbations of membrane fluidity. The amplitude of the Ca0562 influx during the first minutes of a temperature stress modulated the intensity of the HSR, and Ca0562 channel blockers prevented HSR and the onset of thermotolerance. Our data suggest that early sensing of mild temperature increments occurs at the plasma membrane of plant cells independently from cytosolic protein unfolding. The heat signal is translated into an effective HSR by way of a specific membrane-regulated Ca0562 influx, leading to thermotolerance.
DOI:10.1104/pp.113.234401URLPMID:24569847 [本文引用: 1]
Microsporogenesis in rice (Oryza sativa) plants is susceptible to moderate low temperature (LT; approximately 19 C) that disrupts pollen development and causes severe reductions in grain yields. Although considerable research has been invested in the study of cool-temperature injury, a full understanding of the molecular mechanism has not been achieved. Here, we show that endogenous levels of the bioactive gibberellins (GAs) GA4 and GA7, and expression levels of the GA biosynthesis genes GA20ox3 and GA3ox1, decrease in the developing anthers by exposure to LT. By contrast, the levels of precursor GA12 were higher in response to LT. In addition, the expression of the dehydration-responsive element-binding protein DREB2B and SLENDER RICE1 (SLR1)/DELLA was up-regulated in response to LT. Mutants involved in GA biosynthetic and response pathways were hypersensitive to LT stress, including the semidwarf mutants sd1 and d35, the gain-of-function mutant slr1-d, and gibberellin insensitive dwarf1. The reduction in the number of sporogenous cells and the abnormal enlargement of tapetal cells occurred most severely in the GA-insensitive mutant. Application of exogenous GA significantly reversed the male sterility caused by LT, and simultaneous application of exogenous GA with sucrose substantially improved the extent of normal pollen development. Modern rice varieties carrying the sd1 mutation are widely cultivated, and the sd1 mutation is considered one of the greatest achievements of the Green Revolution. The protective strategy achieved by our work may help sustain steady yields of rice under global climate change.
DOI:10.1073/pnas.1000869107URL [本文引用: 1]
DOI:10.1016/j.jplph.2015.02.002URLPMID:25747289 [本文引用: 1]
Accumulation of compatible solutes is considered a key adaptation mechanism in many plants in response to abiotic stress. The expression of four genes, involved in sucrose metabolism (SPS and SuSy), biosynthesis of galactinol (GoLS1) and proline accumulation (P5CS) was compared: at meiosis (MM), vacuolated and mature stages of pollen development in heat tolerant and heat sensitive tomato genotypes. The results showed differences in gene expression across tomato genotypes and stages of pollen development. Three genes (P5CS, SPS and SuSy) were up regulated in heat tolerant genotype CLN1621L at the mature stage and one gene (P5CS) in genotype CLN5915-93D at the MM stage. Two genes (SPS and GoLS1) were down regulated in heat sensitive genotype CA4 and one gene (GoLS1) in genotype CLN2498E at the MM stage. Additionally, the continuous exposure of tomato genotypes to temperatures of 35 C/28 C day/night completely impaired flower development in genotypes CA4 and CLN2498E but not in genotypes CLN1621L and CLN5915-93D. Tomato genotypes CLN1621L and CLN5915-93D produced fully developed flowers containing mixture of non viable pollens and very few viable pollens grains. Membrane permeability was affected at all stages of development under heat stress with heat tolerant genotypes CL5915-93D4, CLN2498E and CLN1621L showing varying degrees of heat acclimation. Significant increases in total chlorophyll were seen in all genotypes in response to heat stress. The expression of compatible solute genes at MM is more critical than at mature stage for the development of viable pollen grain.
DOI:10.1093/aob/mcl037URLPMID:2803419 [本文引用: 1]
61 Background and Aims Global warming is gaining significance as a threat to natural and managed ecosystems since temperature is one of the major environmental factors affecting plant productivity. Hence, the effects of moderate temperature increase on the growth and development of the tomato plant (Lycopersicon esculentum) were investigated. 61 Methods Plants were grown at 32/26°C as a moderately elevated temperature stress (METS) treatment or at 28/22°C (day/night temperatures) as a control with natural light conditions. Vegetative growth and reproductive development as well as sugar content and metabolism, proline content and translocation in the androecium were investigated. 61 Key Results METS did not cause a significant change in biomass, the number of flowers, or the number of pollen grains produced, but there was a significant decrease in the number of fruit set, pollen viability and the number of pollen grains released. Glucose and fructose contents in the androecium (i.e. all stamens from one flower) were generally higher in the control than METS, but sucrose was higher in METS. Coincidently, the mRNA transcript abundance of acid invertase in the androecium was decreased by METS. Proline contents in the androecium were almost the same in the control and METS, while the mRNA transcript level of proline transporter 1, which expresses specifically at the surface of microspores, was significantly decreased by METS. 61 Conclusions The research indicated that failure of tomato fruit set under a moderately increased temperature above optimal is due to the disruption of sugar metabolism and proline translocation during the narrow window of male reproductive development.
[本文引用: 1]
DOI:10.1111/pce.13146URLPMID:29336039 [本文引用: 1]
Abstract High temperature during flowering in rice causes spikelet sterility and is a major threat to rice productivity in tropical and subtropical regions, where hybrid rice development is increasingly contributing to sustain food security. However, the sensitivity of hybrids to increasing temperature and physiological responses in terms of dynamic fertilization processes are unknown. To address these questions, several promising hybrids and inbreds were exposed to control and high day-time temperature (HDT) in Experiment 1 and hybrids having contrasting heat tolerance were selected for Experiment 2 for further physiological investigation under high day and night temperature treatments. The day-time temperature played a dominant role in determining spikelet fertility compared with night-time temperature. HDT significantly induced spikelet sterility in tested hybrids and hybrids had higher heat susceptibility than the high-yielding inbred varieties. Poor pollen germination was strongly associated with sterility under high temperature. Our novel observations capturing the series of dynamic fertilization processes demonstrated that pollen tubes not reaching the viable embryo sac was the major cause for spikelet sterility under heat exposure. Our findings highlight the urgent need to improve heat tolerance in hybrids and incorporating early-morning flowering as a promising trait for mitigating HDT stress impact at flowering.
DOI:10.4161/psb.6.7.15315URLPMID:3257764 [本文引用: 1]
Reproductive development in sexual plants is substantially more sensitive to high temperature stress than vegetative development, resulting in negative implications for food and fiber production under the moderate temperature increases projected to result from global climate change. High temperature exposure either during early pollen development or during the progamic phase of pollen development will negatively impact pollen performance and reproductive output; both phases of pollen development are considered exceptionally sensitive to moderate heat stress. However, moderately elevated temperatures either before or during the progamic phase can limit fertilization by negatively impacting important pollen pistil interactions required for successful pollen tube growth toward the ovules. This mini-review identifies the impacts of heat stress on pollen-pistil interactions and sexual reproduction in angiosperms. A special emphasis is placed on the biochemical response of the pistil to moderately high temperature and the resultant influence on in vivo pollen performance and fertilization.
DOI:10.1016/j.jplph.2010.12.011URLPMID:21256621 [本文引用: 1]
It has recently been reported that high temperature slows in vivo pollen tube growth rates in Gossypium hirsutum pistils under field conditions. Although numerous physical and biochemical pollen istil interactions are necessary for in vivo pollen tube growth to occur, studies investigating the influence of heat-induced changes in pistil biochemistry on in vivo pollen tube growth rates are lacking. We hypothesized that high temperature would alter diurnal pistil biochemistry and that pollen tube growth rates would be dependent upon the soluble carbohydrate content of the pistil during pollen tube growth. G. hirsutum seeds were sown on different dates to obtain flowers exposed to contrasting ambient temperatures but at the same developmental stage. Diurnal pistil measurements included carbohydrate balance, glutathione reductase (GR; EC 1.8.1.7), soluble protein, superoxide dismutase (SOD; EC 1.15.1.1), NADPH oxidase (NOX; EC 1.6.3.1), adenosine triphosphate (ATP), and water-soluble calcium. Soluble carbohydrate levels in cotton pistils were as much as 67.5% lower under high temperature conditions (34.6 C maximum air temperature; August 4, 2009) than under cooler conditions (29.9 C maximum air temperature; August 14, 2009). Regression analysis revealed that pollen tube growth rates were highly correlated with the soluble carbohydrate content of the pistil during pollen tube growth ( r 2 = 0.932). Higher ambient temperature conditions on August 4 increased GR activity in the pistil only during periods not associated with in vivo pollen tube growth; pistil protein content declined earlier in the day under high temperatures; SOD and NOX were unaffected by either sample date or time of day; pistil ATP and water soluble calcium were unaffected by the warmer temperatures. We conclude that moderate heat stress significantly alters diurnal carbohydrate balance in the pistil and suggest that pollen tube growth rate through the style may be limited by soluble carbohydrate supply in the pistil.
