李瑞雪 1 ,
李纪强 1 ,
蒲腾飞 1 ,
张晓丽 1 , 2 , 3 ,
赵喜亭 1 , 2 , 3 ,
李俊华 1 , 2 , 3 ,
李明军 1 , 2 , 3 , * , 1 河南师范大学生命科学学院, 新乡 4530072 河南省道地药材保育及利用工程技术研究中心, 新乡 453007 3 绿色药材生物技术河南省工程实验室, 新乡 453007Li Ruixue 1 ,
Li Jiqiang 1 ,
Pu Tengfei 1 ,
Zhang Xiaoli 1 , 2 , 3 ,
Zhao Xiting 1 , 2 , 3 ,
Li Junhua 1 , 2 , 3 ,
Li Mingjun 1 , 2 , 3 , * , 1 College of Life Sciences, Henan Normal University, Xinxiang 453007, China2 Engineering Technology Research Center of Nursing and Utilization of Genuine Chinese Crude Drugs in Henan Province, Xinxiang 453007, China3 Henan Province Engineering Laboratory of Green Medicinal Plant Biotechnology, Xinxiang 453007, China引用本文 李瑞雪 ,
李纪强 ,
蒲腾飞 ,
张晓丽 ,
赵喜亭 ,
李俊华 ,
李明军 . 怀山药类原球茎的诱导形成与植株再生. 植物学报, 2018, 53(3): 334-340
贡献者
* 通讯作者。E-mail: limingjun2002@263.net 基金资助
国家自然科学基金(No.81274019)、国家中医药管理局中医药行业科研专项子课题(No.201407005-08)和河南省创新型科技人才队伍建设工程(No.C20130037) ;
接受日期:2017-11-6接受日期:2018-01-12网络出版日期:2018-05-20
-->Copyright
2018《植物学报》编辑部
Contributors
* Author for correspondence. E-mail: limingjun2002@263.net History
Received :
Accepted :
Online :
摘要: 为提高山药离体繁殖的速度, 缩短繁殖周期, 以铁棍山药(Dioscorea opposita cv. ‘Tiegun’)带腋芽茎段为材料, 对类原球茎的诱导、增殖、分化与植株再生进行了研究。结果表明, 铁棍山药类原球茎诱导的最适培养基为MS+1.0 mg·L-1 TDZ+30 g·L-1 蔗糖, 增殖的最适培养基为MS+9 mg·L-1 6-BA+30 g·L-1 蔗糖, 分化的最适培养基为MS+2 mg·L-1 KT+0.02 mg·L-1 NAA+30 g·L-1 蔗糖, 最适生根培养基为1/4MS+0.05 mg·L-1 NAA+1.0 mg·L-1 PP333+15 g·L-1 蔗糖, 生根率达80%, 移栽成活率可达85%。类原球茎的诱导形成及植株再生体系的建立为怀山药种苗的快速繁殖提供了一条新途径。 关键词 :
铁棍山药 ;
诱导 ;
植株再生 ;
增殖 ;
类原球茎 Abstract :
To improve the speed of in vitro propagation of Dioscorea opposita and shorten the breeding cycle, we used nodal segments with axillary buds of D. opposita cv. ‘Tiegun’ as the starting material. The optimal conditions for protocorm-like body (PLB) induction, proliferation, differentiation and plantlet regeneration were analyzed. The optimal culture medium for PLB induction was MS+1.0 mg·L-1 TDZ+30 g·L-1 sucrose, the optimal culture medium for PLB proliferation was MS+9 mg·L-1 6-BA+30 g·L-1 sucrose, and the optimal culture medium for PLB differentiation was MS+2 mg·L-1 KT+0.02 mg·L-1 NAA+30 g·L-1 sucrose. In culture medium containing 1/4MS+0.05 mg·L-1 NAA+1.0 mg·L-1 PP333+15 g·L-1 sucrose, the rooting rate of regenerated plantlets from PLBs was 80%, and the survival rate of regenerated plantlets was up to 85% after being sown in soil. The induction of protocorm like bodies and the establishment of plant regeneration system provide a new way for rapid propagation of D. opposita seedlings. Key words :
Dioscorea opposita ;
induction ;
plant regeneration ;
proliferation ;
protocorm-like body 铁棍山药(
Dioscorea opposita cv. ‘Tiegun’)为薯蓣科薯蓣属多年生草本植物, 主产于河南省焦作市温县、武陟等地, 是我国著名的“四大怀药”之一。因其形似铁棍, 运输过程不易折断, 耐储藏, 故又名“铁棍怀山药”(
韩晓勇等, 2013 )。铁棍山药富含氨基酸、维生素、蛋白质和矿物质等(
李青, 2016 ), 是怀山药中的“极品”(
刘雯, 2014 )。但目前铁棍山药离体繁殖速度慢、周期长, 不利于其脱毒种苗的快速繁殖与推广应用, 故有必要探索一条新的山药繁殖途径。
类原球茎(protocorm-like body, PLB)最早于1960年由Morel提出(
姜福星等, 2012 ), 用来描述兰科植物组培过程中产生的类似种子萌发的过渡形态。由于植物类原球茎具有高效的分化再生能力, 并综合了胚性细胞和茎尖分生组织的优点以及其自身的独特性, 因此可广泛应用于植株再生、种质资源保存、基因工程育种、药用次生代谢产物的生产、人工种子的制备以及植物发育生物学研究等领域(
姜福星等, 2012 )。目前, 类原球茎的诱导大多运用于兰科植物, 但随着生物技术的不断发展, 半夏(
Pinellia ternata ) (
江艳华等, 2013 )和狗蔷薇(
Rosa canina ) (
毕玲等, 2012 )等植物也可诱导出类原球茎。研究表明, 不同植物可通过不同的外植体实现类原球茎的诱导, 如茎尖(
Tian et al., 2008 )、根(
罗剑飘等, 2015 )、茎段(
林江波等, 2016 )、根尖(
詹忠根等, 2005 )、种子(
古碧珠等, 2008 )和叶片(
乔永旭等, 2010 )等均可作为诱导类原球茎的材料, 其中叶片应用最为广泛。
薯蓣属植物类原球茎的研究在大薯(
D. alata ) (
许云等, 2014 )与盾叶薯蓣(
D. zingiberensis )中(
彭晓英等, 2010 )已有报道。本实验室从20世纪90年代开始即对怀山药(
D. opposita )的组织培养及其应用进行了系统研究(
李明军, 2004 ), 并建立了怀山药脱毒快繁技术体系(
李明军等, 2003 ,
2004b ;
张晓丽等, 2013 )、种质资源离体保存技术体系(
洪森荣和李明军, 2006 ;
Li et al., 2009 ;
李海兵等, 2010 ;
赵喜亭等, 2012 )和微型块茎诱导形成技术体系(
李明军等, 2000a ,
2004a ,
2008 ;
Li et al., 2014 )。2007年, 我们在研究中首次发现铁棍山药在特殊的诱导条件下也可形成类原球茎, 并对类原球茎的形成规律(
李纪强, 2013 )、相关基因(
SERK )的克隆及遗传转化(
韩林林等, 2016 )等进行了系统研究。本研究通过建立类原球茎的诱导形成与植株再生技术体系, 为怀山药优良品种和脱毒种苗的快繁及工厂化生产提供了一条新途径。
1 植物材料以河南省焦作市温县农业科学研究所种质资源圃的铁棍山药(
Dioscorea opposita Thunb. cv. ‘Tiegun’)大田苗为实验材料。
2 培养基成分与培养条件2.1 无菌苗的获得从大田取回的铁棍山药茎段, 取其顶芽, 在自来水下冲洗30分钟, 转移至超净工作台, 用75%乙醇灭菌30秒, 然后用0.1%升汞浸泡10分钟, 再用无菌水冲洗5次。在无菌条件下, 将材料接种到MS+2 mg·L
-1 KT+0.02 mg·L
-1 NAA+30 g·L
-1 蔗糖的固体培养基上。接种30天后, 将固体培养基换为液体培养基(培养基配方同上), 继续培养, 从而获得无菌试管苗。培养条件均为: 光照强度180 μmol·m
-2 ·s
-1 , 光照时间每天14小时, 温度(25±2)°C。
2.2 类原球茎的诱导形成取1-2 cm试管苗带腋芽茎段, 接种于添加不同浓度(0.1、0.5、1.0、1.5和2.0 mg·L
-1 ) TDZ与30 g·L
-1 蔗糖的MS固体培养基上。接种45天时观察类原球茎的诱导形成情况, 并统计诱导率和平均类原球茎数。
类原球茎诱导率=诱导出类原球茎的外植体数/外植体总数;
平均类原球茎数=诱导出类原球茎的总数/外植体总数。
2.3 类原球茎的增殖将生长良好的类原球茎切成长约0.5 cm的小段, 接种于类原球茎增殖培养基上, 培养基为添加了不同浓度(5、7、9、12和15 mg·L
-1 ) 6-BA与30 g·L
-1 蔗糖的MS培养基。接种30天时观察类原球茎的增殖生长情况并统计增殖率和分化率。
增殖系数=(增殖后团块PLBs数总和-增殖前团块PLBs数总和)/增殖团块数;
分化率=(分化的增殖团块数总和/增殖团块数总和)×100% (下同)。
2.4 类原球茎的分化将增殖后的类原球茎分别接种到MS+30 g·L
-1 蔗糖固体培养基、MS+30 g·L
-1 蔗糖液体培养基以及MS+2 mg·L
-1 KT+0.02 mg·L
-1 NAA+30 g·L
-1 蔗糖液体培养基上, 观察并记录每种培养基上类原球茎的分化率及平均株高。
2.5 类原球茎分化苗的生根与移栽将类原球茎分化苗接种到1/4MS+0.05 mg·L
-1 NAA+ 1.0 mg·L
-1 PP333+15 g·L
-1 蔗糖培养基上, 观察其生根情况, 45天时统计生根率。
生根率=(生根的外植体数/接种外植体数)×100%。
类原球茎分化苗在生根培养基上培养42天后, 打开封口膜移至自然光下, 3-5天后, 将幼苗根部的培养基在自来水下冲洗干净, 移栽到装有蛭石的营养钵中, 在温室中培养, 用营养液浇灌, 观察其生长状况, 待其生长健壮后移栽至大田中, 30天时统计移栽成活率。
2.6 数据统计及处理应用Excel和SPSS 13.0软件对统计结果进行分析。采用多重比较LSD法进行差异显著性分析, 不同小写字母表示在
P <0.05水平上差异显著。
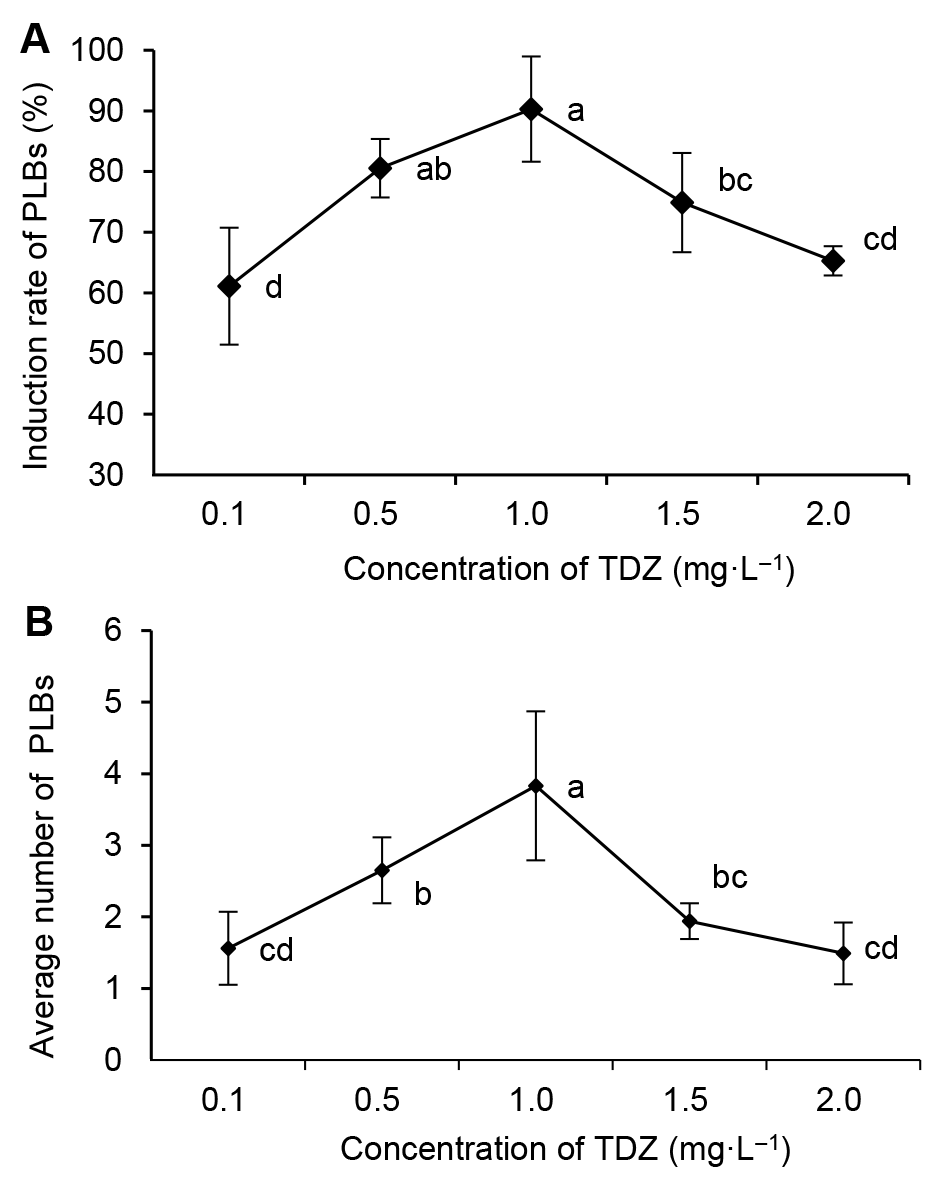
3 结果与讨论3.1 类原球茎的诱导形成将铁棍山药带腋芽茎段接种到添加不同浓度TDZ的培养基上(
图1A ), 5天时部分茎段明显膨大, 7天时有乳白色的点状圆形球体出现。随着培养天数的增加, 球体逐渐增大, 且分裂出新的球体。接种45天, 不同浓度TDZ对铁棍山药类原球茎诱导形成的影响见
图2A 和B。随着TDZ浓度的增加, 类原球茎的诱导率(
图2A )与平均类原球茎数(
图2B )呈现先升高后降低的趋势。当TDZ浓度为1.0 mg·L
-1 时, 类原球茎诱导率与平均类原球茎数均最高, 分别为90.28%和3.83个, 且类原球茎生长最好, 形态饱满, 颜色淡绿, 个头较大, 无分化现象(
图1B ); 该浓度下的诱导率与0.5 mg·L
-1 TDZ处理的差异不显著, 但与其它各浓度的诱导率差异均达显著水平; 平均类原球茎数与各浓度TDZ处理的均差异显著。当TDZ浓度为0.1与0.5 mg·L
-1 时, 分化出的类原球茎较少, 颜色浅绿, 个头较小, 且分化现象较严重。而TDZ浓度为1.5与2.0 mg·L
-1 时, 类原球茎个头虽大, 但数量较少, 且为白色, 甚至有畸形现象出现。故诱导类原球茎的最适TDZ浓度为1.0 mg·L
-1 。
