Abstract In order to identify long-distance signals under iron (Fe) deficiency in rice (Oryza sativa), TMT label technique was used to study the protein profile of phloem sap under different iron concentrations. A total of 206 differentially ex- pressed proteins were identified: 54 were upregulated and 152 were downregulated. Most of these proteins are involved in hormone signal transduction, carbon metabolism, glutathione metabolism and mRNA transport. In addition, we mea- sured the physiological indicators according to the differentially expressed proteins. It was found that phytohormones, sucrose, glutathione and transporters were significantly changed under iron deficiency. To further study the function of these proteins can help to reveal the long distance signaling pathway of rice in response to Fe deficiency. Keywords:rice (Oryza sativa);iron deficiency;phloem;long-distance signal;TMT
PDF (1490KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 陈琳, 林焱, 陈鹏飞, 王绍华, 丁艳锋. 水稻响应缺铁的韧皮部汁液蛋白质组学分析. 植物学报, 2019, 54(2): 194-207 doi:10.11983/CBB18184 Chen Lin, Lin Yan, Chen Pengfei, Wang Shaohua, Ding Yanfeng. Effect of Iron Deficiency on the Protein Profile of Rice (Oryza sativa) Phloem Sap. Chinese Bulletin of Botany, 2019, 54(2): 194-207 doi:10.11983/CBB18184
铁是植物生长必需的矿质营养。为适应低铁环境, 植物进化出2种不同的吸收机制: 还原途径(机理I)和螯合途径(机理II)。通过机理I吸收铁的植物(以下简称机理I植物)包括双子叶植物和非禾本科单子叶植物; 通过机理II吸收铁的植物(以下简称机理II植物)主要是禾本科植物。东京大学Nishizawa的研究发现, 机理II植物水稻(Oryza sativa)也能通过机理I的方式吸收铁(Ishimaru et al., 2006), 即水稻兼具2条途径进行铁的吸收, 该结论后经吴平教授实验室用突变体证实(Cheng et al., 2007)。
在感受到缺铁信号时, 机理I植物根部表皮会诱导产生3种反应: (1) H+-ATPase大量表达, 酸化根际土壤环境, 从而提高根际铁的有效性; (2) 铁还原酶FRO (ferric reductase oxidase)基因表达上调, 催化根表皮的三价铁(Fe(III))还原成二价铁(Fe(II)); (3) Fe(II)的转运体IRT (iron-regulated transporter)表达上调, 将被FRO还原成的Fe(II)转运入根部细胞(吴慧兰等, 2007)。该过程中的关键基因FRO (Robinson et al., 1999)和IRT (Eide et al., 1996)最早于20世纪90年代在拟南芥(Arabidopsis thaliana)中克隆得到, 之后相继报道了它们在多种植物中的同源基因(Con- nolly et al., 2003; Ishimaru et al., 2007)。
水稻的另一种铁吸收方式为螯合方式, 该吸收途径通过合成一种高亲和的与三价铁结合的物质——植物铁载体PS (phytosiderophore), 分泌到根际土壤环境中, 之后与Fe(III)结合生成Fe(III)-PS复合体(Ne- gishi et al., 2002)。同时, 植物表皮细胞质膜上的一种多肽转运体OPT (oligo-peptide transporter)家族成员YS (Yellow Stripe)转运体(Curie et al., 2001)可将上述根表皮细胞附近的Fe(III)-PS复合体跨膜转运到植物体内。OsYSL15是水稻吸收铁过程中Fe(III)-PS的主要转运体(Inoue et al., 2009; Lee et al., 2009), 其受缺铁诱导上调表达。
红色表示蛋白丰度上调, 黄色表示蛋白丰度下调。实线表示直接作用, 虚线表示中间有省略过程。箭头表示促进作用(正效应), 短竖线表示抑制作用(负效应)。 Figure 2Schematic representation of plant hormone signal transduction in phloem of rice under iron deficiency
Red represents an increase of protein expression abundance, and yellow represents a decrease. The solid lines indicate direct action and the dotted lines indicate omission process. The arrows mean a positive effect and the short vertical lines mean inhibited effect.
蛋白激酶和蛋白磷酸酶(PPs)介导的可逆磷酸化是普遍存在于生物体内的信号转导机制。蛋白磷酸酶包括PP1、PP2A、PP2B和PP2C, PP2Cs是植物中最大的蛋白磷酸酶家族(Kerk et al., 2002)。在高等植物中, PP2C参与多种信号转导途径, 包括ABA、环境胁迫以及生长发育等(Tougane et al., 2010)。ABA信号转导途径主要有PP2Cs和SnRK2两种家族蛋白参与。ABA含量较高时, 其受体能够与PYR/PYL家族蛋白结合, 进而结合PP2Cs蛋白的催化位点, 抑制其活性; 相应地, SnRK2家族蛋白的活性得以激活, 从而激活ABA信号途径(Soon et al., 2012)。本研究中, 缺铁条件下水稻韧皮部中参与脱落酸信号转导的蛋白磷酸酶(protein phosphatase 2C 30)表达丰度下调(图2), 表明韧皮部中ABA的含量可能增多, 且转运增强。ABA在植物面临高温和干旱等逆境时会被显著诱导, 是植物遭受逆境胁迫的重要标志之一。同样, 在植物面临缺铁胁迫时, 其内源ABA含量也可能激增。
黄色表示蛋白丰度下调。实线表示直接作用, 虚线表示中间有省略过程。箭头表示促进作用(正效应)。 Figure 4Metabolic pathways of carbon (A) and glutathione (B) related proteins in phloem exudates of rice under iron deficiency
Yellow represents a decrease of protein expression abundance. The solid lines indicate direct action and the dotted lines indicate omission process. The arrows mean a positive effect.
柱形图代表缺铁与对照基因相对表达量的比值。 Figure 5The relative expression levels of OsIRT1, OsYSL15, OsNAS1 and OsNAS2 in root (A) and the relative expression levels of OsYSL2 and OsNAS1 in shoot (B) after three days of iron deficiency treatment
The column diagram represents the relative gene expression level ratio of iron deficiency induced to the control.
2.3 讨论
长距离调节一直是植物营养学的研究热点, 但是其长距离反馈调控机制并不清楚。嫁接和分根实验是研究长距离信号的常用手段。已有研究显示, 将拟南芥根分开, 一部分放入NO3-饥饿处理, 另一部分正常供氮, 发现正常供氮的根系中AtNRT2.1转录水平上调(Mol et al., 1994), 表明植物营养的长距离运输是客观存在的。但信号物质如何进行调控, 以及与地上部是否相关目前还不清楚。前期我们通过分根和去除地上部实验验证了水稻缺铁后, 长距离运输信号客观存在, 并且该信号强度与地上部功能叶相关。缺铁条件下, 水稻地上部会产生信号物质并通过韧皮部传输到根系, 进一步调控与铁吸收相关的标志基因, 从而调节铁的代谢平衡(Chen et al., 2018a)。
ThimmO, EssigmannB, KloskaS, AltmannT, BuckhoutTJ ( 2001). Response of Arabidopsis to iron deficiency stress as revealed by microarray analysis Plant Physiol 127, 1030-1043. DOI:10.1104/pp.010191URL [本文引用: 1]
TiffinLO ( 1966). Iron translocation II. Citrate/iron ratios in plant stem exudates Plant Physiol 41, 515-518. DOI:10.1104/pp.41.3.515URL [本文引用: 1]
TouganeK, KomatsuK, BhyanSB, SakataY, IshizakiK, YamatoKT, KohchiT, TakezawaD ( 2010). Evolutionarily conserved regulatory mechanisms of abscisic acid signa- ling in land plants: characterization of ABSCISIC ACID INSENSITIVE1-like type 2C protein phosphatase in the liverwort Marchantia polymorpha Plant Physiol 152, 1529-1543. [本文引用: 1]
VasiliouV, VasiliouK, NebertDW ( 2009). Human ATP- binding cassette (ABC) transporter family Hum Genomics 3, 281-290. [本文引用: 2]
VertGA, BriatJF, CurieC ( 2003). Dual regulation of the Arabidopsis high-affinity root iron uptake system by local and long-distance signals Plant Physiol 132, 796-804. DOI:10.1104/pp.102.016089URL [本文引用: 2]
von WirénN, KlairS, BansalS, BriatJF, KhodrH, ShioiriT, LeighRA, HiderRC ( 1999). Nicotianamine chelates both Fe III and Fe II. Implications for metal transport in plants Plant Physiol 119, 1107-1114. DOI:10.1104/pp.119.3.1107URL [本文引用: 1]
WangBL, WeiHF, XueZ, ZhangWH ( 2017). Gibberellins regulate iron deficiency-response by influencing iron trans- port and translocation in rice seedlings (Oryza sativa) Ann Bot 119, 945-956. [本文引用: 1]
YokoshoK, YamajiN, UenoD, MitaniN, MaJF ( 2009). OsFRDL1 is a citrate transporter required for efficient translocation of iron in rice Plant Physiol 149, 297-305. DOI:10.1104/pp.108.128132URL [本文引用: 1]
AgarwalaSC, SharmaCP ( 1961). The relation of iron supply to the tissue concentration of iron, chlorophyll and catalase in barley plants grown in sand culture Physiol Plant 14, 275-283. DOI:10.1111/ppl.1961.14.issue-2URL [本文引用: 1]
AoyamaT, KobayashiT, TakahashiM, NagasakaS, UsudaK, KakeiY, IshimaruY, NakanishiH, MoriS, NishizawaNK ( 2009). OsYSL18 is a rice iron(III)-deoxy- mugineic acid transporter specifically expressed in reproductive organs and phloem of lamina joints Plant Mol Biol 70, 681-692. DOI:10.1007/s11103-009-9500-3URL [本文引用: 1]
BaoF, LiJY ( 2002). Evidence that the auxin signaling path- way interacts with plant stress response Acta Bot Sin 44, 532-536. [本文引用: 1]
BashirK, NagasakaS, ItaiRN, KobayashiT, TakahashiM, NakanishiH, MoriS, NishizawaNK ( 2007). Expres- sion and enzyme activity of glutathione reductase is upregulated by Fe-deficiency in graminaceous plants Plant Mol Biol 65, 277-284. DOI:10.1007/s11103-007-9216-1URL [本文引用: 1]
BrownJC, ChaneyRL ( 1971). Effect of iron on the transport of citrate into the xylem of soybeans and tomatoes Plant Physiol 47, 836-840. DOI:10.1104/pp.47.6.836URL [本文引用: 1]
ChenL, DingCQ, ZhaoXF, XuJX, MohammadAA, WangSH, DingYF ( 2015). Differential regulation of proteins in rice ( Oryza sativa L.) under iron deficiency Plant Cell Rep 34, 83-96. [本文引用: 1]
ChenL, WangGP, ChenPF, ZhuHL, WangSH, DingYF ( 2018a). Shoot-root communication plays a key role in physiological alterations of rice (Oryza sativa) under iron deficiency Front Plant Sci 9, 757. [本文引用: 3]
ChenPF, ChenL, JiangZR, WangGP, WangSH, DingYF ( 2018b). Sucrose is involved in the regulation of iron deficiency responses in rice ( Oryza sativa L.) . Plant Cell Rep 37, 789-798. [本文引用: 1]
ChengLJ, WangF, ShouHX, HuangFL, ZhengLQ, HeF, LiJH, ZhaoFJ, UenoD, MaJF, WuP ( 2007). Mutation in nicotianamine aminotransferase stimulated the Fe(II) acquisition system and led to iron accumulation in rice Plant Physiol 145, 1647-1657. DOI:10.1104/pp.107.107912URL [本文引用: 1]
ConnollyEL, CampbellN, GrotzN, PrichardCL, GuerinotML ( 2003). Overexpression of the FRO2 iron reductase confers tolerance to growth on low iron and uncovers post-transcriptional control Plant Physiol 133, 1102-1110. DOI:10.1104/pp.103.025122URL [本文引用: 1]
CurieC, CassinG, CounchD, DivolF, HiguchiK, JeanML, MissonJ, SchikoraA, CzernicP, MariS ( 2009). Metal movement within the plant: contribution of nico- tianamine and yellow stripe 1-like transporters Ann Bot 103, 1-11. DOI:10.1093/aob/mcn207URL [本文引用: 1]
CurieC, PanavieneZ, LoulergueC, DellaportaSL, BriatJF, WalkerEL ( 2001). Maize yellow stripe1 encodes a membrane protein directly involved in Fe(III) uptake Nature 409, 346-349. [本文引用: 1]
DeanM, HamonY, ChiminiG ( 2001). The human ATP- binding cassette (ABC) transporter superfamily J Lipid Res 42, 1007-1017. [本文引用: 1]
