黄淦, 王潇, 金学锋, 王小菁, 王亚琴*, 华南师范大学生命科学学院, 广东省植物发育生物工程重点实验室, 广州 510631 HuangGan, WangXiao, JinXuefeng, WangXiaojing, WangYaqin*, Guangdong Provincial Key Laboratory of Biotechnology for Plant Development, School of Life Sciences, South China Normal University, Guangzhou 510631, China 引用本文 黄淦, 王潇, 金学锋, 王小菁, 王亚琴. 拟南芥谷氧还蛋白GRXC9负调控叶片大小. , 2017, 52(5): 550-559
Abstract: Glutaredoxin (GRX), a kind of small molecule and heat-stable protein with a CXXC/S motif as an active site, is involved in a variety of glutathione-dependent redox reactions. In this study, we performed cloning and expression, subcellular localization and functional analysis of GRXC9. GRXC9 could be detected in tissues and organs such as root, stem, leaf, flower and silique, which was consistent with GUS assay. GRXC9-GFP was localized in the nucleus and cytoplasm. GRXC9 overexpression lines showed smaller leaves than the wild type, and further observations showed smaller palisade cells in overexpression lines than the wild type, but the total number of cells was similar. The expression of the leaf size-related genes AN, LNG1 and LNG2 was significantly decreased in overexpression lines, so GRXC9 plays a negative role by inhibiting their expression and inducing small leaves. Taken together, GRXC9 may play a crucial role in regulating leaf development.
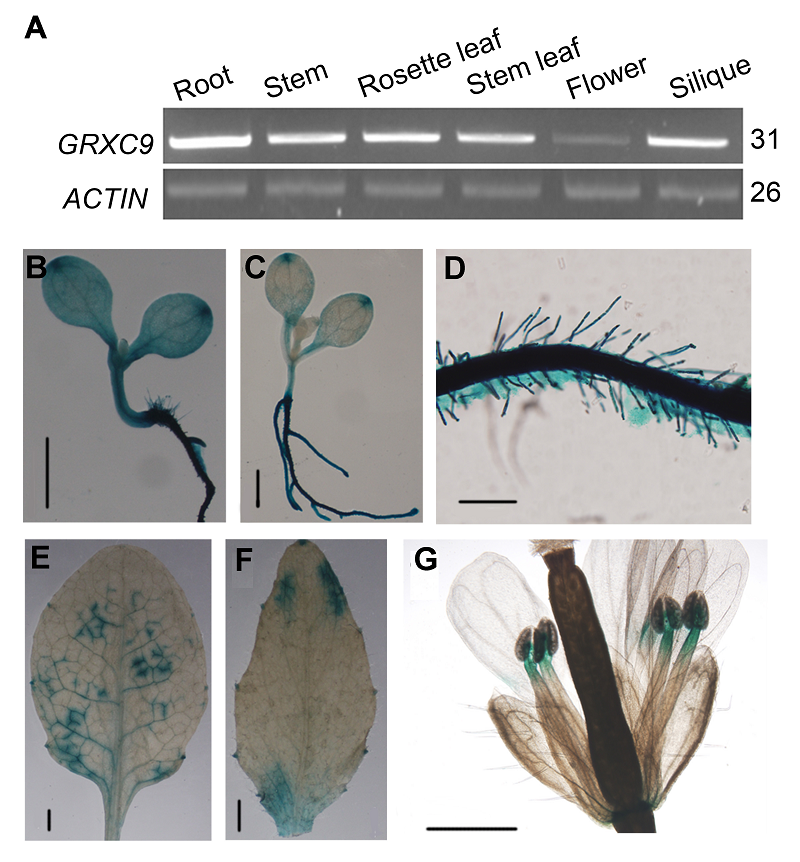
图1 拟南芥GRXC9的组织表达特性 (A) GRXC9在不同组织中的表达情况(以ACTIN为内参; PCR反应循环数标注在图片右侧); (B)-(G) GRXC9的组织化学定位((B) 子叶期(Bar=1 mm); (C) 四叶期(Bar=1 mm); (D) 根(Bar=0.1 mm); (E) 莲座叶; (F) 茎生叶; (G) 花)。(E)-(G) Bar=1 mm Figure 1 Tissue-specific expression of GRXC9 in Arabi- dopsis thaliana (A) Expression level of GRXC9 in root, stem, rosette leaf, stem leaf, flower and silique (ACTIN was used as internal reference; The number of PCR cycle was listed on the right of the images); (B)-(G) Histochemical localization of GUS activity directed by GRXC9::GUS fusions ((B) Cotyledon stage (Bar=1 mm); (C) Four-leaf stage (Bar=1 mm); (D) Root (Bar=0.1 mm); (E) Rosette leaf; (F) Stem leaf; (G) Flower). (E)-(G) Bar=1 mm
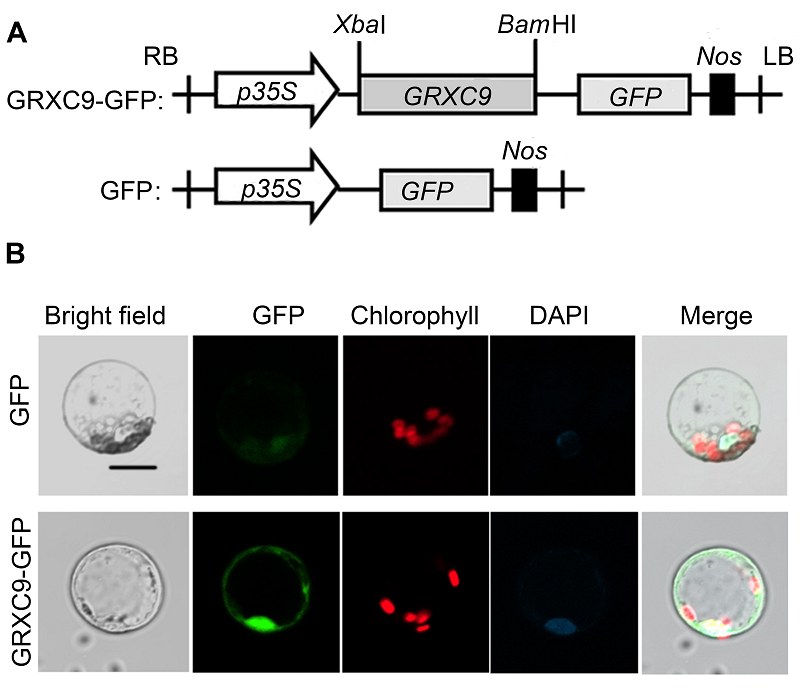
2.2 GRXC9蛋白的亚细胞定位为确定GRXC9蛋白在细胞中的分布, 构建了GRXC9- GFP蛋白融合表达载体(图2A), 采用转化原生质体瞬时表达该融合蛋白, 确定其亚细胞定位。结果显示, 在细胞核和细胞质中均出现荧光信号(图2B), 说明GRXC9蛋白主要定位于细胞核和细胞质中。 图2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-550/img_2.png<b>图2</b> 拟南芥GRXC9的亚细胞定位<br/>(A) p35S::GRXC9-GFP载体结构; (B) GRXC9和GFP的亚细胞共定位(GFP表示GFP荧光信号; Bright field表示明场; DAPI表示DAPI染色荧光信号; Merge表示整合后的图片) (Bar=10 μm)。<br/><b>Figure 2</b> Subcellular localization of GRXC9 of <i>Arabidop- sis thaliana<br/></i>(A) Schematic representation of the p35S::GRXC9-GFP construct; (B) Co-localization of GFP fused to GRXC9 (GFP indicates confocal images under the GFP channel; Bright field indicates confocal images of the same cells with transmitted light; DAPI indicates images under the DAPI channel; Merge indicates the merged images of GFP confocal images and transmitted light) (Bar=10 μm). Figure 2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-550/img_2.png<b>图2</b> 拟南芥GRXC9的亚细胞定位<br/>(A) p35S::GRXC9-GFP载体结构; (B) GRXC9和GFP的亚细胞共定位(GFP表示GFP荧光信号; Bright field表示明场; DAPI表示DAPI染色荧光信号; Merge表示整合后的图片) (Bar=10 μm)。<br/><b>Figure 2</b> Subcellular localization of GRXC9 of <i>Arabidop- sis thaliana<br/></i>(A) Schematic representation of the p35S::GRXC9-GFP construct; (B) Co-localization of GFP fused to GRXC9 (GFP indicates confocal images under the GFP channel; Bright field indicates confocal images of the same cells with transmitted light; DAPI indicates images under the DAPI channel; Merge indicates the merged images of GFP confocal images and transmitted light) (Bar=10 μm).
图2 拟南芥GRXC9的亚细胞定位 (A) p35S::GRXC9-GFP载体结构; (B) GRXC9和GFP的亚细胞共定位(GFP表示GFP荧光信号; Bright field表示明场; DAPI表示DAPI染色荧光信号; Merge表示整合后的图片) (Bar=10 μm)。 Figure 2 Subcellular localization of GRXC9 of Arabidop- sis thaliana (A) Schematic representation of the p35S::GRXC9-GFP construct; (B) Co-localization of GFP fused to GRXC9 (GFP indicates confocal images under the GFP channel; Bright field indicates confocal images of the same cells with transmitted light; DAPI indicates images under the DAPI channel; Merge indicates the merged images of GFP confocal images and transmitted light) (Bar=10 μm).