图1 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-334/img_1.png <b>图1</b> 铁棍山药类原球茎(PLBs)的诱导及植株再生<br/>(A) 外植体; (B) 类原球茎的诱导形成; (C) 类原球茎的增殖; (D) 类原球茎的分化; (E) 类原球茎再生植株; (F) 移栽的再生植株<br/><b>Figure 1</b> Induction and regeneration of <i>Dioscorea opposita</i> cv. ‘Tiegun’ protocorm-like bodies (PLBs)<br/>(A) Explants; (B) Induction of PLBs; (C) Proliferation of PLBs; (D) Differentiation of PLBs; (E) Plants regenerated from PLBs; (F) Plants produced from PLBs after being sown in soil Figure 1 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-334/img_1.png <b>图1</b> 铁棍山药类原球茎(PLBs)的诱导及植株再生<br/>(A) 外植体; (B) 类原球茎的诱导形成; (C) 类原球茎的增殖; (D) 类原球茎的分化; (E) 类原球茎再生植株; (F) 移栽的再生植株<br/><b>Figure 1</b> Induction and regeneration of <i>Dioscorea opposita</i> cv. ‘Tiegun’ protocorm-like bodies (PLBs)<br/>(A) Explants; (B) Induction of PLBs; (C) Proliferation of PLBs; (D) Differentiation of PLBs; (E) Plants regenerated from PLBs; (F) Plants produced from PLBs after being sown in soil 在新窗口打开 下载 下载原图ZIP 生成PPT 图1
铁棍山药类原球茎(PLBs)的诱导及植株再生
(A) 外植体; (B) 类原球茎的诱导形成; (C) 类原球茎的增殖; (D) 类原球茎的分化; (E) 类原球茎再生植株; (F) 移栽的再生植株
Figure 1
Induction and regeneration of
Dioscorea opposita cv. ‘Tiegun’ protocorm-like bodies (PLBs)
(A) Explants; (B) Induction of PLBs; (C) Proliferation of PLBs; (D) Differentiation of PLBs; (E) Plants regenerated from PLBs; (F) Plants produced from PLBs after being sown in soil
图2 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-334/img_2.png <b>图2</b> 不同浓度TDZ对铁棍山药类原球茎(PLBs)诱导率(A)和诱导个数(B)的影响<br/>不同小写字母表示不同处理间差异显著(<i>P</i><0.05)。<br/><b>Figure 2</b> Influence of TDZ concentration on the induction rate (A) and the induction number (B) of <i>Dioscorea opposita</i> cv. ‘Tiegun’ protocorm-like bodies (PLBs)<br/>Different lowercase letters indicate significant differences at <i>P</i>< 0.05 among different treatments. Figure 2 https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-334/img_2.png <b>图2</b> 不同浓度TDZ对铁棍山药类原球茎(PLBs)诱导率(A)和诱导个数(B)的影响<br/>不同小写字母表示不同处理间差异显著(<i>P</i><0.05)。<br/><b>Figure 2</b> Influence of TDZ concentration on the induction rate (A) and the induction number (B) of <i>Dioscorea opposita</i> cv. ‘Tiegun’ protocorm-like bodies (PLBs)<br/>Different lowercase letters indicate significant differences at <i>P</i>< 0.05 among different treatments. 在新窗口打开 下载 下载原图ZIP 生成PPT 图2
不同浓度TDZ对铁棍山药类原球茎(PLBs)诱导率(A)和诱导个数(B)的影响
不同小写字母表示不同处理间差异显著(
P <0.05)。
Figure 2
Influence of TDZ concentration on the induction rate (A) and the induction number (B) of
Dioscorea opposita cv. ‘Tiegun’ protocorm-like bodies (PLBs)
Different lowercase letters indicate significant differences at
P < 0.05 among different treatments.
3.2 类原球茎的增殖不同浓度6-BA对类原球茎增殖的影响见
表1 。随着6-BA浓度的增加, 类原球茎的增殖系数呈先增高后降低的趋势, 但各处理间差异不显著。当6-BA浓度为9 mg·L
-1 时, 增殖系数最高, 分化率相对较低, 分别为5.42与8.33; 当6-BA浓度为5和7 mg·L
-1 时, 类原球茎分化率太高, 且后期类原球茎分化出芽, 无增殖现象; 当6-BA浓度为12 mg·L
-1 时, 增殖系数相比9 mg·L
-1 6-BA处理时低, 而15 mg·L
-1 6-BA处理时, 增殖 类原球茎体积小, 且增殖系数最低。故6-BA浓度为9 mg·L
-1 时最适合类原球茎的增殖(
图1C )。
表1 Table 1 表1 表1 不同浓度6-BA对铁棍山药类原球茎(PLBs)增殖的影响
Table 1 Influence of 6-BA concentration on the multiplication of
Dioscorea opposita cv. ‘Tiegun' protocorm-like bodies (PLBs)
Concentration of 6-BA-1 ) Multiplication index Differentiation The growth of PLBs 5 4.50±1.15 a 50.00 There was no PLB proliferation during the later stage. 2-3 buds were differentiated from the PLBs 7 4.67±0.95 a 33.33 There was no PLB proliferation during the later stage. 4-5 buds were differentiated from the PLBs 9 5.42±1.61 a 8.33 Light green PLBs with larger size and bud tips differentiated 12 4.92±0.14 a 8.33 Pale green PLBs with larger size and significantly differentiated bud tips 15 4.17±1.13 a 8.33 Green PLBs with smaller size and no differentiation
同列数字后相同小写字母表示不同处理间差异不显著。 The same lowercase letters behind the data in the same column indicate no significant difference among different treatments. 表1
不同浓度6-BA对铁棍山药类原球茎(PLBs)增殖的影响
Table 1
Influence of 6-BA concentration on the multiplication of
Dioscorea opposita cv. ‘Tiegun' protocorm-like bodies (PLBs)
3.3 类原球茎的分化不同培养基对类原球茎分化成苗的影响见
表2 。由
表2 可知, 3种不同的培养方式均可使类原球茎的分化率达到100%, 分化后植株的株高差异显著, 配方为MS+2 mg·L
-1 KT+0.02 mg·L
-1 NAA+30 g·L
-1 蔗糖的培养基类原球茎分化出的植株株高最高, 且植株的生长状况最好(
图1D )。故培养基MS+2 mg·L
-1 KT+0.02 mg·L
-1 NAA+30 g·L
-1 蔗糖最适合铁棍山药类原球茎的分化。
表2 Table 2 表2 表2 不同培养基对铁棍山药类原球茎(PLBs)分化成苗的影响(平均值±标准误)
Table 2 Influence of culture mediums on the differentiation of
Dioscorea opposita cv. ‘Tiegun’ protocorm-like bodies (PLBs) (means±SE)
Treatments Differentiation rate (%) Average plant height (cm) The growth of plant 1 100 3.1±0.43 c Plants with fewer branches and slightly unfolded leaves 2 100 1.4±0.54 b Plants with more branches, slightly unfolded leaves and horizontal stems 3 100 5.0±1.13 a Plants with the most branches, slightly unfolded leaves and vertical stems
1: MS+30 g·L-1 蔗糖固体培养基; 2: MS+30 g·L-1 蔗糖液体培养基; 3: MS+2 mg·L-1 KT+0.02 mg·L-1 NAA+30 g·L-1 蔗糖液体培养基。同列数字后不同小写字母表示不同处理间差异显著(P <0.05)。 1: Solid MS+30 g·L-1 sucrose medium; 2: Liquid MS+30 g·L-1 sucrose medium; 3: Liquid MS+2 mg·L-1 KT+0.02 mg·L-1 NAA+30 g·L-1 sucrose medium. Different lowercase letters behind the data in the same column indicate significant differences at P <0.05 among different treatments. 表2
不同培养基对铁棍山药类原球茎(PLBs)分化成苗的影响(平均值±标准误)
Table 2
Influence of culture mediums on the differentiation of
Dioscorea opposita cv. ‘Tiegun’ protocorm-like bodies (PLBs) (means±SE)
3.4 类原球茎分化苗的生根与炼苗移栽将类原球茎分化苗接种到生根培养基上, 9天时株高不再明显增加, 且开始出现气生根。45天, 统计得到类原球茎分化苗的生根率高达80%, 且根的生长状况良好(
图1E )。
将炼苗后的类原球茎再生植株移栽至盛有蛭石的营养钵中, 30天其成活率可达85% (
图1F )。
3.5 讨论目前, 已有很多关于山药组织培养的研究报道。李明军研究团队在怀山药组织培养方面进行了一系列探索。例如, 以茎段为材料, 通过多芽体等途径实现了怀山药无菌苗的快繁(
李明军等, 1997b ,
1999 ), 并在此基础上对铁棍山药快繁培养基进行了优化(
张晓丽等, 2013 ); 另外, 他们还发现山药可以通过诱导形成愈伤组织(
于倩和李明军, 2004 )或微型块茎(
李明军等, 2000a ), 进而形成再生植株; 并对诱导愈伤组织(
李明军等, 2000b )及微型块茎的影响因子(
李明军等, 2008 )进行了研究。
张振霞等(2016) 以广山药(
D. opposita )块茎为外植体, 进行了组织培养技术研究。
马林等(2004) 以带腋芽茎段为外植体, 通过诱导丛生芽实现了黄山药(
D. panthaica )的快速繁殖。而本研究以铁棍山药带腋芽茎段为材料, 首次成功诱导出类原球茎, 并在此基础上建立了类原球茎分化与植株再生体系, 使山药离体培养技术体系得到了进一步完善, 对加快山药良种选育或脱毒种苗的推广应用及产业化进程具有重要意义。
植物类原球茎诱导的影响因素有很多, 其中植物激素种类及其配比是最重要的因素。2,4-D是霍山石斛(
Dendrobium huoshanense )类原球茎诱导过程中的主要影响因素, 而诱导蝴蝶兰(
Phalaenopsis aphrodite ) (
崔广荣等, 2007 )、石斛兰(
D. nobile ) (
王丽萍和梁淑云, 2010 )及文心兰(
Oncidium hybridum ) (
陈晓旋等, 2010 )等兰花类原球茎主要利用细胞分裂素(6-BA或KT)与生长素(2,4-D)的组合(
姜福星等, 2012 )。在大薯诱导类原球茎的研究中发现, 适宜浓度的6-BA与NAA组合可诱导类原球茎的形成(
许云等, 2014 ); 适宜浓度的6-BA与KT组合可诱导盾叶薯蓣的形成(
彭晓英等, 2010 )。本实验以铁棍山药带腋芽茎段为材料, 探讨了不同浓度TDZ对铁棍山药类原球茎诱导形成的影响, 发现适宜浓度(1.0 mg·L
-1 )的TDZ有利于铁棍山药类原球茎的形成, 而较低浓度的TDZ则有利于芽的诱导, 较高浓度的TDZ虽然也能诱导类原球茎的形成, 但类原球茎为白色, 并且可能会导致畸形类原球茎的产生, 这与
张玉等(2002) 发现TDZ比6-BA更有利于蝴蝶兰PLBs诱导的结果一致。可见, 不同植物诱导类原球茎所需的植物激素种类及组合存在差异, 在诱导植物类原球茎过程中, 要全面考虑不同植物激素在其中的作用效果, 同时不能忽视植物激素间的协同效应(
崔广荣等, 2009 )。