DongJ, PiñerosMA, LiX, YangH, LiuY, MurphyAS, KochianLV, LiuD ( 2017). An Arabidopsis ABC trans- porter mediates phosphate deficiency-induced remodeling of root architecture by modulating iron homeostasis in roots Mol Plant 10, 244-259. DOI:10.1016/j.molp.2016.11.001URL [本文引用: 1]
EideD, BroderiusM, FettJ, GuerinotML ( 1996). A novel iron-regulated metal transporter from plants identified by functional expression in yeast Proc Natl Acad Sci USA 93, 5624-5628. DOI:10.1073/pnas.93.11.5624URL [本文引用: 1]
EnomotoY, HodoshimaH, ShimadaH, ShojiK, Yoshi- haraT, GotoF ( 2007). Long-distance signals positively regulate the expression of iron uptake genes in tobacco roots Planta 227, 81-89. DOI:10.1007/s00425-007-0596-xURL [本文引用: 2]
EspenL, Dell'OrtoM, NisiPD, ZocchiG ( 2000). Metabolic responses in cucumber ( Cucumis sativus L.) roots under Fe-deficiency: a 31P-nuclear magnetic resonance in-vivo study Planta 210, 985-992. [本文引用: 1]
FalkaJ, AndersenaG, KernebeckbB, KrupinskaaK ( 2003). Constitutive overexpression of barley 4-hydroxyp henylpyruvate dioxygenase in tobacco results in elevation of the vitamin E content in seeds but not in leaves FEBS Lett 540, 35-40. DOI:10.1016/S0014-5793(03)00166-2URL [本文引用: 1]
FordeBG ( 2002). The role of long-distance signaling in plant responses to nitrate and other nutrients J Exp Bot 53, 39-43. [本文引用: 1]
GarcíaMJ, RomeraFJ, StaceyMG, StaceyG, VillarE, AlcántaraE, Pérez-VicenteR ( 2013). Shoot to root com- munication is necessary to control the expression of iron- acquisition genes in Strategy I plants Planta 237, 65-75. DOI:10.1007/s00425-012-1757-0URL [本文引用: 1]
GreenLS, RogersEE ( 2004). FRD3 controls iron localiza- tion in Arabidopsis Plant Physiol 136, 2523-2531. [本文引用: 1]
HagenG, UhrhammerN, GuilfoyleTJ ( 1988). Regulation of expression of an auxin-induced soybean sequence by cadmium J Biol Chem 263, 6442-6446. [本文引用: 1]
HellR, StephanUW ( 2003). Iron uptake, trafficking and homeostasis in plants Planta 216, 541-551. [本文引用: 1]
HindtMN, GuerinotML ( 2012). Getting a sense for signals: regulation of the plant iron deficiency response Biochim Biophys Acta 1823, 1521-1530. DOI:10.1016/j.bbamcr.2012.03.010URL [本文引用: 1]
InoueH, KobayashiT, NozoyeT, TakahashiM, KakeiY, SuzukiK, NakazonoM, NakanishiH, MoriS, NishizawaNK ( 2009). Rice OsYSL15 is an iron-regulated iron(III)- deoxymugineic acid transporter expressed in the roots and is essential for iron uptake in early growth of the seedlings J Biol Chem 284, 3470-3479. DOI:10.1074/jbc.M806042200URL [本文引用: 2]
IshimaruY, KimSA, TsukamotoT, OkiH, KobayashiT, WatanabeS, MatsuhashiS, TakahashiM, NakanishiH, MoriS, NishizawaNK ( 2007). Mutational reconstructed ferric chelate reductase confers enhanced tolerance in rice to iron deficiency in calcareous soil Proc Natl Acad Sci USA 104, 7373-7378. DOI:10.1073/pnas.0610555104URL [本文引用: 1]
IshimaruY, SuzukiM, TsukamotoT, SuzukiK, NakazonoM, KobayashiT, WadaY, WatanabeS, MatsuhashiS, TakahashiM, NakanishiH, MoriS, NishizawaNK ( 2006). Rice plants take up iron as an Fe 3+-phytosidero- phore and as Fe 2+ Plant J 45, 335-346. DOI:10.1111/tpj.2006.45.issue-3URL [本文引用: 2]
JiangCF, GaoXH, LiaoLL, HarberdNP, FuXD ( 2007). Phosphate starvation root architecture and anthocyanin accumulation responses are modulated by the gibberellin- DELLA signaling pathway in Arabidopsis Plant Physiol 145, 1460-1470. DOI:10.1104/pp.107.103788URL [本文引用: 1]
KakeiY, YamaguchiI, KobayashiT, TakahashiM, Naka- nishiH, YamakawaT, NishizawaNK ( 2009). A highly sensitive, quick and simple quantification method for nicotianamine and 2’-deoxymugineic acid from minimum samp- les using LC/ESI-TOF-MS achieves functional analysis of these components in plants Plant Cell Physiol 50, 1988-1993. DOI:10.1093/pcp/pcp141URL [本文引用: 1]
KerkD, BulgrienJ, SmithDW, BarsamB, VeretnikS, GribskovM ( 2002). The complement of protein phos- phatase catalytic subunits encoded in the genome of Arabidopsis Plant Physiol 129, 908-925. DOI:10.1104/pp.004002URL [本文引用: 1]
LeeS, ChieckoJC, KimSA, WalkerEL, LeeY, GuerinotML, AnG ( 2009). Disruption of OsYSL15 leads to iron inefficiency in rice plants Plant Physiol 150, 786-800. DOI:10.1104/pp.109.135418URL [本文引用: 1]
LeiGJ, ZhuXF, WangZW, DongF, DongNY, ZhengSJ ( 2014). Abscisic acid alleviates iron deficiency by promo- ting root iron reutilization and transport from root to shoot in Arabidopsis Plant Cell Environ 37, 852-863. DOI:10.1111/pce.12203URL [本文引用: 1]
LiJ, WuXD, HaoST, WangXJ, LingHQ ( 2008). Proteomic response to iron deficiency in tomato root Proteomics 8, 2299-2311. DOI:10.1002/(ISSN)1615-9861URL [本文引用: 1]
LiuKD, YueRQ, YuanCC, LiuJX, ZhangL, SunT, YangYJ, TieSG, ShenCJ ( 2015). Auxin signaling is involved in iron deficiency-induced photosynthetic inhibition and shoot growth defect in rice ( Oryza sativa L.) J Plant Biol 58, 391-401. [本文引用: 1]
López-MillánAF, MoralesF, AndaluzS, GogorcenaY, AbadíaA, De Las RivasJ, AbadíaJ ( 2000). Responses of sugar beet roots to iron deficiency. Changes in carbon assimilation and oxygen use Plant Physiol 124, 885-897. DOI:10.1104/pp.124.2.885URL [本文引用: 1]
LucasWJ, YooBC, KraglerF ( 2001). RNA as a long- distance information macromolecule in plants Nat Rev Mol Cell Biol 2, 849-857. [本文引用: 1]
MajerusV, BertinP, LuttsS ( 2009). Abscisic acid and oxidative stress implications in overall ferritin synthesis by African rice ( Oryza glaberrima Steud.) seedlings exposed to short term iron toxicity Plant Soil 324, 253-265. [本文引用: 1]
MatsuokaK, FurukawaJ, BidadiH, AsahinaM, Yama- guchiS, SatohS ( 2014). Gibberellin-induced expression of Fe uptake-related genes in Arabidopsis Plant Cell Phy- siol 55, 87-98. DOI:10.1093/pcp/pct160URL [本文引用: 1]
MolPC, ParkHM, MullinsJT, CabibE ( 1994). A GTP- binding protein regulates the activity of (1→3)-β-glucan synthase, an enzyme directly involved in yeast cell wall morphogenesis J Biol Chem 269, 31267-31274. [本文引用: 1]
NegishiT, NakanishiH, YazakiJ, KishimotoN, FujiiF, ShimboK, YamamotoK, SakataK, SasakiT, KikuchiS, MoriS, NishizawaNK ( 2002). cDNA microarray analysis of gene expression during Fe-deficiency stress in barley suggests that polar transport of vesicles is implicated in phytosiderophore secretion in Fe-deficient barley roots Plant J 30, 83-94. DOI:10.1046/j.1365-313X.2002.01270.xURL [本文引用: 1]
NguyenVNT, MoonS, JungKH ( 2014). Genome-wide expression analysis of rice ABC transporter family across spatio-temporal samples and in response to abiotic stress- ses J Plant Physiol 171, 1276-1288. DOI:10.1016/j.jplph.2014.05.006URL [本文引用: 1]
QiYH, WangSK, ShenCJ, ZhangSN, ChenY, XuYX, LiuY, WuYR, JiangDA ( 2012). OsARF12, a transcription activator on auxin response gene, regulates root elonga- tion and affects iron accumulation in rice (Oryza sativa) New Phytol 193, 109-120. [本文引用: 1]
RobinsonNJ, ProcterCM, ConnollyEL, GuerinotML ( 1999). A ferric-chelate reductase for iron uptake from soils Nature 397, 694-697. DOI:10.1038/17800 [本文引用: 1]
ShanmugamV, WangYW, TsedneeM, KarunakaranK, YehKC ( 2015). Glutathione plays an essential role in nitric oxide-mediated iron-deficiency signaling and iron-defi- ciency tolerance in Arabidopsis Plant J 84, 464-477. DOI:10.1111/tpj.2015.84.issue-3URL [本文引用: 1]
TakahashiM, TeradaY, NakaiI, NakanishiH, YoshimuraE, MoriS, NishizawaNK ( 2003). Role of nicotianamine in the intracellular delivery of metals and plant reproductive development Plant Cell 15, 1263-1280. [本文引用: 1]
YosephT, DavidKS, DeanDP ( 2002). Overexpression of the enzyme p -hydroxyphenolpyruvate dioxygenase in Ara- bidopsis and its relation to tocopherol biosynthesis Plant Physiol Biochem 40, 913-920. [本文引用: 1]
TanakaN, Abe-DohmaeS, IwamotoN, FitzgeraldML, YokoyamaS ( 2010). Helical apolipoproteins of high- density lipoprotein enhance phagocytosis by stabilizing ATP-binding cassette transporter A7 J Lipid Res 51, 2591-2599. DOI:10.1194/jlr.M006049URL [本文引用: 1]
Response of Arabidopsis to iron deficiency stress as revealed by microarray analysis 1 2001
... ABC转运体参与植物的多种生理代谢过程, 主要利用ATP水解的能量转运金属离子等相关物质, 以及参与非生物胁迫反应和矿质元素缺乏等生理过程.前人研究表明, ABC转运体的转运底物包括无机离子、金属离子、肽、疏水化合物、核苷和糖(Vasiliou et al., 2009).水稻中ABC家族的16个基因在矿质营养(包括镉和铁)缺乏情况下上调或下调表达(Nguyen et al., 2014).我们前期研究表明, 缺铁水稻叶片和根系中, 与能量相关的蛋白代谢(如ATP合成、糖酵解和三羧酸循环)有所上调(Chen et al., 2015).这与拟南芥和甜菜(Beta vulgaris)缺铁处理后的组学研究结果一致, 缺铁的甜菜中ATP含量是对照根系的5倍(López- Millán et al., 2000; Thimm et al., 2001); 同样在番茄(Lycopersicon esculentum)根中, 其ATP合成酶在缺铁时表达丰度显著上调(Li et al., 2008).在黄瓜(Cucumis sativus)中, 缺铁会诱导储存的碳水化合物分解生成NADPH、ATP和磷酸丙酮酸等能量物质(Espen et al., 2000).产生的能量物质将被卡尔文循环和糖酵解途径利用来补偿因光合下降而减少的能量物质.本研究中, 缺铁水稻韧皮部汁液中的ABCA7蛋白丰度上调, 可能是由于缺铁后水稻ATP含量增加, 为ABCA7蛋白提供了充分的能量转运底物, 最终导致铁离子的转运增强.缺铁条件下, 植株从环境中吸收的铁减少, 可能通过增加植株内储存的铁再分配来维持其正常生理代谢功能.缺铁后, 水稻地上部与地下部铁吸收转运相关的基因显著上调表达, 也进一步验证了地上部铁转运再分配能力增强.近期, 也有研究表明, 拟南芥中ABC转运体可以通过调控根系铁的富集来调节缺磷响应(Dong et al., 2017), 而ABC转运体在缺铁水稻植株中的功能还有待进一步探索. ...