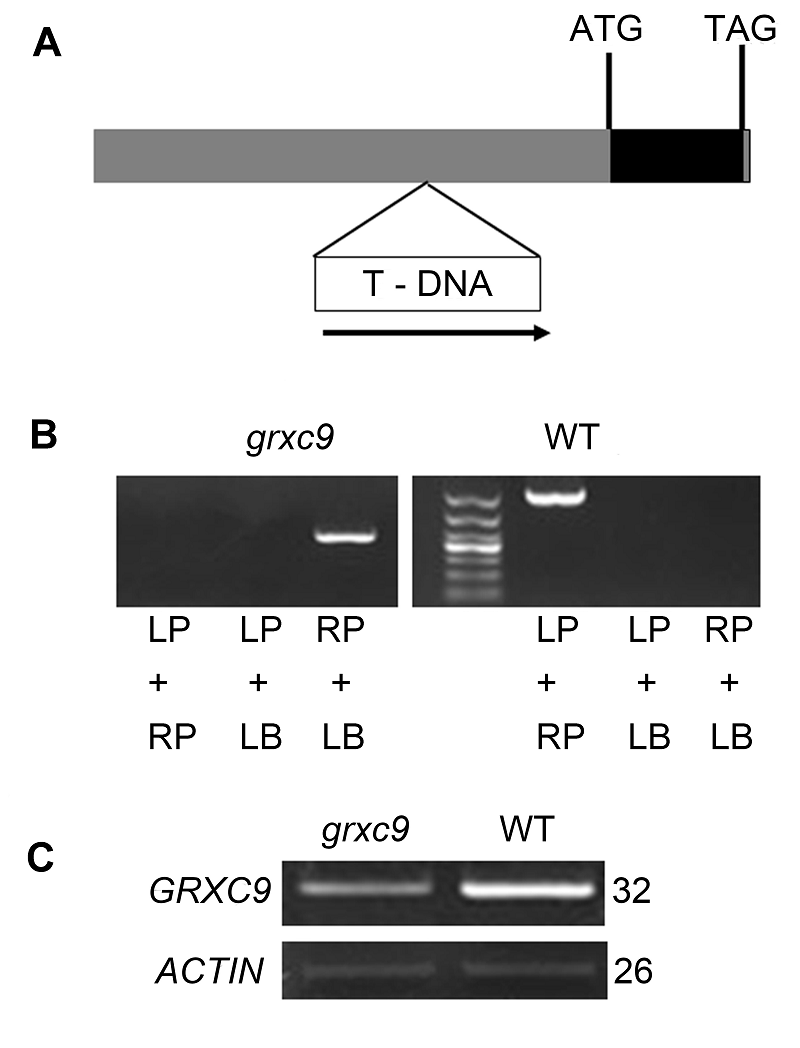
2.3 grxc9突变体纯合株系的鉴定从TAIR (The Arabidopsis Information Resource)和SALK研究院遗传分析实验室(Salk Institute Geno- mic Analysis Laboratory)购买的突变体grxc9 (种子编号: SAIL_435B09.v1)的插入位点在AT1G28480.1基因上游的启动子区域(图3A)。根据插入位点分别设计引物RP和LP, 结合GEBD (Genome Express Br- owser Server)提供的T-DNA筛选引物LB进行纯合体筛选(表1)。 图3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-550/img_3.png<b>图3</b> 拟南芥<i>grxc9</i>突变体的鉴定<br/>(A) <i>grxc9</i>的T-DNA插入位点示意图; (B) <i>grxc9</i>纯合株系的三引物鉴定; (C) RT-PCR分析<i>GRXC9</i>在野生型和<i>grxc9</i>突变体中的表达<br/><b>Figure 3</b> Molecular identification of Arabidopsis <i>grxc9</i> mu- tant<br/>(A) T-DNA insertion sites in <i>grxc9</i>; (B) Identification of <i>grxc9</i> using the three primer method; (C) Reverse trans- criptase PCR analysis of <i>GRXC9 </i>expression in WT and <i>grxc9</i> Figure 3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-550/img_3.png<b>图3</b> 拟南芥<i>grxc9</i>突变体的鉴定<br/>(A) <i>grxc9</i>的T-DNA插入位点示意图; (B) <i>grxc9</i>纯合株系的三引物鉴定; (C) RT-PCR分析<i>GRXC9</i>在野生型和<i>grxc9</i>突变体中的表达<br/><b>Figure 3</b> Molecular identification of Arabidopsis <i>grxc9</i> mu- tant<br/>(A) T-DNA insertion sites in <i>grxc9</i>; (B) Identification of <i>grxc9</i> using the three primer method; (C) Reverse trans- criptase PCR analysis of <i>GRXC9 </i>expression in WT and <i>grxc9</i>
图3 拟南芥grxc9突变体的鉴定 (A) grxc9的T-DNA插入位点示意图; (B) grxc9纯合株系的三引物鉴定; (C) RT-PCR分析GRXC9在野生型和grxc9突变体中的表达 Figure 3 Molecular identification of Arabidopsis grxc9 mu- tant (A) T-DNA insertion sites in grxc9; (B) Identification of grxc9 using the three primer method; (C) Reverse trans- criptase PCR analysis of GRXC9 expression in WT and grxc9
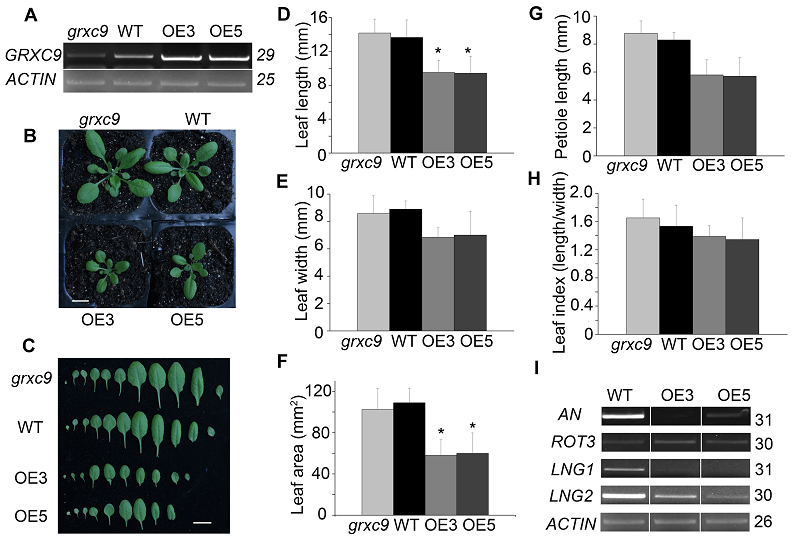
2.4 过表达GRXC9导致拟南芥叶片短小及叶片大小相关基因的表达前人的研究表明, 拟南芥谷氧还蛋白GRXC9基因参与激素SA/JA和JA/ET信号调控网络。基于此, 我们构建了35S:GRXC9表达载体, 以进一步探究GRXC9是否参与植物生长发育的调控。通过卡那霉素(Kanamy- cin, Kan)抗性筛选获得6株过表达植株。进一步的抗性筛选显示, 此6个株系的后代都出现3:1的分离比。T3代纯合株系的表达量检测结果显示, 6个株系中GRXC9表达量均明显高于野生型和突变体。选取其中表达量较高的2个株系(OE3和OE5)用于后续实验(图4A)。 图4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-550/img_4.png<b>图4</b> 拟南芥3种基因型植株的表型分析及叶片大小相关基因的表达<br/>(A) 3种基因型中<i>GRXC9</i>基因的表达水平检测; (B) 3种基因型植株生长28天的表型观察(Bar=1 cm); (C) 3种基因型代表性叶片的比较(Bar=1 cm); (D)-(H) 不同株系叶片长(D)、宽(E)、叶面积(F)、叶柄(G)和叶片系数(H)的统计分析(数据为平均值±标准误, <i>n</i>> 10, *表示差异显著(<i>P</i><0.05, <i>t</i>检验)); (I) 叶片大小相关基因的表达<br/><b>Figure 4</b> Phenotypic analysis and expression level of leaf size related genes in three genotypes of <i>Arabidopsis thaliana<br/></i>(A) Identification of <i>GRXC9</i> gene expression in <i>grxc9</i>,<i> 35S::GRXC9</i> and wild type; (B) The phenotype of three kinds of genotypes after 28 days grown under long-day conditions (Bar=1 cm); (C) The comparison of representative leaves among <i>grxc9</i>, <i>35S::GRXC9</i> and wild type (Bar=1 cm); (D)-(H) Statistical analysis of leaf blade length (D), width (E), area (F), leaf petiole length (G) and leaf index (H) among <i>grxc9</i>, two transgenic plants and wild type (Value=means±SE, <i>n</i> >10, * Student’s <i>t</i>-test significant difference compared with WT plants (<i>P</i><0.05) ); (I) Expression level of leaf size related genes Figure 4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-550/img_4.png<b>图4</b> 拟南芥3种基因型植株的表型分析及叶片大小相关基因的表达<br/>(A) 3种基因型中<i>GRXC9</i>基因的表达水平检测; (B) 3种基因型植株生长28天的表型观察(Bar=1 cm); (C) 3种基因型代表性叶片的比较(Bar=1 cm); (D)-(H) 不同株系叶片长(D)、宽(E)、叶面积(F)、叶柄(G)和叶片系数(H)的统计分析(数据为平均值±标准误, <i>n</i>> 10, *表示差异显著(<i>P</i><0.05, <i>t</i>检验)); (I) 叶片大小相关基因的表达<br/><b>Figure 4</b> Phenotypic analysis and expression level of leaf size related genes in three genotypes of <i>Arabidopsis thaliana<br/></i>(A) Identification of <i>GRXC9</i> gene expression in <i>grxc9</i>,<i> 35S::GRXC9</i> and wild type; (B) The phenotype of three kinds of genotypes after 28 days grown under long-day conditions (Bar=1 cm); (C) The comparison of representative leaves among <i>grxc9</i>, <i>35S::GRXC9</i> and wild type (Bar=1 cm); (D)-(H) Statistical analysis of leaf blade length (D), width (E), area (F), leaf petiole length (G) and leaf index (H) among <i>grxc9</i>, two transgenic plants and wild type (Value=means±SE, <i>n</i> >10, * Student’s <i>t</i>-test significant difference compared with WT plants (<i>P</i><0.05) ); (I) Expression level of leaf size related genes
图4 拟南芥3种基因型植株的表型分析及叶片大小相关基因的表达 (A) 3种基因型中GRXC9基因的表达水平检测; (B) 3种基因型植株生长28天的表型观察(Bar=1 cm); (C) 3种基因型代表性叶片的比较(Bar=1 cm); (D)-(H) 不同株系叶片长(D)、宽(E)、叶面积(F)、叶柄(G)和叶片系数(H)的统计分析(数据为平均值±标准误, n> 10, *表示差异显著(P<0.05, t检验)); (I) 叶片大小相关基因的表达 Figure 4 Phenotypic analysis and expression level of leaf size related genes in three genotypes of Arabidopsis thaliana (A) Identification of GRXC9 gene expression in grxc9, 35S::GRXC9 and wild type; (B) The phenotype of three kinds of genotypes after 28 days grown under long-day conditions (Bar=1 cm); (C) The comparison of representative leaves among grxc9, 35S::GRXC9 and wild type (Bar=1 cm); (D)-(H) Statistical analysis of leaf blade length (D), width (E), area (F), leaf petiole length (G) and leaf index (H) among grxc9, two transgenic plants and wild type (Value=means±SE, n >10, * Student’s t-test significant difference compared with WT plants (P<0.05) ); (I) Expression level of leaf size related genes
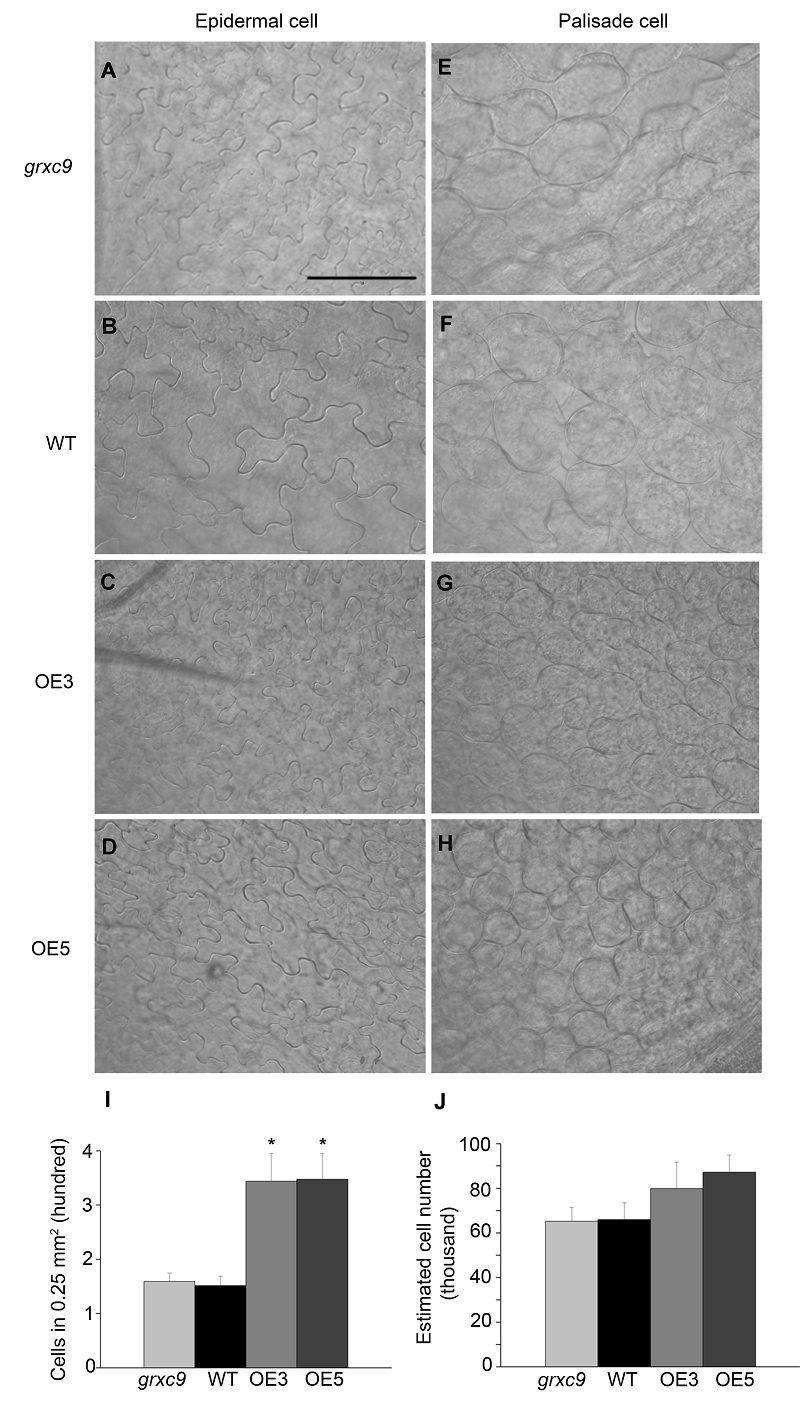
2.5 表皮细胞和栅栏细胞的观察叶片细胞的大小和数目是决定叶片大小的2个主要因素。为了确定GRXC9导致叶片短小是源于前者还是后者, 我们选取生长28天不同基因型植株的第5片莲座叶近轴端的中部相同位置, 进行脱色和透明等处理, 之后在微分干涉差显微镜下观察表皮细胞(图5A-D)和栅栏细胞(图5E-H)。从图5可以看出, 过表达株系表皮细胞的排列比野生型紧密, 突变体与野生型相比差异不显著; 而栅栏细胞在过表达株系中明显比野生型和突变体小。 图5https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-550/img_5.png<b>图5</b> 拟南芥3种基因型植株叶片的表皮细胞和栅栏细胞形态观察<br/>(A)-(D) 3种基因型植株叶片的表皮细胞形态观察; (E)-(H) 3种基因型植株叶片的栅栏细胞形态观察; (I) 每0.25 mm<sup>2</sup>叶片栅栏细胞的数目统计; (J) 估算的叶片第1层栅栏细胞总数(数据为平均值±标准误, <i>n</i> >10, *表示差异显著(<i>P</i><0.05, <i>t</i>检验))<br/><b>Figure 5</b> Morphological characterization of epidermal cells and palisade cells in three genotypes of <i>Arabidopsis thaliana<br/></i>(A)-(D) Morphological characterization of epidermal cells in <i>grxc9</i>, two transgenic plants and wild type; (E)-(H) Morphological characterization of palisade cells in <i>grxc9</i>, two transgenic plants and wild type; (I) The statistics of palisade cells in a 0.25 mm<sup>2</sup> area of <i>grxc9</i>, two transgenic plants and wild type; (J) Based on average cell number per 0.25 mm<sup>2</sup> area and leaf area, the total number of the first layer of palisade cells was estimated (Value=means±SE, <i>n</i> >10, * Student’s <i>t</i>-test significant difference compared with WT plants (<i>P</i><0.05)) Figure 5https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-550/img_5.png<b>图5</b> 拟南芥3种基因型植株叶片的表皮细胞和栅栏细胞形态观察<br/>(A)-(D) 3种基因型植株叶片的表皮细胞形态观察; (E)-(H) 3种基因型植株叶片的栅栏细胞形态观察; (I) 每0.25 mm<sup>2</sup>叶片栅栏细胞的数目统计; (J) 估算的叶片第1层栅栏细胞总数(数据为平均值±标准误, <i>n</i> >10, *表示差异显著(<i>P</i><0.05, <i>t</i>检验))<br/><b>Figure 5</b> Morphological characterization of epidermal cells and palisade cells in three genotypes of <i>Arabidopsis thaliana<br/></i>(A)-(D) Morphological characterization of epidermal cells in <i>grxc9</i>, two transgenic plants and wild type; (E)-(H) Morphological characterization of palisade cells in <i>grxc9</i>, two transgenic plants and wild type; (I) The statistics of palisade cells in a 0.25 mm<sup>2</sup> area of <i>grxc9</i>, two transgenic plants and wild type; (J) Based on average cell number per 0.25 mm<sup>2</sup> area and leaf area, the total number of the first layer of palisade cells was estimated (Value=means±SE, <i>n</i> >10, * Student’s <i>t</i>-test significant difference compared with WT plants (<i>P</i><0.05))
图5 拟南芥3种基因型植株叶片的表皮细胞和栅栏细胞形态观察 (A)-(D) 3种基因型植株叶片的表皮细胞形态观察; (E)-(H) 3种基因型植株叶片的栅栏细胞形态观察; (I) 每0.25 mm2叶片栅栏细胞的数目统计; (J) 估算的叶片第1层栅栏细胞总数(数据为平均值±标准误, n >10, *表示差异显著(P<0.05, t检验)) Figure 5 Morphological characterization of epidermal cells and palisade cells in three genotypes of Arabidopsis thaliana (A)-(D) Morphological characterization of epidermal cells in grxc9, two transgenic plants and wild type; (E)-(H) Morphological characterization of palisade cells in grxc9, two transgenic plants and wild type; (I) The statistics of palisade cells in a 0.25 mm2 area of grxc9, two transgenic plants and wild type; (J) Based on average cell number per 0.25 mm2 area and leaf area, the total number of the first layer of palisade cells was estimated (Value=means±SE, n >10, * Student’s t-test significant difference compared with WT plants (P<0.05))
BaiY, FalkS, SchnittgerA, JakobyMJ, HülskampM (2010). Tissue layer specific regulation of leaf length and width in Arabidopsis as revealed by the cell autonomous action of ANGUSTIFOLIA.Plant J 61, 191-199. [本文引用: 1]
[3]
BarM, OriN (2015). Compound leaf development in model plant species.Curr Opin Plant Biol 23, 61-69. [本文引用: 1]
[4]
BarkoulasM, GalinhaC, GriggSP, TsiantisM (2007). From genes to shape: regulatory interactions in leaf development.Curr Opin Plant Biol 10, 660-666. [本文引用: 1]
ChengNH, HirschiKD (2003). Cloning and characterization of CXIP1, a novel PICOT domain-containing Arabidopsis protein that associates with CAX1.J Biol Chem 278, 6503-6509. [本文引用: 1]
[7]
ChengNH, LiuJZ, BrockA, NelsonRS, HirschiKD (2006). AtGRXcp, an Arabidopsis chloroplastic glutare- doxin, is critical for protection against protein oxidative da- mage.J Biol Chem 281, 26280-26288. [本文引用: 2]
[8]
CloughSJ, BentAF (1998). Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidop- sis thaliana.Plant J 16, 735-743. [本文引用: 1]
[9]
DaviesPJ (1995). The plant hormones: their nature, occurr- ence, and functions. In: Davies PJ, ed. Plant Hormones. Berlin: Springer. pp. 1-12. [本文引用: 1]
[10]
FolkersU, KirikV, SchöbingerU, FalkS, KrishnakumarS, PollockMA, OppenheimerDG, DayI, ReddyAR, Jür- gensG, HülskampM (2002). The cell morphogenesis gene ANGUSTIFOLIA encodes a CtBP/BARS-like protein and is involved in the control of the microtubule cytosk- eleton.EMBO J 21, 1280-1288. [本文引用: 2]
[11]
HansonJ, JohannessonH, EngströmP (2001). Sugar- dependent alterations in cotyledon and leaf development in transgenic plants expressing the HDZhdip gene ATHB- 13.Plant Mol Biol 45, 247-262. [本文引用: 1]
[12]
Herrera-VásquezA, CarvalloL, BlancoF, TobarM, Villarroel-CandiaE, Vicente-CarbajosaJ, SalinasP, HoluigueL (2015). Transcriptional control of glutaredoxin GRXC9 expression by a salicylic acid-dependent and NPR1-independent pathway in Arabidopsis.Plant Mol Biol Rep 33, 624-637. [本文引用: 2]
[13]
HolmgrenA (1976). Hydrogen donor system for Escherichia coli ribonucleoside-diphosphate reductase dependent up- on glutathione.Proc Natl Acad Sci USA 73, 2275-2279. [本文引用: 2]
[14]
HolmgrenA (1979a). Glutathione-dependent synthesis of deoxyribonucleotides. Characterization of the enzymatic mechanism of Escherichia coli glutaredoxin.J Biol Chem 254, 3672-3678. [本文引用: 1]
[15]
HolmgrenA (1979b). Glutathione-dependent synthesis of deoxyribonucleotides. Purification and characterization of glutaredoxin from Escherichia coli.J Biol Chem 254, 3664-3671. [本文引用: 1]
[16]
HoriguchiG, KimGT, TsukayaH (2005). The transcription factor AtGRF5 and the transcription coactivator AN3 re- gulate cell proliferation in leaf primordia of Arabidopsis tha- liana.Plant J 43, 68-78. [本文引用: 2]
[17]
KimGT, ShodaK, TsugeT, ChoKH, UchimiyaH, YokoyamaR, NishitaniK, TsukayaH (2002). The ANGUSTIFOLIA gene of Arabidopsis, a plant CtBP gene, regulates leaf-cell expansion, the arrangement of cortical mi- crotubules in leaf cells and expression of a gene involved in cell-wall formation.EMBO J 21, 1267-1279. [本文引用: 3]
[18]
KimGT, TsukayaH, UchimiyaH (1998). The ROTUNDI- FOLIA3 gene of Arabidopsis thaliana encodes a new member of the cytochrome P-450 family that is required for the regulated polar elongation of leaf cells.Genes Dev 12, 2381-2391. [本文引用: 2]
[19]
KimJH, KendeH (2004). A transcriptional coactivator, AtGIF1, is involved in regulating leaf growth and morpho- logy in Arabidopsis.Proc Natl Acad Sci USA 101, 13374-13379. [本文引用: 2]
LaporteD, OlateE, SalinasP, SalazarM, JordanaX, HoluigueL (2012). Glutaredoxin GRXS13 plays a key role in protection against photooxidative stress in Arabidopsis.J Exp Bot 63, 503-515. [本文引用: 1]
[22]
LeeYK, KimGT, KimIJ, ParkJ, KwakSS, ChoiG, ChungWI (2006). LONGIFOLIA1 and LONGIFOLIA2, two homologous genes, regulate longitudinal cell elongation in Arabidopsis.Development 133, 4305-4314. [本文引用: 2]
[23]
LemaireSD (2004). The glutaredoxin family in oxygenic photosynthetic organisms.Photosynth Res 79, 305-318. [本文引用: 1]
MeyerY, BuchananBB, VignolsF, ReichheldJP (2009). Thioredoxins and glutaredoxins: unifying elements in redox biology.Annu Rev Genet 43, 335-367. [本文引用: 1]
[26]
MizukamiY (2001). A matter of size: developmental control of organ size in plants.Curr Opin Plant Biol 4, 533-539. [本文引用: 2]
[27]
MizukamiY, FischerRL (2000). Plant organ size control: AINTEGUMENTA regulates growth and cell numbers during organogenesis.Proc Natl Acad Sci USA 97, 942-947.