类原球茎增殖过程中, 主要利用植物生长调节剂进行处理。6-BA可以促进类原球茎分生区细胞的分裂和生长, 是诱导类原球茎增殖的关键因素之一, 也是应用最广泛的细胞分裂素。
刘晓芳等(2009) 在春兰(
Cymbidium goeringii )类原球茎增殖的研究中发现, 6-BA与NAA的影响最大, 且达极显著水平;
崔广荣等(2009) 在诱导文心兰类原球茎增殖过程中也有类似发现; 而药用植物金线莲(
Anoectochilus formosanus )用来进行PLBs增殖的最适植物生长调节剂为6- BA与NAA (
陈钢等, 1994 )。本实验在增殖培养基中单独添加6-BA, 实现了铁棍山药类原球茎的增殖, 而
张超等(2010) 也在文心兰类原球茎增殖中发现, 单独使用6-BA的增殖效果较与其它植物生长调节剂配合使用的效果更好。此外, 我们在研究中还发现, 较低浓度的6-BA在培养后期会引起类原球茎的分化, 而较高浓度6-BA处理增殖后的类原球茎体积较小, 整齐度较低, 这与
杨美纯等(2000) 以及
罗剑飘等(2015) 的研究结果一致。
本实验探讨了不同培养方式对铁棍山药类原球茎分化的影响, 发现不添加任何植物激素的固体培养比液体培养更适合类原球茎的分化, 推测其原因可能为类原球茎完全浸没在液体培养基中氧气不足, 从而影响其分化。但也有研究表明, 固体培养褐化严重, 而液体培养能稀释培养过程中培养物所产生的醌类物质, 从而可以降低褐化物对其分化的抑制作用(
蔡建荣, 2008 )。本研究也表明, 添加了KT和NAA的液体培养基中铁棍山药类原球茎分化的效果最好, 说明适宜浓度的植物生长调节剂可以促进铁棍山药类原球茎在液体中的分化。
在植物组织培养过程中, 植物的生根情况会直接影响植物的成活率。影响植物生根的因素有很多, 如环境、品种差异以及植物生长调节剂的使用等。其中, 植物生长调节剂的种类和浓度起着至关重要的作用。常用的植物生根调节剂有PP333、NAA、IAA和IBA。据报道, NAA为最经济高效的生根激素, 多数花卉NAA浓度为0.1-1.0 mg·L
-1 时培养15天左右就能生根(
张瑞姿, 2012 )。PP333又称多效唑, 具有延缓植物生长、抑制茎枝伸长、促进分蘖、促进成花和坐果、增强抗旱及抗寒性、提高耐盐性和延缓植物衰老等多种效用(
李明军, 1995 )。
李明军等(1997a) 研究发现, 适宜浓度的PP333可有效促进玉米(
Zea mays )再生苗根系的生长, 并使玉米再生苗的移栽成活率由原来的不到30%提高到85%以上。本实验使用0.05 mg·L
-1 NAA和1.0 mg·L
-1 PP333达到了促进铁棍山药类原球茎分化苗生根的目的。
综上, 本研究成功建立了铁棍山药类原球茎诱导形成与植株再生技术体系, 为怀山药优良品种或脱毒试管苗的离体快繁提供了一条有效途径, 同时也为山药的遗传转化提供了一个良好的受体系统。
The authors have declared that no competing interests exist. 参考文献 文献选项 原文顺序 文献年度倒序 文中引用次数倒序 被引期刊影响因子 [1] 毕玲 , 刘凤栾 , 董爱香 , 周晓阳 , 马男 , 赵梁军 (2012 ). 狗蔷薇类原球茎遗传转化体系的建立. 核农学报 26, 270 -274 .[2] 蔡建荣 (2008 ). 山药组织培养褐化反应的研究. 中国农学通报 24(8 ), 118 -120 .[3] 陈钢 , 林兰英 , 王建勤 (1994 ). 药用金线莲组培类原球茎研究初报. 福建中医药 25(4 ), 23 -24 .[4] 陈晓旋 , 李洪波 , 叶春海 , 丰锋 , 卢淑仪 , 李柳菁 (2010 ). 文心兰类原球茎的诱导与增殖技术的优化. 热带作物学报 31, 1464 -1468 .[5] 崔广荣 , 侯喜林 , 张子学 , 张从宇 , 胡能兵 , 刘跃成 (2007 ). 蝴蝶兰叶片离体培养胚状体的发生及组织学观察. 园艺学报 34, 431 -436 .[6] 崔广荣 , 张子学 , 张从宇 , 胡能兵 (2009 ). 文心兰、蝴蝶兰类原球茎薄切片高频诱导PLBs. 安徽科技学院学报 23(2 ), 21 -27 .[7] 古碧珠 , 刘柏涛 , 何嘉碧 , 梁红 (2008 ). 石斛兰和秋花独蒜兰种子的原球茎诱导及其生长分化. 仲恺农业技术学院学报 21(4 ), 17 -21 .[8] 韩林林 , 李俊华 , 赵喜亭 , 张晓丽 , 宋志辉 , 刘世宇 , 李明军 (2016 ). 农杆菌介导的怀山药叶片瞬时表达方法的建立. 河南师范大学学报(自然科学版) 44(6 ), 135 -139 .[9] 韩晓勇 , 闫瑞霞 , 殷剑美 , 张培通 , 郭文琦 , 李春宏 (2013 ). 铁棍山药组织培养快繁及试管珠芽离体再生体系研究. 西北植物学报 33, 2120 -2125 .[10] 洪森荣 , 李明军 (2006 ). 玻璃化法超低温保存怀山药种质的技术研究. 中草药 37, 1715 -1718 .[11] 姜福星 , 刘凤栾 , 赵梁军 (2012 ). 植物类原球茎的研究与应用. 核农学报 26, 478 -483 .[12] 江艳华 , 李艾莲 , 陈彩霞 , 崔思然 , 赵锋 (2013 ). 半夏类原球茎的诱导及其植株再生研究. 中国农学通报 29(19 ), 153 -158 .[13] 李海兵 , 周娜 , 赵姣 , 李翔 , 冯秋妍 , 赵喜亭 , 李明军 (2010 ). 怀山药种质资源的包埋玻璃化超低温保存与植株再生. 植物学报 45, 379 -383 .[14] 李纪强 (2013 ). 铁棍山药类原球茎诱导增殖及植株再生的研究. 硕士论文. 新乡: 河南师范大学 . pp. 1 -43 .[15] 李明军 (1995 ). 多效唑——一种优良的植物生长调节剂. 植物学通报 12(2 ), 27 -31 .[16] 李明军 (2004 ). 怀山药组织培养及其应用 . 北京: 科学出版社. pp. 5 -160 .[17] 李明军 , 陈明霞 , 郭君丽 , 洪森荣 , 张晓丽 , 徐鑫 , 刘萍 (2004 a). 生长调节物质和糖对怀山药微型块茎诱导形成的影响. 华北农学报 19, 69 -72 .[18] 李明军 , 李金亭 , 朱命炜 , 张嘉宝 (1999 ). 怀山药的离体繁殖. 中草药 30, 296 -298 .[19] 李明军 , 刘纪华 , 张嘉宝 , 施介村 , 郭仲琛 (1997 a). 多效唑(MET)在玉米组培中的作用. 作物学报 23, 759 -761 .[20] 李明军 , 刘萍 , 张嘉宝 (2000 a). 怀山药微型块茎的离体诱导. 植物生理学通讯 36, 41 -42 .[21] 李明军 , 刘欣英 , 李萍 , 张晓丽 , 赵喜亭 , 柳俊 , 谢从华 (2008 ). 山药微型块茎诱导形成的影响因子研究. 中草药 39, 905 -910 .[22] 李明军 , 薛建平 , 陈明霞 , 郭闽英 , 邱卫花 , 张嘉宝 (2000 b). 不同因子对山药愈伤组织诱导的影响. 广西植物 20, 156 -160 .[23] 李明军 , 杨建伟 , 张嘉宝 (1997 b). 怀山药的茎段培养和快速繁殖. 植物生理学报 33, 275 -276 .[24] 李明军 , 张宝华 , 王天亮 , 张峰 , 陈明霞 , 张嘉宝 (2004b ). 怀山药脱毒快繁技术研究. 见:陈振光. 全国植物组织培养、脱毒快繁及工厂化生产学术研讨会 . 北京: 中国农业科技出版社. pp. 92 -97 .[25] 李明军 , 张峰 , 陈明霞 , 于相丽 (2003 ). 怀山药病毒病的研究. 中草药 34, 附3-附5. [26] 李青 (2016 ). 绿色食品铁棍山药栽培技术. 河南农业 11, 31 -32 .[27] 林江波 , 王伟英 , 李海明 , 吴水金 , 邹晖 , 李跃森 , 戴艺民 (2016 ). 铁皮石斛茎段原球茎的诱导、分化与植株再生. 福建农业学报 31, 1075 -1079 .[28] 刘雯 (2014 ). 铁棍山药微型块茎形成的形态解剖及生理生化变化研究. 硕士论文. 新乡: 河南师范大学 . pp. 1 -34 .[29] 刘晓芳 , 黄闽敏 , 姜敏 , 牛俊丽 (2009 ). 离体培养春兰原球茎增殖的影响因子研究. 西北林学院学报 24(3 ), 88 -90 .[30] 罗剑飘 , 吴坤林 , 翁殊斐 (2015 ). 麒麟火焰兰叶片及根类原球茎的诱导研究. 浙江农业学报 27, 1173 -1177 .[31] 马林 , 张玲 , 李卫锋 (2004 ). 黄山药丛生芽诱导与植株快速繁殖. 生物技术 14(2 ), 53 -54 .[32] 彭晓英 , 周朴华 , 张良波 , 彭尽晖 , 周双德 (2010 ). 盾叶薯蓣类原球茎的离体诱导及快繁体系的建立. 植物遗传资源学报 11, 629 -634 .[33] 乔永旭 , 张永平 , 陈超 , 韩兴辉 , 王旭 , 刘桂萍 (2010 ). 蝴蝶兰组培苗类原球茎的诱导与分化. 北方园艺 (4 ), 138 -141 .[34] 王丽萍 , 梁淑云 (2010 ). 铁皮石斛原球茎诱导与增殖研究. 中国农学通报 26, 265 -268 .[35] 许云 , 高艳强 , 高洪昌 , 夏赟 , 吴文嫱 , 黄东益 (2014 ). 大薯类原球茎的离体诱导及再生体系的建立. 植物生理学报 50, 1027 -1032 .[36] 杨美纯 , 周歧伟 , 许鸿源 , 卢美英 (2000 ). 外部因子对蝴蝶兰叶片原球茎状体发生的影响. 广西植物 20, 42 -46 .[37] 于倩 , 李明军 (2004 ). 怀山药微型块茎愈伤组织的诱导形成及高频率再生. 生态学报 24, 1022 -1026 .[38] 詹忠根 , 徐程 , 张铭 , 罗紫娟 (2005 ). 铁皮石斛离体根尖经体细胞胚再生植株研究. 浙江大学学报(农业与生命科学版) 31, 579 -580 .[39] 张超 , 荣松 , 张茜茜 , 楼楠男 (2010 ). 基本培养基、碳源及植物生长调节剂对文心兰原球茎增殖的影响. 浙江农业科学 1, 55 -59 .[40] 张瑞姿 (2012 ). 植物生长调节剂在观赏植物组织培养中的应用. 园林绿化 (5 ), 39 -40 .[41] 张晓丽 , 王医鹏 , 刘雯 , 郭晓博 , 李书杰 , 李明军 (2013 ). 铁棍山药试管苗快繁培养基的优化. 河南师范大学学报(自然科学版) 41, 123 -126 .[42] 张玉 , 李艳敏 , 张和臣 , 王利民 , 王慧娟 , 张鸽香 (2012 ). 蝴蝶兰叶片诱导类原球茎初探. 河南农业科学 41, 126 -128 .[43] 张振霞 , 叶静鹏 , 郑莉 , 陈贵豪 , 郑玉忠 (2016 ). 广山药组织培养技术的研究. 江苏农业科学 44(2 ), 76 -80 .[44] 赵喜亭 , 王苗 , 邵换娟 , 李翔 , 李海兵 , 李明军 (2012 ). 山药种质包埋玻璃化超低温保存再生植株的稳定性分析. 华北农学报 27, 234 -238 .[45] Li MJ Li JH Liu W Liu LY Lu J Niu J Liu XY Yang QX 2014 ). A protocol for in vitro production of microtubers in Chinese yam ( Dioscorea opposita ). Biosci Biotechnol Bio- chem 78, 1005 -1009 .[46] Li MJ Zhao XT Hong SR Zhang XL Li P Liu J Xie CH 2009 ). Cryopreservation of plantlet nodes of Dioscorea opposita Thunb. using a vitrification method. CryoLetters 30, 19 -28 .[47] Tian CW Chen Y Zhao XL Zhao LJ 2008 ). Plant regene- ration through protocorm-like bodies induced from rhi- zoids using leaf explants of Rosa spp. Plant Cell Rep 823 -831 .
狗蔷薇类原球茎遗传转化体系的建立
1
2012
... 类原球茎(protocorm-like body, PLB)最早于1960年由Morel提出(
姜福星等, 2012 ), 用来描述兰科植物组培过程中产生的类似种子萌发的过渡形态.由于植物类原球茎具有高效的分化再生能力, 并综合了胚性细胞和茎尖分生组织的优点以及其自身的独特性, 因此可广泛应用于植株再生、种质资源保存、基因工程育种、药用次生代谢产物的生产、人工种子的制备以及植物发育生物学研究等领域(
姜福星等, 2012 ).目前, 类原球茎的诱导大多运用于兰科植物, 但随着生物技术的不断发展, 半夏(
Pinellia ternata ) (
江艳华等, 2013 )和狗蔷薇(
Rosa canina ) (
毕玲等, 2012 )等植物也可诱导出类原球茎.研究表明, 不同植物可通过不同的外植体实现类原球茎的诱导, 如茎尖(
Tian et al., 2008 )、根(
罗剑飘等, 2015 )、茎段(
林江波等, 2016 )、根尖(
詹忠根等, 2005 )、种子(
古碧珠等, 2008 )和叶片(
乔永旭等, 2010 )等均可作为诱导类原球茎的材料, 其中叶片应用最为广泛. ...
山药组织培养褐化反应的研究
1
2008
... 本实验探讨了不同培养方式对铁棍山药类原球茎分化的影响, 发现不添加任何植物激素的固体培养比液体培养更适合类原球茎的分化, 推测其原因可能为类原球茎完全浸没在液体培养基中氧气不足, 从而影响其分化.但也有研究表明, 固体培养褐化严重, 而液体培养能稀释培养过程中培养物所产生的醌类物质, 从而可以降低褐化物对其分化的抑制作用(
蔡建荣, 2008 ).本研究也表明, 添加了KT和NAA的液体培养基中铁棍山药类原球茎分化的效果最好, 说明适宜浓度的植物生长调节剂可以促进铁棍山药类原球茎在液体中的分化. ...