Iron translocation II. Citrate/iron ratios in plant stem exudates 1 1966
... 铁进入水稻根系后, 主要通过蒸腾作用和主动转运由地下向地上运输.铁在植物体内的运输一般以复合体的形式进行, 主要与柠檬酸(citrate) (Tiffin, 1966; Brown and Chaney, 1971)、尼克酰胺(NA) (Hell and Stephan, 2003; Takahashi et al., 2003)和麦根酸(MAs) (Aoyama et al., 2009; Kakei et al., 2009)等结合.在木质部中, Fe(III)-柠檬酸是主要的运输形式.FRD3属于MATE家族成员, 定位于根质膜(Green and Rogers, 2004), 其主要功能是将根部的柠檬酸转运到地上木质部.水稻中有6个FRD3的同源基因, 敲除OsFRDL1会导致叶片失绿(Yokosho et al., 2009).韧皮部中铁的螯合体主要是NA, NA不仅能够与Fe(II)络合, 也可以络合Fe(III) (von Wirén et al., 1999).在韧皮部中, Fe-NA由YSL家族成员转运(Haydon and Cobbett, 2007; Curie et al., 2009).水稻的YSL表达模式分析表明, 有些成员同时在地上部和地下部表达(如OsYSL6、OsYSL14和OsYSL16).OsYSL2和OsYSL15受缺铁响应调控, OsYSL2转运Fe-NA, 而OsYSL15转运Fe-PS (Inoue et al., 2009). ...
Evolutionarily conserved regulatory mechanisms of abscisic acid signa- ling in land plants: characterization of ABSCISIC ACID INSENSITIVE1-like type 2C protein phosphatase in the liverwort Marchantia polymorpha 1 2010
... 蛋白激酶和蛋白磷酸酶(PPs)介导的可逆磷酸化是普遍存在于生物体内的信号转导机制.蛋白磷酸酶包括PP1、PP2A、PP2B和PP2C, PP2Cs是植物中最大的蛋白磷酸酶家族(Kerk et al., 2002).在高等植物中, PP2C参与多种信号转导途径, 包括ABA、环境胁迫以及生长发育等(Tougane et al., 2010).ABA信号转导途径主要有PP2Cs和SnRK2两种家族蛋白参与.ABA含量较高时, 其受体能够与PYR/PYL家族蛋白结合, 进而结合PP2Cs蛋白的催化位点, 抑制其活性; 相应地, SnRK2家族蛋白的活性得以激活, 从而激活ABA信号途径(Soon et al., 2012).本研究中, 缺铁条件下水稻韧皮部中参与脱落酸信号转导的蛋白磷酸酶(protein phosphatase 2C 30)表达丰度下调(图2), 表明韧皮部中ABA的含量可能增多, 且转运增强.ABA在植物面临高温和干旱等逆境时会被显著诱导, 是植物遭受逆境胁迫的重要标志之一.同样, 在植物面临缺铁胁迫时, 其内源ABA含量也可能激增. ...
Human ATP- binding cassette (ABC) transporter family 2 2009
... ATP结合盒式蛋白(ATP-binding cassette transporter, ABC)是以ATP水解释放的能量为能源进行主动转运的载体蛋白家族.ABCA7 (ABC transporter A family member 7)是ABC转运蛋白基因超家族成员, 利用ATP水解的能量在细胞脂质膜和细胞内细胞器膜上主动运输一系列底物(Dean et al., 2001; Vasiliou et al., 2009; Tanaka et al., 2010).本研究中, ABCA7蛋白表达丰度上调, 表明铁离子的转运增强.对缺铁处理3天后铁吸收转运相关基因的表达水平进行测定, 发现参与还原途径的基因OsIRT1几乎不被诱导, 而参与螯合途径的OsYSL5、OsNAS1和OsNAS2基因表达被显著诱导(图5A), 地上部铁转运相关基因(OsYSL2和OsNAS1)的表达也被显著诱导(图5B), 表明铁的转运增加.此外, 参与蛋白质合成和核糖体代谢的相关蛋白表达丰度也发生变化(表1), 表明缺铁会影响水稻植株mRNA的转录及翻译过程, 但该结论有待进一步实验验证. ...
... ABC转运体参与植物的多种生理代谢过程, 主要利用ATP水解的能量转运金属离子等相关物质, 以及参与非生物胁迫反应和矿质元素缺乏等生理过程.前人研究表明, ABC转运体的转运底物包括无机离子、金属离子、肽、疏水化合物、核苷和糖(Vasiliou et al., 2009).水稻中ABC家族的16个基因在矿质营养(包括镉和铁)缺乏情况下上调或下调表达(Nguyen et al., 2014).我们前期研究表明, 缺铁水稻叶片和根系中, 与能量相关的蛋白代谢(如ATP合成、糖酵解和三羧酸循环)有所上调(Chen et al., 2015).这与拟南芥和甜菜(Beta vulgaris)缺铁处理后的组学研究结果一致, 缺铁的甜菜中ATP含量是对照根系的5倍(López- Millán et al., 2000; Thimm et al., 2001); 同样在番茄(Lycopersicon esculentum)根中, 其ATP合成酶在缺铁时表达丰度显著上调(Li et al., 2008).在黄瓜(Cucumis sativus)中, 缺铁会诱导储存的碳水化合物分解生成NADPH、ATP和磷酸丙酮酸等能量物质(Espen et al., 2000).产生的能量物质将被卡尔文循环和糖酵解途径利用来补偿因光合下降而减少的能量物质.本研究中, 缺铁水稻韧皮部汁液中的ABCA7蛋白丰度上调, 可能是由于缺铁后水稻ATP含量增加, 为ABCA7蛋白提供了充分的能量转运底物, 最终导致铁离子的转运增强.缺铁条件下, 植株从环境中吸收的铁减少, 可能通过增加植株内储存的铁再分配来维持其正常生理代谢功能.缺铁后, 水稻地上部与地下部铁吸收转运相关的基因显著上调表达, 也进一步验证了地上部铁转运再分配能力增强.近期, 也有研究表明, 拟南芥中ABC转运体可以通过调控根系铁的富集来调节缺磷响应(Dong et al., 2017), 而ABC转运体在缺铁水稻植株中的功能还有待进一步探索. ...
Dual regulation of the Arabidopsis high-affinity root iron uptake system by local and long-distance signals 2 2003
... 地上部到地下部的信号传递会影响植物对养分的吸收利用效率(Forde, 2002; Enomoto et al., 2007).冠-根和根-冠组成长距离信号传递通道, 当根系感受到养分状况变化时, 会产生相应信号并通过木质部传输到地上部, 进而激发地上部产生信号并通过韧皮部传输到根系, 从而对植物根系生长进行反馈调节, 使植物对养分的吸收与地上部代谢相平衡(Vert et al., 2003; 熊长明等, 2014). ...
... Vert等(2003)在拟南芥中提出了2种与铁吸收相关的模型: 促进模型和抑制模型.促进模型指地上部产生的信号传到地下部, 诱导缺铁根系铁吸收相关基因的表达, 而在正常供铁情况下, 地上部并无信号传到地下部.与之相反的抑制模型是在铁正常供给情况下, 地上部会产生抑制信号并传输到地下部, 进而抑制根系对铁的吸收, 而缺铁时地上部则不产生信号.由此可见, 地上部长距离信号对植物缺铁响应有重要调控作用(Vert et al., 2003; Enomoto et al., 2007; García et al., 2013).韧皮部是连接地上和地下部的重要维管组织, 不仅可以传递植物生长发育相关信息, 还可以运输相关同化产物.韧皮部汁液成分包括大分子糖、氨基酸、有机复合物、蛋白质、矿质元素以及一些信号分子, 如激素、系统伤流信号和小RNA等(Lucas et al., 2001, 2013), 这些物质参与多种胁迫(包括矿质元素缺乏)响应. ...
Nicotianamine chelates both Fe III and Fe II. Implications for metal transport in plants 1 1999
... 铁进入水稻根系后, 主要通过蒸腾作用和主动转运由地下向地上运输.铁在植物体内的运输一般以复合体的形式进行, 主要与柠檬酸(citrate) (Tiffin, 1966; Brown and Chaney, 1971)、尼克酰胺(NA) (Hell and Stephan, 2003; Takahashi et al., 2003)和麦根酸(MAs) (Aoyama et al., 2009; Kakei et al., 2009)等结合.在木质部中, Fe(III)-柠檬酸是主要的运输形式.FRD3属于MATE家族成员, 定位于根质膜(Green and Rogers, 2004), 其主要功能是将根部的柠檬酸转运到地上木质部.水稻中有6个FRD3的同源基因, 敲除OsFRDL1会导致叶片失绿(Yokosho et al., 2009).韧皮部中铁的螯合体主要是NA, NA不仅能够与Fe(II)络合, 也可以络合Fe(III) (von Wirén et al., 1999).在韧皮部中, Fe-NA由YSL家族成员转运(Haydon and Cobbett, 2007; Curie et al., 2009).水稻的YSL表达模式分析表明, 有些成员同时在地上部和地下部表达(如OsYSL6、OsYSL14和OsYSL16).OsYSL2和OsYSL15受缺铁响应调控, OsYSL2转运Fe-NA, 而OsYSL15转运Fe-PS (Inoue et al., 2009). ...
Gibberellins regulate iron deficiency-response by influencing iron trans- port and translocation in rice seedlings (Oryza sativa) 1 2017
... 赤霉素是一种多功能的植物激素, 不仅参与拟南芥的缺磷响应(Jiang et al., 2007), 而且在缺铁响应中也扮演重要角色.例如, 缺铁条件下, 对赤霉素缺失的拟南芥突变体进行外源赤霉素处理后, 参与缺铁响应的基因上调表达(Matsuoka et al., 2014).在水稻中, 赤霉素通过调控OsYSL2的表达抑制铁向地上部转运和再分配过程.缺铁会导致水稻植株体内赤霉素含量和活性降低, 外源喷施赤霉素抑制铁向地上部转运(Wang et al., 2017).本研究中, 缺铁水稻植株地上与地下部赤霉素含量均下降, 与前人研究结果一致.同时, 水稻韧皮部中赤霉素调控的蛋白表达丰度发生变化, 表明赤霉素信号可能参与水稻响应缺铁过程. ...
OsFRDL1 is a citrate transporter required for efficient translocation of iron in rice 1 2009
... 铁进入水稻根系后, 主要通过蒸腾作用和主动转运由地下向地上运输.铁在植物体内的运输一般以复合体的形式进行, 主要与柠檬酸(citrate) (Tiffin, 1966; Brown and Chaney, 1971)、尼克酰胺(NA) (Hell and Stephan, 2003; Takahashi et al., 2003)和麦根酸(MAs) (Aoyama et al., 2009; Kakei et al., 2009)等结合.在木质部中, Fe(III)-柠檬酸是主要的运输形式.FRD3属于MATE家族成员, 定位于根质膜(Green and Rogers, 2004), 其主要功能是将根部的柠檬酸转运到地上木质部.水稻中有6个FRD3的同源基因, 敲除OsFRDL1会导致叶片失绿(Yokosho et al., 2009).韧皮部中铁的螯合体主要是NA, NA不仅能够与Fe(II)络合, 也可以络合Fe(III) (von Wirén et al., 1999).在韧皮部中, Fe-NA由YSL家族成员转运(Haydon and Cobbett, 2007; Curie et al., 2009).水稻的YSL表达模式分析表明, 有些成员同时在地上部和地下部表达(如OsYSL6、OsYSL14和OsYSL16).OsYSL2和OsYSL15受缺铁响应调控, OsYSL2转运Fe-NA, 而OsYSL15转运Fe-PS (Inoue et al., 2009). ...
OsYSL18 is a rice iron(III)-deoxy- mugineic acid transporter specifically expressed in reproductive organs and phloem of lamina joints 1 2009
... 铁进入水稻根系后, 主要通过蒸腾作用和主动转运由地下向地上运输.铁在植物体内的运输一般以复合体的形式进行, 主要与柠檬酸(citrate) (Tiffin, 1966; Brown and Chaney, 1971)、尼克酰胺(NA) (Hell and Stephan, 2003; Takahashi et al., 2003)和麦根酸(MAs) (Aoyama et al., 2009; Kakei et al., 2009)等结合.在木质部中, Fe(III)-柠檬酸是主要的运输形式.FRD3属于MATE家族成员, 定位于根质膜(Green and Rogers, 2004), 其主要功能是将根部的柠檬酸转运到地上木质部.水稻中有6个FRD3的同源基因, 敲除OsFRDL1会导致叶片失绿(Yokosho et al., 2009).韧皮部中铁的螯合体主要是NA, NA不仅能够与Fe(II)络合, 也可以络合Fe(III) (von Wirén et al., 1999).在韧皮部中, Fe-NA由YSL家族成员转运(Haydon and Cobbett, 2007; Curie et al., 2009).水稻的YSL表达模式分析表明, 有些成员同时在地上部和地下部表达(如OsYSL6、OsYSL14和OsYSL16).OsYSL2和OsYSL15受缺铁响应调控, OsYSL2转运Fe-NA, 而OsYSL15转运Fe-PS (Inoue et al., 2009). ...