DOI:10.1093/jxb/ers298URLPMID:3504494 [本文引用: 1]
Stress-induced plant cell reprogramming involves changes in global genome organization, being the epigenetic modifications key factors in the regulation of genome flexibility. DNA methylation, accomplished by DNA methyltransferases, constitutes a prominent epigenetic modification of the chromatin fibre which is locked in a transcriptionally inactive conformation. Changes in DNA methylation accompany the reorganization of the nuclear architecture during plant cell differentiation and proliferation. After a stress treatment,in vitro-cultured microspores are reprogrammed and change their gametophytic developmental pathway towards embryogenesis, the process constituting a useful system of reprogramming in isolated cells for applied and basic research. Gene expression driven by developmental and stress cues often depends on DNA methylation; however, global DNA methylation and genome-wide expression patterns relationship is still poorly understood. In this work, the dynamics of DNA methylation patterns in relation to nuclear architecture and the expression of BnMET1a-likeDNA methyltransferase genes have been analysed during pollen development and pollen reprogramming to embryogenesis inBrassica napusL. by a multidisciplinary approach. Results showed an epigenetic reprogramming after microspore embryogenesis induction which involved a decrease of global DNA methylation and its nuclear redistribution with the change of developmental programme and the activation of cell proliferation, while DNA methylation increases with pollen and embryo differentiation in a cell-type-specific manner. Changes in the presence, abundance, and distribution of BnMET1a-liketranscripts highly correlated with variations in DNA methylation. Mature zygotic and pollen embryos presented analogous patterns of DNA methylation andMET1a-likeexpression, providing new evidence of the similarities between both developmental embryogenic programmes.
DOI:10.1007/s00425-015-2259-7URLPMID:25672505 [本文引用: 1]
Main conclusion Pollen tube growth in styles was strongly inhibited by temperature above 35 C, and the yield of cotton decreased because of the adverse effect of high temperatures during square development. High-temperature stress during flowering influences the square development of upland cotton ( Gossypium hirsutum L.) and cotton yield. Although it is well known that square development is sensitive to high temperature, high-temperature sensitive stages of square development and the effects of high temperature on pollen tube growth in the styles are unknown. The effect of high temperature on anther development corresponding to pollen vigor is unknown during anther development. The objectives of this study were to identify the stages of square development that are sensitive to high temperatures (37/30 and 40/34 C), to determine whether the abnormal development of squares influenced by high temperature is responsible for the variation in the in vitro germination percent of pollen grains at anthesis, to identify the effect of high temperature on pollen germination in the styles, and to determine pollen thermotolerance heterosis. Our results show that the stages from the sporogenous cell to tetrad stage (square length <6.0mm) were the most sensitive to high temperature, and the corresponding pollen viability at anthesis was consistent with the changes in the square development stage. Pollen tube growth in the styles was strongly inhibited by temperature above 35 C, and the yield of cotton decreased because of the effect of high temperature during square development. The thermotolerance of hybrid F 1 pollen showed heterosis, and pollen viability could be used as a criterion for screening for high-temperature tolerance cultivars. These results can be used in breeding to develop new cotton cultivars that can withstand high-temperature conditions, particularly in a future warmer climate.
DOI:10.1007/s004970100071URL [本文引用: 1]
Pollen sterility was induced by heat stress applied about 10 days before flowering in the snap bean Phaseolus vulgaris L. Cytohistological changes in the tapetum during early development of the anther were studied to identify the tissues most sensitive to high temperature stress. The first distinct structural abnormalities were detected in the distribution pattern of the rough endoplasmic reticulum (RER) in the tapetum at the early microspore stage under high temperature conditions. Stacks of RER were frequently observed in the tapetum under optimal conditions, but rarely occurred under high temperature conditions. Various patterns of endoplasmic reticulum (ER) arrangement linear, wavy, looped or circular were observed in the tapetum. Two types of circular ER were observed at the microspore stage under high temperature conditions, RER with ribosomes on the surface and smooth endoplasmic reticulum (SER) lacking ribosomes. The tapetum underwent degenerative changes under high temperature conditions earlier than under optimal conditions. The structural abnormalities of the microspore were associated with tapetal degeneration. We concluded that high air temperature affected the ER structure and blocked its function in the tapetum, and then induced earlier than usual degeneration of tapetum. Pollen sterility is associated with tapetal degeneration.
DOI:10.1007/s10725-007-9225-8URL [本文引用: 1]
In order to explore the possible physiological mechanism of high temperature induced sterility in rice, we examined the floret sterility and endogenous plant growth regulator contents in pollens of two hybrid rice cultivars Shanyou63 and Teyou559 that are tolerant and susceptible to high temperature, respectively. Indexes of floret sterility, pollen activity, and variation of endogenous indole-3-acetic acid (IAA), gibberellic acids (GAs), abscisic acid (ABA), free proline and soluble proteins in anthers were measured. We found that during the course of high temperature treatment, both cultivars exhibited a marked decrease in pollen activity, pollen germination and floret fertility; however, the high temperature tolerant Shanyou63 showed a much slower rate of decrease than the high temperature susceptible Teyou559. In addition, anthers of both cultivars displayed a decrease in the contents of IAA, GAs, free proline and soluble proteins but an increase in the ABA content. Yet compared to Teyou559, Shanyou63 retained significantly higher levels of free praline and GAs and a lower level of ABA, along with higher pollen vigour and pollen germination rate even after prolonged high temperature treatment. Our study suggests a possible correlation between pollen viability/floret sterility and high temperature-caused changes in IAA, GAs, ABA, free proline and soluble protein contents. The severity in these changes may reflect the variation of rice cultivars in their heat stress sensitivities for floret development.
DOI:10.1104/pp.112.206888URLPMID:23370720 [本文引用: 1]
Cyclic nucleotide-gated channels (CNGCs) have been implicated in diverse aspects of plant growth and development, including responses to biotic and abiotic stress, as well as pollen tube growth and fertility. Here, genetic evidence identifies CNGC16 in Arabidopsis (Arabidopsis thaliana) as critical for pollen fertility under conditions of heat stress and drought. Two independent transfer DNA disruptions of cngc16 resulted in a greater than 10-fold stress-dependent reduction in pollen fitness and seed set. This phenotype was fully rescued through pollen expression of a CNGC16 transgene, indicating that cngc16-1 and 16-2 were both loss-of-function null alleles. The most stress-sensitive period for cngc16 pollen was during germination and the initiation of pollen tube tip growth. Pollen viability assays indicate that mutant pollen are also hypersensitive to external calcium chloride, a phenomenon analogous to calcium chloride hypersensitivities observed in other cngc mutants. A heat stress was found to increase concentrations of 3',5'-cyclic guanyl monophosphate in both pollen and leaves, as detected using an antibody-binding assay. A quantitative PCR analysis indicates that cngc16 mutant pollen have attenuated expression of several heat-stress response genes, including two heat shock transcription factor genes, HsfA2 and HsfB1. Together, these results provide evidence for a heat stress response pathway in pollen that connects a cyclic nucleotide signal, a Ca2+-permeable ion channel, and a signaling network that activates a downstream transcriptional heat shock response.
DOI:10.1007/s00497-010-0157-5URLPMID:21103996 [本文引用: 1]
During angiosperm male gametophyte development, the male germline is segregated by an asymmetric cell division of the haploid microspore. This review encompasses recent advances in understanding the genetic and molecular mechanisms involved in generating the male germline from this pluripotent germline initial and in specifying the production of the twin sperm cells required for double fertilization. Genetic studies and access to the transcriptome of isolated gametes have enabled remarkable progress in understanding some of the key regulators that control and integrate germ cell cycle progression with germline specification, and an emerging regulatory model is presented. Rapid advances have also been made in understanding epigenetic regulation and small RNA pathways in the male gametophyte and germline that impact on genome integrity and gamete development, traits that are shared with animal germlines. The review concludes with a perspective of the outstanding issues and directions of future research that will further our understanding of germline specification and the gametophytic control of pollen development.
DOI:10.1186/s12870-016-0771-yURLPMID:27079791 [本文引用: 1]
Being sessile organisms, plants are often exposed to a wide array of abiotic and biotic stresses. Abiotic stress conditions include drought, heat, cold and salinity, whereas biotic stress arises mainly from bacteria, fungi, viruses, nematodes and insects. To adapt to such adverse situations, plants have evolved well-developed mechanisms that help to perceive the stress signal and enable optimal growth response. Phytohormones play critical roles in helping the plants to adapt to adverse environmental conditions. The elaborate hormone signaling networks and their ability to crosstalk make them ideal candidates for mediating defense responses. Recent research findings have helped to clarify the elaborate signaling networks and the sophisticated crosstalk occurring among the different hormone signaling pathways. In this review, we summarize the roles of the major plant hormones in regulating abiotic and biotic stress responses with special focus on the significance of crosstalk between different hormones in generating a sophisticated and efficient stress response. We divided the discussion into the roles of ABA, salicylic acid, jasmonates and ethylene separately at the start of the review. Subsequently, we have discussed the crosstalk among them, followed by crosstalk with growth promoting hormones (gibberellins, auxins and cytokinins). These have been illustrated with examples drawn from selected abiotic and biotic stress responses. The discussion on seed dormancy and germination serves to illustrate the fine balance that can be enforced by the two key hormones ABA and GA in regulating plant responses to environmental signals. The intricate web of crosstalk among the often redundant multitudes of signaling intermediates is just beginning to be understood. Future research employing genome-scale systems biology approaches to solve problems of such magnitude will undoubtedly lead to a better understanding of plant development. Therefore, discovering additional crosstalk mechanisms among various hormones in coordinating growth under stress will be an important theme in the field of abiotic stress research. Such efforts will help to reveal important points of genetic control that can be useful to engineer stress tolerant crops.