药用金线莲组培类原球茎研究初报
1
1994
... 类原球茎增殖过程中, 主要利用植物生长调节剂进行处理.6-BA可以促进类原球茎分生区细胞的分裂和生长, 是诱导类原球茎增殖的关键因素之一, 也是应用最广泛的细胞分裂素.
刘晓芳等(2009) 在春兰(
Cymbidium goeringii )类原球茎增殖的研究中发现, 6-BA与NAA的影响最大, 且达极显著水平;
崔广荣等(2009) 在诱导文心兰类原球茎增殖过程中也有类似发现; 而药用植物金线莲(
Anoectochilus formosanus )用来进行PLBs增殖的最适植物生长调节剂为6- BA与NAA (
陈钢等, 1994 ).本实验在增殖培养基中单独添加6-BA, 实现了铁棍山药类原球茎的增殖, 而
张超等(2010) 也在文心兰类原球茎增殖中发现, 单独使用6-BA的增殖效果较与其它植物生长调节剂配合使用的效果更好.此外, 我们在研究中还发现, 较低浓度的6-BA在培养后期会引起类原球茎的分化, 而较高浓度6-BA处理增殖后的类原球茎体积较小, 整齐度较低, 这与
杨美纯等(2000) 以及
罗剑飘等(2015) 的研究结果一致. ...
文心兰类原球茎的诱导与增殖技术的优化
1
2010
... 植物类原球茎诱导的影响因素有很多, 其中植物激素种类及其配比是最重要的因素.2,4-D是霍山石斛(
Dendrobium huoshanense )类原球茎诱导过程中的主要影响因素, 而诱导蝴蝶兰(
Phalaenopsis aphrodite ) (
崔广荣等, 2007 )、石斛兰(
D. nobile ) (
王丽萍和梁淑云, 2010 )及文心兰(
Oncidium hybridum ) (
陈晓旋等, 2010 )等兰花类原球茎主要利用细胞分裂素(6-BA或KT)与生长素(2,4-D)的组合(
姜福星等, 2012 ).在大薯诱导类原球茎的研究中发现, 适宜浓度的6-BA与NAA组合可诱导类原球茎的形成(
许云等, 2014 ); 适宜浓度的6-BA与KT组合可诱导盾叶薯蓣的形成(
彭晓英等, 2010 ).本实验以铁棍山药带腋芽茎段为材料, 探讨了不同浓度TDZ对铁棍山药类原球茎诱导形成的影响, 发现适宜浓度(1.0 mg·L
-1 )的TDZ有利于铁棍山药类原球茎的形成, 而较低浓度的TDZ则有利于芽的诱导, 较高浓度的TDZ虽然也能诱导类原球茎的形成, 但类原球茎为白色, 并且可能会导致畸形类原球茎的产生, 这与
张玉等(2002) 发现TDZ比6-BA更有利于蝴蝶兰PLBs诱导的结果一致.可见, 不同植物诱导类原球茎所需的植物激素种类及组合存在差异, 在诱导植物类原球茎过程中, 要全面考虑不同植物激素在其中的作用效果, 同时不能忽视植物激素间的协同效应(
崔广荣等, 2009 ). ...
蝴蝶兰叶片离体培养胚状体的发生及组织学观察
1
2007
... 植物类原球茎诱导的影响因素有很多, 其中植物激素种类及其配比是最重要的因素.2,4-D是霍山石斛(
Dendrobium huoshanense )类原球茎诱导过程中的主要影响因素, 而诱导蝴蝶兰(
Phalaenopsis aphrodite ) (
崔广荣等, 2007 )、石斛兰(
D. nobile ) (
王丽萍和梁淑云, 2010 )及文心兰(
Oncidium hybridum ) (
陈晓旋等, 2010 )等兰花类原球茎主要利用细胞分裂素(6-BA或KT)与生长素(2,4-D)的组合(
姜福星等, 2012 ).在大薯诱导类原球茎的研究中发现, 适宜浓度的6-BA与NAA组合可诱导类原球茎的形成(
许云等, 2014 ); 适宜浓度的6-BA与KT组合可诱导盾叶薯蓣的形成(
彭晓英等, 2010 ).本实验以铁棍山药带腋芽茎段为材料, 探讨了不同浓度TDZ对铁棍山药类原球茎诱导形成的影响, 发现适宜浓度(1.0 mg·L
-1 )的TDZ有利于铁棍山药类原球茎的形成, 而较低浓度的TDZ则有利于芽的诱导, 较高浓度的TDZ虽然也能诱导类原球茎的形成, 但类原球茎为白色, 并且可能会导致畸形类原球茎的产生, 这与
张玉等(2002) 发现TDZ比6-BA更有利于蝴蝶兰PLBs诱导的结果一致.可见, 不同植物诱导类原球茎所需的植物激素种类及组合存在差异, 在诱导植物类原球茎过程中, 要全面考虑不同植物激素在其中的作用效果, 同时不能忽视植物激素间的协同效应(
崔广荣等, 2009 ). ...
文心兰、蝴蝶兰类原球茎薄切片高频诱导PLBs
2
2009
... 植物类原球茎诱导的影响因素有很多, 其中植物激素种类及其配比是最重要的因素.2,4-D是霍山石斛(
Dendrobium huoshanense )类原球茎诱导过程中的主要影响因素, 而诱导蝴蝶兰(
Phalaenopsis aphrodite ) (
崔广荣等, 2007 )、石斛兰(
D. nobile ) (
王丽萍和梁淑云, 2010 )及文心兰(
Oncidium hybridum ) (
陈晓旋等, 2010 )等兰花类原球茎主要利用细胞分裂素(6-BA或KT)与生长素(2,4-D)的组合(
姜福星等, 2012 ).在大薯诱导类原球茎的研究中发现, 适宜浓度的6-BA与NAA组合可诱导类原球茎的形成(
许云等, 2014 ); 适宜浓度的6-BA与KT组合可诱导盾叶薯蓣的形成(
彭晓英等, 2010 ).本实验以铁棍山药带腋芽茎段为材料, 探讨了不同浓度TDZ对铁棍山药类原球茎诱导形成的影响, 发现适宜浓度(1.0 mg·L
-1 )的TDZ有利于铁棍山药类原球茎的形成, 而较低浓度的TDZ则有利于芽的诱导, 较高浓度的TDZ虽然也能诱导类原球茎的形成, 但类原球茎为白色, 并且可能会导致畸形类原球茎的产生, 这与
张玉等(2002) 发现TDZ比6-BA更有利于蝴蝶兰PLBs诱导的结果一致.可见, 不同植物诱导类原球茎所需的植物激素种类及组合存在差异, 在诱导植物类原球茎过程中, 要全面考虑不同植物激素在其中的作用效果, 同时不能忽视植物激素间的协同效应(
崔广荣等, 2009 ). ...
... 类原球茎增殖过程中, 主要利用植物生长调节剂进行处理.6-BA可以促进类原球茎分生区细胞的分裂和生长, 是诱导类原球茎增殖的关键因素之一, 也是应用最广泛的细胞分裂素.
刘晓芳等(2009) 在春兰(
Cymbidium goeringii )类原球茎增殖的研究中发现, 6-BA与NAA的影响最大, 且达极显著水平;
崔广荣等(2009) 在诱导文心兰类原球茎增殖过程中也有类似发现; 而药用植物金线莲(
Anoectochilus formosanus )用来进行PLBs增殖的最适植物生长调节剂为6- BA与NAA (
陈钢等, 1994 ).本实验在增殖培养基中单独添加6-BA, 实现了铁棍山药类原球茎的增殖, 而
张超等(2010) 也在文心兰类原球茎增殖中发现, 单独使用6-BA的增殖效果较与其它植物生长调节剂配合使用的效果更好.此外, 我们在研究中还发现, 较低浓度的6-BA在培养后期会引起类原球茎的分化, 而较高浓度6-BA处理增殖后的类原球茎体积较小, 整齐度较低, 这与
杨美纯等(2000) 以及
罗剑飘等(2015) 的研究结果一致. ...
石斛兰和秋花独蒜兰种子的原球茎诱导及其生长分化
1
2008
... 类原球茎(protocorm-like body, PLB)最早于1960年由Morel提出(
姜福星等, 2012 ), 用来描述兰科植物组培过程中产生的类似种子萌发的过渡形态.由于植物类原球茎具有高效的分化再生能力, 并综合了胚性细胞和茎尖分生组织的优点以及其自身的独特性, 因此可广泛应用于植株再生、种质资源保存、基因工程育种、药用次生代谢产物的生产、人工种子的制备以及植物发育生物学研究等领域(
姜福星等, 2012 ).目前, 类原球茎的诱导大多运用于兰科植物, 但随着生物技术的不断发展, 半夏(
Pinellia ternata ) (
江艳华等, 2013 )和狗蔷薇(
Rosa canina ) (
毕玲等, 2012 )等植物也可诱导出类原球茎.研究表明, 不同植物可通过不同的外植体实现类原球茎的诱导, 如茎尖(
Tian et al., 2008 )、根(
罗剑飘等, 2015 )、茎段(
林江波等, 2016 )、根尖(
詹忠根等, 2005 )、种子(
古碧珠等, 2008 )和叶片(
乔永旭等, 2010 )等均可作为诱导类原球茎的材料, 其中叶片应用最为广泛. ...
农杆菌介导的怀山药叶片瞬时表达方法的建立
1
2016
... 薯蓣属植物类原球茎的研究在大薯(
D. alata ) (
许云等, 2014 )与盾叶薯蓣(
D. zingiberensis )中(
彭晓英等, 2010 )已有报道.本实验室从20世纪90年代开始即对怀山药(
D. opposita )的组织培养及其应用进行了系统研究(
李明军, 2004 ), 并建立了怀山药脱毒快繁技术体系(
李明军等, 2003 ,
2004b ;
张晓丽等, 2013 )、种质资源离体保存技术体系(
洪森荣和李明军, 2006 ;
Li et al., 2009 ;
李海兵等, 2010 ;
赵喜亭等, 2012 )和微型块茎诱导形成技术体系(
李明军等, 2000a ,
2004a ,
2008 ;
Li et al., 2014 ).2007年, 我们在研究中首次发现铁棍山药在特殊的诱导条件下也可形成类原球茎, 并对类原球茎的形成规律(
李纪强, 2013 )、相关基因(
SERK )的克隆及遗传转化(
韩林林等, 2016 )等进行了系统研究.本研究通过建立类原球茎的诱导形成与植株再生技术体系, 为怀山药优良品种和脱毒种苗的快繁及工厂化生产提供了一条新途径. ...
铁棍山药组织培养快繁及试管珠芽离体再生体系研究
1
2013
... 铁棍山药(
Dioscorea opposita cv. ‘Tiegun’)为薯蓣科薯蓣属多年生草本植物, 主产于河南省焦作市温县、武陟等地, 是我国著名的“四大怀药”之一.因其形似铁棍, 运输过程不易折断, 耐储藏, 故又名“铁棍怀山药”(
韩晓勇等, 2013 ).铁棍山药富含氨基酸、维生素、蛋白质和矿物质等(
李青, 2016 ), 是怀山药中的“极品”(
刘雯, 2014 ).但目前铁棍山药离体繁殖速度慢、周期长, 不利于其脱毒种苗的快速繁殖与推广应用, 故有必要探索一条新的山药繁殖途径. ...
玻璃化法超低温保存怀山药种质的技术研究
1
2006
... 薯蓣属植物类原球茎的研究在大薯(
D. alata ) (
许云等, 2014 )与盾叶薯蓣(
D. zingiberensis )中(
彭晓英等, 2010 )已有报道.本实验室从20世纪90年代开始即对怀山药(
D. opposita )的组织培养及其应用进行了系统研究(
李明军, 2004 ), 并建立了怀山药脱毒快繁技术体系(
李明军等, 2003 ,
2004b ;
张晓丽等, 2013 )、种质资源离体保存技术体系(
洪森荣和李明军, 2006 ;
Li et al., 2009 ;
李海兵等, 2010 ;
赵喜亭等, 2012 )和微型块茎诱导形成技术体系(
李明军等, 2000a ,
2004a ,
2008 ;
Li et al., 2014 ).2007年, 我们在研究中首次发现铁棍山药在特殊的诱导条件下也可形成类原球茎, 并对类原球茎的形成规律(
李纪强, 2013 )、相关基因(
SERK )的克隆及遗传转化(
韩林林等, 2016 )等进行了系统研究.本研究通过建立类原球茎的诱导形成与植株再生技术体系, 为怀山药优良品种和脱毒种苗的快繁及工厂化生产提供了一条新途径. ...
植物类原球茎的研究与应用
3
2012
... 类原球茎(protocorm-like body, PLB)最早于1960年由Morel提出(
姜福星等, 2012 ), 用来描述兰科植物组培过程中产生的类似种子萌发的过渡形态.由于植物类原球茎具有高效的分化再生能力, 并综合了胚性细胞和茎尖分生组织的优点以及其自身的独特性, 因此可广泛应用于植株再生、种质资源保存、基因工程育种、药用次生代谢产物的生产、人工种子的制备以及植物发育生物学研究等领域(
姜福星等, 2012 ).目前, 类原球茎的诱导大多运用于兰科植物, 但随着生物技术的不断发展, 半夏(
Pinellia ternata ) (
江艳华等, 2013 )和狗蔷薇(
Rosa canina ) (
毕玲等, 2012 )等植物也可诱导出类原球茎.研究表明, 不同植物可通过不同的外植体实现类原球茎的诱导, 如茎尖(
Tian et al., 2008 )、根(
罗剑飘等, 2015 )、茎段(
林江波等, 2016 )、根尖(
詹忠根等, 2005 )、种子(
古碧珠等, 2008 )和叶片(
乔永旭等, 2010 )等均可作为诱导类原球茎的材料, 其中叶片应用最为广泛. ...