Evidence that the auxin signaling path- way interacts with plant stress response 1 2002
... 生长素原初响应基因主要分为3类: Aux/IAAs、GH3s (Gretchen Hagen 3)和SAURs (Small Auxin Up RNAs).植物主要通过GH3调控体内生长素的动态平衡, GH3基因与植物响应环境胁迫密切相关(Hagen et al., 1988; Bao and Li, 2002), 且GH3基因的表达受植株体内生长素水平的影响.本研究中, 缺铁条件下水稻韧皮部中参与生长素信号转导的吲哚乙酸酰胺合成酶(indole-3-acetic acid-amido synthetase GH3.6)表达丰度下调(图2), 表明植株韧皮部中的生长素含量偏低.生长素主要在幼嫩组织(叶片)中合成, 然后转运至根系调控植物生长发育.缺铁条件下, 韧皮部中生长素含量降低, 导致从地上部向地下部转运的生长素减少.另外, 参与色氨酸生物合成的色氨酸合成酶(tryptophan synthase alpha chain)也下调表达.生长素的合成需要色氨酸参与, 其合成酶下调表达也表明生长素合成有所减弱. ...
Expres- sion and enzyme activity of glutathione reductase is upregulated by Fe-deficiency in graminaceous plants 1 2007
Effect of iron on the transport of citrate into the xylem of soybeans and tomatoes 1 1971
... 铁进入水稻根系后, 主要通过蒸腾作用和主动转运由地下向地上运输.铁在植物体内的运输一般以复合体的形式进行, 主要与柠檬酸(citrate) (Tiffin, 1966; Brown and Chaney, 1971)、尼克酰胺(NA) (Hell and Stephan, 2003; Takahashi et al., 2003)和麦根酸(MAs) (Aoyama et al., 2009; Kakei et al., 2009)等结合.在木质部中, Fe(III)-柠檬酸是主要的运输形式.FRD3属于MATE家族成员, 定位于根质膜(Green and Rogers, 2004), 其主要功能是将根部的柠檬酸转运到地上木质部.水稻中有6个FRD3的同源基因, 敲除OsFRDL1会导致叶片失绿(Yokosho et al., 2009).韧皮部中铁的螯合体主要是NA, NA不仅能够与Fe(II)络合, 也可以络合Fe(III) (von Wirén et al., 1999).在韧皮部中, Fe-NA由YSL家族成员转运(Haydon and Cobbett, 2007; Curie et al., 2009).水稻的YSL表达模式分析表明, 有些成员同时在地上部和地下部表达(如OsYSL6、OsYSL14和OsYSL16).OsYSL2和OsYSL15受缺铁响应调控, OsYSL2转运Fe-NA, 而OsYSL15转运Fe-PS (Inoue et al., 2009). ...
Differential regulation of proteins in rice ( Oryza sativa L.) under iron deficiency 1 2015
... ABC转运体参与植物的多种生理代谢过程, 主要利用ATP水解的能量转运金属离子等相关物质, 以及参与非生物胁迫反应和矿质元素缺乏等生理过程.前人研究表明, ABC转运体的转运底物包括无机离子、金属离子、肽、疏水化合物、核苷和糖(Vasiliou et al., 2009).水稻中ABC家族的16个基因在矿质营养(包括镉和铁)缺乏情况下上调或下调表达(Nguyen et al., 2014).我们前期研究表明, 缺铁水稻叶片和根系中, 与能量相关的蛋白代谢(如ATP合成、糖酵解和三羧酸循环)有所上调(Chen et al., 2015).这与拟南芥和甜菜(Beta vulgaris)缺铁处理后的组学研究结果一致, 缺铁的甜菜中ATP含量是对照根系的5倍(López- Millán et al., 2000; Thimm et al., 2001); 同样在番茄(Lycopersicon esculentum)根中, 其ATP合成酶在缺铁时表达丰度显著上调(Li et al., 2008).在黄瓜(Cucumis sativus)中, 缺铁会诱导储存的碳水化合物分解生成NADPH、ATP和磷酸丙酮酸等能量物质(Espen et al., 2000).产生的能量物质将被卡尔文循环和糖酵解途径利用来补偿因光合下降而减少的能量物质.本研究中, 缺铁水稻韧皮部汁液中的ABCA7蛋白丰度上调, 可能是由于缺铁后水稻ATP含量增加, 为ABCA7蛋白提供了充分的能量转运底物, 最终导致铁离子的转运增强.缺铁条件下, 植株从环境中吸收的铁减少, 可能通过增加植株内储存的铁再分配来维持其正常生理代谢功能.缺铁后, 水稻地上部与地下部铁吸收转运相关的基因显著上调表达, 也进一步验证了地上部铁转运再分配能力增强.近期, 也有研究表明, 拟南芥中ABC转运体可以通过调控根系铁的富集来调节缺磷响应(Dong et al., 2017), 而ABC转运体在缺铁水稻植株中的功能还有待进一步探索. ...
Shoot-root communication plays a key role in physiological alterations of rice (Oryza sativa) under iron deficiency 3 2018
Sucrose is involved in the regulation of iron deficiency responses in rice ( Oryza sativa L.) 1 2018
... 质体醌和维生素E生物合成途径的关键步骤由4-羟基苯基丙酮酸双加氧酶(4-hydroxyphenylpyruvate dioxygenase, HPPD)催化, 目前已在胡萝卜(Daucus carota)、拟南芥和大麦(Hordeum vulgare)等植物中鉴定出HPPD基因(Falka et al., 2003).研究表明, 在拟南芥中过表达hppd基因使其种子和叶片中的生育酚含量增加(Yoseph et al., 2002).质体醌参与光合作用过程中光反应的电子传递.缺铁水稻植株韧皮部汁液中, 4-羟基苯基丙酮酸双加氧酶蛋白丰度下调, 水稻维生素E的含量也减少, 光合系统被破坏.前期的研究表明, 缺铁后水稻植株的光合速率显著下降(Chen et al., 2018b), 推测可能是缺铁使光反应效率降低, 最终导致光合速率下降. ...
Mutation in nicotianamine aminotransferase stimulated the Fe(II) acquisition system and led to iron accumulation in rice 1 2007
... 铁是植物生长必需的矿质营养.为适应低铁环境, 植物进化出2种不同的吸收机制: 还原途径(机理I)和螯合途径(机理II).通过机理I吸收铁的植物(以下简称机理I植物)包括双子叶植物和非禾本科单子叶植物; 通过机理II吸收铁的植物(以下简称机理II植物)主要是禾本科植物.东京大学Nishizawa的研究发现, 机理II植物水稻(Oryza sativa)也能通过机理I的方式吸收铁(Ishimaru et al., 2006), 即水稻兼具2条途径进行铁的吸收, 该结论后经吴平教授实验室用突变体证实(Cheng et al., 2007). ...
Overexpression of the FRO2 iron reductase confers tolerance to growth on low iron and uncovers post-transcriptional control 1 2003
... 在感受到缺铁信号时, 机理I植物根部表皮会诱导产生3种反应: (1) H+-ATPase大量表达, 酸化根际土壤环境, 从而提高根际铁的有效性; (2) 铁还原酶FRO (ferric reductase oxidase)基因表达上调, 催化根表皮的三价铁(Fe(III))还原成二价铁(Fe(II)); (3) Fe(II)的转运体IRT (iron-regulated transporter)表达上调, 将被FRO还原成的Fe(II)转运入根部细胞(吴慧兰等, 2007).该过程中的关键基因FRO (Robinson et al., 1999)和IRT (Eide et al., 1996)最早于20世纪90年代在拟南芥(Arabidopsis thaliana)中克隆得到, 之后相继报道了它们在多种植物中的同源基因(Con- nolly et al., 2003; Ishimaru et al., 2007). ...
Metal movement within the plant: contribution of nico- tianamine and yellow stripe 1-like transporters 1 2009
... 铁进入水稻根系后, 主要通过蒸腾作用和主动转运由地下向地上运输.铁在植物体内的运输一般以复合体的形式进行, 主要与柠檬酸(citrate) (Tiffin, 1966; Brown and Chaney, 1971)、尼克酰胺(NA) (Hell and Stephan, 2003; Takahashi et al., 2003)和麦根酸(MAs) (Aoyama et al., 2009; Kakei et al., 2009)等结合.在木质部中, Fe(III)-柠檬酸是主要的运输形式.FRD3属于MATE家族成员, 定位于根质膜(Green and Rogers, 2004), 其主要功能是将根部的柠檬酸转运到地上木质部.水稻中有6个FRD3的同源基因, 敲除OsFRDL1会导致叶片失绿(Yokosho et al., 2009).韧皮部中铁的螯合体主要是NA, NA不仅能够与Fe(II)络合, 也可以络合Fe(III) (von Wirén et al., 1999).在韧皮部中, Fe-NA由YSL家族成员转运(Haydon and Cobbett, 2007; Curie et al., 2009).水稻的YSL表达模式分析表明, 有些成员同时在地上部和地下部表达(如OsYSL6、OsYSL14和OsYSL16).OsYSL2和OsYSL15受缺铁响应调控, OsYSL2转运Fe-NA, 而OsYSL15转运Fe-PS (Inoue et al., 2009). ...
Maize yellow stripe1 encodes a membrane protein directly involved in Fe(III) uptake 1 2001
... 水稻的另一种铁吸收方式为螯合方式, 该吸收途径通过合成一种高亲和的与三价铁结合的物质——植物铁载体PS (phytosiderophore), 分泌到根际土壤环境中, 之后与Fe(III)结合生成Fe(III)-PS复合体(Ne- gishi et al., 2002).同时, 植物表皮细胞质膜上的一种多肽转运体OPT (oligo-peptide transporter)家族成员YS (Yellow Stripe)转运体(Curie et al., 2001)可将上述根表皮细胞附近的Fe(III)-PS复合体跨膜转运到植物体内.OsYSL15是水稻吸收铁过程中Fe(III)-PS的主要转运体(Inoue et al., 2009; Lee et al., 2009), 其受缺铁诱导上调表达. ...
The human ATP- binding cassette (ABC) transporter superfamily 1 2001
... ATP结合盒式蛋白(ATP-binding cassette transporter, ABC)是以ATP水解释放的能量为能源进行主动转运的载体蛋白家族.ABCA7 (ABC transporter A family member 7)是ABC转运蛋白基因超家族成员, 利用ATP水解的能量在细胞脂质膜和细胞内细胞器膜上主动运输一系列底物(Dean et al., 2001; Vasiliou et al., 2009; Tanaka et al., 2010).本研究中, ABCA7蛋白表达丰度上调, 表明铁离子的转运增强.对缺铁处理3天后铁吸收转运相关基因的表达水平进行测定, 发现参与还原途径的基因OsIRT1几乎不被诱导, 而参与螯合途径的OsYSL5、OsNAS1和OsNAS2基因表达被显著诱导(图5A), 地上部铁转运相关基因(OsYSL2和OsNAS1)的表达也被显著诱导(图5B), 表明铁的转运增加.此外, 参与蛋白质合成和核糖体代谢的相关蛋白表达丰度也发生变化(表1), 表明缺铁会影响水稻植株mRNA的转录及翻译过程, 但该结论有待进一步实验验证. ...
An Arabidopsis ABC trans- porter mediates phosphate deficiency-induced remodeling of root architecture by modulating iron homeostasis in roots 1 2017
... ABC转运体参与植物的多种生理代谢过程, 主要利用ATP水解的能量转运金属离子等相关物质, 以及参与非生物胁迫反应和矿质元素缺乏等生理过程.前人研究表明, ABC转运体的转运底物包括无机离子、金属离子、肽、疏水化合物、核苷和糖(Vasiliou et al., 2009).水稻中ABC家族的16个基因在矿质营养(包括镉和铁)缺乏情况下上调或下调表达(Nguyen et al., 2014).我们前期研究表明, 缺铁水稻叶片和根系中, 与能量相关的蛋白代谢(如ATP合成、糖酵解和三羧酸循环)有所上调(Chen et al., 2015).这与拟南芥和甜菜(Beta vulgaris)缺铁处理后的组学研究结果一致, 缺铁的甜菜中ATP含量是对照根系的5倍(López- Millán et al., 2000; Thimm et al., 2001); 同样在番茄(Lycopersicon esculentum)根中, 其ATP合成酶在缺铁时表达丰度显著上调(Li et al., 2008).在黄瓜(Cucumis sativus)中, 缺铁会诱导储存的碳水化合物分解生成NADPH、ATP和磷酸丙酮酸等能量物质(Espen et al., 2000).产生的能量物质将被卡尔文循环和糖酵解途径利用来补偿因光合下降而减少的能量物质.本研究中, 缺铁水稻韧皮部汁液中的ABCA7蛋白丰度上调, 可能是由于缺铁后水稻ATP含量增加, 为ABCA7蛋白提供了充分的能量转运底物, 最终导致铁离子的转运增强.缺铁条件下, 植株从环境中吸收的铁减少, 可能通过增加植株内储存的铁再分配来维持其正常生理代谢功能.缺铁后, 水稻地上部与地下部铁吸收转运相关的基因显著上调表达, 也进一步验证了地上部铁转运再分配能力增强.近期, 也有研究表明, 拟南芥中ABC转运体可以通过调控根系铁的富集来调节缺磷响应(Dong et al., 2017), 而ABC转运体在缺铁水稻植株中的功能还有待进一步探索. ...