DOI:10.1007/s11103-005-0339-yURLPMID:15821976 [本文引用: 1]
Abstract In plants small heat shock proteins (sHsp) are abundantly expressed upon heat stress in vegetative tissue, however, sHsp expression is also developmentally induced in pollen. The developmental induction of sHsp has been related to the potential for stress-induced microspore embryogenesis. We investigated the polymorphism among sHsp and their expression during pollen development and after heat stress in tobacco. Real-time RT-PCR was used for quantification of mRNA of two known and nine newly isolated cDNAs representing cytosolic sHsp. At normal temperature most of these genes are not transcribed in vegetative tissues, however, all genes were expressed during pollen development. Low levels of mRNAs were found for sHsp-1A and -1B in early-unicellular stage, increasing four to sevenfold in mature pollen. Nine other genes are up-regulated in unicellular and down-regulated in bicellular pollen; three these genes show stage-specific expression. Western analysis revealed that cytosolic class I and II sHsp are developmentally expressed during all stages of pollen development. Different subsets of cytosolic sHsp genes are expressed in a stage-specific fashion suggesting that certain sHsp genes may play specific roles in early, others during later stages of pollen development. Heat stress results in a relatively weak and incomplete response in pollen: (i) the heat-induced levels of mRNA (excepting sHsp-2B, -3C and -6) are much lower than in leaves, (ii) several sHsp are not detected after heat stress in pollen, although, they are heat-inducibly expressed in leaves. Application of heat stress, cold, and starvation, which induce microspore embryogenesis, modify mRNA levels and the patterns of 2-D-separated sHsp, but only heat stress enhances the expression of sHsp in microspores. There is no correlation of the expression of specific sHsp with the potential for microspore embryogenesis.
DOI:10.1038/s41598-017-05661-xURLPMID:28706219 [本文引用: 1]
Abstract Temperature change is of potential to trigger the formation of unreduced gametes. In this study, we showed that short periods of high temperature treatment can induce the production of 2n pollen in Populus pseudo-simonii Kitag. The meiotic stage, duration of treatment, and temperature have significant effects on the induction of 2n pollen. Heat stress resulted in meiotic abnormalities, including failure of chromosome separation, chromosome stickiness, laggards and micronuclei. Spindle disorientations in the second meiotic division, such as parallel, fused, and tripolar spindles, either increased in frequency or were induced de novo by high temperature treatment. We found that the high temperature treatment induced depolymerisation of meiotic microtubular cytoskeleton, resulting in the failure of chromosome segregation. New microtubular cytoskeletons were able to repolymerise in some heat-treated cells after transferring them to normal conditions. However, aberrant cytokinesis occurred owing to defects of new radial microtubule systems, leading to production of monads, dyads, triads, and polyads. This suggested that depolymerisation and incomplete restoration of microtubules may be important for high temperature-induction of unreduced gametes. These findings might help us understand how polyploidisation is induced by temperature-related stress and support the potential effects of global climate change on reproductive development of plants.
[本文引用: 1]
DOI:10.1186/s12870-014-0327-yURLPMID:4262988 [本文引用: 1]
Background To accelerate the application of the CRISPR/Cas9 (clustered regularly interspaced short palindromic repeats/ CRISPR-associated protein 9) system to a variety of plant species, a toolkit with additional plant selectable markers, more gRNA modules, and easier methods for the assembly of one or more gRNA expression cassettes is required. Results We developed a CRISPR/Cas9 binary vector set based on the pGreen or pCAMBIA backbone, as well as a gRNA (guide RNA) module vector set, as a toolkit for multiplex genome editing in plants. This toolkit requires no restriction enzymes besides BsaI to generate final constructs harboring maize-codon optimized Cas9 and one or more gRNAs with high efficiency in as little as one cloning step. The toolkit was validated using maize protoplasts, transgenic maize lines, and transgenic Arabidopsis lines and was shown to exhibit high efficiency and specificity. More importantly, using this toolkit, targeted mutations of three Arabidopsis genes were detected in transgenic seedlings of the T1 generation. Moreover, the multiple-gene mutations could be inherited by the next generation. Conclusions We developed a toolkit that facilitates transient or stable expression of the CRISPR/Cas9 system in a variety of plant species, which will facilitate plant research, as it enables high efficiency generation of mutants bearing multiple gene mutations.
DOI:10.1007/s11032-017-0664-2URLPMID:5395597 [本文引用: 1]
Global warming has become a worldwide concern due to its adverse effects on agricultural output. In particular, long-term mildly high temperatures interfere with sexual reproduction and thus fruit and seed set. To uncover the genetic basis of observed variation in tolerance against heat, a bi-parental F2mapping population from two contrasting cultivars, i.e. Nagcarlang and NCHS-1, was generated and phenotyped under continuous mild heat conditions for a number of traits underlying reproductive success, i.e. pollen viability, pollen number, style length, anther length, style protrusion, female fertility and flowering characteristics, i.e. inflorescence number and flowers per inflorescence. Quantitative trait loci (QTLs) were identified for most of these traits, including a single, highly significant one for pollen viability, which accounted for 36% of phenotypic variation in the population and modified pollen viability under high temperature with around 20%. QTLs for some traits colocalised, indicating trait dependency or pleiotropic-effect loci. We conclude that a limited set of major genes determines differences in performance of reproductive traits under continuous mild heat in tomato. The results contribute to our fundamental understanding of pollen thermotolerance and may support development of more heat-tolerant tomato varieties. The online version of this article (doi:10.1007/s11032-017-0664-2) contains supplementary material, which is available to authorized users.
.
[本文引用: 1]
[本文引用: 1]
DOI:10.1186/s12284-018-0206-5URLPMID:29532187 [本文引用: 1]
Background Pollen tube elongation in the pistil is a key step for pollination success in plants, and auxins play an important role in this process. However, the function of auxins in pollen tube...
DOI:10.1016/j.jgg.2011.08.001URLPMID:21930097 [本文引用: 1]
Microsporogenesis and male gametogenesis are essential for the alternating life cycle of flowering plants between diploid sporophyte and haploid gametophyte generations. Rice ( Oryza sativa) is the world major staple food, and manipulation of pollen fertility is particularly important for the demands to increase rice grain yield. Towards a better understanding of the mechanisms controlling rice male reproductive development, we describe here the cytological changes of anther development through 14 stages, including cell division, differentiation and degeneration of somatic tissues consisting of four concentric cell layers surrounding and supporting reproductive cells as they form mature pollen grains through meiosis and mitosis. Furthermore, we compare the morphological difference of anthers and pollen grains in both monocot rice and eudicot Arabidopsis thaliana. Additionally, we describe the key genes identified to date critical for rice anther development and pollen formation.
DOI:10.1104/pp.108.133744URL [本文引用: 1]
DOI:10.1007/s00299-018-2264-yURLPMID:29464319 [本文引用: 2]
Key message HT-induced ROS burst in developing anther is closely related to the lowered CAT activity as the result of the markedly suppressed OsCATBtranscript, thereby causing severe fertility...