[28]
MurmuJ, BushMJ, DeLongC, LiST, XuML, KhanM, MalcolmsonC, FobertPR, ZachgoS, HepworthSR (2010). Arabidopsis basic leucine-zipper transcription factors TGA9 and TGA10 interact with floral glutaredoxins ROXY1 and ROXY2 and are redundantly required for anther development.Plant Physiol 154, 1492-1504. [本文引用: 1]
[29]
NaritaNN, MooreS, HoriguchiG, KuboM, DemuraT, FukudaH, GoodrichJ, TsukayaH (2004). Overexpres- sion of a novel small peptide ROTUNDIFOLIA4 decre- ases cell proliferation and alters leaf shape in Arabidopsis thaliana.Plant J 38, 699-713. [本文引用: 1]
[30]
NdamukongI, Al AbdallatA, ThurowC, FodeB, ZanderM, WeigelR, GatzC (2007). SA-inducible Arabidopsis glutaredoxin interacts with TGA factors and suppresses JA-responsive PDF1.2 transcription.Plant J 50, 128-139. [本文引用: 2]
[31]
QiuJL, JilkR, MarksMD, SzymanskiDB (2002). The Ara- bidopsis SPIKE1 gene is required for normal cell shape control and tissue development.Plant Cell 14, 101-118. [本文引用: 2]
RouhierN, GelhayeE, JacquotJP (2004). Plant glutaredoxins: still mysterious reducing systems.Cell Mol Life Sci 61, 1266-1277. [本文引用: 1]
[34]
RouhierN, LemaireSD, JacquotJP (2008). The role of glutathione in photosynthetic organisms: emerging functions for glutaredoxins and glutathionylation.Annu Rev Plant Biol 59, 143-166. [本文引用: 1]
[35]
Sugimoto-ShirasuK, RobertsK (2003). “Big it up”: endoreduplication and cell-size control in plants.Curr Opin Plant Biol 6, 544-553. [本文引用: 1]
[36]
TsugeT, TsukayaH, UchimiyaH (1996). Two independent and polarized processes of cell elongation regulate leaf blade expansion in Arabidopsis thaliana (L) Heynh.Development 122, 1589-1600. [本文引用: 2]
[37]
TsukayaH (2005). Leaf shape: genetic controls and envi- ronmental factors.Inter J Dev Biol 49, 547-555. [本文引用: 1]
VermaPK, VermaS, PandeV, MallickS, TripathiRD, DhankherOP, ChakrabartyD (2016). Overexpression of rice glutaredoxin OsGrx_C7 and OsGrx_C2.1 reduces intracellular arsenic accumulation and increases tolerance in Arabidopsis thaliana.Front Plant Sci 7, 740. [本文引用: 1]
[40]
VernouxT, AutranD, TraasJ (2000). Developmental control of cell division patterns in the shoot apex. In: Inzé D, ed. The Plant Cell Cycle. Dordrecht: Springer. pp. 25-37. [本文引用: 1]
[41]
XingS, LauriA, ZachgoS (2006). Redox regulation and flower development: a novel function for glutaredoxins.Plant Biol 8, 547-555. [本文引用: 1]
[42]
XingSP, RossoMG, ZachgoS (2005). ROXY1, a member of the plant glutaredoxin family, is required for petal deve- lopment in Arabidopsis thaliana.Development 132, 1555-1565. [本文引用: 2]
[43]
XingSP, ZachgoS (2008). ROXY1 and ROXY2, two Arabidopsis glutaredoxin genes, are required for anther development.Plant J 53, 790-801. [本文引用: 1]
[44]
YooSD, ChoYH, SheenJ (2007). Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis.Nat Protoc 2, 1565-1572. [本文引用: 1]
[45]
ZanderM, ChenSX, ImkampeJ, ThurowC, GatzC (2012). Repression of the Arabidopsis thaliana jasmonic acid/ethylene-induced defense pathway by TGA-interac- ting glutaredoxins depends on their C-terminal ALWL motif.Mol Plant 5, 831-840. [本文引用: 2]
[46]
ZhangSD, ShenZJ, LiuXM, LiZ, ZhangQW, LiuXX (2016). Molecular identification of three novel glutaredoxin genes that play important roles in antioxidant defense in Helicoverpa armigera.Insect Biochem Mol Biol 75, 107-116.
非洲菊微管相关蛋白基因GMAP65-1功能分析 1 2015
... 植物器官的发育依赖于细胞的分裂、扩张及分化, 器官的最终大小由细胞数目和细胞大小决定(Mizu- kami, 2001; 李凌飞等, 2015).细胞分裂在植物器官发育过程中起重要作用(Vernoux et al., 2000).细胞周期调控因子CDKA和CycD3与一些基因(如拟南芥ANT基因)协同表达调控细胞分裂, 进而影响器官的发育(Mizukami and Fischer, 2000 ).细胞扩张与细胞分裂相比影响器官发育的方式更为复杂.植物细胞的最终大小与其倍性及核内复制相关, 但是具体分子机制尚不清楚(Kondorosi et al., 2000; Sugimoto- Shirasu and Roberts, 2003).植物一些器官(尤其是叶片)的大小和形状由极性细胞的扩张决定.例如, 拟南芥an突变体叶片较窄、厚, 原因是决定叶片宽度的细胞扩张受损(Kim et al., 2002); 拟南芥rot3突变体呈现出叶片较宽的表型, 是由于决定叶片长度的细胞扩张受损(Kim et al., 1998).此外, 细胞的伸长和扩张还受到赤霉素(GA)与油菜素内酯(BR)等多种激素的调控(Davies, 1995), 它们可能相互作用共同调控植物的器官发育和大小(Mizukami, 2001).综上, GRXC9基因可能通过抑制控制细胞扩张的基因AN以及调控叶形态的基因LNG1和LNG2的表达使叶片短小, 对拟南芥叶片的发育起负调控作用.然而, GRXC9是否也受到GA和BR等多种激素的调控, 以及是否有上游转录因子对其功能进行调节等均有待进一步研究. ...
1 2010
... 前人运用基因组和表达序列数据库等手段, 对拟南芥GRX基因家族进行了研究, 发现GRX家族的很多基因参与了植物的生长发育过程(Lemaire, 2004; Rouhier et al., 2004; Xing et al., 2006).本研究表明, GRXC9基因过表达株系叶片出现短小的表型, 且叶片短小的原因是由于叶片细胞体积变小.进一步检测叶片大小相关基因的表达情况, 发现过表达株系中AN、LNG1和LNG2的表达量明显下降.Tsuge等(1996)报道了AN基因通过调控特异的极性细胞伸长控制叶的形态.Kim等(2002)的研究发现, AN不仅控制叶细胞的极性伸长, 而且还可能通过调控细胞壁形成相关基因参与叶形态的调控.同时, AN是以一种非自主的模式控制细胞分裂(Bai et al., 2010).Lee等(2006)报道了LNG1和LNG2基因突变体叶片短小, 两基因共同参与叶片长度的调控.GRXC9基因过表达株系叶片短小可能是通过抑制参与叶形态调控基因的表达实现.之前有报道显示, GRXC9参与SA、JA和ET等多种激素调控途径, 在植物对外界环境的响应中发挥作用, 但具体机制尚不清楚(Ndamukong et al., 2007; Zander et al., 2012; Herrera-Vásquez et al., 2015).本研究表明, GRXC9可能通过影响AN、LNG1和LNG2等基因的表达参与叶片的发育, 但此过程是否与激素SA/JA和JA/ET的信号调控网络有关, 目前尚不明确. ...