A novel iron-regulated metal transporter from plants identified by functional expression in yeast 1 1996
... 在感受到缺铁信号时, 机理I植物根部表皮会诱导产生3种反应: (1) H+-ATPase大量表达, 酸化根际土壤环境, 从而提高根际铁的有效性; (2) 铁还原酶FRO (ferric reductase oxidase)基因表达上调, 催化根表皮的三价铁(Fe(III))还原成二价铁(Fe(II)); (3) Fe(II)的转运体IRT (iron-regulated transporter)表达上调, 将被FRO还原成的Fe(II)转运入根部细胞(吴慧兰等, 2007).该过程中的关键基因FRO (Robinson et al., 1999)和IRT (Eide et al., 1996)最早于20世纪90年代在拟南芥(Arabidopsis thaliana)中克隆得到, 之后相继报道了它们在多种植物中的同源基因(Con- nolly et al., 2003; Ishimaru et al., 2007). ...
Long-distance signals positively regulate the expression of iron uptake genes in tobacco roots 2 2007
... 地上部到地下部的信号传递会影响植物对养分的吸收利用效率(Forde, 2002; Enomoto et al., 2007).冠-根和根-冠组成长距离信号传递通道, 当根系感受到养分状况变化时, 会产生相应信号并通过木质部传输到地上部, 进而激发地上部产生信号并通过韧皮部传输到根系, 从而对植物根系生长进行反馈调节, 使植物对养分的吸收与地上部代谢相平衡(Vert et al., 2003; 熊长明等, 2014). ...
... Vert等(2003)在拟南芥中提出了2种与铁吸收相关的模型: 促进模型和抑制模型.促进模型指地上部产生的信号传到地下部, 诱导缺铁根系铁吸收相关基因的表达, 而在正常供铁情况下, 地上部并无信号传到地下部.与之相反的抑制模型是在铁正常供给情况下, 地上部会产生抑制信号并传输到地下部, 进而抑制根系对铁的吸收, 而缺铁时地上部则不产生信号.由此可见, 地上部长距离信号对植物缺铁响应有重要调控作用(Vert et al., 2003; Enomoto et al., 2007; García et al., 2013).韧皮部是连接地上和地下部的重要维管组织, 不仅可以传递植物生长发育相关信息, 还可以运输相关同化产物.韧皮部汁液成分包括大分子糖、氨基酸、有机复合物、蛋白质、矿质元素以及一些信号分子, 如激素、系统伤流信号和小RNA等(Lucas et al., 2001, 2013), 这些物质参与多种胁迫(包括矿质元素缺乏)响应. ...
Metabolic responses in cucumber ( Cucumis sativus L.) roots under Fe-deficiency: a 31P-nuclear magnetic resonance in-vivo study 1 2000
... ABC转运体参与植物的多种生理代谢过程, 主要利用ATP水解的能量转运金属离子等相关物质, 以及参与非生物胁迫反应和矿质元素缺乏等生理过程.前人研究表明, ABC转运体的转运底物包括无机离子、金属离子、肽、疏水化合物、核苷和糖(Vasiliou et al., 2009).水稻中ABC家族的16个基因在矿质营养(包括镉和铁)缺乏情况下上调或下调表达(Nguyen et al., 2014).我们前期研究表明, 缺铁水稻叶片和根系中, 与能量相关的蛋白代谢(如ATP合成、糖酵解和三羧酸循环)有所上调(Chen et al., 2015).这与拟南芥和甜菜(Beta vulgaris)缺铁处理后的组学研究结果一致, 缺铁的甜菜中ATP含量是对照根系的5倍(López- Millán et al., 2000; Thimm et al., 2001); 同样在番茄(Lycopersicon esculentum)根中, 其ATP合成酶在缺铁时表达丰度显著上调(Li et al., 2008).在黄瓜(Cucumis sativus)中, 缺铁会诱导储存的碳水化合物分解生成NADPH、ATP和磷酸丙酮酸等能量物质(Espen et al., 2000).产生的能量物质将被卡尔文循环和糖酵解途径利用来补偿因光合下降而减少的能量物质.本研究中, 缺铁水稻韧皮部汁液中的ABCA7蛋白丰度上调, 可能是由于缺铁后水稻ATP含量增加, 为ABCA7蛋白提供了充分的能量转运底物, 最终导致铁离子的转运增强.缺铁条件下, 植株从环境中吸收的铁减少, 可能通过增加植株内储存的铁再分配来维持其正常生理代谢功能.缺铁后, 水稻地上部与地下部铁吸收转运相关的基因显著上调表达, 也进一步验证了地上部铁转运再分配能力增强.近期, 也有研究表明, 拟南芥中ABC转运体可以通过调控根系铁的富集来调节缺磷响应(Dong et al., 2017), 而ABC转运体在缺铁水稻植株中的功能还有待进一步探索. ...
Constitutive overexpression of barley 4-hydroxyp henylpyruvate dioxygenase in tobacco results in elevation of the vitamin E content in seeds but not in leaves 1 2003
... 质体醌和维生素E生物合成途径的关键步骤由4-羟基苯基丙酮酸双加氧酶(4-hydroxyphenylpyruvate dioxygenase, HPPD)催化, 目前已在胡萝卜(Daucus carota)、拟南芥和大麦(Hordeum vulgare)等植物中鉴定出HPPD基因(Falka et al., 2003).研究表明, 在拟南芥中过表达hppd基因使其种子和叶片中的生育酚含量增加(Yoseph et al., 2002).质体醌参与光合作用过程中光反应的电子传递.缺铁水稻植株韧皮部汁液中, 4-羟基苯基丙酮酸双加氧酶蛋白丰度下调, 水稻维生素E的含量也减少, 光合系统被破坏.前期的研究表明, 缺铁后水稻植株的光合速率显著下降(Chen et al., 2018b), 推测可能是缺铁使光反应效率降低, 最终导致光合速率下降. ...
The role of long-distance signaling in plant responses to nitrate and other nutrients 1 2002
... 地上部到地下部的信号传递会影响植物对养分的吸收利用效率(Forde, 2002; Enomoto et al., 2007).冠-根和根-冠组成长距离信号传递通道, 当根系感受到养分状况变化时, 会产生相应信号并通过木质部传输到地上部, 进而激发地上部产生信号并通过韧皮部传输到根系, 从而对植物根系生长进行反馈调节, 使植物对养分的吸收与地上部代谢相平衡(Vert et al., 2003; 熊长明等, 2014). ...
Shoot to root com- munication is necessary to control the expression of iron- acquisition genes in Strategy I plants 1 2013
... Vert等(2003)在拟南芥中提出了2种与铁吸收相关的模型: 促进模型和抑制模型.促进模型指地上部产生的信号传到地下部, 诱导缺铁根系铁吸收相关基因的表达, 而在正常供铁情况下, 地上部并无信号传到地下部.与之相反的抑制模型是在铁正常供给情况下, 地上部会产生抑制信号并传输到地下部, 进而抑制根系对铁的吸收, 而缺铁时地上部则不产生信号.由此可见, 地上部长距离信号对植物缺铁响应有重要调控作用(Vert et al., 2003; Enomoto et al., 2007; García et al., 2013).韧皮部是连接地上和地下部的重要维管组织, 不仅可以传递植物生长发育相关信息, 还可以运输相关同化产物.韧皮部汁液成分包括大分子糖、氨基酸、有机复合物、蛋白质、矿质元素以及一些信号分子, 如激素、系统伤流信号和小RNA等(Lucas et al., 2001, 2013), 这些物质参与多种胁迫(包括矿质元素缺乏)响应. ...
FRD3 controls iron localiza- tion in Arabidopsis 1 2004
... 铁进入水稻根系后, 主要通过蒸腾作用和主动转运由地下向地上运输.铁在植物体内的运输一般以复合体的形式进行, 主要与柠檬酸(citrate) (Tiffin, 1966; Brown and Chaney, 1971)、尼克酰胺(NA) (Hell and Stephan, 2003; Takahashi et al., 2003)和麦根酸(MAs) (Aoyama et al., 2009; Kakei et al., 2009)等结合.在木质部中, Fe(III)-柠檬酸是主要的运输形式.FRD3属于MATE家族成员, 定位于根质膜(Green and Rogers, 2004), 其主要功能是将根部的柠檬酸转运到地上木质部.水稻中有6个FRD3的同源基因, 敲除OsFRDL1会导致叶片失绿(Yokosho et al., 2009).韧皮部中铁的螯合体主要是NA, NA不仅能够与Fe(II)络合, 也可以络合Fe(III) (von Wirén et al., 1999).在韧皮部中, Fe-NA由YSL家族成员转运(Haydon and Cobbett, 2007; Curie et al., 2009).水稻的YSL表达模式分析表明, 有些成员同时在地上部和地下部表达(如OsYSL6、OsYSL14和OsYSL16).OsYSL2和OsYSL15受缺铁响应调控, OsYSL2转运Fe-NA, 而OsYSL15转运Fe-PS (Inoue et al., 2009). ...
Regulation of expression of an auxin-induced soybean sequence by cadmium 1 1988
... 生长素原初响应基因主要分为3类: Aux/IAAs、GH3s (Gretchen Hagen 3)和SAURs (Small Auxin Up RNAs).植物主要通过GH3调控体内生长素的动态平衡, GH3基因与植物响应环境胁迫密切相关(Hagen et al., 1988; Bao and Li, 2002), 且GH3基因的表达受植株体内生长素水平的影响.本研究中, 缺铁条件下水稻韧皮部中参与生长素信号转导的吲哚乙酸酰胺合成酶(indole-3-acetic acid-amido synthetase GH3.6)表达丰度下调(图2), 表明植株韧皮部中的生长素含量偏低.生长素主要在幼嫩组织(叶片)中合成, 然后转运至根系调控植物生长发育.缺铁条件下, 韧皮部中生长素含量降低, 导致从地上部向地下部转运的生长素减少.另外, 参与色氨酸生物合成的色氨酸合成酶(tryptophan synthase alpha chain)也下调表达.生长素的合成需要色氨酸参与, 其合成酶下调表达也表明生长素合成有所减弱. ...
Transporters of ligands for essential metal ions in plants 1 2007
... 铁进入水稻根系后, 主要通过蒸腾作用和主动转运由地下向地上运输.铁在植物体内的运输一般以复合体的形式进行, 主要与柠檬酸(citrate) (Tiffin, 1966; Brown and Chaney, 1971)、尼克酰胺(NA) (Hell and Stephan, 2003; Takahashi et al., 2003)和麦根酸(MAs) (Aoyama et al., 2009; Kakei et al., 2009)等结合.在木质部中, Fe(III)-柠檬酸是主要的运输形式.FRD3属于MATE家族成员, 定位于根质膜(Green and Rogers, 2004), 其主要功能是将根部的柠檬酸转运到地上木质部.水稻中有6个FRD3的同源基因, 敲除OsFRDL1会导致叶片失绿(Yokosho et al., 2009).韧皮部中铁的螯合体主要是NA, NA不仅能够与Fe(II)络合, 也可以络合Fe(III) (von Wirén et al., 1999).在韧皮部中, Fe-NA由YSL家族成员转运(Haydon and Cobbett, 2007; Curie et al., 2009).水稻的YSL表达模式分析表明, 有些成员同时在地上部和地下部表达(如OsYSL6、OsYSL14和OsYSL16).OsYSL2和OsYSL15受缺铁响应调控, OsYSL2转运Fe-NA, 而OsYSL15转运Fe-PS (Inoue et al., 2009). ...