DOI:10.1016/j.plaphy.2017.11.009URLPMID:29202329 [本文引用: 1]
Anther culture in the breeding process of winter wheat. III. Ability of winter wheat F1 populations with the two heterozygous 1AL–IAS/1AL–IRS and 1BL–1BS/1BL–IRS chromosome pairs Application of anther culture to four F1 hybrids between the IBL–IRS (‘Amigo’) and several 1BL–IRS wheat-rye translocation forms yielded 129 green pollen plants in an average embryo induction frequency of 17.6 %. A... [Show full abstract]
DOI:10.1093/jxb/erq053URL [本文引用: 1]
被子植物生殖细胞与精细胞的分离方法
1
2014
... 随着全球气候逐渐变暖, 温度变化对生态环境的影响日益明显(
Thermal stress impacts reproduce- tive development and grain yield in rice
1
2017
... 在进行花粉高温胁迫研究时, 需要考虑胁迫温度、处理方式以及花粉取材时期.首先需要选择合适的温度处理范围, 不同的温度处理会产生不同的结果.温和高温可以使植物获得高温耐受能力, 严重高温则直接破坏植物细胞, 导致植株不可逆损伤甚至死亡.通过总结常见作物的花粉高温胁迫研究, 发现常见作物的高温胁迫温度通常选择30-39°C.尽管番茄花粉高温胁迫研究也有选择45°C高温的情况, 但处理时间相对较短(
The effect of drought and heat stress on reproductive processes in cereals
1
2008
... 通常情况下, 在高于最适温度5°C以上的环境下, 花粉就会遭受高温伤害.许多重要作物如小麦(Tri- ticum aestivum)、水稻(Oryza sativa)和玉米(Zea mays), 其开花期均集中在夏季, 极易遭遇连续或严重的高温天气, 从而导致花粉发育异常(
Tracking maize pollen development by the Leaf Collar Method
1
2017
... 在设计花粉高温胁迫相关实验时, 还需要考虑花粉所处的发育阶段.不同发育阶段的花粉对高温胁迫的敏感性和响应机制可能不同.在实际研究中, 通常把花粉发育过程分为4个阶段: (1) 花粉母细胞到四分体时期; (2) 小孢子形成到早期的二细胞时期; (3) 二细胞末期/三细胞期到成熟花粉期; (4) 花粉萌发期.目前, 可以通过一些形态指标对常见作物花粉发育阶段进行判断.例如, 水稻花粉的发育阶段可以通过花芽长度进行判断; 而玉米花粉的发育阶段则可以通过茎秆的叶领(leaf collar)数进行判断(
Thermal stress effects on grain yield in Brachypodium distachyon occur via H2A.Z-nucleosomes
1
2013
... H2A.Z是一种组蛋白变体, 包含H2A.Z的核小体与DNA结合更加紧密.研究表明, 高温处理可以减少含有H2A.Z的核小体在基因转录起始位点的占有率, 从而调控相关基因的表达(
Identification of novel small ncRNAs in pollen of tomato
1
2015
... 研究作物花粉高温胁迫应答机制并提高作物花粉的高温耐受能力, 对于稳定和提高作物产量具有重要意义.目前, 人们已对高温导致的花粉败育进行了系统的形态学观察和多种组学分析, 并且已鉴定到一些参与花粉高温胁迫响应的因子, 但对于花粉高温胁迫应答调控机制的认知仍然有限(
Enhancement of reproductive heat tolerance in plants
1
2015
... 花粉感知环境中的高温信号后, 通过信号转导引起代谢、转录甚至表观遗传水平的改变, 从而实现对高温胁迫的响应.其中热激转录因子是高温胁迫应答网络中的关键组分, 可以诱导高温胁迫应答重要效应物热激蛋白的表达, 通过维持蛋白的构型稳定减少由高温导致的蛋白变性和聚集, 从而提高植物耐热性.在烟草(Nicotiana tabacum)花粉发育过程中, 高温诱导不同的HSPs表达(
Heat-treatment-responsive proteins in different developmental stages of tomato pollen detected by targeted mass accuracy precursor alignment (tMAPA)
1
2015
... 尽管目前对花粉高温胁迫响应机制认识有限, 但是随着各种组学技术、全基因组关联分析(genome- wide association study, GWAS)和CRISPR-Cas9等基因编辑技术在花粉高温胁迫研究中的应用(
Epigenetic events in plant male germ cell heat stress responses
2
2016
... 研究作物花粉高温胁迫应答机制并提高作物花粉的高温耐受能力, 对于稳定和提高作物产量具有重要意义.目前, 人们已对高温导致的花粉败育进行了系统的形态学观察和多种组学分析, 并且已鉴定到一些参与花粉高温胁迫响应的因子, 但对于花粉高温胁迫应答调控机制的认知仍然有限(
... ;
Resilience of rice ( Oryza spp.) pollen germination and tube growth to temperature stress
1
2016
... 高温通过改变花粉和柱头细胞中生长素和糖类的平衡影响花粉管在柱头内的生长, 表现出花粉萌发失败以及花粉管末端膨大、扭曲和破裂等现象, 最终导致受精过程无法完成(
Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants
1
2006
... 综上, 在应对高温胁迫时, 花粉至少可以通过4种机制感受高温信号并利用多种机制响应胁迫, 这些机制和途径之间也存在“交叉对话”.例如, 内流的钙离子可以参与多个途径; ROS可以影响UPR; ABA则影响糖类代谢; 而糖信号通路又可以影响ROS的平衡.这些机制之间的相互作用也增加了花粉高温胁迫研究的复杂性(
The Arabidopsis mutantjason produces unreduced first division restitution male gametes through a parallel/fused spindle mechanism in meiosis II
1
2011
... 高温影响花粉发育全过程, 导致花粉结构和发育异常, 产生没有活性的花粉, 并最终导致受精失败(
The impact of environmental stress on male reproductive development in plants: biological processes and molecular mechanisms
4
2014
... 高温影响花粉发育全过程, 导致花粉结构和发育异常, 产生没有活性的花粉, 并最终导致受精失败(
... ), 推测高温影响这3个基因的表达, 导致产生异常倍性的小孢子(
... 高温还可以促进绒毡层细胞提前发生程序性死亡(programmed cell death, PCD), 进而影响花粉发育、活性以及散粉过程(
... 研究作物花粉高温胁迫应答机制并提高作物花粉的高温耐受能力, 对于稳定和提高作物产量具有重要意义.目前, 人们已对高温导致的花粉败育进行了系统的形态学观察和多种组学分析, 并且已鉴定到一些参与花粉高温胁迫响应的因子, 但对于花粉高温胁迫应答调控机制的认知仍然有限(
IRE1, a component of the unfolded protein response signaling pathway, protects pollen development in Arabidopsis from heat stress
1
2016
... 绒毡层或者花粉细胞内的UPR反应也可以感受高温胁迫, 并通过ER-UPR (内质网中未折叠蛋白反应)提高内质网对蛋白质的折叠能力, 降低未折叠蛋白质的数量, 从而提高花粉的胁迫耐受能力.一些转录因子和RNA剪切因子也参与ER-UPR, 并在花粉发育过程中发挥重要作用, 其中erdj2a、bip1/2/3和pod1等突变体均表现出花粉发育异常表型(
Mutations in AtPS1 (Arabidopsis thaliana parallel spindle 1) lead to the production of diploid pollen grains
1
2008
... 高温影响花粉发育全过程, 导致花粉结构和发育异常, 产生没有活性的花粉, 并最终导致受精失败(
microRNAs involved in auxin signaling modulate male sterility under high-tem- perature stress in cotton (Gossypium hirsutum)
1
2017
... 高温胁迫下, 植物体内的激素水平也会发生显著变化(
Short periods of high temperature during meiosis prevent normal meiotic progression and reduce grain number in hexaploid wheat (Triticum aestivum L.)