... ), 用来描述兰科植物组培过程中产生的类似种子萌发的过渡形态.由于植物类原球茎具有高效的分化再生能力, 并综合了胚性细胞和茎尖分生组织的优点以及其自身的独特性, 因此可广泛应用于植株再生、种质资源保存、基因工程育种、药用次生代谢产物的生产、人工种子的制备以及植物发育生物学研究等领域(
姜福星等, 2012 ).目前, 类原球茎的诱导大多运用于兰科植物, 但随着生物技术的不断发展, 半夏(
Pinellia ternata ) (
江艳华等, 2013 )和狗蔷薇(
Rosa canina ) (
毕玲等, 2012 )等植物也可诱导出类原球茎.研究表明, 不同植物可通过不同的外植体实现类原球茎的诱导, 如茎尖(
Tian et al., 2008 )、根(
罗剑飘等, 2015 )、茎段(
林江波等, 2016 )、根尖(
詹忠根等, 2005 )、种子(
古碧珠等, 2008 )和叶片(
乔永旭等, 2010 )等均可作为诱导类原球茎的材料, 其中叶片应用最为广泛. ...
... 植物类原球茎诱导的影响因素有很多, 其中植物激素种类及其配比是最重要的因素.2,4-D是霍山石斛(
Dendrobium huoshanense )类原球茎诱导过程中的主要影响因素, 而诱导蝴蝶兰(
Phalaenopsis aphrodite ) (
崔广荣等, 2007 )、石斛兰(
D. nobile ) (
王丽萍和梁淑云, 2010 )及文心兰(
Oncidium hybridum ) (
陈晓旋等, 2010 )等兰花类原球茎主要利用细胞分裂素(6-BA或KT)与生长素(2,4-D)的组合(
姜福星等, 2012 ).在大薯诱导类原球茎的研究中发现, 适宜浓度的6-BA与NAA组合可诱导类原球茎的形成(
许云等, 2014 ); 适宜浓度的6-BA与KT组合可诱导盾叶薯蓣的形成(
彭晓英等, 2010 ).本实验以铁棍山药带腋芽茎段为材料, 探讨了不同浓度TDZ对铁棍山药类原球茎诱导形成的影响, 发现适宜浓度(1.0 mg·L
-1 )的TDZ有利于铁棍山药类原球茎的形成, 而较低浓度的TDZ则有利于芽的诱导, 较高浓度的TDZ虽然也能诱导类原球茎的形成, 但类原球茎为白色, 并且可能会导致畸形类原球茎的产生, 这与
张玉等(2002) 发现TDZ比6-BA更有利于蝴蝶兰PLBs诱导的结果一致.可见, 不同植物诱导类原球茎所需的植物激素种类及组合存在差异, 在诱导植物类原球茎过程中, 要全面考虑不同植物激素在其中的作用效果, 同时不能忽视植物激素间的协同效应(
崔广荣等, 2009 ). ...
半夏类原球茎的诱导及其植株再生研究
1
2013
... 类原球茎(protocorm-like body, PLB)最早于1960年由Morel提出(
姜福星等, 2012 ), 用来描述兰科植物组培过程中产生的类似种子萌发的过渡形态.由于植物类原球茎具有高效的分化再生能力, 并综合了胚性细胞和茎尖分生组织的优点以及其自身的独特性, 因此可广泛应用于植株再生、种质资源保存、基因工程育种、药用次生代谢产物的生产、人工种子的制备以及植物发育生物学研究等领域(
姜福星等, 2012 ).目前, 类原球茎的诱导大多运用于兰科植物, 但随着生物技术的不断发展, 半夏(
Pinellia ternata ) (
江艳华等, 2013 )和狗蔷薇(
Rosa canina ) (
毕玲等, 2012 )等植物也可诱导出类原球茎.研究表明, 不同植物可通过不同的外植体实现类原球茎的诱导, 如茎尖(
Tian et al., 2008 )、根(
罗剑飘等, 2015 )、茎段(
林江波等, 2016 )、根尖(
詹忠根等, 2005 )、种子(
古碧珠等, 2008 )和叶片(
乔永旭等, 2010 )等均可作为诱导类原球茎的材料, 其中叶片应用最为广泛. ...
怀山药种质资源的包埋玻璃化超低温保存与植株再生
1
2010
... 薯蓣属植物类原球茎的研究在大薯(
D. alata ) (
许云等, 2014 )与盾叶薯蓣(
D. zingiberensis )中(
彭晓英等, 2010 )已有报道.本实验室从20世纪90年代开始即对怀山药(
D. opposita )的组织培养及其应用进行了系统研究(
李明军, 2004 ), 并建立了怀山药脱毒快繁技术体系(
李明军等, 2003 ,
2004b ;
张晓丽等, 2013 )、种质资源离体保存技术体系(
洪森荣和李明军, 2006 ;
Li et al., 2009 ;
李海兵等, 2010 ;
赵喜亭等, 2012 )和微型块茎诱导形成技术体系(
李明军等, 2000a ,
2004a ,
2008 ;
Li et al., 2014 ).2007年, 我们在研究中首次发现铁棍山药在特殊的诱导条件下也可形成类原球茎, 并对类原球茎的形成规律(
李纪强, 2013 )、相关基因(
SERK )的克隆及遗传转化(
韩林林等, 2016 )等进行了系统研究.本研究通过建立类原球茎的诱导形成与植株再生技术体系, 为怀山药优良品种和脱毒种苗的快繁及工厂化生产提供了一条新途径. ...
铁棍山药类原球茎诱导增殖及植株再生的研究
1
2013
... 薯蓣属植物类原球茎的研究在大薯(
D. alata ) (
许云等, 2014 )与盾叶薯蓣(
D. zingiberensis )中(
彭晓英等, 2010 )已有报道.本实验室从20世纪90年代开始即对怀山药(
D. opposita )的组织培养及其应用进行了系统研究(
李明军, 2004 ), 并建立了怀山药脱毒快繁技术体系(
李明军等, 2003 ,
2004b ;
张晓丽等, 2013 )、种质资源离体保存技术体系(
洪森荣和李明军, 2006 ;
Li et al., 2009 ;
李海兵等, 2010 ;
赵喜亭等, 2012 )和微型块茎诱导形成技术体系(
李明军等, 2000a ,
2004a ,
2008 ;
Li et al., 2014 ).2007年, 我们在研究中首次发现铁棍山药在特殊的诱导条件下也可形成类原球茎, 并对类原球茎的形成规律(
李纪强, 2013 )、相关基因(
SERK )的克隆及遗传转化(
韩林林等, 2016 )等进行了系统研究.本研究通过建立类原球茎的诱导形成与植株再生技术体系, 为怀山药优良品种和脱毒种苗的快繁及工厂化生产提供了一条新途径. ...
多效唑——一种优良的植物生长调节剂
1
1995
... 在植物组织培养过程中, 植物的生根情况会直接影响植物的成活率.影响植物生根的因素有很多, 如环境、品种差异以及植物生长调节剂的使用等.其中, 植物生长调节剂的种类和浓度起着至关重要的作用.常用的植物生根调节剂有PP333、NAA、IAA和IBA.据报道, NAA为最经济高效的生根激素, 多数花卉NAA浓度为0.1-1.0 mg·L
-1 时培养15天左右就能生根(
张瑞姿, 2012 ).PP333又称多效唑, 具有延缓植物生长、抑制茎枝伸长、促进分蘖、促进成花和坐果、增强抗旱及抗寒性、提高耐盐性和延缓植物衰老等多种效用(
李明军, 1995 ).
李明军等(1997a) 研究发现, 适宜浓度的PP333可有效促进玉米(
Zea mays )再生苗根系的生长, 并使玉米再生苗的移栽成活率由原来的不到30%提高到85%以上.本实验使用0.05 mg·L
-1 NAA和1.0 mg·L
-1 PP333达到了促进铁棍山药类原球茎分化苗生根的目的. ...
0
2004
a). 生长调节物质和糖对怀山药微型块茎诱导形成的影响
2
2004
... 薯蓣属植物类原球茎的研究在大薯(
D. alata ) (
许云等, 2014 )与盾叶薯蓣(
D. zingiberensis )中(
彭晓英等, 2010 )已有报道.本实验室从20世纪90年代开始即对怀山药(
D. opposita )的组织培养及其应用进行了系统研究(
李明军, 2004 ), 并建立了怀山药脱毒快繁技术体系(
李明军等, 2003 ,
2004b ;
张晓丽等, 2013 )、种质资源离体保存技术体系(
洪森荣和李明军, 2006 ;
Li et al., 2009 ;
李海兵等, 2010 ;
赵喜亭等, 2012 )和微型块茎诱导形成技术体系(
李明军等, 2000a ,
2004a ,
2008 ;
Li et al., 2014 ).2007年, 我们在研究中首次发现铁棍山药在特殊的诱导条件下也可形成类原球茎, 并对类原球茎的形成规律(
李纪强, 2013 )、相关基因(
SERK )的克隆及遗传转化(
韩林林等, 2016 )等进行了系统研究.本研究通过建立类原球茎的诱导形成与植株再生技术体系, 为怀山药优良品种和脱毒种苗的快繁及工厂化生产提供了一条新途径. ...
... ,
2004a ,
2008 ;
Li et al., 2014 ).2007年, 我们在研究中首次发现铁棍山药在特殊的诱导条件下也可形成类原球茎, 并对类原球茎的形成规律(
李纪强, 2013 )、相关基因(
SERK )的克隆及遗传转化(
韩林林等, 2016 )等进行了系统研究.本研究通过建立类原球茎的诱导形成与植株再生技术体系, 为怀山药优良品种和脱毒种苗的快繁及工厂化生产提供了一条新途径. ...
怀山药的离体繁殖
1
1999
... 目前, 已有很多关于山药组织培养的研究报道.李明军研究团队在怀山药组织培养方面进行了一系列探索.例如, 以茎段为材料, 通过多芽体等途径实现了怀山药无菌苗的快繁(
李明军等, 1997b ,
1999 ), 并在此基础上对铁棍山药快繁培养基进行了优化(
张晓丽等, 2013 ); 另外, 他们还发现山药可以通过诱导形成愈伤组织(
于倩和李明军, 2004 )或微型块茎(
李明军等, 2000a ), 进而形成再生植株; 并对诱导愈伤组织(
李明军等, 2000b )及微型块茎的影响因子(
李明军等, 2008 )进行了研究.
张振霞等(2016) 以广山药(
D. opposita )块茎为外植体, 进行了组织培养技术研究.
马林等(2004) 以带腋芽茎段为外植体, 通过诱导丛生芽实现了黄山药(
D. panthaica )的快速繁殖.而本研究以铁棍山药带腋芽茎段为材料, 首次成功诱导出类原球茎, 并在此基础上建立了类原球茎分化与植株再生体系, 使山药离体培养技术体系得到了进一步完善, 对加快山药良种选育或脱毒种苗的推广应用及产业化进程具有重要意义. ...
a). 多效唑(MET)在玉米组培中的作用
1
1997
... 在植物组织培养过程中, 植物的生根情况会直接影响植物的成活率.影响植物生根的因素有很多, 如环境、品种差异以及植物生长调节剂的使用等.其中, 植物生长调节剂的种类和浓度起着至关重要的作用.常用的植物生根调节剂有PP333、NAA、IAA和IBA.据报道, NAA为最经济高效的生根激素, 多数花卉NAA浓度为0.1-1.0 mg·L
-1 时培养15天左右就能生根(
张瑞姿, 2012 ).PP333又称多效唑, 具有延缓植物生长、抑制茎枝伸长、促进分蘖、促进成花和坐果、增强抗旱及抗寒性、提高耐盐性和延缓植物衰老等多种效用(
李明军, 1995 ).
李明军等(1997a) 研究发现, 适宜浓度的PP333可有效促进玉米(
Zea mays )再生苗根系的生长, 并使玉米再生苗的移栽成活率由原来的不到30%提高到85%以上.本实验使用0.05 mg·L
-1 NAA和1.0 mg·L
-1 PP333达到了促进铁棍山药类原球茎分化苗生根的目的. ...
a). 怀山药微型块茎的离体诱导
2
2000
... 薯蓣属植物类原球茎的研究在大薯(
D. alata ) (
许云等, 2014 )与盾叶薯蓣(
D. zingiberensis )中(
彭晓英等, 2010 )已有报道.本实验室从20世纪90年代开始即对怀山药(
D. opposita )的组织培养及其应用进行了系统研究(
李明军, 2004 ), 并建立了怀山药脱毒快繁技术体系(
李明军等, 2003 ,
2004b ;
张晓丽等, 2013 )、种质资源离体保存技术体系(
洪森荣和李明军, 2006 ;
Li et al., 2009 ;
李海兵等, 2010 ;
赵喜亭等, 2012 )和微型块茎诱导形成技术体系(
李明军等, 2000a ,
2004a ,
2008 ;
Li et al., 2014 ).2007年, 我们在研究中首次发现铁棍山药在特殊的诱导条件下也可形成类原球茎, 并对类原球茎的形成规律(
李纪强, 2013 )、相关基因(
SERK )的克隆及遗传转化(
韩林林等, 2016 )等进行了系统研究.本研究通过建立类原球茎的诱导形成与植株再生技术体系, 为怀山药优良品种和脱毒种苗的快繁及工厂化生产提供了一条新途径. ...
... 目前, 已有很多关于山药组织培养的研究报道.李明军研究团队在怀山药组织培养方面进行了一系列探索.例如, 以茎段为材料, 通过多芽体等途径实现了怀山药无菌苗的快繁(
李明军等, 1997b ,
1999 ), 并在此基础上对铁棍山药快繁培养基进行了优化(
张晓丽等, 2013 ); 另外, 他们还发现山药可以通过诱导形成愈伤组织(
于倩和李明军, 2004 )或微型块茎(
李明军等, 2000a ), 进而形成再生植株; 并对诱导愈伤组织(
李明军等, 2000b )及微型块茎的影响因子(
李明军等, 2008 )进行了研究.