Iron uptake, trafficking and homeostasis in plants 1 2003
... 铁进入水稻根系后, 主要通过蒸腾作用和主动转运由地下向地上运输.铁在植物体内的运输一般以复合体的形式进行, 主要与柠檬酸(citrate) (Tiffin, 1966; Brown and Chaney, 1971)、尼克酰胺(NA) (Hell and Stephan, 2003; Takahashi et al., 2003)和麦根酸(MAs) (Aoyama et al., 2009; Kakei et al., 2009)等结合.在木质部中, Fe(III)-柠檬酸是主要的运输形式.FRD3属于MATE家族成员, 定位于根质膜(Green and Rogers, 2004), 其主要功能是将根部的柠檬酸转运到地上木质部.水稻中有6个FRD3的同源基因, 敲除OsFRDL1会导致叶片失绿(Yokosho et al., 2009).韧皮部中铁的螯合体主要是NA, NA不仅能够与Fe(II)络合, 也可以络合Fe(III) (von Wirén et al., 1999).在韧皮部中, Fe-NA由YSL家族成员转运(Haydon and Cobbett, 2007; Curie et al., 2009).水稻的YSL表达模式分析表明, 有些成员同时在地上部和地下部表达(如OsYSL6、OsYSL14和OsYSL16).OsYSL2和OsYSL15受缺铁响应调控, OsYSL2转运Fe-NA, 而OsYSL15转运Fe-PS (Inoue et al., 2009). ...
Getting a sense for signals: regulation of the plant iron deficiency response 1 2012
... 生长素(IAA)作为调控植物生长的重要激素, 对植物形态的塑造有重要作用.当水稻面临缺铁胁迫时, 植株根系伸长、根毛增多, 以增强对环境中铁的吸收, 而地上部生长则减慢以降低植株自身对铁的需求, 这些变化都需要生长素参与.已在拟南芥中证实缺铁会导致生长素含量升高, 并诱导拟南芥铁吸收相关基因(FRO2)表达上调(Hindt and Guerinot, 2012).此外, 生长素也被报道参与水稻缺铁响应, 生长素响应因子OsARF12参与水稻缺铁信号转导(Qi et al., 2012); 外源生长素处理会上调水稻植株地上部缺铁响应基因的表达, 而外源施用生长素抑制剂则会有相反的效应(Liu et al., 2015).本研究表明, 缺铁处理后地上部生长素积累增多而根系积累减少(图3), 推测可能是缺铁后水稻植株生长素的转运受到抑制, 即从地上部向地下部转运减少, 从而诱导根系响应缺铁. ...
Rice OsYSL15 is an iron-regulated iron(III)- deoxymugineic acid transporter expressed in the roots and is essential for iron uptake in early growth of the seedlings 2 2009
... 水稻的另一种铁吸收方式为螯合方式, 该吸收途径通过合成一种高亲和的与三价铁结合的物质——植物铁载体PS (phytosiderophore), 分泌到根际土壤环境中, 之后与Fe(III)结合生成Fe(III)-PS复合体(Ne- gishi et al., 2002).同时, 植物表皮细胞质膜上的一种多肽转运体OPT (oligo-peptide transporter)家族成员YS (Yellow Stripe)转运体(Curie et al., 2001)可将上述根表皮细胞附近的Fe(III)-PS复合体跨膜转运到植物体内.OsYSL15是水稻吸收铁过程中Fe(III)-PS的主要转运体(Inoue et al., 2009; Lee et al., 2009), 其受缺铁诱导上调表达. ...
... 铁进入水稻根系后, 主要通过蒸腾作用和主动转运由地下向地上运输.铁在植物体内的运输一般以复合体的形式进行, 主要与柠檬酸(citrate) (Tiffin, 1966; Brown and Chaney, 1971)、尼克酰胺(NA) (Hell and Stephan, 2003; Takahashi et al., 2003)和麦根酸(MAs) (Aoyama et al., 2009; Kakei et al., 2009)等结合.在木质部中, Fe(III)-柠檬酸是主要的运输形式.FRD3属于MATE家族成员, 定位于根质膜(Green and Rogers, 2004), 其主要功能是将根部的柠檬酸转运到地上木质部.水稻中有6个FRD3的同源基因, 敲除OsFRDL1会导致叶片失绿(Yokosho et al., 2009).韧皮部中铁的螯合体主要是NA, NA不仅能够与Fe(II)络合, 也可以络合Fe(III) (von Wirén et al., 1999).在韧皮部中, Fe-NA由YSL家族成员转运(Haydon and Cobbett, 2007; Curie et al., 2009).水稻的YSL表达模式分析表明, 有些成员同时在地上部和地下部表达(如OsYSL6、OsYSL14和OsYSL16).OsYSL2和OsYSL15受缺铁响应调控, OsYSL2转运Fe-NA, 而OsYSL15转运Fe-PS (Inoue et al., 2009). ...
Mutational reconstructed ferric chelate reductase confers enhanced tolerance in rice to iron deficiency in calcareous soil 1 2007
... 在感受到缺铁信号时, 机理I植物根部表皮会诱导产生3种反应: (1) H+-ATPase大量表达, 酸化根际土壤环境, 从而提高根际铁的有效性; (2) 铁还原酶FRO (ferric reductase oxidase)基因表达上调, 催化根表皮的三价铁(Fe(III))还原成二价铁(Fe(II)); (3) Fe(II)的转运体IRT (iron-regulated transporter)表达上调, 将被FRO还原成的Fe(II)转运入根部细胞(吴慧兰等, 2007).该过程中的关键基因FRO (Robinson et al., 1999)和IRT (Eide et al., 1996)最早于20世纪90年代在拟南芥(Arabidopsis thaliana)中克隆得到, 之后相继报道了它们在多种植物中的同源基因(Con- nolly et al., 2003; Ishimaru et al., 2007). ...
Rice plants take up iron as an Fe 3+-phytosidero- phore and as Fe 2+ 2 2006
... 铁是植物生长必需的矿质营养.为适应低铁环境, 植物进化出2种不同的吸收机制: 还原途径(机理I)和螯合途径(机理II).通过机理I吸收铁的植物(以下简称机理I植物)包括双子叶植物和非禾本科单子叶植物; 通过机理II吸收铁的植物(以下简称机理II植物)主要是禾本科植物.东京大学Nishizawa的研究发现, 机理II植物水稻(Oryza sativa)也能通过机理I的方式吸收铁(Ishimaru et al., 2006), 即水稻兼具2条途径进行铁的吸收, 该结论后经吴平教授实验室用突变体证实(Cheng et al., 2007). ...
Phosphate starvation root architecture and anthocyanin accumulation responses are modulated by the gibberellin- DELLA signaling pathway in Arabidopsis 1 2007
... 赤霉素是一种多功能的植物激素, 不仅参与拟南芥的缺磷响应(Jiang et al., 2007), 而且在缺铁响应中也扮演重要角色.例如, 缺铁条件下, 对赤霉素缺失的拟南芥突变体进行外源赤霉素处理后, 参与缺铁响应的基因上调表达(Matsuoka et al., 2014).在水稻中, 赤霉素通过调控OsYSL2的表达抑制铁向地上部转运和再分配过程.缺铁会导致水稻植株体内赤霉素含量和活性降低, 外源喷施赤霉素抑制铁向地上部转运(Wang et al., 2017).本研究中, 缺铁水稻植株地上与地下部赤霉素含量均下降, 与前人研究结果一致.同时, 水稻韧皮部中赤霉素调控的蛋白表达丰度发生变化, 表明赤霉素信号可能参与水稻响应缺铁过程. ...
A highly sensitive, quick and simple quantification method for nicotianamine and 2’-deoxymugineic acid from minimum samp- les using LC/ESI-TOF-MS achieves functional analysis of these components in plants 1 2009
... 铁进入水稻根系后, 主要通过蒸腾作用和主动转运由地下向地上运输.铁在植物体内的运输一般以复合体的形式进行, 主要与柠檬酸(citrate) (Tiffin, 1966; Brown and Chaney, 1971)、尼克酰胺(NA) (Hell and Stephan, 2003; Takahashi et al., 2003)和麦根酸(MAs) (Aoyama et al., 2009; Kakei et al., 2009)等结合.在木质部中, Fe(III)-柠檬酸是主要的运输形式.FRD3属于MATE家族成员, 定位于根质膜(Green and Rogers, 2004), 其主要功能是将根部的柠檬酸转运到地上木质部.水稻中有6个FRD3的同源基因, 敲除OsFRDL1会导致叶片失绿(Yokosho et al., 2009).韧皮部中铁的螯合体主要是NA, NA不仅能够与Fe(II)络合, 也可以络合Fe(III) (von Wirén et al., 1999).在韧皮部中, Fe-NA由YSL家族成员转运(Haydon and Cobbett, 2007; Curie et al., 2009).水稻的YSL表达模式分析表明, 有些成员同时在地上部和地下部表达(如OsYSL6、OsYSL14和OsYSL16).OsYSL2和OsYSL15受缺铁响应调控, OsYSL2转运Fe-NA, 而OsYSL15转运Fe-PS (Inoue et al., 2009). ...
The complement of protein phos- phatase catalytic subunits encoded in the genome of Arabidopsis 1 2002
... 蛋白激酶和蛋白磷酸酶(PPs)介导的可逆磷酸化是普遍存在于生物体内的信号转导机制.蛋白磷酸酶包括PP1、PP2A、PP2B和PP2C, PP2Cs是植物中最大的蛋白磷酸酶家族(Kerk et al., 2002).在高等植物中, PP2C参与多种信号转导途径, 包括ABA、环境胁迫以及生长发育等(Tougane et al., 2010).ABA信号转导途径主要有PP2Cs和SnRK2两种家族蛋白参与.ABA含量较高时, 其受体能够与PYR/PYL家族蛋白结合, 进而结合PP2Cs蛋白的催化位点, 抑制其活性; 相应地, SnRK2家族蛋白的活性得以激活, 从而激活ABA信号途径(Soon et al., 2012).本研究中, 缺铁条件下水稻韧皮部中参与脱落酸信号转导的蛋白磷酸酶(protein phosphatase 2C 30)表达丰度下调(图2), 表明韧皮部中ABA的含量可能增多, 且转运增强.ABA在植物面临高温和干旱等逆境时会被显著诱导, 是植物遭受逆境胁迫的重要标志之一.同样, 在植物面临缺铁胁迫时, 其内源ABA含量也可能激增. ...
Disruption of OsYSL15 leads to iron inefficiency in rice plants 1 2009
... 水稻的另一种铁吸收方式为螯合方式, 该吸收途径通过合成一种高亲和的与三价铁结合的物质——植物铁载体PS (phytosiderophore), 分泌到根际土壤环境中, 之后与Fe(III)结合生成Fe(III)-PS复合体(Ne- gishi et al., 2002).同时, 植物表皮细胞质膜上的一种多肽转运体OPT (oligo-peptide transporter)家族成员YS (Yellow Stripe)转运体(Curie et al., 2001)可将上述根表皮细胞附近的Fe(III)-PS复合体跨膜转运到植物体内.OsYSL15是水稻吸收铁过程中Fe(III)-PS的主要转运体(Inoue et al., 2009; Lee et al., 2009), 其受缺铁诱导上调表达. ...
Abscisic acid alleviates iron deficiency by promo- ting root iron reutilization and transport from root to shoot in Arabidopsis 1 2014
... 脱落酸(ABA)在植物胁迫响应中扮演重要角色.研究表明, 铁毒害下, ABA参与水稻铁蛋白合成, 但外源ABA并不影响缺铁条件下水稻铁蛋白的积累(Majer- us et al., 2009).此外, 在拟南芥中, ABA可通过促进铁从根系向地上部转运和再分配来激活缺铁响应.具体表现为缺铁会诱导拟南芥根系ABA的积累, 外源ABA也会促进铁向地上部转运, 从而减缓叶片萎黄症状(Lei et al., 2014).本研究表明, 缺铁会促进水稻根系ABA的积累(图3), 而韧皮部中与ABA信号转导相关的蛋白表达水平发生变化也进一步支持了我们的推测.将根系或老叶中的养分再分配是植物应对养分胁迫的一种策略, 尤其对于易移动的营养元素, 如氮、磷、钾.铁在植物体内转移性较差, 因此, 根系铁的积累远高于地上部.缺铁时, 植物将根系中的铁转移到地上部进行再分配对调控水稻籽粒中铁的富集具有一定意义, 而ABA在水稻中是否也促进铁转运尚待深入研究. ...