1 2015
... 谷氧还蛋白家族基因在植物发育方面的作用已有报道, 但在植物叶片形态大小方面的研究尚少.叶片形成需要经历形态发生和组织分化两个重要过程(Bar and Ori, 2015).叶片器官形态发生可分为3个阶段: 起始阶段、叶片极性的建立和细胞扩张(Tsukaya, 2005, 2006).这3个阶段受不同基因的调控(Byrne, 2005; Barkoulas et al., 2007).AN3/AtGIF1和At- GRF5的功能缺失突变体出现窄叶表型, 该表型由叶片细胞数目变少引起(Kim and Kende, 2004; Horigu- chi et al., 2005).与野生型相比, 过表达AN3和AtGRF5的植株叶片明显变大20%-30% (Horiguchi et al., 2005).在细胞扩张阶段, ROT3和ROT4基因参与调控叶片长度, rot3突变体表现出叶片发育畸形, ROT3过表达植株表现出叶片变长, 但宽度没有变化(Tsuge et al., 1996; Kim et al., 1998).ROT4基因编码膜结合的短肽, 其突变体的表型与rot3类似, 但叶片短小是由细胞增殖减少引起(Narita et al., 2004).LNG1和LNG2也参与调控叶片长度, 其突变体叶片短小(Lee et al., 2006).还有一些基因, 如AN蛋白基因参与叶片细胞的扩张(Folkers et al., 2002); SPIK- E1参与细胞骨架的构建, 最终决定细胞的形状和组织发育(Qiu et al., 2002).AN、SPIKE1、AN3、AtGIF1和AtHB13都参与调控叶片宽度, 且突变体叶片宽度都比较小(Hanson et al., 2001; Folkers et al., 2002; Kim et al., 2002; Qiu et al., 2002; Kim and Kende, 2004). ...
1 2007
... 谷氧还蛋白家族基因在植物发育方面的作用已有报道, 但在植物叶片形态大小方面的研究尚少.叶片形成需要经历形态发生和组织分化两个重要过程(Bar and Ori, 2015).叶片器官形态发生可分为3个阶段: 起始阶段、叶片极性的建立和细胞扩张(Tsukaya, 2005, 2006).这3个阶段受不同基因的调控(Byrne, 2005; Barkoulas et al., 2007).AN3/AtGIF1和At- GRF5的功能缺失突变体出现窄叶表型, 该表型由叶片细胞数目变少引起(Kim and Kende, 2004; Horigu- chi et al., 2005).与野生型相比, 过表达AN3和AtGRF5的植株叶片明显变大20%-30% (Horiguchi et al., 2005).在细胞扩张阶段, ROT3和ROT4基因参与调控叶片长度, rot3突变体表现出叶片发育畸形, ROT3过表达植株表现出叶片变长, 但宽度没有变化(Tsuge et al., 1996; Kim et al., 1998).ROT4基因编码膜结合的短肽, 其突变体的表型与rot3类似, 但叶片短小是由细胞增殖减少引起(Narita et al., 2004).LNG1和LNG2也参与调控叶片长度, 其突变体叶片短小(Lee et al., 2006).还有一些基因, 如AN蛋白基因参与叶片细胞的扩张(Folkers et al., 2002); SPIK- E1参与细胞骨架的构建, 最终决定细胞的形状和组织发育(Qiu et al., 2002).AN、SPIKE1、AN3、AtGIF1和AtHB13都参与调控叶片宽度, 且突变体叶片宽度都比较小(Hanson et al., 2001; Folkers et al., 2002; Kim et al., 2002; Qiu et al., 2002; Kim and Kende, 2004). ...
1 2005
... 谷氧还蛋白家族基因在植物发育方面的作用已有报道, 但在植物叶片形态大小方面的研究尚少.叶片形成需要经历形态发生和组织分化两个重要过程(Bar and Ori, 2015).叶片器官形态发生可分为3个阶段: 起始阶段、叶片极性的建立和细胞扩张(Tsukaya, 2005, 2006).这3个阶段受不同基因的调控(Byrne, 2005; Barkoulas et al., 2007).AN3/AtGIF1和At- GRF5的功能缺失突变体出现窄叶表型, 该表型由叶片细胞数目变少引起(Kim and Kende, 2004; Horigu- chi et al., 2005).与野生型相比, 过表达AN3和AtGRF5的植株叶片明显变大20%-30% (Horiguchi et al., 2005).在细胞扩张阶段, ROT3和ROT4基因参与调控叶片长度, rot3突变体表现出叶片发育畸形, ROT3过表达植株表现出叶片变长, 但宽度没有变化(Tsuge et al., 1996; Kim et al., 1998).ROT4基因编码膜结合的短肽, 其突变体的表型与rot3类似, 但叶片短小是由细胞增殖减少引起(Narita et al., 2004).LNG1和LNG2也参与调控叶片长度, 其突变体叶片短小(Lee et al., 2006).还有一些基因, 如AN蛋白基因参与叶片细胞的扩张(Folkers et al., 2002); SPIK- E1参与细胞骨架的构建, 最终决定细胞的形状和组织发育(Qiu et al., 2002).AN、SPIKE1、AN3、AtGIF1和AtHB13都参与调控叶片宽度, 且突变体叶片宽度都比较小(Hanson et al., 2001; Folkers et al., 2002; Kim et al., 2002; Qiu et al., 2002; Kim and Kende, 2004). ...
1 2003
... 拟南芥GRX家族的很多基因已被广泛研究, 它们在植物发育、激素信号途径以及抗逆反应等多个方面发挥重要作用.GRXC7/ROXY1基因参与花瓣的发育, 其突变体表现出花瓣和花原基异常的表型(Xing et al., 2005).GRXC8/ROXY2与GRXC7功能冗余, 且其可与TGA因子互作参与花药的发育(Murmu et al., 2010).表型观察发现, roxy1和roxy2单突变体花药发育正常, roxy1/roxy2双突变体花药发育存在缺陷.Ndamukong等(2007)的研究表明, GRXC9可与TGA转录因子互作, 受水杨酸(SA)诱导, 抑制茉莉酸(JA)途径的转录因子PDF1.2的转录, 推测GRXC9可能参与激素SA/JA的信号调控网络.之后, Zander等(2012)报道GRXC9与TGA因子互作能抑制JA/ET (乙烯)诱导的信号通路; 通过GRXC9的异位表达, 他们发现其负调控JA/ET信号途径的基因表达, 但具体机制还不十分清楚.GRXS13参与抗光氧化胁迫, 过表达GRXS13的植株在MV和高光处理后损伤程度比野生型有所减弱, 说明GRXS13的表达使ROS受到限制, 其可能参与拟南芥细胞内ROS的清除和抗光氧化的调控网络(Laporte et al., 2012).GRXS14通过与CAX1的N端互作, 参与调控Ca2+的转运(Cheng and Hirschi, 2003)和氧化胁迫(Cheng et al., 2006).Zhang等(2016)报道了棉铃虫中谷氧还蛋白HaGrx、 HaGrx3和HaGrx5受不同温度胁迫及过氧化氢(H2O2)的诱导; 进一步检测发现, 敲减植株中超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)、抗坏血酸和总谷胱甘肽(total GSH)等含量升高, 说明HaGrx、HaGrx3和HaGrx5具有抗氧化防御的功能.另外, Verma等(2016)发现水稻(Oryza sativa)中谷氧还蛋白OsGrx_C7以及OsGrx_C2.1对重金属砷敏感, 转化拟南芥的过表达植株表现出对砷的抗性增强, 说明在谷氧还蛋白家族中有些基因参与砷胁迫应答反应. ...
2 2006
... 拟南芥GRX家族的很多基因已被广泛研究, 它们在植物发育、激素信号途径以及抗逆反应等多个方面发挥重要作用.GRXC7/ROXY1基因参与花瓣的发育, 其突变体表现出花瓣和花原基异常的表型(Xing et al., 2005).GRXC8/ROXY2与GRXC7功能冗余, 且其可与TGA因子互作参与花药的发育(Murmu et al., 2010).表型观察发现, roxy1和roxy2单突变体花药发育正常, roxy1/roxy2双突变体花药发育存在缺陷.Ndamukong等(2007)的研究表明, GRXC9可与TGA转录因子互作, 受水杨酸(SA)诱导, 抑制茉莉酸(JA)途径的转录因子PDF1.2的转录, 推测GRXC9可能参与激素SA/JA的信号调控网络.之后, Zander等(2012)报道GRXC9与TGA因子互作能抑制JA/ET (乙烯)诱导的信号通路; 通过GRXC9的异位表达, 他们发现其负调控JA/ET信号途径的基因表达, 但具体机制还不十分清楚.GRXS13参与抗光氧化胁迫, 过表达GRXS13的植株在MV和高光处理后损伤程度比野生型有所减弱, 说明GRXS13的表达使ROS受到限制, 其可能参与拟南芥细胞内ROS的清除和抗光氧化的调控网络(Laporte et al., 2012).GRXS14通过与CAX1的N端互作, 参与调控Ca2+的转运(Cheng and Hirschi, 2003)和氧化胁迫(Cheng et al., 2006).Zhang等(2016)报道了棉铃虫中谷氧还蛋白HaGrx、 HaGrx3和HaGrx5受不同温度胁迫及过氧化氢(H2O2)的诱导; 进一步检测发现, 敲减植株中超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)、抗坏血酸和总谷胱甘肽(total GSH)等含量升高, 说明HaGrx、HaGrx3和HaGrx5具有抗氧化防御的功能.另外, Verma等(2016)发现水稻(Oryza sativa)中谷氧还蛋白OsGrx_C7以及OsGrx_C2.1对重金属砷敏感, 转化拟南芥的过表达植株表现出对砷的抗性增强, 说明在谷氧还蛋白家族中有些基因参与砷胁迫应答反应. ... ... 基因精确的时空表达是其正常行使功能的基础与前提, 基因在不同时间空间的表达差异暗示了其功能的不同.因此, 研究GRXC9基因的时空表达, 将为探明该基因的功能提供重要线索.目前, 对GRX基因家族表达部位的研究较少.已有的研究表明, CC型的RO- XY1 (GRXC7)基因在花序、根以及果荚中强烈表达, 而在茎叶中不表达(Xing et al., 2005). ROXY2 (GR- XC8)基因在花原基和分生组织中表达较少, 而在花药发育时期表达量最高, 这与其参与雄蕊发育的功能相对应(Xing and Zachgo, 2008).GRXS14是组成型表达基因, 其在叶片、维管束、根、茎和花中均有表达, 但在叶片中的表达量最高(Cheng et al., 2006).通过RT-PCR分析及构建GUS启动子融合表达载体两种手段, 我们研究了GRXC9的时空表达模式, 结果发现GRXC9基因的表达不具有组织特异性, 其在植物根、茎、叶、花和角果中均有表达, 且在花中的表达主要是花丝和花药部位, 暗示GRXC9基因可能参与雄蕊的生长发育. ...