1
2017
... 高温影响花粉发育全过程, 导致花粉结构和发育异常, 产生没有活性的花粉, 并最终导致受精失败(
High temperatures cause male sterility in rice plants with transcriptional alterations during pollen development
2
2009
... 通常情况下, 在高于最适温度5°C以上的环境下, 花粉就会遭受高温伤害.许多重要作物如小麦(Tri- ticum aestivum)、水稻(Oryza sativa)和玉米(Zea mays), 其开花期均集中在夏季, 极易遭遇连续或严重的高温天气, 从而导致花粉发育异常(
... 高温还可以促进绒毡层细胞提前发生程序性死亡(programmed cell death, PCD), 进而影响花粉发育、活性以及散粉过程(
A novel pollen-pistil interaction conferring high-temperature tolerance during reproduction via CLE45 signaling
1
2013
... 高温通过改变花粉和柱头细胞中生长素和糖类的平衡影响花粉管在柱头内的生长, 表现出花粉萌发失败以及花粉管末端膨大、扭曲和破裂等现象, 最终导致受精过程无法完成(
Heat stress in wheat during reproductive and grain-filling phases
1
2011
... 通常情况下, 在高于最适温度5°C以上的环境下, 花粉就会遭受高温伤害.许多重要作物如小麦(Tri- ticum aestivum)、水稻(Oryza sativa)和玉米(Zea mays), 其开花期均集中在夏季, 极易遭遇连续或严重的高温天气, 从而导致花粉发育异常(
HOP3, a member of the HOP family in Arabidopsis, interacts with BiP and plays a major role in the ER stress response
1
2017
... 绒毡层或者花粉细胞内的UPR反应也可以感受高温胁迫, 并通过ER-UPR (内质网中未折叠蛋白反应)提高内质网对蛋白质的折叠能力, 降低未折叠蛋白质的数量, 从而提高花粉的胁迫耐受能力.一些转录因子和RNA剪切因子也参与ER-UPR, 并在花粉发育过程中发挥重要作用, 其中erdj2a、bip1/2/3和pod1等突变体均表现出花粉发育异常表型(
Ethylene is involved in maintaining tomato (Solanum lycopersicum) pollen quality under heat-stress conditions
2
2012
... 高温胁迫下, 植物体内的激素水平也会发生显著变化(
... ;
Pollen grains of heat tolerant tomato cultivars retain higher carbohydrate concentration under heat stress conditions
1
2006
... 花粉作为有性生殖过程中光合产物重要的“库”, 其淀粉含量通常在二细胞花粉期达到最高值, 而可溶性糖的含量在散粉期达到最高值.积累的糖类物质不仅为花粉发育和花粉管形成提供能量, 也可以作为渗透物质提高花粉对胁迫的响应.通过比较不同的番茄品种可以发现, 耐高温品种的花粉中含有更多的淀粉和可溶性糖(
Unfolded protein response in pollen development and heat stress tolerance
1
2016
... 绒毡层或者花粉细胞内的UPR反应也可以感受高温胁迫, 并通过ER-UPR (内质网中未折叠蛋白反应)提高内质网对蛋白质的折叠能力, 降低未折叠蛋白质的数量, 从而提高花粉的胁迫耐受能力.一些转录因子和RNA剪切因子也参与ER-UPR, 并在花粉发育过程中发挥重要作用, 其中erdj2a、bip1/2/3和pod1等突变体均表现出花粉发育异常表型(
HsfA2 controls the activity of developmentally and stress-regulated heat stress protection mechanisms in tomato male reproductive tissues
2
2016
... 花粉感知环境中的高温信号后, 通过信号转导引起代谢、转录甚至表观遗传水平的改变, 从而实现对高温胁迫的响应.其中热激转录因子是高温胁迫应答网络中的关键组分, 可以诱导高温胁迫应答重要效应物热激蛋白的表达, 通过维持蛋白的构型稳定减少由高温导致的蛋白变性和聚集, 从而提高植物耐热性.在烟草(Nicotiana tabacum)花粉发育过程中, 高温诱导不同的HSPs表达(
... ;
Prospects of engineering thermotolerance in crops through modulation of heat stress transcription factor and heat shock protein networks
1
2015
... 在进行花粉高温胁迫研究时, 需要考虑胁迫温度、处理方式以及花粉取材时期.首先需要选择合适的温度处理范围, 不同的温度处理会产生不同的结果.温和高温可以使植物获得高温耐受能力, 严重高温则直接破坏植物细胞, 导致植株不可逆损伤甚至死亡.通过总结常见作物的花粉高温胁迫研究, 发现常见作物的高温胁迫温度通常选择30-39°C.尽管番茄花粉高温胁迫研究也有选择45°C高温的情况, 但处理时间相对较短(
Pollen tetrad-based visual assay for meiotic recombination in Arabidopsis
2
2007
... 高温影响花粉发育全过程, 导致花粉结构和发育异常, 产生没有活性的花粉, 并最终导致受精失败(
... ).染色体倍性异常的小孢子, 如二价体(diploid dyads)的产生则是由于高温导致维管骨架解聚以及纺锤体错误排列引起的(
Transcriptional profiling of maturing tomato (Solanum lycopersicum L.) microspores reveals the involvement of heat shock proteins, ROS scavengers, hormones, and sugars in the heat stress response
3
2009
... ROS是一类化学性质活泼且具有较高氧化活性的分子, 高温胁迫会导致植物体内产生大量ROS.积累的ROS不仅会造成核酸和蛋白质等重要生物大分子的氧化损伤, 也可以作为信号分子激活植物体内的抗氧化系统, 从而调控植物多个器官(包括花粉)响应高温胁迫.对小麦花粉进行高温处理(42°C, 2小时)后, 发现花粉中ROS和超氧化物歧化酶(superoxide dismutase, SOD)、过氧化物酶(peroxidase, POD)和过氧化氢酶(catalase, CAT)含量显著升高, 同时伴随着热激蛋白(heat shock proteins, HSPs) HSP70、HSP90、HSP17和HSP26含量增加(Kumar et al., 2013).番茄(Lycopersicon esculentum)和水稻花粉中也有类似机制(
... 花粉感知环境中的高温信号后, 通过信号转导引起代谢、转录甚至表观遗传水平的改变, 从而实现对高温胁迫的响应.其中热激转录因子是高温胁迫应答网络中的关键组分, 可以诱导高温胁迫应答重要效应物热激蛋白的表达, 通过维持蛋白的构型稳定减少由高温导致的蛋白变性和聚集, 从而提高植物耐热性.在烟草(Nicotiana tabacum)花粉发育过程中, 高温诱导不同的HSPs表达(
... 活性氧清除系统、激素以及糖类物质在花粉的高温应答中也发挥重要作用(
A heat-activated calcium-perme- able channel—Arabidopsis cyclic nucleotide-gated ion channel 6—is involved in heat shock responses
2
2012
... 质膜上的钙离子通道是植物细胞重要的高温感受器.电化学和生化等实验证实, 温度升高可以触发质膜上钙离子通道打开, 导致胞外钙离子内流, 使胞质内钙离子增加, 引起细胞产生热激反应(heat shock response, HSR) (
... 花粉感知环境中的高温信号后, 通过信号转导引起代谢、转录甚至表观遗传水平的改变, 从而实现对高温胁迫的响应.其中热激转录因子是高温胁迫应答网络中的关键组分, 可以诱导高温胁迫应答重要效应物热激蛋白的表达, 通过维持蛋白的构型稳定减少由高温导致的蛋白变性和聚集, 从而提高植物耐热性.在烟草(Nicotiana tabacum)花粉发育过程中, 高温诱导不同的HSPs表达(
Developmental and heat stress- regulated expression of HsfA2 and small heat shock proteins in tomato anthers
1
2010
... 花粉感知环境中的高温信号后, 通过信号转导引起代谢、转录甚至表观遗传水平的改变, 从而实现对高温胁迫的响应.其中热激转录因子是高温胁迫应答网络中的关键组分, 可以诱导高温胁迫应答重要效应物热激蛋白的表达, 通过维持蛋白的构型稳定减少由高温导致的蛋白变性和聚集, 从而提高植物耐热性.在烟草(Nicotiana tabacum)花粉发育过程中, 高温诱导不同的HSPs表达(
Genome-wide transcriptome analysis during anthesis reveals new insights into the molecular basis of heat stress responses in tolerant and sensitive rice varieties
1
2016
... 由于花粉取材相对困难, 因此针对花粉高温应答机制的研究相对较少.但随着分离技术的发展以及多种组学技术的应用, 人们对花粉的高温应答机制已有一定认识(
The evolution of the evidence base for observed impacts of climate change
1
2015
... 随着全球气候逐渐变暖, 温度变化对生态环境的影响日益明显(
High temperature injury and auxin biosynthesis in microsporogenesis
1
2013
... 高温胁迫下, 植物体内的激素水平也会发生显著变化(
Rice MADS3 regulates ROS homeostasis during late anther development
1
2011
... ROS是一类化学性质活泼且具有较高氧化活性的分子, 高温胁迫会导致植物体内产生大量ROS.积累的ROS不仅会造成核酸和蛋白质等重要生物大分子的氧化损伤, 也可以作为信号分子激活植物体内的抗氧化系统, 从而调控植物多个器官(包括花粉)响应高温胁迫.对小麦花粉进行高温处理(42°C, 2小时)后, 发现花粉中ROS和超氧化物歧化酶(superoxide dismutase, SOD)、过氧化物酶(peroxidase, POD)和过氧化氢酶(catalase, CAT)含量显著升高, 同时伴随着热激蛋白(heat shock proteins, HSPs) HSP70、HSP90、HSP17和HSP26含量增加(Kumar et al., 2013).番茄(Lycopersicon esculentum)和水稻花粉中也有类似机制(
Physiological and proteomic approaches to address heat tolerance during anthesis in rice (Oryza sativa L.)