Proteomic response to iron deficiency in tomato root 1 2008
... ABC转运体参与植物的多种生理代谢过程, 主要利用ATP水解的能量转运金属离子等相关物质, 以及参与非生物胁迫反应和矿质元素缺乏等生理过程.前人研究表明, ABC转运体的转运底物包括无机离子、金属离子、肽、疏水化合物、核苷和糖(Vasiliou et al., 2009).水稻中ABC家族的16个基因在矿质营养(包括镉和铁)缺乏情况下上调或下调表达(Nguyen et al., 2014).我们前期研究表明, 缺铁水稻叶片和根系中, 与能量相关的蛋白代谢(如ATP合成、糖酵解和三羧酸循环)有所上调(Chen et al., 2015).这与拟南芥和甜菜(Beta vulgaris)缺铁处理后的组学研究结果一致, 缺铁的甜菜中ATP含量是对照根系的5倍(López- Millán et al., 2000; Thimm et al., 2001); 同样在番茄(Lycopersicon esculentum)根中, 其ATP合成酶在缺铁时表达丰度显著上调(Li et al., 2008).在黄瓜(Cucumis sativus)中, 缺铁会诱导储存的碳水化合物分解生成NADPH、ATP和磷酸丙酮酸等能量物质(Espen et al., 2000).产生的能量物质将被卡尔文循环和糖酵解途径利用来补偿因光合下降而减少的能量物质.本研究中, 缺铁水稻韧皮部汁液中的ABCA7蛋白丰度上调, 可能是由于缺铁后水稻ATP含量增加, 为ABCA7蛋白提供了充分的能量转运底物, 最终导致铁离子的转运增强.缺铁条件下, 植株从环境中吸收的铁减少, 可能通过增加植株内储存的铁再分配来维持其正常生理代谢功能.缺铁后, 水稻地上部与地下部铁吸收转运相关的基因显著上调表达, 也进一步验证了地上部铁转运再分配能力增强.近期, 也有研究表明, 拟南芥中ABC转运体可以通过调控根系铁的富集来调节缺磷响应(Dong et al., 2017), 而ABC转运体在缺铁水稻植株中的功能还有待进一步探索. ...
Auxin signaling is involved in iron deficiency-induced photosynthetic inhibition and shoot growth defect in rice ( Oryza sativa L.) 1 2015
... 生长素(IAA)作为调控植物生长的重要激素, 对植物形态的塑造有重要作用.当水稻面临缺铁胁迫时, 植株根系伸长、根毛增多, 以增强对环境中铁的吸收, 而地上部生长则减慢以降低植株自身对铁的需求, 这些变化都需要生长素参与.已在拟南芥中证实缺铁会导致生长素含量升高, 并诱导拟南芥铁吸收相关基因(FRO2)表达上调(Hindt and Guerinot, 2012).此外, 生长素也被报道参与水稻缺铁响应, 生长素响应因子OsARF12参与水稻缺铁信号转导(Qi et al., 2012); 外源生长素处理会上调水稻植株地上部缺铁响应基因的表达, 而外源施用生长素抑制剂则会有相反的效应(Liu et al., 2015).本研究表明, 缺铁处理后地上部生长素积累增多而根系积累减少(图3), 推测可能是缺铁后水稻植株生长素的转运受到抑制, 即从地上部向地下部转运减少, 从而诱导根系响应缺铁. ...
Responses of sugar beet roots to iron deficiency. Changes in carbon assimilation and oxygen use 1 2000
... ABC转运体参与植物的多种生理代谢过程, 主要利用ATP水解的能量转运金属离子等相关物质, 以及参与非生物胁迫反应和矿质元素缺乏等生理过程.前人研究表明, ABC转运体的转运底物包括无机离子、金属离子、肽、疏水化合物、核苷和糖(Vasiliou et al., 2009).水稻中ABC家族的16个基因在矿质营养(包括镉和铁)缺乏情况下上调或下调表达(Nguyen et al., 2014).我们前期研究表明, 缺铁水稻叶片和根系中, 与能量相关的蛋白代谢(如ATP合成、糖酵解和三羧酸循环)有所上调(Chen et al., 2015).这与拟南芥和甜菜(Beta vulgaris)缺铁处理后的组学研究结果一致, 缺铁的甜菜中ATP含量是对照根系的5倍(López- Millán et al., 2000; Thimm et al., 2001); 同样在番茄(Lycopersicon esculentum)根中, 其ATP合成酶在缺铁时表达丰度显著上调(Li et al., 2008).在黄瓜(Cucumis sativus)中, 缺铁会诱导储存的碳水化合物分解生成NADPH、ATP和磷酸丙酮酸等能量物质(Espen et al., 2000).产生的能量物质将被卡尔文循环和糖酵解途径利用来补偿因光合下降而减少的能量物质.本研究中, 缺铁水稻韧皮部汁液中的ABCA7蛋白丰度上调, 可能是由于缺铁后水稻ATP含量增加, 为ABCA7蛋白提供了充分的能量转运底物, 最终导致铁离子的转运增强.缺铁条件下, 植株从环境中吸收的铁减少, 可能通过增加植株内储存的铁再分配来维持其正常生理代谢功能.缺铁后, 水稻地上部与地下部铁吸收转运相关的基因显著上调表达, 也进一步验证了地上部铁转运再分配能力增强.近期, 也有研究表明, 拟南芥中ABC转运体可以通过调控根系铁的富集来调节缺磷响应(Dong et al., 2017), 而ABC转运体在缺铁水稻植株中的功能还有待进一步探索. ...
The plant vascular sys- tem: evolution, development and functions 1 2013
... Vert等(2003)在拟南芥中提出了2种与铁吸收相关的模型: 促进模型和抑制模型.促进模型指地上部产生的信号传到地下部, 诱导缺铁根系铁吸收相关基因的表达, 而在正常供铁情况下, 地上部并无信号传到地下部.与之相反的抑制模型是在铁正常供给情况下, 地上部会产生抑制信号并传输到地下部, 进而抑制根系对铁的吸收, 而缺铁时地上部则不产生信号.由此可见, 地上部长距离信号对植物缺铁响应有重要调控作用(Vert et al., 2003; Enomoto et al., 2007; García et al., 2013).韧皮部是连接地上和地下部的重要维管组织, 不仅可以传递植物生长发育相关信息, 还可以运输相关同化产物.韧皮部汁液成分包括大分子糖、氨基酸、有机复合物、蛋白质、矿质元素以及一些信号分子, 如激素、系统伤流信号和小RNA等(Lucas et al., 2001, 2013), 这些物质参与多种胁迫(包括矿质元素缺乏)响应. ...
RNA as a long- distance information macromolecule in plants 1 2001
... Vert等(2003)在拟南芥中提出了2种与铁吸收相关的模型: 促进模型和抑制模型.促进模型指地上部产生的信号传到地下部, 诱导缺铁根系铁吸收相关基因的表达, 而在正常供铁情况下, 地上部并无信号传到地下部.与之相反的抑制模型是在铁正常供给情况下, 地上部会产生抑制信号并传输到地下部, 进而抑制根系对铁的吸收, 而缺铁时地上部则不产生信号.由此可见, 地上部长距离信号对植物缺铁响应有重要调控作用(Vert et al., 2003; Enomoto et al., 2007; García et al., 2013).韧皮部是连接地上和地下部的重要维管组织, 不仅可以传递植物生长发育相关信息, 还可以运输相关同化产物.韧皮部汁液成分包括大分子糖、氨基酸、有机复合物、蛋白质、矿质元素以及一些信号分子, 如激素、系统伤流信号和小RNA等(Lucas et al., 2001, 2013), 这些物质参与多种胁迫(包括矿质元素缺乏)响应. ...
Abscisic acid and oxidative stress implications in overall ferritin synthesis by African rice ( Oryza glaberrima Steud.) seedlings exposed to short term iron toxicity 1 2009
... 脱落酸(ABA)在植物胁迫响应中扮演重要角色.研究表明, 铁毒害下, ABA参与水稻铁蛋白合成, 但外源ABA并不影响缺铁条件下水稻铁蛋白的积累(Majer- us et al., 2009).此外, 在拟南芥中, ABA可通过促进铁从根系向地上部转运和再分配来激活缺铁响应.具体表现为缺铁会诱导拟南芥根系ABA的积累, 外源ABA也会促进铁向地上部转运, 从而减缓叶片萎黄症状(Lei et al., 2014).本研究表明, 缺铁会促进水稻根系ABA的积累(图3), 而韧皮部中与ABA信号转导相关的蛋白表达水平发生变化也进一步支持了我们的推测.将根系或老叶中的养分再分配是植物应对养分胁迫的一种策略, 尤其对于易移动的营养元素, 如氮、磷、钾.铁在植物体内转移性较差, 因此, 根系铁的积累远高于地上部.缺铁时, 植物将根系中的铁转移到地上部进行再分配对调控水稻籽粒中铁的富集具有一定意义, 而ABA在水稻中是否也促进铁转运尚待深入研究. ...
Gibberellin-induced expression of Fe uptake-related genes in Arabidopsis 1 2014
... 赤霉素是一种多功能的植物激素, 不仅参与拟南芥的缺磷响应(Jiang et al., 2007), 而且在缺铁响应中也扮演重要角色.例如, 缺铁条件下, 对赤霉素缺失的拟南芥突变体进行外源赤霉素处理后, 参与缺铁响应的基因上调表达(Matsuoka et al., 2014).在水稻中, 赤霉素通过调控OsYSL2的表达抑制铁向地上部转运和再分配过程.缺铁会导致水稻植株体内赤霉素含量和活性降低, 外源喷施赤霉素抑制铁向地上部转运(Wang et al., 2017).本研究中, 缺铁水稻植株地上与地下部赤霉素含量均下降, 与前人研究结果一致.同时, 水稻韧皮部中赤霉素调控的蛋白表达丰度发生变化, 表明赤霉素信号可能参与水稻响应缺铁过程. ...
A GTP- binding protein regulates the activity of (1→3)-β-glucan synthase, an enzyme directly involved in yeast cell wall morphogenesis 1 1994
cDNA microarray analysis of gene expression during Fe-deficiency stress in barley suggests that polar transport of vesicles is implicated in phytosiderophore secretion in Fe-deficient barley roots 1 2002
... 水稻的另一种铁吸收方式为螯合方式, 该吸收途径通过合成一种高亲和的与三价铁结合的物质——植物铁载体PS (phytosiderophore), 分泌到根际土壤环境中, 之后与Fe(III)结合生成Fe(III)-PS复合体(Ne- gishi et al., 2002).同时, 植物表皮细胞质膜上的一种多肽转运体OPT (oligo-peptide transporter)家族成员YS (Yellow Stripe)转运体(Curie et al., 2001)可将上述根表皮细胞附近的Fe(III)-PS复合体跨膜转运到植物体内.OsYSL15是水稻吸收铁过程中Fe(III)-PS的主要转运体(Inoue et al., 2009; Lee et al., 2009), 其受缺铁诱导上调表达. ...
Genome-wide expression analysis of rice ABC transporter family across spatio-temporal samples and in response to abiotic stress- ses 1 2014
... ABC转运体参与植物的多种生理代谢过程, 主要利用ATP水解的能量转运金属离子等相关物质, 以及参与非生物胁迫反应和矿质元素缺乏等生理过程.前人研究表明, ABC转运体的转运底物包括无机离子、金属离子、肽、疏水化合物、核苷和糖(Vasiliou et al., 2009).水稻中ABC家族的16个基因在矿质营养(包括镉和铁)缺乏情况下上调或下调表达(Nguyen et al., 2014).我们前期研究表明, 缺铁水稻叶片和根系中, 与能量相关的蛋白代谢(如ATP合成、糖酵解和三羧酸循环)有所上调(Chen et al., 2015).这与拟南芥和甜菜(Beta vulgaris)缺铁处理后的组学研究结果一致, 缺铁的甜菜中ATP含量是对照根系的5倍(López- Millán et al., 2000; Thimm et al., 2001); 同样在番茄(Lycopersicon esculentum)根中, 其ATP合成酶在缺铁时表达丰度显著上调(Li et al., 2008).在黄瓜(Cucumis sativus)中, 缺铁会诱导储存的碳水化合物分解生成NADPH、ATP和磷酸丙酮酸等能量物质(Espen et al., 2000).产生的能量物质将被卡尔文循环和糖酵解途径利用来补偿因光合下降而减少的能量物质.本研究中, 缺铁水稻韧皮部汁液中的ABCA7蛋白丰度上调, 可能是由于缺铁后水稻ATP含量增加, 为ABCA7蛋白提供了充分的能量转运底物, 最终导致铁离子的转运增强.缺铁条件下, 植株从环境中吸收的铁减少, 可能通过增加植株内储存的铁再分配来维持其正常生理代谢功能.缺铁后, 水稻地上部与地下部铁吸收转运相关的基因显著上调表达, 也进一步验证了地上部铁转运再分配能力增强.近期, 也有研究表明, 拟南芥中ABC转运体可以通过调控根系铁的富集来调节缺磷响应(Dong et al., 2017), 而ABC转运体在缺铁水稻植株中的功能还有待进一步探索. ...