1 1998
... 采用热激法将构建好的过表达载体导入农杆菌EHA105感受态.然后, 采用农杆菌介导的花粉管导入法(floral-dip) (Clough and Bent, 1998)转化拟南芥野生型植株.经卡那霉素抗性筛选后获得转基因纯合株系, 将其与同一批收获的野生型种子播种在同一个MS培养基平板上, 在相同的培养环境中观察表型. ...
1 1995
... 植物器官的发育依赖于细胞的分裂、扩张及分化, 器官的最终大小由细胞数目和细胞大小决定(Mizu- kami, 2001; 李凌飞等, 2015).细胞分裂在植物器官发育过程中起重要作用(Vernoux et al., 2000).细胞周期调控因子CDKA和CycD3与一些基因(如拟南芥ANT基因)协同表达调控细胞分裂, 进而影响器官的发育(Mizukami and Fischer, 2000 ).细胞扩张与细胞分裂相比影响器官发育的方式更为复杂.植物细胞的最终大小与其倍性及核内复制相关, 但是具体分子机制尚不清楚(Kondorosi et al., 2000; Sugimoto- Shirasu and Roberts, 2003).植物一些器官(尤其是叶片)的大小和形状由极性细胞的扩张决定.例如, 拟南芥an突变体叶片较窄、厚, 原因是决定叶片宽度的细胞扩张受损(Kim et al., 2002); 拟南芥rot3突变体呈现出叶片较宽的表型, 是由于决定叶片长度的细胞扩张受损(Kim et al., 1998).此外, 细胞的伸长和扩张还受到赤霉素(GA)与油菜素内酯(BR)等多种激素的调控(Davies, 1995), 它们可能相互作用共同调控植物的器官发育和大小(Mizukami, 2001).综上, GRXC9基因可能通过抑制控制细胞扩张的基因AN以及调控叶形态的基因LNG1和LNG2的表达使叶片短小, 对拟南芥叶片的发育起负调控作用.然而, GRXC9是否也受到GA和BR等多种激素的调控, 以及是否有上游转录因子对其功能进行调节等均有待进一步研究. ...
2 2002
... 谷氧还蛋白家族基因在植物发育方面的作用已有报道, 但在植物叶片形态大小方面的研究尚少.叶片形成需要经历形态发生和组织分化两个重要过程(Bar and Ori, 2015).叶片器官形态发生可分为3个阶段: 起始阶段、叶片极性的建立和细胞扩张(Tsukaya, 2005, 2006).这3个阶段受不同基因的调控(Byrne, 2005; Barkoulas et al., 2007).AN3/AtGIF1和At- GRF5的功能缺失突变体出现窄叶表型, 该表型由叶片细胞数目变少引起(Kim and Kende, 2004; Horigu- chi et al., 2005).与野生型相比, 过表达AN3和AtGRF5的植株叶片明显变大20%-30% (Horiguchi et al., 2005).在细胞扩张阶段, ROT3和ROT4基因参与调控叶片长度, rot3突变体表现出叶片发育畸形, ROT3过表达植株表现出叶片变长, 但宽度没有变化(Tsuge et al., 1996; Kim et al., 1998).ROT4基因编码膜结合的短肽, 其突变体的表型与rot3类似, 但叶片短小是由细胞增殖减少引起(Narita et al., 2004).LNG1和LNG2也参与调控叶片长度, 其突变体叶片短小(Lee et al., 2006).还有一些基因, 如AN蛋白基因参与叶片细胞的扩张(Folkers et al., 2002); SPIK- E1参与细胞骨架的构建, 最终决定细胞的形状和组织发育(Qiu et al., 2002).AN、SPIKE1、AN3、AtGIF1和AtHB13都参与调控叶片宽度, 且突变体叶片宽度都比较小(Hanson et al., 2001; Folkers et al., 2002; Kim et al., 2002; Qiu et al., 2002; Kim and Kende, 2004). ... ... ; Folkers et al., 2002; Kim et al., 2002; Qiu et al., 2002; Kim and Kende, 2004). ...
1 2001
... 谷氧还蛋白家族基因在植物发育方面的作用已有报道, 但在植物叶片形态大小方面的研究尚少.叶片形成需要经历形态发生和组织分化两个重要过程(Bar and Ori, 2015).叶片器官形态发生可分为3个阶段: 起始阶段、叶片极性的建立和细胞扩张(Tsukaya, 2005, 2006).这3个阶段受不同基因的调控(Byrne, 2005; Barkoulas et al., 2007).AN3/AtGIF1和At- GRF5的功能缺失突变体出现窄叶表型, 该表型由叶片细胞数目变少引起(Kim and Kende, 2004; Horigu- chi et al., 2005).与野生型相比, 过表达AN3和AtGRF5的植株叶片明显变大20%-30% (Horiguchi et al., 2005).在细胞扩张阶段, ROT3和ROT4基因参与调控叶片长度, rot3突变体表现出叶片发育畸形, ROT3过表达植株表现出叶片变长, 但宽度没有变化(Tsuge et al., 1996; Kim et al., 1998).ROT4基因编码膜结合的短肽, 其突变体的表型与rot3类似, 但叶片短小是由细胞增殖减少引起(Narita et al., 2004).LNG1和LNG2也参与调控叶片长度, 其突变体叶片短小(Lee et al., 2006).还有一些基因, 如AN蛋白基因参与叶片细胞的扩张(Folkers et al., 2002); SPIK- E1参与细胞骨架的构建, 最终决定细胞的形状和组织发育(Qiu et al., 2002).AN、SPIKE1、AN3、AtGIF1和AtHB13都参与调控叶片宽度, 且突变体叶片宽度都比较小(Hanson et al., 2001; Folkers et al., 2002; Kim et al., 2002; Qiu et al., 2002; Kim and Kende, 2004). ...
2 2015
... 前人的研究表明, 拟南芥谷氧还蛋白GRXC9基因参与激素SA/JA和JA/ET的信号调控网络(Ndamu- kong et al., 2007; Zander et al., 2012; Herrera- Vásquez et al., 2015).本研究通过探讨GRX的时空表达模式、亚细胞定位及对植物生长发育的调控, 发现其具有负调控叶片大小的功能.研究结果丰富了人们对GRXC9基因功能的认识, 拓宽了谷氧还蛋白家族在植物叶片发育方面的研究价值. ... ... 前人运用基因组和表达序列数据库等手段, 对拟南芥GRX基因家族进行了研究, 发现GRX家族的很多基因参与了植物的生长发育过程(Lemaire, 2004; Rouhier et al., 2004; Xing et al., 2006).本研究表明, GRXC9基因过表达株系叶片出现短小的表型, 且叶片短小的原因是由于叶片细胞体积变小.进一步检测叶片大小相关基因的表达情况, 发现过表达株系中AN、LNG1和LNG2的表达量明显下降.Tsuge等(1996)报道了AN基因通过调控特异的极性细胞伸长控制叶的形态.Kim等(2002)的研究发现, AN不仅控制叶细胞的极性伸长, 而且还可能通过调控细胞壁形成相关基因参与叶形态的调控.同时, AN是以一种非自主的模式控制细胞分裂(Bai et al., 2010).Lee等(2006)报道了LNG1和LNG2基因突变体叶片短小, 两基因共同参与叶片长度的调控.GRXC9基因过表达株系叶片短小可能是通过抑制参与叶形态调控基因的表达实现.之前有报道显示, GRXC9参与SA、JA和ET等多种激素调控途径, 在植物对外界环境的响应中发挥作用, 但具体机制尚不清楚(Ndamukong et al., 2007; Zander et al., 2012; Herrera-Vásquez et al., 2015).本研究表明, GRXC9可能通过影响AN、LNG1和LNG2等基因的表达参与叶片的发育, 但此过程是否与激素SA/JA和JA/ET的信号调控网络有关, 目前尚不明确. ...
2 1976
... 谷氧还蛋白(glutaredoxin, GRX)是一类谷胱甘肽(glutathione, GSH)依赖的氧化还原酶, 最早被发现可恢复大肠杆菌硫氧还蛋白(TRX)突变体的生长发育, 且在二硫键形成过程中能循环催化核苷酸还原酶(Holmgren, 1976, 1979a, 1979b).研究表明, GRX在人类健康、菌类及植物中均发挥重要作用(Holm- gren, 1976; Lillig et al., 2008).拟南芥(Arabidopsis thaliana)中GRX家族成员共有31个(Rouhier et al., 2006).根据它们的活性位点序列可分为3类, 即CP- YC型、CGFS型和CC型.其中CC型也被称作ROXY型, 主要存在于高等植物中.拟南芥中大多数GRX (21个)为CC类型(Rouhier et al., 2008), 此外, 6个为CPYC型, 4个为CGFS型(Meyer et al., 2009). ... ... ).研究表明, GRX在人类健康、菌类及植物中均发挥重要作用(Holm- gren, 1976; Lillig et al., 2008).拟南芥(Arabidopsis thaliana)中GRX家族成员共有31个(Rouhier et al., 2006).根据它们的活性位点序列可分为3类, 即CP- YC型、CGFS型和CC型.其中CC型也被称作ROXY型, 主要存在于高等植物中.拟南芥中大多数GRX (21个)为CC类型(Rouhier et al., 2008), 此外, 6个为CPYC型, 4个为CGFS型(Meyer et al., 2009). ...
1 1979
... 谷氧还蛋白(glutaredoxin, GRX)是一类谷胱甘肽(glutathione, GSH)依赖的氧化还原酶, 最早被发现可恢复大肠杆菌硫氧还蛋白(TRX)突变体的生长发育, 且在二硫键形成过程中能循环催化核苷酸还原酶(Holmgren, 1976, 1979a, 1979b).研究表明, GRX在人类健康、菌类及植物中均发挥重要作用(Holm- gren, 1976; Lillig et al., 2008).拟南芥(Arabidopsis thaliana)中GRX家族成员共有31个(Rouhier et al., 2006).根据它们的活性位点序列可分为3类, 即CP- YC型、CGFS型和CC型.其中CC型也被称作ROXY型, 主要存在于高等植物中.拟南芥中大多数GRX (21个)为CC类型(Rouhier et al., 2008), 此外, 6个为CPYC型, 4个为CGFS型(Meyer et al., 2009). ...
1 1979
... 谷氧还蛋白(glutaredoxin, GRX)是一类谷胱甘肽(glutathione, GSH)依赖的氧化还原酶, 最早被发现可恢复大肠杆菌硫氧还蛋白(TRX)突变体的生长发育, 且在二硫键形成过程中能循环催化核苷酸还原酶(Holmgren, 1976, 1979a, 1979b).研究表明, GRX在人类健康、菌类及植物中均发挥重要作用(Holm- gren, 1976; Lillig et al., 2008).拟南芥(Arabidopsis thaliana)中GRX家族成员共有31个(Rouhier et al., 2006).根据它们的活性位点序列可分为3类, 即CP- YC型、CGFS型和CC型.其中CC型也被称作ROXY型, 主要存在于高等植物中.拟南芥中大多数GRX (21个)为CC类型(Rouhier et al., 2008), 此外, 6个为CPYC型, 4个为CGFS型(Meyer et al., 2009). ...