1
2010
... 由于花粉取材相对困难, 因此针对花粉高温应答机制的研究相对较少.但随着分离技术的发展以及多种组学技术的应用, 人们对花粉的高温应答机制已有一定认识(
Short- term high temperature growth conditions during vegetative-to-reproductive phase transition irreversibly compromise cell wall invertase-mediated sucrose catalysis and microspore meiosis in grain sorghum (Sorghum bicolor)
1
2010
... 高温还影响小孢子的发育.研究表明, 蔗糖转化酶(invertases, INVs)在高温胁迫下小孢子发育过程中发挥重要作用.高温导致小孢子和花药中INVs基因表达量下降, 从而影响蔗糖水解过程, 造成小孢子中淀粉积累异常, 产生没有活性的花粉(
Ethylene production and signaling in tomato (Solanum lycopersicum) pollen grains is responsive to heat stress conditions
1
2018
... 高温胁迫下, 植物体内的激素水平也会发生显著变化(
Differences inin vitro pollen germination and pollen tube growth of cotton cultivars in response to high temperature
1
2005
... 高温通过改变花粉和柱头细胞中生长素和糖类的平衡影响花粉管在柱头内的生长, 表现出花粉萌发失败以及花粉管末端膨大、扭曲和破裂等现象, 最终导致受精过程无法完成(
Alternative splicing in tomato pollen in response to heat stress
2
2017
... 由于花粉取材相对困难, 因此针对花粉高温应答机制的研究相对较少.但随着分离技术的发展以及多种组学技术的应用, 人们对花粉的高温应答机制已有一定认识(
... 花粉感知环境中的高温信号后, 通过信号转导引起代谢、转录甚至表观遗传水平的改变, 从而实现对高温胁迫的响应.其中热激转录因子是高温胁迫应答网络中的关键组分, 可以诱导高温胁迫应答重要效应物热激蛋白的表达, 通过维持蛋白的构型稳定减少由高温导致的蛋白变性和聚集, 从而提高植物耐热性.在烟草(Nicotiana tabacum)花粉发育过程中, 高温诱导不同的HSPs表达(
The coupling of transcriptome and proteome adaptation during development and heat stress response of tomato pollen
1
2018
... 由于花粉取材相对困难, 因此针对花粉高温应答机制的研究相对较少.但随着分离技术的发展以及多种组学技术的应用, 人们对花粉的高温应答机制已有一定认识(
Quantitative shotgun proteomics analysis of rice anther proteins after exposure to high temperature
1
2015
... 尽管目前对花粉高温胁迫响应机制认识有限, 但是随着各种组学技术、全基因组关联分析(genome- wide association study, GWAS)和CRISPR-Cas9等基因编辑技术在花粉高温胁迫研究中的应用(
Complexity of the heat stress response in plants
2
2007
... 植物进化出完整而复杂的胁迫响应机制来适应高温并维持自身的生长发育(
... 综上, 在应对高温胁迫时, 花粉至少可以通过4种机制感受高温信号并利用多种机制响应胁迫, 这些机制和途径之间也存在“交叉对话”.例如, 内流的钙离子可以参与多个途径; ROS可以影响UPR; ABA则影响糖类代谢; 而糖信号通路又可以影响ROS的平衡.这些机制之间的相互作用也增加了花粉高温胁迫研究的复杂性(
Expression analysis of ClpB/ Hsp100 gene in faba bean (Vicia faba L.) plants in response to heat stress
1
2016
... 花粉感知环境中的高温信号后, 通过信号转导引起代谢、转录甚至表观遗传水平的改变, 从而实现对高温胁迫的响应.其中热激转录因子是高温胁迫应答网络中的关键组分, 可以诱导高温胁迫应答重要效应物热激蛋白的表达, 通过维持蛋白的构型稳定减少由高温导致的蛋白变性和聚集, 从而提高植物耐热性.在烟草(Nicotiana tabacum)花粉发育过程中, 高温诱导不同的HSPs表达(
Ascorbic acid at pre-anthesis modulate the thermotolerance level of wheat (Triticum aestivum) pollen under heat stress
1
2014
... 活性氧清除系统、激素以及糖类物质在花粉的高温应答中也发挥重要作用(
H2A.Z-containing nucleo- somes mediate the thermosensory response in Arabidopsis
2
2010
... 植物进化出完整而复杂的胁迫响应机制来适应高温并维持自身的生长发育(
... H2A.Z是一种组蛋白变体, 包含H2A.Z的核小体与DNA结合更加紧密.研究表明, 高温处理可以减少含有H2A.Z的核小体在基因转录起始位点的占有率, 从而调控相关基因的表达(
Transgenerational inheritance and resetting of stress- induced loss of epigenetic gene silencing in Arabidopsis
1
2010
... 研究作物花粉高温胁迫应答机制并提高作物花粉的高温耐受能力, 对于稳定和提高作物产量具有重要意义.目前, 人们已对高温导致的花粉败育进行了系统的形态学观察和多种组学分析, 并且已鉴定到一些参与花粉高温胁迫响应的因子, 但对于花粉高温胁迫应答调控机制的认知仍然有限(
Core genome responses involved in acclimation to high temperature
1
2008
... 花粉感知环境中的高温信号后, 通过信号转导引起代谢、转录甚至表观遗传水平的改变, 从而实现对高温胁迫的响应.其中热激转录因子是高温胁迫应答网络中的关键组分, 可以诱导高温胁迫应答重要效应物热激蛋白的表达, 通过维持蛋白的构型稳定减少由高温导致的蛋白变性和聚集, 从而提高植物耐热性.在烟草(Nicotiana tabacum)花粉发育过程中, 高温诱导不同的HSPs表达(
The riceTapetum degeneration retardation gene is required for tapetum degradation and anther development
1
2006
... 高温还可以促进绒毡层细胞提前发生程序性死亡(programmed cell death, PCD), 进而影响花粉发育、活性以及散粉过程(
Functional characterization ofArabidopsis thaliana WRKY39 in heat stress
1
2010
... 高温影响花粉发育全过程, 导致花粉结构和发育异常, 产生没有活性的花粉, 并最终导致受精失败(
The type II Arabidopsis formin14 interacts with microtubules and microfilaments to regulate cell division
1
2010
... 质膜上的钙离子通道是植物细胞重要的高温感受器.电化学和生化等实验证实, 温度升高可以触发质膜上钙离子通道打开, 导致胞外钙离子内流, 使胞质内钙离子增加, 引起细胞产生热激反应(heat shock response, HSR) (
Genetic and epigenetic control of plant heat responses
1
2015
... 通常情况下, 在高于最适温度5°C以上的环境下, 花粉就会遭受高温伤害.许多重要作物如小麦(Tri- ticum aestivum)、水稻(Oryza sativa)和玉米(Zea mays), 其开花期均集中在夏季, 极易遭遇连续或严重的高温天气, 从而导致花粉发育异常(
Climate trends and global crop production since 1980
1
2011
... 随着全球气候逐渐变暖, 温度变化对生态环境的影响日益明显(
Effects of high temperatures on mature pollen grains in wild and cultivated maize accessions
1
1991
... 通常情况下, 在高于最适温度5°C以上的环境下, 花粉就会遭受高温伤害.许多重要作物如小麦(Tri- ticum aestivum)、水稻(Oryza sativa)和玉米(Zea mays), 其开花期均集中在夏季, 极易遭遇连续或严重的高温天气, 从而导致花粉发育异常(
The THERMOSENSITIVE MALE STERILE 1 interacts with the BiPs via DnaJ domain and stimulates their atpase enzyme activities in Arabidopsis
1
2015
... 绒毡层或者花粉细胞内的UPR反应也可以感受高温胁迫, 并通过ER-UPR (内质网中未折叠蛋白反应)提高内质网对蛋白质的折叠能力, 降低未折叠蛋白质的数量, 从而提高花粉的胁迫耐受能力.一些转录因子和RNA剪切因子也参与ER-UPR, 并在花粉发育过程中发挥重要作用, 其中erdj2a、bip1/2/3和pod1等突变体均表现出花粉发育异常表型(
Heat stress regimes for the investigation of pollen thermotolerance in crop plants
3
2016
... 在进行花粉高温胁迫研究时, 需要考虑胁迫温度、处理方式以及花粉取材时期.首先需要选择合适的温度处理范围, 不同的温度处理会产生不同的结果.温和高温可以使植物获得高温耐受能力, 严重高温则直接破坏植物细胞, 导致植株不可逆损伤甚至死亡.通过总结常见作物的花粉高温胁迫研究, 发现常见作物的高温胁迫温度通常选择30-39°C.尽管番茄花粉高温胁迫研究也有选择45°C高温的情况, 但处理时间相对较短(
... ).其次, 还需考虑胁迫施加的方式(
... ).对植物直接施加高温(direct applied heat stress, DAHS), 主要用来研究植物的基础高温胁迫耐受性(basal thermo tolerance, BTT).如果在对植物施加高温处理前先预施加一个温和高温(pre-induction heat stress, PIHS), 则主要用于研究植物获得性高温胁迫耐受性(acquired thermotolerance, ATT).例如, 在番茄花粉的ATT研究中, 可以将花粉32°C预处理1小时, 然后置于25°C恢复1小时, 最后再于50°C的高温条件下进行处理.也可以利用温和高温(30- 36°C)对植物处理几周甚至几个月, 这种处理方式被称为温和的慢性高温处理(mild chronic heat stress, MCHS), 可以引起植物产生温和高温胁迫耐受性(mild heat thermotolerance, MHTT).由于MCHS处理方式可以更好地模拟大气环境中温度的变化并覆盖花粉整个发育阶段, 更有利于人们对花粉发育的各个时期的发育状态和生理指标进行监测, 因此已成为花粉高温处理的主要方式(
Transgenerational phenotypic and epigenetic changes in response to heat stress in Arabidopsis thaliana
1
2014
... 研究作物花粉高温胁迫应答机制并提高作物花粉的高温耐受能力, 对于稳定和提高作物产量具有重要意义.目前, 人们已对高温导致的花粉败育进行了系统的形态学观察和多种组学分析, 并且已鉴定到一些参与花粉高温胁迫响应的因子, 但对于花粉高温胁迫应答调控机制的认知仍然有限(
Sugar and auxin signa- ling pathways respond to high-temperature stress during anther development as revealed by transcript profiling analysis in cotton
1
2014
... 活性氧清除系统、激素以及糖类物质在花粉的高温应答中也发挥重要作用(
How do plants feel the heat?