张振霞等(2016) 以广山药(
D. opposita )块茎为外植体, 进行了组织培养技术研究.
马林等(2004) 以带腋芽茎段为外植体, 通过诱导丛生芽实现了黄山药(
D. panthaica )的快速繁殖.而本研究以铁棍山药带腋芽茎段为材料, 首次成功诱导出类原球茎, 并在此基础上建立了类原球茎分化与植株再生体系, 使山药离体培养技术体系得到了进一步完善, 对加快山药良种选育或脱毒种苗的推广应用及产业化进程具有重要意义. ...
山药微型块茎诱导形成的影响因子研究
2
2008
... 薯蓣属植物类原球茎的研究在大薯(
D. alata ) (
许云等, 2014 )与盾叶薯蓣(
D. zingiberensis )中(
彭晓英等, 2010 )已有报道.本实验室从20世纪90年代开始即对怀山药(
D. opposita )的组织培养及其应用进行了系统研究(
李明军, 2004 ), 并建立了怀山药脱毒快繁技术体系(
李明军等, 2003 ,
2004b ;
张晓丽等, 2013 )、种质资源离体保存技术体系(
洪森荣和李明军, 2006 ;
Li et al., 2009 ;
李海兵等, 2010 ;
赵喜亭等, 2012 )和微型块茎诱导形成技术体系(
李明军等, 2000a ,
2004a ,
2008 ;
Li et al., 2014 ).2007年, 我们在研究中首次发现铁棍山药在特殊的诱导条件下也可形成类原球茎, 并对类原球茎的形成规律(
李纪强, 2013 )、相关基因(
SERK )的克隆及遗传转化(
韩林林等, 2016 )等进行了系统研究.本研究通过建立类原球茎的诱导形成与植株再生技术体系, 为怀山药优良品种和脱毒种苗的快繁及工厂化生产提供了一条新途径. ...
... 目前, 已有很多关于山药组织培养的研究报道.李明军研究团队在怀山药组织培养方面进行了一系列探索.例如, 以茎段为材料, 通过多芽体等途径实现了怀山药无菌苗的快繁(
李明军等, 1997b ,
1999 ), 并在此基础上对铁棍山药快繁培养基进行了优化(
张晓丽等, 2013 ); 另外, 他们还发现山药可以通过诱导形成愈伤组织(
于倩和李明军, 2004 )或微型块茎(
李明军等, 2000a ), 进而形成再生植株; 并对诱导愈伤组织(
李明军等, 2000b )及微型块茎的影响因子(
李明军等, 2008 )进行了研究.
张振霞等(2016) 以广山药(
D. opposita )块茎为外植体, 进行了组织培养技术研究.
马林等(2004) 以带腋芽茎段为外植体, 通过诱导丛生芽实现了黄山药(
D. panthaica )的快速繁殖.而本研究以铁棍山药带腋芽茎段为材料, 首次成功诱导出类原球茎, 并在此基础上建立了类原球茎分化与植株再生体系, 使山药离体培养技术体系得到了进一步完善, 对加快山药良种选育或脱毒种苗的推广应用及产业化进程具有重要意义. ...
b). 不同因子对山药愈伤组织诱导的影响
1
2000
... 目前, 已有很多关于山药组织培养的研究报道.李明军研究团队在怀山药组织培养方面进行了一系列探索.例如, 以茎段为材料, 通过多芽体等途径实现了怀山药无菌苗的快繁(
李明军等, 1997b ,
1999 ), 并在此基础上对铁棍山药快繁培养基进行了优化(
张晓丽等, 2013 ); 另外, 他们还发现山药可以通过诱导形成愈伤组织(
于倩和李明军, 2004 )或微型块茎(
李明军等, 2000a ), 进而形成再生植株; 并对诱导愈伤组织(
李明军等, 2000b )及微型块茎的影响因子(
李明军等, 2008 )进行了研究.
张振霞等(2016) 以广山药(
D. opposita )块茎为外植体, 进行了组织培养技术研究.
马林等(2004) 以带腋芽茎段为外植体, 通过诱导丛生芽实现了黄山药(
D. panthaica )的快速繁殖.而本研究以铁棍山药带腋芽茎段为材料, 首次成功诱导出类原球茎, 并在此基础上建立了类原球茎分化与植株再生体系, 使山药离体培养技术体系得到了进一步完善, 对加快山药良种选育或脱毒种苗的推广应用及产业化进程具有重要意义. ...
b). 怀山药的茎段培养和快速繁殖
1
1997
... 目前, 已有很多关于山药组织培养的研究报道.李明军研究团队在怀山药组织培养方面进行了一系列探索.例如, 以茎段为材料, 通过多芽体等途径实现了怀山药无菌苗的快繁(
李明军等, 1997b ,
1999 ), 并在此基础上对铁棍山药快繁培养基进行了优化(
张晓丽等, 2013 ); 另外, 他们还发现山药可以通过诱导形成愈伤组织(
于倩和李明军, 2004 )或微型块茎(
李明军等, 2000a ), 进而形成再生植株; 并对诱导愈伤组织(
李明军等, 2000b )及微型块茎的影响因子(
李明军等, 2008 )进行了研究.
张振霞等(2016) 以广山药(
D. opposita )块茎为外植体, 进行了组织培养技术研究.
马林等(2004) 以带腋芽茎段为外植体, 通过诱导丛生芽实现了黄山药(
D. panthaica )的快速繁殖.而本研究以铁棍山药带腋芽茎段为材料, 首次成功诱导出类原球茎, 并在此基础上建立了类原球茎分化与植株再生体系, 使山药离体培养技术体系得到了进一步完善, 对加快山药良种选育或脱毒种苗的推广应用及产业化进程具有重要意义. ...
怀山药脱毒快繁技术研究
1
2004b
... 薯蓣属植物类原球茎的研究在大薯(
D. alata ) (
许云等, 2014 )与盾叶薯蓣(
D. zingiberensis )中(
彭晓英等, 2010 )已有报道.本实验室从20世纪90年代开始即对怀山药(
D. opposita )的组织培养及其应用进行了系统研究(
李明军, 2004 ), 并建立了怀山药脱毒快繁技术体系(
李明军等, 2003 ,
2004b ;
张晓丽等, 2013 )、种质资源离体保存技术体系(
洪森荣和李明军, 2006 ;
Li et al., 2009 ;
李海兵等, 2010 ;
赵喜亭等, 2012 )和微型块茎诱导形成技术体系(
李明军等, 2000a ,
2004a ,
2008 ;
Li et al., 2014 ).2007年, 我们在研究中首次发现铁棍山药在特殊的诱导条件下也可形成类原球茎, 并对类原球茎的形成规律(
李纪强, 2013 )、相关基因(
SERK )的克隆及遗传转化(
韩林林等, 2016 )等进行了系统研究.本研究通过建立类原球茎的诱导形成与植株再生技术体系, 为怀山药优良品种和脱毒种苗的快繁及工厂化生产提供了一条新途径. ...
怀山药病毒病的研究
1
2003
... 薯蓣属植物类原球茎的研究在大薯(
D. alata ) (
许云等, 2014 )与盾叶薯蓣(
D. zingiberensis )中(
彭晓英等, 2010 )已有报道.本实验室从20世纪90年代开始即对怀山药(
D. opposita )的组织培养及其应用进行了系统研究(
李明军, 2004 ), 并建立了怀山药脱毒快繁技术体系(
李明军等, 2003 ,
2004b ;
张晓丽等, 2013 )、种质资源离体保存技术体系(
洪森荣和李明军, 2006 ;
Li et al., 2009 ;
李海兵等, 2010 ;
赵喜亭等, 2012 )和微型块茎诱导形成技术体系(
李明军等, 2000a ,
2004a ,
2008 ;
Li et al., 2014 ).2007年, 我们在研究中首次发现铁棍山药在特殊的诱导条件下也可形成类原球茎, 并对类原球茎的形成规律(
李纪强, 2013 )、相关基因(
SERK )的克隆及遗传转化(
韩林林等, 2016 )等进行了系统研究.本研究通过建立类原球茎的诱导形成与植株再生技术体系, 为怀山药优良品种和脱毒种苗的快繁及工厂化生产提供了一条新途径. ...
绿色食品铁棍山药栽培技术
1
2016
... 铁棍山药(
Dioscorea opposita cv. ‘Tiegun’)为薯蓣科薯蓣属多年生草本植物, 主产于河南省焦作市温县、武陟等地, 是我国著名的“四大怀药”之一.因其形似铁棍, 运输过程不易折断, 耐储藏, 故又名“铁棍怀山药”(
韩晓勇等, 2013 ).铁棍山药富含氨基酸、维生素、蛋白质和矿物质等(
李青, 2016 ), 是怀山药中的“极品”(
刘雯, 2014 ).但目前铁棍山药离体繁殖速度慢、周期长, 不利于其脱毒种苗的快速繁殖与推广应用, 故有必要探索一条新的山药繁殖途径. ...
铁皮石斛茎段原球茎的诱导、分化与植株再生
1
2016
... 类原球茎(protocorm-like body, PLB)最早于1960年由Morel提出(
姜福星等, 2012 ), 用来描述兰科植物组培过程中产生的类似种子萌发的过渡形态.由于植物类原球茎具有高效的分化再生能力, 并综合了胚性细胞和茎尖分生组织的优点以及其自身的独特性, 因此可广泛应用于植株再生、种质资源保存、基因工程育种、药用次生代谢产物的生产、人工种子的制备以及植物发育生物学研究等领域(
姜福星等, 2012 ).目前, 类原球茎的诱导大多运用于兰科植物, 但随着生物技术的不断发展, 半夏(
Pinellia ternata ) (
江艳华等, 2013 )和狗蔷薇(
Rosa canina ) (
毕玲等, 2012 )等植物也可诱导出类原球茎.研究表明, 不同植物可通过不同的外植体实现类原球茎的诱导, 如茎尖(
Tian et al., 2008 )、根(
罗剑飘等, 2015 )、茎段(
林江波等, 2016 )、根尖(
詹忠根等, 2005 )、种子(
古碧珠等, 2008 )和叶片(
乔永旭等, 2010 )等均可作为诱导类原球茎的材料, 其中叶片应用最为广泛. ...
铁棍山药微型块茎形成的形态解剖及生理生化变化研究
1
2014
... 铁棍山药(
Dioscorea opposita cv. ‘Tiegun’)为薯蓣科薯蓣属多年生草本植物, 主产于河南省焦作市温县、武陟等地, 是我国著名的“四大怀药”之一.因其形似铁棍, 运输过程不易折断, 耐储藏, 故又名“铁棍怀山药”(
韩晓勇等, 2013 ).铁棍山药富含氨基酸、维生素、蛋白质和矿物质等(
李青, 2016 ), 是怀山药中的“极品”(
刘雯, 2014 ).但目前铁棍山药离体繁殖速度慢、周期长, 不利于其脱毒种苗的快速繁殖与推广应用, 故有必要探索一条新的山药繁殖途径. ...
离体培养春兰原球茎增殖的影响因子研究
1
2009
... 类原球茎增殖过程中, 主要利用植物生长调节剂进行处理.6-BA可以促进类原球茎分生区细胞的分裂和生长, 是诱导类原球茎增殖的关键因素之一, 也是应用最广泛的细胞分裂素.
刘晓芳等(2009) 在春兰(
Cymbidium goeringii )类原球茎增殖的研究中发现, 6-BA与NAA的影响最大, 且达极显著水平;
崔广荣等(2009) 在诱导文心兰类原球茎增殖过程中也有类似发现; 而药用植物金线莲(
Anoectochilus formosanus )用来进行PLBs增殖的最适植物生长调节剂为6- BA与NAA (
陈钢等, 1994 ).本实验在增殖培养基中单独添加6-BA, 实现了铁棍山药类原球茎的增殖, 而
张超等(2010) 也在文心兰类原球茎增殖中发现, 单独使用6-BA的增殖效果较与其它植物生长调节剂配合使用的效果更好.此外, 我们在研究中还发现, 较低浓度的6-BA在培养后期会引起类原球茎的分化, 而较高浓度6-BA处理增殖后的类原球茎体积较小, 整齐度较低, 这与
杨美纯等(2000) 以及
罗剑飘等(2015) 的研究结果一致. ...
麒麟火焰兰叶片及根类原球茎的诱导研究
2
2015
... 类原球茎(protocorm-like body, PLB)最早于1960年由Morel提出(
姜福星等, 2012 ), 用来描述兰科植物组培过程中产生的类似种子萌发的过渡形态.由于植物类原球茎具有高效的分化再生能力, 并综合了胚性细胞和茎尖分生组织的优点以及其自身的独特性, 因此可广泛应用于植株再生、种质资源保存、基因工程育种、药用次生代谢产物的生产、人工种子的制备以及植物发育生物学研究等领域(
姜福星等, 2012 ).目前, 类原球茎的诱导大多运用于兰科植物, 但随着生物技术的不断发展, 半夏(
Pinellia ternata ) (
江艳华等, 2013 )和狗蔷薇(
Rosa canina ) (
毕玲等, 2012 )等植物也可诱导出类原球茎.研究表明, 不同植物可通过不同的外植体实现类原球茎的诱导, 如茎尖(
Tian et al., 2008 )、根(
罗剑飘等, 2015 )、茎段(
林江波等, 2016 )、根尖(
詹忠根等, 2005 )、种子(
古碧珠等, 2008 )和叶片(
乔永旭等, 2010 )等均可作为诱导类原球茎的材料, 其中叶片应用最为广泛. ...
... 类原球茎增殖过程中, 主要利用植物生长调节剂进行处理.6-BA可以促进类原球茎分生区细胞的分裂和生长, 是诱导类原球茎增殖的关键因素之一, 也是应用最广泛的细胞分裂素.