2 2005
... 谷氧还蛋白家族基因在植物发育方面的作用已有报道, 但在植物叶片形态大小方面的研究尚少.叶片形成需要经历形态发生和组织分化两个重要过程(Bar and Ori, 2015).叶片器官形态发生可分为3个阶段: 起始阶段、叶片极性的建立和细胞扩张(Tsukaya, 2005, 2006).这3个阶段受不同基因的调控(Byrne, 2005; Barkoulas et al., 2007).AN3/AtGIF1和At- GRF5的功能缺失突变体出现窄叶表型, 该表型由叶片细胞数目变少引起(Kim and Kende, 2004; Horigu- chi et al., 2005).与野生型相比, 过表达AN3和AtGRF5的植株叶片明显变大20%-30% (Horiguchi et al., 2005).在细胞扩张阶段, ROT3和ROT4基因参与调控叶片长度, rot3突变体表现出叶片发育畸形, ROT3过表达植株表现出叶片变长, 但宽度没有变化(Tsuge et al., 1996; Kim et al., 1998).ROT4基因编码膜结合的短肽, 其突变体的表型与rot3类似, 但叶片短小是由细胞增殖减少引起(Narita et al., 2004).LNG1和LNG2也参与调控叶片长度, 其突变体叶片短小(Lee et al., 2006).还有一些基因, 如AN蛋白基因参与叶片细胞的扩张(Folkers et al., 2002); SPIK- E1参与细胞骨架的构建, 最终决定细胞的形状和组织发育(Qiu et al., 2002).AN、SPIKE1、AN3、AtGIF1和AtHB13都参与调控叶片宽度, 且突变体叶片宽度都比较小(Hanson et al., 2001; Folkers et al., 2002; Kim et al., 2002; Qiu et al., 2002; Kim and Kende, 2004). ... ... 的植株叶片明显变大20%-30% (Horiguchi et al., 2005).在细胞扩张阶段, ROT3和ROT4基因参与调控叶片长度, rot3突变体表现出叶片发育畸形, ROT3过表达植株表现出叶片变长, 但宽度没有变化(Tsuge et al., 1996; Kim et al., 1998).ROT4基因编码膜结合的短肽, 其突变体的表型与rot3类似, 但叶片短小是由细胞增殖减少引起(Narita et al., 2004).LNG1和LNG2也参与调控叶片长度, 其突变体叶片短小(Lee et al., 2006).还有一些基因, 如AN蛋白基因参与叶片细胞的扩张(Folkers et al., 2002); SPIK- E1参与细胞骨架的构建, 最终决定细胞的形状和组织发育(Qiu et al., 2002).AN、SPIKE1、AN3、AtGIF1和AtHB13都参与调控叶片宽度, 且突变体叶片宽度都比较小(Hanson et al., 2001; Folkers et al., 2002; Kim et al., 2002; Qiu et al., 2002; Kim and Kende, 2004). ...
3 2002
... 谷氧还蛋白家族基因在植物发育方面的作用已有报道, 但在植物叶片形态大小方面的研究尚少.叶片形成需要经历形态发生和组织分化两个重要过程(Bar and Ori, 2015).叶片器官形态发生可分为3个阶段: 起始阶段、叶片极性的建立和细胞扩张(Tsukaya, 2005, 2006).这3个阶段受不同基因的调控(Byrne, 2005; Barkoulas et al., 2007).AN3/AtGIF1和At- GRF5的功能缺失突变体出现窄叶表型, 该表型由叶片细胞数目变少引起(Kim and Kende, 2004; Horigu- chi et al., 2005).与野生型相比, 过表达AN3和AtGRF5的植株叶片明显变大20%-30% (Horiguchi et al., 2005).在细胞扩张阶段, ROT3和ROT4基因参与调控叶片长度, rot3突变体表现出叶片发育畸形, ROT3过表达植株表现出叶片变长, 但宽度没有变化(Tsuge et al., 1996; Kim et al., 1998).ROT4基因编码膜结合的短肽, 其突变体的表型与rot3类似, 但叶片短小是由细胞增殖减少引起(Narita et al., 2004).LNG1和LNG2也参与调控叶片长度, 其突变体叶片短小(Lee et al., 2006).还有一些基因, 如AN蛋白基因参与叶片细胞的扩张(Folkers et al., 2002); SPIK- E1参与细胞骨架的构建, 最终决定细胞的形状和组织发育(Qiu et al., 2002).AN、SPIKE1、AN3、AtGIF1和AtHB13都参与调控叶片宽度, 且突变体叶片宽度都比较小(Hanson et al., 2001; Folkers et al., 2002; Kim et al., 2002; Qiu et al., 2002; Kim and Kende, 2004). ... ... 前人运用基因组和表达序列数据库等手段, 对拟南芥GRX基因家族进行了研究, 发现GRX家族的很多基因参与了植物的生长发育过程(Lemaire, 2004; Rouhier et al., 2004; Xing et al., 2006).本研究表明, GRXC9基因过表达株系叶片出现短小的表型, 且叶片短小的原因是由于叶片细胞体积变小.进一步检测叶片大小相关基因的表达情况, 发现过表达株系中AN、LNG1和LNG2的表达量明显下降.Tsuge等(1996)报道了AN基因通过调控特异的极性细胞伸长控制叶的形态.Kim等(2002)的研究发现, AN不仅控制叶细胞的极性伸长, 而且还可能通过调控细胞壁形成相关基因参与叶形态的调控.同时, AN是以一种非自主的模式控制细胞分裂(Bai et al., 2010).Lee等(2006)报道了LNG1和LNG2基因突变体叶片短小, 两基因共同参与叶片长度的调控.GRXC9基因过表达株系叶片短小可能是通过抑制参与叶形态调控基因的表达实现.之前有报道显示, GRXC9参与SA、JA和ET等多种激素调控途径, 在植物对外界环境的响应中发挥作用, 但具体机制尚不清楚(Ndamukong et al., 2007; Zander et al., 2012; Herrera-Vásquez et al., 2015).本研究表明, GRXC9可能通过影响AN、LNG1和LNG2等基因的表达参与叶片的发育, 但此过程是否与激素SA/JA和JA/ET的信号调控网络有关, 目前尚不明确. ... ... 植物器官的发育依赖于细胞的分裂、扩张及分化, 器官的最终大小由细胞数目和细胞大小决定(Mizu- kami, 2001; 李凌飞等, 2015).细胞分裂在植物器官发育过程中起重要作用(Vernoux et al., 2000).细胞周期调控因子CDKA和CycD3与一些基因(如拟南芥ANT基因)协同表达调控细胞分裂, 进而影响器官的发育(Mizukami and Fischer, 2000 ).细胞扩张与细胞分裂相比影响器官发育的方式更为复杂.植物细胞的最终大小与其倍性及核内复制相关, 但是具体分子机制尚不清楚(Kondorosi et al., 2000; Sugimoto- Shirasu and Roberts, 2003).植物一些器官(尤其是叶片)的大小和形状由极性细胞的扩张决定.例如, 拟南芥an突变体叶片较窄、厚, 原因是决定叶片宽度的细胞扩张受损(Kim et al., 2002); 拟南芥rot3突变体呈现出叶片较宽的表型, 是由于决定叶片长度的细胞扩张受损(Kim et al., 1998).此外, 细胞的伸长和扩张还受到赤霉素(GA)与油菜素内酯(BR)等多种激素的调控(Davies, 1995), 它们可能相互作用共同调控植物的器官发育和大小(Mizukami, 2001).综上, GRXC9基因可能通过抑制控制细胞扩张的基因AN以及调控叶形态的基因LNG1和LNG2的表达使叶片短小, 对拟南芥叶片的发育起负调控作用.然而, GRXC9是否也受到GA和BR等多种激素的调控, 以及是否有上游转录因子对其功能进行调节等均有待进一步研究. ...
2 1998
... 谷氧还蛋白家族基因在植物发育方面的作用已有报道, 但在植物叶片形态大小方面的研究尚少.叶片形成需要经历形态发生和组织分化两个重要过程(Bar and Ori, 2015).叶片器官形态发生可分为3个阶段: 起始阶段、叶片极性的建立和细胞扩张(Tsukaya, 2005, 2006).这3个阶段受不同基因的调控(Byrne, 2005; Barkoulas et al., 2007).AN3/AtGIF1和At- GRF5的功能缺失突变体出现窄叶表型, 该表型由叶片细胞数目变少引起(Kim and Kende, 2004; Horigu- chi et al., 2005).与野生型相比, 过表达AN3和AtGRF5的植株叶片明显变大20%-30% (Horiguchi et al., 2005).在细胞扩张阶段, ROT3和ROT4基因参与调控叶片长度, rot3突变体表现出叶片发育畸形, ROT3过表达植株表现出叶片变长, 但宽度没有变化(Tsuge et al., 1996; Kim et al., 1998).ROT4基因编码膜结合的短肽, 其突变体的表型与rot3类似, 但叶片短小是由细胞增殖减少引起(Narita et al., 2004).LNG1和LNG2也参与调控叶片长度, 其突变体叶片短小(Lee et al., 2006).还有一些基因, 如AN蛋白基因参与叶片细胞的扩张(Folkers et al., 2002); SPIK- E1参与细胞骨架的构建, 最终决定细胞的形状和组织发育(Qiu et al., 2002).AN、SPIKE1、AN3、AtGIF1和AtHB13都参与调控叶片宽度, 且突变体叶片宽度都比较小(Hanson et al., 2001; Folkers et al., 2002; Kim et al., 2002; Qiu et al., 2002; Kim and Kende, 2004). ... ... 植物器官的发育依赖于细胞的分裂、扩张及分化, 器官的最终大小由细胞数目和细胞大小决定(Mizu- kami, 2001; 李凌飞等, 2015).细胞分裂在植物器官发育过程中起重要作用(Vernoux et al., 2000).细胞周期调控因子CDKA和CycD3与一些基因(如拟南芥ANT基因)协同表达调控细胞分裂, 进而影响器官的发育(Mizukami and Fischer, 2000 ).细胞扩张与细胞分裂相比影响器官发育的方式更为复杂.植物细胞的最终大小与其倍性及核内复制相关, 但是具体分子机制尚不清楚(Kondorosi et al., 2000; Sugimoto- Shirasu and Roberts, 2003).植物一些器官(尤其是叶片)的大小和形状由极性细胞的扩张决定.例如, 拟南芥an突变体叶片较窄、厚, 原因是决定叶片宽度的细胞扩张受损(Kim et al., 2002); 拟南芥rot3突变体呈现出叶片较宽的表型, 是由于决定叶片长度的细胞扩张受损(Kim et al., 1998).此外, 细胞的伸长和扩张还受到赤霉素(GA)与油菜素内酯(BR)等多种激素的调控(Davies, 1995), 它们可能相互作用共同调控植物的器官发育和大小(Mizukami, 2001).综上, GRXC9基因可能通过抑制控制细胞扩张的基因AN以及调控叶形态的基因LNG1和LNG2的表达使叶片短小, 对拟南芥叶片的发育起负调控作用.然而, GRXC9是否也受到GA和BR等多种激素的调控, 以及是否有上游转录因子对其功能进行调节等均有待进一步研究. ...