1
2012
... 植物进化出完整而复杂的胁迫响应机制来适应高温并维持自身的生长发育(
Acclimation to high temperature during pollen development
2
2016
... 高温还可以促进绒毡层细胞提前发生程序性死亡(programmed cell death, PCD), 进而影响花粉发育、活性以及散粉过程(
... 活性氧清除系统、激素以及糖类物质在花粉的高温应答中也发挥重要作用(
Abiotic stress enhances androgenesis from isolated microspores of some legume species (Fabaceae)
1
2009
... 高温还影响小孢子的发育.研究表明, 蔗糖转化酶(invertases, INVs)在高温胁迫下小孢子发育过程中发挥重要作用.高温导致小孢子和花药中INVs基因表达量下降, 从而影响蔗糖水解过程, 造成小孢子中淀粉积累异常, 产生没有活性的花粉(
ABA regulates apoplastic sugar transport and is a potential signal for cold-induced pollen sterility in rice
1
2007
... 高温胁迫下, 植物体内的激素水平也会发生显著变化(
The influence of terminal heat stress on meiosis abnormalities in pollen mother cells of wheat
1
2014
... 高温影响花粉发育全过程, 导致花粉结构和发育异常, 产生没有活性的花粉, 并最终导致受精失败(
Premature progression of anther early developmental programs accompanied by comprehensive alterations in transcription du- ring high-temperature injury in barley plants
1
2007
... 高温影响花粉发育全过程, 导致花粉结构和发育异常, 产生没有活性的花粉, 并最终导致受精失败(
Tapetal development and abiotic stress: a centre of vulnerability
1
2012
... 高温还可以促进绒毡层细胞提前发生程序性死亡(programmed cell death, PCD), 进而影响花粉发育、活性以及散粉过程(
Heat stress affects the cytoskeleton and the delivery of sucrose synthase in tobacco pollen tubes
1
2016
... 高温通过改变花粉和柱头细胞中生长素和糖类的平衡影响花粉管在柱头内的生长, 表现出花粉萌发失败以及花粉管末端膨大、扭曲和破裂等现象, 最终导致受精过程无法完成(
The metabolic basis of pollen thermo-tolerance: perspectives for breeding
1
2014
... 尽管目前对花粉高温胁迫响应机制认识有限, 但是随着各种组学技术、全基因组关联分析(genome- wide association study, GWAS)和CRISPR-Cas9等基因编辑技术在花粉高温胁迫研究中的应用(
Stress-induced chromatin changes: a critical view on their heritability
1
2012
... 研究作物花粉高温胁迫应答机制并提高作物花粉的高温耐受能力, 对于稳定和提高作物产量具有重要意义.目前, 人们已对高温导致的花粉败育进行了系统的形态学观察和多种组学分析, 并且已鉴定到一些参与花粉高温胁迫响应的因子, 但对于花粉高温胁迫应答调控机制的认知仍然有限(
Polyploidization mechanisms: temperature environment can induce diploid gamete formation inRosa sp
3
2011
... 高温影响花粉发育全过程, 导致花粉结构和发育异常, 产生没有活性的花粉, 并最终导致受精失败(
... ;
... ).这种由于高温引起小孢子母细胞减数分裂异常, 进而导致花粉多倍体化(polyploidization)的现象对植物进化具有重要意义(
Effects of high-temperature stress on microsporogenesis in heat-sensitive and heat-tolerant genotypes of Phaseolus vulgaris
1
2001
... 高温影响花粉发育全过程, 导致花粉结构和发育异常, 产生没有活性的花粉, 并最终导致受精失败(
Species, ecotype and cultivar differences in spikelet fertility and harvest index of rice in response to high temperature stress
1
2006
... 通常情况下, 在高于最适温度5°C以上的环境下, 花粉就会遭受高温伤害.许多重要作物如小麦(Tri- ticum aestivum)、水稻(Oryza sativa)和玉米(Zea mays), 其开花期均集中在夏季, 极易遭遇连续或严重的高温天气, 从而导致花粉发育异常(
The effect of heat stress on tomato pollen characteristics is associated with changes in carbohydrate concentration in the developing anthers
1
2002
... 花粉作为有性生殖过程中光合产物重要的“库”, 其淀粉含量通常在二细胞花粉期达到最高值, 而可溶性糖的含量在散粉期达到最高值.积累的糖类物质不仅为花粉发育和花粉管形成提供能量, 也可以作为渗透物质提高花粉对胁迫的响应.通过比较不同的番茄品种可以发现, 耐高温品种的花粉中含有更多的淀粉和可溶性糖(
Melatonin alleviates high temperature-induced pollen abortion in Solanum lycopersicum
1
2018
... 花粉作为有性生殖过程中光合产物重要的“库”, 其淀粉含量通常在二细胞花粉期达到最高值, 而可溶性糖的含量在散粉期达到最高值.积累的糖类物质不仅为花粉发育和花粉管形成提供能量, 也可以作为渗透物质提高花粉对胁迫的响应.通过比较不同的番茄品种可以发现, 耐高温品种的花粉中含有更多的淀粉和可溶性糖(
Heat stress-responsive transcriptome analysis in heat susceptible and tolerant wheat (Triticum aestivum L.) by using Wheat Genome Array
1
2008
... 由于花粉取材相对困难, 因此针对花粉高温应答机制的研究相对较少.但随着分离技术的发展以及多种组学技术的应用, 人们对花粉的高温应答机制已有一定认识(
A comparison of heat-stress transcriptome changes between wild-type Arabidopsis pollen and a heat-sensitive mutant harboring a knockout of cyclic nucleotide-gated cation channel 16 (cn- gc16)
1
2018
... 质膜上的钙离子通道是植物细胞重要的高温感受器.电化学和生化等实验证实, 温度升高可以触发质膜上钙离子通道打开, 导致胞外钙离子内流, 使胞质内钙离子增加, 引起细胞产生热激反应(heat shock response, HSR) (
A new link between stress response and nucleolar function during pollen development in Arabidopsis mediated by AtREN1 protein
1
2014
... 花粉感知环境中的高温信号后, 通过信号转导引起代谢、转录甚至表观遗传水平的改变, 从而实现对高温胁迫的响应.其中热激转录因子是高温胁迫应答网络中的关键组分, 可以诱导高温胁迫应答重要效应物热激蛋白的表达, 通过维持蛋白的构型稳定减少由高温导致的蛋白变性和聚集, 从而提高植物耐热性.在烟草(Nicotiana tabacum)花粉发育过程中, 高温诱导不同的HSPs表达(
Abscisic acid prevents pollen abortion under high-temperature stress by mediating sugar metabolism in rice spikelets
1
2018
... 高温胁迫下, 植物体内的激素水平也会发生显著变化(
The heat shock response in moss plants is regulated by specific calcium-permeable channels in the plasma membrane
1
2009
... 质膜上的钙离子通道是植物细胞重要的高温感受器.电化学和生化等实验证实, 温度升高可以触发质膜上钙离子通道打开, 导致胞外钙离子内流, 使胞质内钙离子增加, 引起细胞产生热激反应(heat shock response, HSR) (
Reduction of gibberellin by low temperature disrupts pollen development in rice
1
2014
... 高温胁迫下, 植物体内的激素水平也会发生显著变化(
Auxins reverse plant male sterility caused by high temperatures
1
2010
... 高温胁迫下, 植物体内的激素水平也会发生显著变化(
Expression of genes for the biosynthesis of compatible solutes during pollen development under heat stress in tomato (Solanum lycopersicum)
1
2015
... 花粉作为有性生殖过程中光合产物重要的“库”, 其淀粉含量通常在二细胞花粉期达到最高值, 而可溶性糖的含量在散粉期达到最高值.积累的糖类物质不仅为花粉发育和花粉管形成提供能量, 也可以作为渗透物质提高花粉对胁迫的响应.通过比较不同的番茄品种可以发现, 耐高温品种的花粉中含有更多的淀粉和可溶性糖(
Moderate increase of mean daily temperature adversely affects fruit set of lycopersicon esculentum by disrupting specific physiological processes in male reproductive development
1
2006
... 高温影响花粉发育全过程, 导致花粉结构和发育异常, 产生没有活性的花粉, 并最终导致受精失败(
Determining critical pre- and post-anthesis periods and physiological pro- cesses in Lycopersicon esculentum Mill. exposed to mode- rately elevated temperatures
1
2002
... 高温还可以促进绒毡层细胞提前发生程序性死亡(programmed cell death, PCD), 进而影响花粉发育、活性以及散粉过程(
Pollen germination andin vivo fertilization in response to high-temperature during flowering in hybrid and inbred rice
1
2018
... 高温通过改变花粉和柱头细胞中生长素和糖类的平衡影响花粉管在柱头内的生长, 表现出花粉萌发失败以及花粉管末端膨大、扭曲和破裂等现象, 最终导致受精过程无法完成(
How does timing, duration and severity of heat stress influence pollen-pistil interactions in angiosperms?