OsARF12, a transcription activator on auxin response gene, regulates root elonga- tion and affects iron accumulation in rice (Oryza sativa) 1 2012
... 生长素(IAA)作为调控植物生长的重要激素, 对植物形态的塑造有重要作用.当水稻面临缺铁胁迫时, 植株根系伸长、根毛增多, 以增强对环境中铁的吸收, 而地上部生长则减慢以降低植株自身对铁的需求, 这些变化都需要生长素参与.已在拟南芥中证实缺铁会导致生长素含量升高, 并诱导拟南芥铁吸收相关基因(FRO2)表达上调(Hindt and Guerinot, 2012).此外, 生长素也被报道参与水稻缺铁响应, 生长素响应因子OsARF12参与水稻缺铁信号转导(Qi et al., 2012); 外源生长素处理会上调水稻植株地上部缺铁响应基因的表达, 而外源施用生长素抑制剂则会有相反的效应(Liu et al., 2015).本研究表明, 缺铁处理后地上部生长素积累增多而根系积累减少(图3), 推测可能是缺铁后水稻植株生长素的转运受到抑制, 即从地上部向地下部转运减少, 从而诱导根系响应缺铁. ...
A ferric-chelate reductase for iron uptake from soils 1 1999
... 在感受到缺铁信号时, 机理I植物根部表皮会诱导产生3种反应: (1) H+-ATPase大量表达, 酸化根际土壤环境, 从而提高根际铁的有效性; (2) 铁还原酶FRO (ferric reductase oxidase)基因表达上调, 催化根表皮的三价铁(Fe(III))还原成二价铁(Fe(II)); (3) Fe(II)的转运体IRT (iron-regulated transporter)表达上调, 将被FRO还原成的Fe(II)转运入根部细胞(吴慧兰等, 2007).该过程中的关键基因FRO (Robinson et al., 1999)和IRT (Eide et al., 1996)最早于20世纪90年代在拟南芥(Arabidopsis thaliana)中克隆得到, 之后相继报道了它们在多种植物中的同源基因(Con- nolly et al., 2003; Ishimaru et al., 2007). ...
Glutathione plays an essential role in nitric oxide-mediated iron-deficiency signaling and iron-defi- ciency tolerance in Arabidopsis 1 2015
Molecular mimicry regulates ABA signaling by SnRK2 kinases and PP2C phosphatases 1 2012
... 蛋白激酶和蛋白磷酸酶(PPs)介导的可逆磷酸化是普遍存在于生物体内的信号转导机制.蛋白磷酸酶包括PP1、PP2A、PP2B和PP2C, PP2Cs是植物中最大的蛋白磷酸酶家族(Kerk et al., 2002).在高等植物中, PP2C参与多种信号转导途径, 包括ABA、环境胁迫以及生长发育等(Tougane et al., 2010).ABA信号转导途径主要有PP2Cs和SnRK2两种家族蛋白参与.ABA含量较高时, 其受体能够与PYR/PYL家族蛋白结合, 进而结合PP2Cs蛋白的催化位点, 抑制其活性; 相应地, SnRK2家族蛋白的活性得以激活, 从而激活ABA信号途径(Soon et al., 2012).本研究中, 缺铁条件下水稻韧皮部中参与脱落酸信号转导的蛋白磷酸酶(protein phosphatase 2C 30)表达丰度下调(图2), 表明韧皮部中ABA的含量可能增多, 且转运增强.ABA在植物面临高温和干旱等逆境时会被显著诱导, 是植物遭受逆境胁迫的重要标志之一.同样, 在植物面临缺铁胁迫时, 其内源ABA含量也可能激增. ...
Role of nicotianamine in the intracellular delivery of metals and plant reproductive development 1 2003
... 铁进入水稻根系后, 主要通过蒸腾作用和主动转运由地下向地上运输.铁在植物体内的运输一般以复合体的形式进行, 主要与柠檬酸(citrate) (Tiffin, 1966; Brown and Chaney, 1971)、尼克酰胺(NA) (Hell and Stephan, 2003; Takahashi et al., 2003)和麦根酸(MAs) (Aoyama et al., 2009; Kakei et al., 2009)等结合.在木质部中, Fe(III)-柠檬酸是主要的运输形式.FRD3属于MATE家族成员, 定位于根质膜(Green and Rogers, 2004), 其主要功能是将根部的柠檬酸转运到地上木质部.水稻中有6个FRD3的同源基因, 敲除OsFRDL1会导致叶片失绿(Yokosho et al., 2009).韧皮部中铁的螯合体主要是NA, NA不仅能够与Fe(II)络合, 也可以络合Fe(III) (von Wirén et al., 1999).在韧皮部中, Fe-NA由YSL家族成员转运(Haydon and Cobbett, 2007; Curie et al., 2009).水稻的YSL表达模式分析表明, 有些成员同时在地上部和地下部表达(如OsYSL6、OsYSL14和OsYSL16).OsYSL2和OsYSL15受缺铁响应调控, OsYSL2转运Fe-NA, 而OsYSL15转运Fe-PS (Inoue et al., 2009). ...
Overexpression of the enzyme p -hydroxyphenolpyruvate dioxygenase in Ara- bidopsis and its relation to tocopherol biosynthesis 1 2002
... 质体醌和维生素E生物合成途径的关键步骤由4-羟基苯基丙酮酸双加氧酶(4-hydroxyphenylpyruvate dioxygenase, HPPD)催化, 目前已在胡萝卜(Daucus carota)、拟南芥和大麦(Hordeum vulgare)等植物中鉴定出HPPD基因(Falka et al., 2003).研究表明, 在拟南芥中过表达hppd基因使其种子和叶片中的生育酚含量增加(Yoseph et al., 2002).质体醌参与光合作用过程中光反应的电子传递.缺铁水稻植株韧皮部汁液中, 4-羟基苯基丙酮酸双加氧酶蛋白丰度下调, 水稻维生素E的含量也减少, 光合系统被破坏.前期的研究表明, 缺铁后水稻植株的光合速率显著下降(Chen et al., 2018b), 推测可能是缺铁使光反应效率降低, 最终导致光合速率下降. ...
Helical apolipoproteins of high- density lipoprotein enhance phagocytosis by stabilizing ATP-binding cassette transporter A7 1 2010
... ATP结合盒式蛋白(ATP-binding cassette transporter, ABC)是以ATP水解释放的能量为能源进行主动转运的载体蛋白家族.ABCA7 (ABC transporter A family member 7)是ABC转运蛋白基因超家族成员, 利用ATP水解的能量在细胞脂质膜和细胞内细胞器膜上主动运输一系列底物(Dean et al., 2001; Vasiliou et al., 2009; Tanaka et al., 2010).本研究中, ABCA7蛋白表达丰度上调, 表明铁离子的转运增强.对缺铁处理3天后铁吸收转运相关基因的表达水平进行测定, 发现参与还原途径的基因OsIRT1几乎不被诱导, 而参与螯合途径的OsYSL5、OsNAS1和OsNAS2基因表达被显著诱导(图5A), 地上部铁转运相关基因(OsYSL2和OsNAS1)的表达也被显著诱导(图5B), 表明铁的转运增加.此外, 参与蛋白质合成和核糖体代谢的相关蛋白表达丰度也发生变化(表1), 表明缺铁会影响水稻植株mRNA的转录及翻译过程, 但该结论有待进一步实验验证. ...
 ,*南京农业大学农学院, 南京 210095
,*南京农业大学农学院, 南京 210095
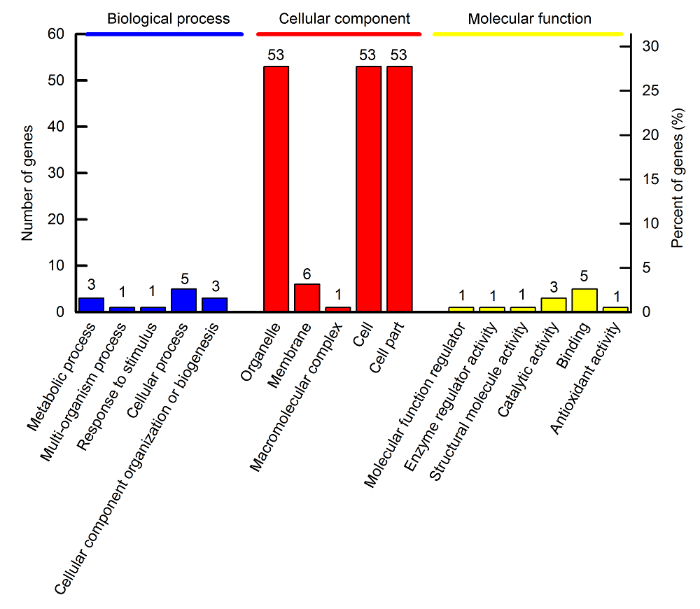 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT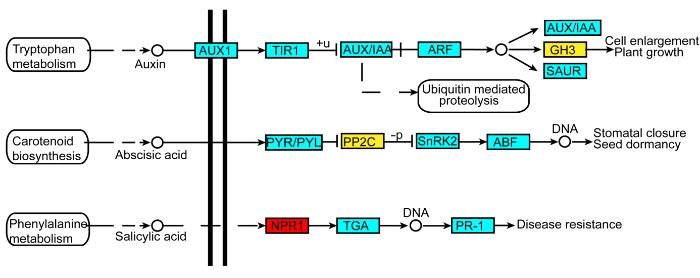 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT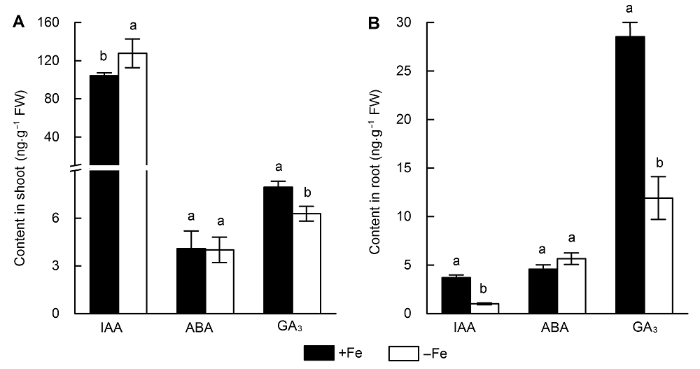 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT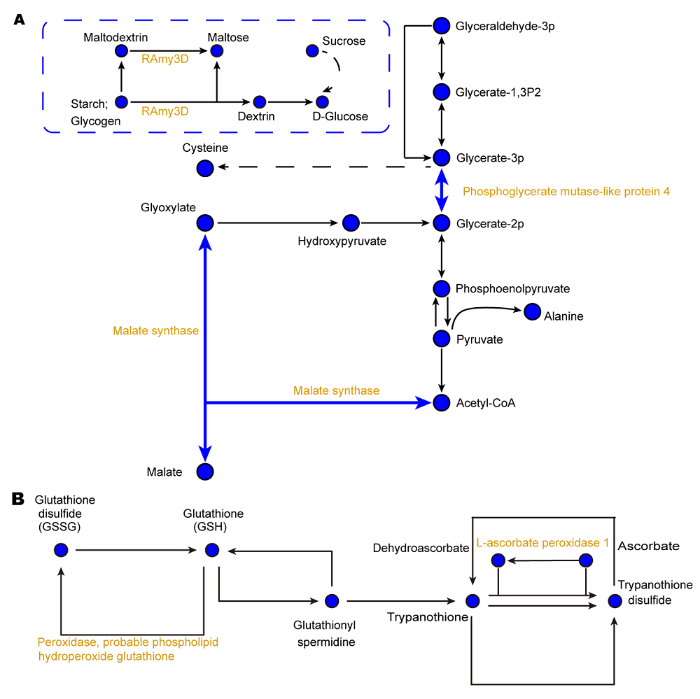 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT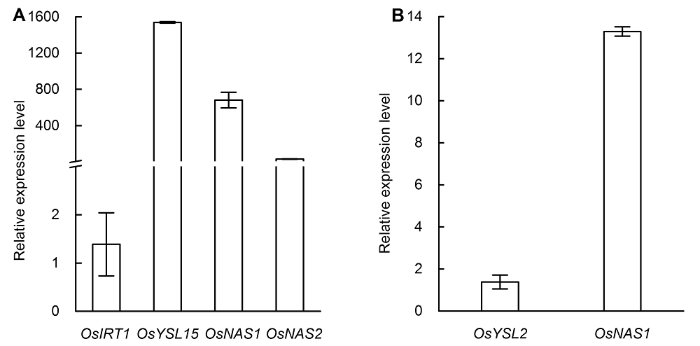 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}