刘晓芳等(2009) 在春兰(
Cymbidium goeringii )类原球茎增殖的研究中发现, 6-BA与NAA的影响最大, 且达极显著水平;
崔广荣等(2009) 在诱导文心兰类原球茎增殖过程中也有类似发现; 而药用植物金线莲(
Anoectochilus formosanus )用来进行PLBs增殖的最适植物生长调节剂为6- BA与NAA (
陈钢等, 1994 ).本实验在增殖培养基中单独添加6-BA, 实现了铁棍山药类原球茎的增殖, 而
张超等(2010) 也在文心兰类原球茎增殖中发现, 单独使用6-BA的增殖效果较与其它植物生长调节剂配合使用的效果更好.此外, 我们在研究中还发现, 较低浓度的6-BA在培养后期会引起类原球茎的分化, 而较高浓度6-BA处理增殖后的类原球茎体积较小, 整齐度较低, 这与
杨美纯等(2000) 以及
罗剑飘等(2015) 的研究结果一致. ...
黄山药丛生芽诱导与植株快速繁殖
1
2004
... 目前, 已有很多关于山药组织培养的研究报道.李明军研究团队在怀山药组织培养方面进行了一系列探索.例如, 以茎段为材料, 通过多芽体等途径实现了怀山药无菌苗的快繁(
李明军等, 1997b ,
1999 ), 并在此基础上对铁棍山药快繁培养基进行了优化(
张晓丽等, 2013 ); 另外, 他们还发现山药可以通过诱导形成愈伤组织(
于倩和李明军, 2004 )或微型块茎(
李明军等, 2000a ), 进而形成再生植株; 并对诱导愈伤组织(
李明军等, 2000b )及微型块茎的影响因子(
李明军等, 2008 )进行了研究.
张振霞等(2016) 以广山药(
D. opposita )块茎为外植体, 进行了组织培养技术研究.
马林等(2004) 以带腋芽茎段为外植体, 通过诱导丛生芽实现了黄山药(
D. panthaica )的快速繁殖.而本研究以铁棍山药带腋芽茎段为材料, 首次成功诱导出类原球茎, 并在此基础上建立了类原球茎分化与植株再生体系, 使山药离体培养技术体系得到了进一步完善, 对加快山药良种选育或脱毒种苗的推广应用及产业化进程具有重要意义. ...
盾叶薯蓣类原球茎的离体诱导及快繁体系的建立
2
2010
... 薯蓣属植物类原球茎的研究在大薯(
D. alata ) (
许云等, 2014 )与盾叶薯蓣(
D. zingiberensis )中(
彭晓英等, 2010 )已有报道.本实验室从20世纪90年代开始即对怀山药(
D. opposita )的组织培养及其应用进行了系统研究(
李明军, 2004 ), 并建立了怀山药脱毒快繁技术体系(
李明军等, 2003 ,
2004b ;
张晓丽等, 2013 )、种质资源离体保存技术体系(
洪森荣和李明军, 2006 ;
Li et al., 2009 ;
李海兵等, 2010 ;
赵喜亭等, 2012 )和微型块茎诱导形成技术体系(
李明军等, 2000a ,
2004a ,
2008 ;
Li et al., 2014 ).2007年, 我们在研究中首次发现铁棍山药在特殊的诱导条件下也可形成类原球茎, 并对类原球茎的形成规律(
李纪强, 2013 )、相关基因(
SERK )的克隆及遗传转化(
韩林林等, 2016 )等进行了系统研究.本研究通过建立类原球茎的诱导形成与植株再生技术体系, 为怀山药优良品种和脱毒种苗的快繁及工厂化生产提供了一条新途径. ...
... 植物类原球茎诱导的影响因素有很多, 其中植物激素种类及其配比是最重要的因素.2,4-D是霍山石斛(
Dendrobium huoshanense )类原球茎诱导过程中的主要影响因素, 而诱导蝴蝶兰(
Phalaenopsis aphrodite ) (
崔广荣等, 2007 )、石斛兰(
D. nobile ) (
王丽萍和梁淑云, 2010 )及文心兰(
Oncidium hybridum ) (
陈晓旋等, 2010 )等兰花类原球茎主要利用细胞分裂素(6-BA或KT)与生长素(2,4-D)的组合(
姜福星等, 2012 ).在大薯诱导类原球茎的研究中发现, 适宜浓度的6-BA与NAA组合可诱导类原球茎的形成(
许云等, 2014 ); 适宜浓度的6-BA与KT组合可诱导盾叶薯蓣的形成(
彭晓英等, 2010 ).本实验以铁棍山药带腋芽茎段为材料, 探讨了不同浓度TDZ对铁棍山药类原球茎诱导形成的影响, 发现适宜浓度(1.0 mg·L
-1 )的TDZ有利于铁棍山药类原球茎的形成, 而较低浓度的TDZ则有利于芽的诱导, 较高浓度的TDZ虽然也能诱导类原球茎的形成, 但类原球茎为白色, 并且可能会导致畸形类原球茎的产生, 这与
张玉等(2002) 发现TDZ比6-BA更有利于蝴蝶兰PLBs诱导的结果一致.可见, 不同植物诱导类原球茎所需的植物激素种类及组合存在差异, 在诱导植物类原球茎过程中, 要全面考虑不同植物激素在其中的作用效果, 同时不能忽视植物激素间的协同效应(
崔广荣等, 2009 ). ...
蝴蝶兰组培苗类原球茎的诱导与分化
1
2010
... 类原球茎(protocorm-like body, PLB)最早于1960年由Morel提出(
姜福星等, 2012 ), 用来描述兰科植物组培过程中产生的类似种子萌发的过渡形态.由于植物类原球茎具有高效的分化再生能力, 并综合了胚性细胞和茎尖分生组织的优点以及其自身的独特性, 因此可广泛应用于植株再生、种质资源保存、基因工程育种、药用次生代谢产物的生产、人工种子的制备以及植物发育生物学研究等领域(
姜福星等, 2012 ).目前, 类原球茎的诱导大多运用于兰科植物, 但随着生物技术的不断发展, 半夏(
Pinellia ternata ) (
江艳华等, 2013 )和狗蔷薇(
Rosa canina ) (
毕玲等, 2012 )等植物也可诱导出类原球茎.研究表明, 不同植物可通过不同的外植体实现类原球茎的诱导, 如茎尖(
Tian et al., 2008 )、根(
罗剑飘等, 2015 )、茎段(
林江波等, 2016 )、根尖(
詹忠根等, 2005 )、种子(
古碧珠等, 2008 )和叶片(
乔永旭等, 2010 )等均可作为诱导类原球茎的材料, 其中叶片应用最为广泛. ...
铁皮石斛原球茎诱导与增殖研究
1
2010
... 植物类原球茎诱导的影响因素有很多, 其中植物激素种类及其配比是最重要的因素.2,4-D是霍山石斛(
Dendrobium huoshanense )类原球茎诱导过程中的主要影响因素, 而诱导蝴蝶兰(
Phalaenopsis aphrodite ) (
崔广荣等, 2007 )、石斛兰(
D. nobile ) (
王丽萍和梁淑云, 2010 )及文心兰(
Oncidium hybridum ) (
陈晓旋等, 2010 )等兰花类原球茎主要利用细胞分裂素(6-BA或KT)与生长素(2,4-D)的组合(
姜福星等, 2012 ).在大薯诱导类原球茎的研究中发现, 适宜浓度的6-BA与NAA组合可诱导类原球茎的形成(
许云等, 2014 ); 适宜浓度的6-BA与KT组合可诱导盾叶薯蓣的形成(
彭晓英等, 2010 ).本实验以铁棍山药带腋芽茎段为材料, 探讨了不同浓度TDZ对铁棍山药类原球茎诱导形成的影响, 发现适宜浓度(1.0 mg·L
-1 )的TDZ有利于铁棍山药类原球茎的形成, 而较低浓度的TDZ则有利于芽的诱导, 较高浓度的TDZ虽然也能诱导类原球茎的形成, 但类原球茎为白色, 并且可能会导致畸形类原球茎的产生, 这与
张玉等(2002) 发现TDZ比6-BA更有利于蝴蝶兰PLBs诱导的结果一致.可见, 不同植物诱导类原球茎所需的植物激素种类及组合存在差异, 在诱导植物类原球茎过程中, 要全面考虑不同植物激素在其中的作用效果, 同时不能忽视植物激素间的协同效应(
崔广荣等, 2009 ). ...
大薯类原球茎的离体诱导及再生体系的建立
2
2014
... 薯蓣属植物类原球茎的研究在大薯(
D. alata ) (
许云等, 2014 )与盾叶薯蓣(
D. zingiberensis )中(
彭晓英等, 2010 )已有报道.本实验室从20世纪90年代开始即对怀山药(
D. opposita )的组织培养及其应用进行了系统研究(
李明军, 2004 ), 并建立了怀山药脱毒快繁技术体系(
李明军等, 2003 ,
2004b ;
张晓丽等, 2013 )、种质资源离体保存技术体系(
洪森荣和李明军, 2006 ;
Li et al., 2009 ;
李海兵等, 2010 ;
赵喜亭等, 2012 )和微型块茎诱导形成技术体系(
李明军等, 2000a ,
2004a ,
2008 ;
Li et al., 2014 ).2007年, 我们在研究中首次发现铁棍山药在特殊的诱导条件下也可形成类原球茎, 并对类原球茎的形成规律(
李纪强, 2013 )、相关基因(
SERK )的克隆及遗传转化(
韩林林等, 2016 )等进行了系统研究.本研究通过建立类原球茎的诱导形成与植株再生技术体系, 为怀山药优良品种和脱毒种苗的快繁及工厂化生产提供了一条新途径. ...
... 植物类原球茎诱导的影响因素有很多, 其中植物激素种类及其配比是最重要的因素.2,4-D是霍山石斛(
Dendrobium huoshanense )类原球茎诱导过程中的主要影响因素, 而诱导蝴蝶兰(
Phalaenopsis aphrodite ) (
崔广荣等, 2007 )、石斛兰(
D. nobile ) (
王丽萍和梁淑云, 2010 )及文心兰(
Oncidium hybridum ) (
陈晓旋等, 2010 )等兰花类原球茎主要利用细胞分裂素(6-BA或KT)与生长素(2,4-D)的组合(
姜福星等, 2012 ).在大薯诱导类原球茎的研究中发现, 适宜浓度的6-BA与NAA组合可诱导类原球茎的形成(
许云等, 2014 ); 适宜浓度的6-BA与KT组合可诱导盾叶薯蓣的形成(
彭晓英等, 2010 ).本实验以铁棍山药带腋芽茎段为材料, 探讨了不同浓度TDZ对铁棍山药类原球茎诱导形成的影响, 发现适宜浓度(1.0 mg·L
-1 )的TDZ有利于铁棍山药类原球茎的形成, 而较低浓度的TDZ则有利于芽的诱导, 较高浓度的TDZ虽然也能诱导类原球茎的形成, 但类原球茎为白色, 并且可能会导致畸形类原球茎的产生, 这与
张玉等(2002) 发现TDZ比6-BA更有利于蝴蝶兰PLBs诱导的结果一致.可见, 不同植物诱导类原球茎所需的植物激素种类及组合存在差异, 在诱导植物类原球茎过程中, 要全面考虑不同植物激素在其中的作用效果, 同时不能忽视植物激素间的协同效应(
崔广荣等, 2009 ). ...
外部因子对蝴蝶兰叶片原球茎状体发生的影响
1
2000
... 类原球茎增殖过程中, 主要利用植物生长调节剂进行处理.6-BA可以促进类原球茎分生区细胞的分裂和生长, 是诱导类原球茎增殖的关键因素之一, 也是应用最广泛的细胞分裂素.
刘晓芳等(2009) 在春兰(
Cymbidium goeringii )类原球茎增殖的研究中发现, 6-BA与NAA的影响最大, 且达极显著水平;
崔广荣等(2009) 在诱导文心兰类原球茎增殖过程中也有类似发现; 而药用植物金线莲(
Anoectochilus formosanus )用来进行PLBs增殖的最适植物生长调节剂为6- BA与NAA (
陈钢等, 1994 ).本实验在增殖培养基中单独添加6-BA, 实现了铁棍山药类原球茎的增殖, 而
张超等(2010) 也在文心兰类原球茎增殖中发现, 单独使用6-BA的增殖效果较与其它植物生长调节剂配合使用的效果更好.此外, 我们在研究中还发现, 较低浓度的6-BA在培养后期会引起类原球茎的分化, 而较高浓度6-BA处理增殖后的类原球茎体积较小, 整齐度较低, 这与
杨美纯等(2000) 以及
罗剑飘等(2015) 的研究结果一致. ...
怀山药微型块茎愈伤组织的诱导形成及高频率再生
1
2004
... 目前, 已有很多关于山药组织培养的研究报道.李明军研究团队在怀山药组织培养方面进行了一系列探索.例如, 以茎段为材料, 通过多芽体等途径实现了怀山药无菌苗的快繁(
李明军等, 1997b ,
1999 ), 并在此基础上对铁棍山药快繁培养基进行了优化(
张晓丽等, 2013 ); 另外, 他们还发现山药可以通过诱导形成愈伤组织(
于倩和李明军, 2004 )或微型块茎(
李明军等, 2000a ), 进而形成再生植株; 并对诱导愈伤组织(
李明军等, 2000b )及微型块茎的影响因子(
李明军等, 2008 )进行了研究.
张振霞等(2016) 以广山药(
D. opposita )块茎为外植体, 进行了组织培养技术研究.
马林等(2004) 以带腋芽茎段为外植体, 通过诱导丛生芽实现了黄山药(
D. panthaica )的快速繁殖.而本研究以铁棍山药带腋芽茎段为材料, 首次成功诱导出类原球茎, 并在此基础上建立了类原球茎分化与植株再生体系, 使山药离体培养技术体系得到了进一步完善, 对加快山药良种选育或脱毒种苗的推广应用及产业化进程具有重要意义. ...