1
2011
... 高温通过改变花粉和柱头细胞中生长素和糖类的平衡影响花粉管在柱头内的生长, 表现出花粉萌发失败以及花粉管末端膨大、扭曲和破裂等现象, 最终导致受精过程无法完成(
High temperature limits in vivo pollen tube growth rates by altering diurnal carbohydrate balance in field- grown Gossypium hirsutum pistils
1
2011
... 高温通过改变花粉和柱头细胞中生长素和糖类的平衡影响花粉管在柱头内的生长, 表现出花粉萌发失败以及花粉管末端膨大、扭曲和破裂等现象, 最终导致受精过程无法完成(
DNA methylation dynamics andMET1a-like gene expression changes during stress-induced pollen reprogramming to embryogenesis
1
2012
... 研究作物花粉高温胁迫应答机制并提高作物花粉的高温耐受能力, 对于稳定和提高作物产量具有重要意义.目前, 人们已对高温导致的花粉败育进行了系统的形态学观察和多种组学分析, 并且已鉴定到一些参与花粉高温胁迫响应的因子, 但对于花粉高温胁迫应答调控机制的认知仍然有限(
Anther response to high- temperature stress during development and pollen thermotolerance heterosis as revealed by pollen tube growth andin vitro pollen vigor analysis in upland cotton
1
2015
... 高温通过改变花粉和柱头细胞中生长素和糖类的平衡影响花粉管在柱头内的生长, 表现出花粉萌发失败以及花粉管末端膨大、扭曲和破裂等现象, 最终导致受精过程无法完成(
Ultrastructural study on degeneration of tapetum in anther of snap bean (Phaseolus vulgaris L.) under heat stress
1
2001
... 高温还可以促进绒毡层细胞提前发生程序性死亡(programmed cell death, PCD), 进而影响花粉发育、活性以及散粉过程(
Possible correlation between high temperature-induced floret sterility and endogenous levels of IAA, GAs and ABA in rice (Oryza sativa L.)
1
2008
... 高温胁迫下, 植物体内的激素水平也会发生显著变化(
A cyclic nucleotide-gated channel (CNGC16) in pollen is critical for stress tolerance in pollen reproductive development
1
2013
... 质膜上的钙离子通道是植物细胞重要的高温感受器.电化学和生化等实验证实, 温度升高可以触发质膜上钙离子通道打开, 导致胞外钙离子内流, 使胞质内钙离子增加, 引起细胞产生热激反应(heat shock response, HSR) (
Male gametogenesis and germline specification in flowering plants
1
2011
... 花粉作为植物高度特化的雄配子体, 其发育过程既短暂又复杂(
Plant hormone- mediated regulation of stress responses
1
2016
... 高温胁迫下, 植物体内的激素水平也会发生显著变化(
Small heat shock proteins are differentially regulated during pollen deve- lopment and following heat stress in tobacco
1
2005
... 花粉感知环境中的高温信号后, 通过信号转导引起代谢、转录甚至表观遗传水平的改变, 从而实现对高温胁迫的响应.其中热激转录因子是高温胁迫应答网络中的关键组分, 可以诱导高温胁迫应答重要效应物热激蛋白的表达, 通过维持蛋白的构型稳定减少由高温导致的蛋白变性和聚集, 从而提高植物耐热性.在烟草(Nicotiana tabacum)花粉发育过程中, 高温诱导不同的HSPs表达(
High temperature-induced production of unreduced pollen and its cytological effects in Populus
1
2017
... 高温影响花粉发育全过程, 导致花粉结构和发育异常, 产生没有活性的花粉, 并最终导致受精失败(
Plant ion chan-nels: gene families, physiology, and functional genomics analyses
1
2009
... 质膜上的钙离子通道是植物细胞重要的高温感受器.电化学和生化等实验证实, 温度升高可以触发质膜上钙离子通道打开, 导致胞外钙离子内流, 使胞质内钙离子增加, 引起细胞产生热激反应(heat shock response, HSR) (
A CRISPR/Cas9 toolkit for multiplex genome editing in plants
1
2014
... 尽管目前对花粉高温胁迫响应机制认识有限, 但是随着各种组学技术、全基因组关联分析(genome- wide association study, GWAS)和CRISPR-Cas9等基因编辑技术在花粉高温胁迫研究中的应用(
Mapping quantitative trait loci for heat tolerance of reproductive traits in tomato (Solanum lyco- persicum)
1
2017
... 尽管目前对花粉高温胁迫响应机制认识有限, 但是随着各种组学技术、全基因组关联分析(genome- wide association study, GWAS)和CRISPR-Cas9等基因编辑技术在花粉高温胁迫研究中的应用(
RNA-seq reveals differentially expressed genes of rice ( Oryza sativa) spikelet in response to temperature interacting with nitrogen at meiosis stage
1
2015
... 由于花粉取材相对困难, 因此针对花粉高温应答机制的研究相对较少.但随着分离技术的发展以及多种组学技术的应用, 人们对花粉的高温应答机制已有一定认识(
A mutation in THERMOSENSITIVE MALE STERILE 1, encoding a heat shock protein with DnaJ and PDI domains, leads to thermosensitive gametophytic male sterility in Arabidopsis
1
2009
... 绒毡层或者花粉细胞内的UPR反应也可以感受高温胁迫, 并通过ER-UPR (内质网中未折叠蛋白反应)提高内质网对蛋白质的折叠能力, 降低未折叠蛋白质的数量, 从而提高花粉的胁迫耐受能力.一些转录因子和RNA剪切因子也参与ER-UPR, 并在花粉发育过程中发挥重要作用, 其中erdj2a、bip1/2/3和pod1等突变体均表现出花粉发育异常表型(
Heat stress induces spikelet sterility in rice at anthesis through inhibition of pollen tube elongation interfering with auxin homeostasis in pollinated pistils
1
2018
... 高温通过改变花粉和柱头细胞中生长素和糖类的平衡影响花粉管在柱头内的生长, 表现出花粉萌发失败以及花粉管末端膨大、扭曲和破裂等现象, 最终导致受精过程无法完成(
Cytological analysis and genetic control of rice anther development
1
2011
... 在设计花粉高温胁迫相关实验时, 还需要考虑花粉所处的发育阶段.不同发育阶段的花粉对高温胁迫的敏感性和响应机制可能不同.在实际研究中, 通常把花粉发育过程分为4个阶段: (1) 花粉母细胞到四分体时期; (2) 小孢子形成到早期的二细胞时期; (3) 二细胞末期/三细胞期到成熟花粉期; (4) 花粉萌发期.目前, 可以通过一些形态指标对常见作物花粉发育阶段进行判断.例如, 水稻花粉的发育阶段可以通过花芽长度进行判断; 而玉米花粉的发育阶段则可以通过茎秆的叶领(leaf collar)数进行判断(
Molecular and genetic evidence for the key role of AtCaM3 in heat-shock signal transduction in Arabidopsis
1
2009
... 质膜上的钙离子通道是植物细胞重要的高温感受器.电化学和生化等实验证实, 温度升高可以触发质膜上钙离子通道打开, 导致胞外钙离子内流, 使胞质内钙离子增加, 引起细胞产生热激反应(heat shock response, HSR) (
Involvement of CAT in the detoxification of HT-induced ROS burst in rice anther and its relation to pollen fertility
2
2018
... ROS是一类化学性质活泼且具有较高氧化活性的分子, 高温胁迫会导致植物体内产生大量ROS.积累的ROS不仅会造成核酸和蛋白质等重要生物大分子的氧化损伤, 也可以作为信号分子激活植物体内的抗氧化系统, 从而调控植物多个器官(包括花粉)响应高温胁迫.对小麦花粉进行高温处理(42°C, 2小时)后, 发现花粉中ROS和超氧化物歧化酶(superoxide dismutase, SOD)、过氧化物酶(peroxidase, POD)和过氧化氢酶(catalase, CAT)含量显著升高, 同时伴随着热激蛋白(heat shock proteins, HSPs) HSP70、HSP90、HSP17和HSP26含量增加(Kumar et al., 2013).番茄(Lycopersicon esculentum)和水稻花粉中也有类似机制(
... 活性氧清除系统、激素以及糖类物质在花粉的高温应答中也发挥重要作用(
Relationship of ROS accumulation and superoxide dismutase isozymes in develop- ping anther with floret fertility of rice under heat stress
1
2018
... 活性氧清除系统、激素以及糖类物质在花粉的高温应答中也发挥重要作用(
Temperature stress and plant sexual reproduction: uncovering the wea- kest links
1
2010
... 在进行花粉高温胁迫研究时, 需要考虑胁迫温度、处理方式以及花粉取材时期.首先需要选择合适的温度处理范围, 不同的温度处理会产生不同的结果.温和高温可以使植物获得高温耐受能力, 严重高温则直接破坏植物细胞, 导致植株不可逆损伤甚至死亡.通过总结常见作物的花粉高温胁迫研究, 发现常见作物的高温胁迫温度通常选择30-39°C.尽管番茄花粉高温胁迫研究也有选择45°C高温的情况, 但处理时间相对较短(
备案号: 京ICP备16067583号-21
版权所有 © 2021 《植物学报》编辑部
地址:北京香山南辛村20号 邮编:100093
电话:010-62836135 010-62836131 E-mail:cbb@ibcas.ac.cn
本系统由北京玛格泰克科技发展有限公司设计开发

{kind=link}
{kind=link}
{kind=link}
{kind=link}