铁皮石斛离体根尖经体细胞胚再生植株研究
1
2005
... 类原球茎(protocorm-like body, PLB)最早于1960年由Morel提出(
姜福星等, 2012 ), 用来描述兰科植物组培过程中产生的类似种子萌发的过渡形态.由于植物类原球茎具有高效的分化再生能力, 并综合了胚性细胞和茎尖分生组织的优点以及其自身的独特性, 因此可广泛应用于植株再生、种质资源保存、基因工程育种、药用次生代谢产物的生产、人工种子的制备以及植物发育生物学研究等领域(
姜福星等, 2012 ).目前, 类原球茎的诱导大多运用于兰科植物, 但随着生物技术的不断发展, 半夏(
Pinellia ternata ) (
江艳华等, 2013 )和狗蔷薇(
Rosa canina ) (
毕玲等, 2012 )等植物也可诱导出类原球茎.研究表明, 不同植物可通过不同的外植体实现类原球茎的诱导, 如茎尖(
Tian et al., 2008 )、根(
罗剑飘等, 2015 )、茎段(
林江波等, 2016 )、根尖(
詹忠根等, 2005 )、种子(
古碧珠等, 2008 )和叶片(
乔永旭等, 2010 )等均可作为诱导类原球茎的材料, 其中叶片应用最为广泛. ...
基本培养基、碳源及植物生长调节剂对文心兰原球茎增殖的影响
1
2010
... 类原球茎增殖过程中, 主要利用植物生长调节剂进行处理.6-BA可以促进类原球茎分生区细胞的分裂和生长, 是诱导类原球茎增殖的关键因素之一, 也是应用最广泛的细胞分裂素.
刘晓芳等(2009) 在春兰(
Cymbidium goeringii )类原球茎增殖的研究中发现, 6-BA与NAA的影响最大, 且达极显著水平;
崔广荣等(2009) 在诱导文心兰类原球茎增殖过程中也有类似发现; 而药用植物金线莲(
Anoectochilus formosanus )用来进行PLBs增殖的最适植物生长调节剂为6- BA与NAA (
陈钢等, 1994 ).本实验在增殖培养基中单独添加6-BA, 实现了铁棍山药类原球茎的增殖, 而
张超等(2010) 也在文心兰类原球茎增殖中发现, 单独使用6-BA的增殖效果较与其它植物生长调节剂配合使用的效果更好.此外, 我们在研究中还发现, 较低浓度的6-BA在培养后期会引起类原球茎的分化, 而较高浓度6-BA处理增殖后的类原球茎体积较小, 整齐度较低, 这与
杨美纯等(2000) 以及
罗剑飘等(2015) 的研究结果一致. ...
植物生长调节剂在观赏植物组织培养中的应用
1
2012
... 在植物组织培养过程中, 植物的生根情况会直接影响植物的成活率.影响植物生根的因素有很多, 如环境、品种差异以及植物生长调节剂的使用等.其中, 植物生长调节剂的种类和浓度起着至关重要的作用.常用的植物生根调节剂有PP333、NAA、IAA和IBA.据报道, NAA为最经济高效的生根激素, 多数花卉NAA浓度为0.1-1.0 mg·L
-1 时培养15天左右就能生根(
张瑞姿, 2012 ).PP333又称多效唑, 具有延缓植物生长、抑制茎枝伸长、促进分蘖、促进成花和坐果、增强抗旱及抗寒性、提高耐盐性和延缓植物衰老等多种效用(
李明军, 1995 ).
李明军等(1997a) 研究发现, 适宜浓度的PP333可有效促进玉米(
Zea mays )再生苗根系的生长, 并使玉米再生苗的移栽成活率由原来的不到30%提高到85%以上.本实验使用0.05 mg·L
-1 NAA和1.0 mg·L
-1 PP333达到了促进铁棍山药类原球茎分化苗生根的目的. ...
铁棍山药试管苗快繁培养基的优化
2
2013
... 薯蓣属植物类原球茎的研究在大薯(
D. alata ) (
许云等, 2014 )与盾叶薯蓣(
D. zingiberensis )中(
彭晓英等, 2010 )已有报道.本实验室从20世纪90年代开始即对怀山药(
D. opposita )的组织培养及其应用进行了系统研究(
李明军, 2004 ), 并建立了怀山药脱毒快繁技术体系(
李明军等, 2003 ,
2004b ;
张晓丽等, 2013 )、种质资源离体保存技术体系(
洪森荣和李明军, 2006 ;
Li et al., 2009 ;
李海兵等, 2010 ;
赵喜亭等, 2012 )和微型块茎诱导形成技术体系(
李明军等, 2000a ,
2004a ,
2008 ;
Li et al., 2014 ).2007年, 我们在研究中首次发现铁棍山药在特殊的诱导条件下也可形成类原球茎, 并对类原球茎的形成规律(
李纪强, 2013 )、相关基因(
SERK )的克隆及遗传转化(
韩林林等, 2016 )等进行了系统研究.本研究通过建立类原球茎的诱导形成与植株再生技术体系, 为怀山药优良品种和脱毒种苗的快繁及工厂化生产提供了一条新途径. ...
... 目前, 已有很多关于山药组织培养的研究报道.李明军研究团队在怀山药组织培养方面进行了一系列探索.例如, 以茎段为材料, 通过多芽体等途径实现了怀山药无菌苗的快繁(
李明军等, 1997b ,
1999 ), 并在此基础上对铁棍山药快繁培养基进行了优化(
张晓丽等, 2013 ); 另外, 他们还发现山药可以通过诱导形成愈伤组织(
于倩和李明军, 2004 )或微型块茎(
李明军等, 2000a ), 进而形成再生植株; 并对诱导愈伤组织(
李明军等, 2000b )及微型块茎的影响因子(
李明军等, 2008 )进行了研究.
张振霞等(2016) 以广山药(
D. opposita )块茎为外植体, 进行了组织培养技术研究.
马林等(2004) 以带腋芽茎段为外植体, 通过诱导丛生芽实现了黄山药(
D. panthaica )的快速繁殖.而本研究以铁棍山药带腋芽茎段为材料, 首次成功诱导出类原球茎, 并在此基础上建立了类原球茎分化与植株再生体系, 使山药离体培养技术体系得到了进一步完善, 对加快山药良种选育或脱毒种苗的推广应用及产业化进程具有重要意义. ...
蝴蝶兰叶片诱导类原球茎初探
1
2012
... 植物类原球茎诱导的影响因素有很多, 其中植物激素种类及其配比是最重要的因素.2,4-D是霍山石斛(
Dendrobium huoshanense )类原球茎诱导过程中的主要影响因素, 而诱导蝴蝶兰(
Phalaenopsis aphrodite ) (
崔广荣等, 2007 )、石斛兰(
D. nobile ) (
王丽萍和梁淑云, 2010 )及文心兰(
Oncidium hybridum ) (
陈晓旋等, 2010 )等兰花类原球茎主要利用细胞分裂素(6-BA或KT)与生长素(2,4-D)的组合(
姜福星等, 2012 ).在大薯诱导类原球茎的研究中发现, 适宜浓度的6-BA与NAA组合可诱导类原球茎的形成(
许云等, 2014 ); 适宜浓度的6-BA与KT组合可诱导盾叶薯蓣的形成(
彭晓英等, 2010 ).本实验以铁棍山药带腋芽茎段为材料, 探讨了不同浓度TDZ对铁棍山药类原球茎诱导形成的影响, 发现适宜浓度(1.0 mg·L
-1 )的TDZ有利于铁棍山药类原球茎的形成, 而较低浓度的TDZ则有利于芽的诱导, 较高浓度的TDZ虽然也能诱导类原球茎的形成, 但类原球茎为白色, 并且可能会导致畸形类原球茎的产生, 这与
张玉等(2002) 发现TDZ比6-BA更有利于蝴蝶兰PLBs诱导的结果一致.可见, 不同植物诱导类原球茎所需的植物激素种类及组合存在差异, 在诱导植物类原球茎过程中, 要全面考虑不同植物激素在其中的作用效果, 同时不能忽视植物激素间的协同效应(
崔广荣等, 2009 ). ...
广山药组织培养技术的研究
1
2016
... 目前, 已有很多关于山药组织培养的研究报道.李明军研究团队在怀山药组织培养方面进行了一系列探索.例如, 以茎段为材料, 通过多芽体等途径实现了怀山药无菌苗的快繁(
李明军等, 1997b ,
1999 ), 并在此基础上对铁棍山药快繁培养基进行了优化(
张晓丽等, 2013 ); 另外, 他们还发现山药可以通过诱导形成愈伤组织(
于倩和李明军, 2004 )或微型块茎(
李明军等, 2000a ), 进而形成再生植株; 并对诱导愈伤组织(
李明军等, 2000b )及微型块茎的影响因子(
李明军等, 2008 )进行了研究.
张振霞等(2016) 以广山药(
D. opposita )块茎为外植体, 进行了组织培养技术研究.
马林等(2004) 以带腋芽茎段为外植体, 通过诱导丛生芽实现了黄山药(
D. panthaica )的快速繁殖.而本研究以铁棍山药带腋芽茎段为材料, 首次成功诱导出类原球茎, 并在此基础上建立了类原球茎分化与植株再生体系, 使山药离体培养技术体系得到了进一步完善, 对加快山药良种选育或脱毒种苗的推广应用及产业化进程具有重要意义. ...
山药种质包埋玻璃化超低温保存再生植株的稳定性分析
1
2012
... 薯蓣属植物类原球茎的研究在大薯(
D. alata ) (
许云等, 2014 )与盾叶薯蓣(
D. zingiberensis )中(
彭晓英等, 2010 )已有报道.本实验室从20世纪90年代开始即对怀山药(
D. opposita )的组织培养及其应用进行了系统研究(
李明军, 2004 ), 并建立了怀山药脱毒快繁技术体系(
李明军等, 2003 ,
2004b ;
张晓丽等, 2013 )、种质资源离体保存技术体系(
洪森荣和李明军, 2006 ;
Li et al., 2009 ;
李海兵等, 2010 ;
赵喜亭等, 2012 )和微型块茎诱导形成技术体系(
李明军等, 2000a ,
2004a ,
2008 ;
Li et al., 2014 ).2007年, 我们在研究中首次发现铁棍山药在特殊的诱导条件下也可形成类原球茎, 并对类原球茎的形成规律(
李纪强, 2013 )、相关基因(
SERK )的克隆及遗传转化(
韩林林等, 2016 )等进行了系统研究.本研究通过建立类原球茎的诱导形成与植株再生技术体系, 为怀山药优良品种和脱毒种苗的快繁及工厂化生产提供了一条新途径. ...
1
2014
... 薯蓣属植物类原球茎的研究在大薯(
D. alata ) (
许云等, 2014 )与盾叶薯蓣(
D. zingiberensis )中(
彭晓英等, 2010 )已有报道.本实验室从20世纪90年代开始即对怀山药(
D. opposita )的组织培养及其应用进行了系统研究(
李明军, 2004 ), 并建立了怀山药脱毒快繁技术体系(
李明军等, 2003 ,
2004b ;
张晓丽等, 2013 )、种质资源离体保存技术体系(
洪森荣和李明军, 2006 ;
Li et al., 2009 ;
李海兵等, 2010 ;
赵喜亭等, 2012 )和微型块茎诱导形成技术体系(
李明军等, 2000a ,
2004a ,
2008 ;
Li et al., 2014 ).2007年, 我们在研究中首次发现铁棍山药在特殊的诱导条件下也可形成类原球茎, 并对类原球茎的形成规律(
李纪强, 2013 )、相关基因(
SERK )的克隆及遗传转化(
韩林林等, 2016 )等进行了系统研究.本研究通过建立类原球茎的诱导形成与植株再生技术体系, 为怀山药优良品种和脱毒种苗的快繁及工厂化生产提供了一条新途径. ...
1
2009
... 薯蓣属植物类原球茎的研究在大薯(
D. alata ) (
许云等, 2014 )与盾叶薯蓣(
D. zingiberensis )中(
彭晓英等, 2010 )已有报道.本实验室从20世纪90年代开始即对怀山药(
D. opposita )的组织培养及其应用进行了系统研究(
李明军, 2004 ), 并建立了怀山药脱毒快繁技术体系(
李明军等, 2003 ,
2004b ;
张晓丽等, 2013 )、种质资源离体保存技术体系(
洪森荣和李明军, 2006 ;
Li et al., 2009 ;
李海兵等, 2010 ;
赵喜亭等, 2012 )和微型块茎诱导形成技术体系(
李明军等, 2000a ,
2004a ,
2008 ;
Li et al., 2014 ).2007年, 我们在研究中首次发现铁棍山药在特殊的诱导条件下也可形成类原球茎, 并对类原球茎的形成规律(
李纪强, 2013 )、相关基因(
SERK )的克隆及遗传转化(
韩林林等, 2016 )等进行了系统研究.本研究通过建立类原球茎的诱导形成与植株再生技术体系, 为怀山药优良品种和脱毒种苗的快繁及工厂化生产提供了一条新途径. ...
1
2008
... 类原球茎(protocorm-like body, PLB)最早于1960年由Morel提出(
姜福星等, 2012 ), 用来描述兰科植物组培过程中产生的类似种子萌发的过渡形态.由于植物类原球茎具有高效的分化再生能力, 并综合了胚性细胞和茎尖分生组织的优点以及其自身的独特性, 因此可广泛应用于植株再生、种质资源保存、基因工程育种、药用次生代谢产物的生产、人工种子的制备以及植物发育生物学研究等领域(
姜福星等, 2012 ).目前, 类原球茎的诱导大多运用于兰科植物, 但随着生物技术的不断发展, 半夏(
Pinellia ternata ) (
江艳华等, 2013 )和狗蔷薇(
Rosa canina ) (
毕玲等, 2012 )等植物也可诱导出类原球茎.研究表明, 不同植物可通过不同的外植体实现类原球茎的诱导, 如茎尖(
Tian et al., 2008 )、根(
罗剑飘等, 2015 )、茎段(
林江波等, 2016 )、根尖(
詹忠根等, 2005 )、种子(
古碧珠等, 2008 )和叶片(
乔永旭等, 2010 )等均可作为诱导类原球茎的材料, 其中叶片应用最为广泛. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}