2 2004
... 谷氧还蛋白家族基因在植物发育方面的作用已有报道, 但在植物叶片形态大小方面的研究尚少.叶片形成需要经历形态发生和组织分化两个重要过程(Bar and Ori, 2015).叶片器官形态发生可分为3个阶段: 起始阶段、叶片极性的建立和细胞扩张(Tsukaya, 2005, 2006).这3个阶段受不同基因的调控(Byrne, 2005; Barkoulas et al., 2007).AN3/AtGIF1和At- GRF5的功能缺失突变体出现窄叶表型, 该表型由叶片细胞数目变少引起(Kim and Kende, 2004; Horigu- chi et al., 2005).与野生型相比, 过表达AN3和AtGRF5的植株叶片明显变大20%-30% (Horiguchi et al., 2005).在细胞扩张阶段, ROT3和ROT4基因参与调控叶片长度, rot3突变体表现出叶片发育畸形, ROT3过表达植株表现出叶片变长, 但宽度没有变化(Tsuge et al., 1996; Kim et al., 1998).ROT4基因编码膜结合的短肽, 其突变体的表型与rot3类似, 但叶片短小是由细胞增殖减少引起(Narita et al., 2004).LNG1和LNG2也参与调控叶片长度, 其突变体叶片短小(Lee et al., 2006).还有一些基因, 如AN蛋白基因参与叶片细胞的扩张(Folkers et al., 2002); SPIK- E1参与细胞骨架的构建, 最终决定细胞的形状和组织发育(Qiu et al., 2002).AN、SPIKE1、AN3、AtGIF1和AtHB13都参与调控叶片宽度, 且突变体叶片宽度都比较小(Hanson et al., 2001; Folkers et al., 2002; Kim et al., 2002; Qiu et al., 2002; Kim and Kende, 2004). ... ... ; Kim and Kende, 2004). ...
1 2000
... 植物器官的发育依赖于细胞的分裂、扩张及分化, 器官的最终大小由细胞数目和细胞大小决定(Mizu- kami, 2001; 李凌飞等, 2015).细胞分裂在植物器官发育过程中起重要作用(Vernoux et al., 2000).细胞周期调控因子CDKA和CycD3与一些基因(如拟南芥ANT基因)协同表达调控细胞分裂, 进而影响器官的发育(Mizukami and Fischer, 2000 ).细胞扩张与细胞分裂相比影响器官发育的方式更为复杂.植物细胞的最终大小与其倍性及核内复制相关, 但是具体分子机制尚不清楚(Kondorosi et al., 2000; Sugimoto- Shirasu and Roberts, 2003).植物一些器官(尤其是叶片)的大小和形状由极性细胞的扩张决定.例如, 拟南芥an突变体叶片较窄、厚, 原因是决定叶片宽度的细胞扩张受损(Kim et al., 2002); 拟南芥rot3突变体呈现出叶片较宽的表型, 是由于决定叶片长度的细胞扩张受损(Kim et al., 1998).此外, 细胞的伸长和扩张还受到赤霉素(GA)与油菜素内酯(BR)等多种激素的调控(Davies, 1995), 它们可能相互作用共同调控植物的器官发育和大小(Mizukami, 2001).综上, GRXC9基因可能通过抑制控制细胞扩张的基因AN以及调控叶形态的基因LNG1和LNG2的表达使叶片短小, 对拟南芥叶片的发育起负调控作用.然而, GRXC9是否也受到GA和BR等多种激素的调控, 以及是否有上游转录因子对其功能进行调节等均有待进一步研究. ...
1 2012
... 拟南芥GRX家族的很多基因已被广泛研究, 它们在植物发育、激素信号途径以及抗逆反应等多个方面发挥重要作用.GRXC7/ROXY1基因参与花瓣的发育, 其突变体表现出花瓣和花原基异常的表型(Xing et al., 2005).GRXC8/ROXY2与GRXC7功能冗余, 且其可与TGA因子互作参与花药的发育(Murmu et al., 2010).表型观察发现, roxy1和roxy2单突变体花药发育正常, roxy1/roxy2双突变体花药发育存在缺陷.Ndamukong等(2007)的研究表明, GRXC9可与TGA转录因子互作, 受水杨酸(SA)诱导, 抑制茉莉酸(JA)途径的转录因子PDF1.2的转录, 推测GRXC9可能参与激素SA/JA的信号调控网络.之后, Zander等(2012)报道GRXC9与TGA因子互作能抑制JA/ET (乙烯)诱导的信号通路; 通过GRXC9的异位表达, 他们发现其负调控JA/ET信号途径的基因表达, 但具体机制还不十分清楚.GRXS13参与抗光氧化胁迫, 过表达GRXS13的植株在MV和高光处理后损伤程度比野生型有所减弱, 说明GRXS13的表达使ROS受到限制, 其可能参与拟南芥细胞内ROS的清除和抗光氧化的调控网络(Laporte et al., 2012).GRXS14通过与CAX1的N端互作, 参与调控Ca2+的转运(Cheng and Hirschi, 2003)和氧化胁迫(Cheng et al., 2006).Zhang等(2016)报道了棉铃虫中谷氧还蛋白HaGrx、 HaGrx3和HaGrx5受不同温度胁迫及过氧化氢(H2O2)的诱导; 进一步检测发现, 敲减植株中超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)、抗坏血酸和总谷胱甘肽(total GSH)等含量升高, 说明HaGrx、HaGrx3和HaGrx5具有抗氧化防御的功能.另外, Verma等(2016)发现水稻(Oryza sativa)中谷氧还蛋白OsGrx_C7以及OsGrx_C2.1对重金属砷敏感, 转化拟南芥的过表达植株表现出对砷的抗性增强, 说明在谷氧还蛋白家族中有些基因参与砷胁迫应答反应. ...
2 2006
... 谷氧还蛋白家族基因在植物发育方面的作用已有报道, 但在植物叶片形态大小方面的研究尚少.叶片形成需要经历形态发生和组织分化两个重要过程(Bar and Ori, 2015).叶片器官形态发生可分为3个阶段: 起始阶段、叶片极性的建立和细胞扩张(Tsukaya, 2005, 2006).这3个阶段受不同基因的调控(Byrne, 2005; Barkoulas et al., 2007).AN3/AtGIF1和At- GRF5的功能缺失突变体出现窄叶表型, 该表型由叶片细胞数目变少引起(Kim and Kende, 2004; Horigu- chi et al., 2005).与野生型相比, 过表达AN3和AtGRF5的植株叶片明显变大20%-30% (Horiguchi et al., 2005).在细胞扩张阶段, ROT3和ROT4基因参与调控叶片长度, rot3突变体表现出叶片发育畸形, ROT3过表达植株表现出叶片变长, 但宽度没有变化(Tsuge et al., 1996; Kim et al., 1998).ROT4基因编码膜结合的短肽, 其突变体的表型与rot3类似, 但叶片短小是由细胞增殖减少引起(Narita et al., 2004).LNG1和LNG2也参与调控叶片长度, 其突变体叶片短小(Lee et al., 2006).还有一些基因, 如AN蛋白基因参与叶片细胞的扩张(Folkers et al., 2002); SPIK- E1参与细胞骨架的构建, 最终决定细胞的形状和组织发育(Qiu et al., 2002).AN、SPIKE1、AN3、AtGIF1和AtHB13都参与调控叶片宽度, 且突变体叶片宽度都比较小(Hanson et al., 2001; Folkers et al., 2002; Kim et al., 2002; Qiu et al., 2002; Kim and Kende, 2004). ... ... 前人运用基因组和表达序列数据库等手段, 对拟南芥GRX基因家族进行了研究, 发现GRX家族的很多基因参与了植物的生长发育过程(Lemaire, 2004; Rouhier et al., 2004; Xing et al., 2006).本研究表明, GRXC9基因过表达株系叶片出现短小的表型, 且叶片短小的原因是由于叶片细胞体积变小.进一步检测叶片大小相关基因的表达情况, 发现过表达株系中AN、LNG1和LNG2的表达量明显下降.Tsuge等(1996)报道了AN基因通过调控特异的极性细胞伸长控制叶的形态.Kim等(2002)的研究发现, AN不仅控制叶细胞的极性伸长, 而且还可能通过调控细胞壁形成相关基因参与叶形态的调控.同时, AN是以一种非自主的模式控制细胞分裂(Bai et al., 2010).Lee等(2006)报道了LNG1和LNG2基因突变体叶片短小, 两基因共同参与叶片长度的调控.GRXC9基因过表达株系叶片短小可能是通过抑制参与叶形态调控基因的表达实现.之前有报道显示, GRXC9参与SA、JA和ET等多种激素调控途径, 在植物对外界环境的响应中发挥作用, 但具体机制尚不清楚(Ndamukong et al., 2007; Zander et al., 2012; Herrera-Vásquez et al., 2015).本研究表明, GRXC9可能通过影响AN、LNG1和LNG2等基因的表达参与叶片的发育, 但此过程是否与激素SA/JA和JA/ET的信号调控网络有关, 目前尚不明确. ...
1 2004
... 前人运用基因组和表达序列数据库等手段, 对拟南芥GRX基因家族进行了研究, 发现GRX家族的很多基因参与了植物的生长发育过程(Lemaire, 2004; Rouhier et al., 2004; Xing et al., 2006).本研究表明, GRXC9基因过表达株系叶片出现短小的表型, 且叶片短小的原因是由于叶片细胞体积变小.进一步检测叶片大小相关基因的表达情况, 发现过表达株系中AN、LNG1和LNG2的表达量明显下降.Tsuge等(1996)报道了AN基因通过调控特异的极性细胞伸长控制叶的形态.Kim等(2002)的研究发现, AN不仅控制叶细胞的极性伸长, 而且还可能通过调控细胞壁形成相关基因参与叶形态的调控.同时, AN是以一种非自主的模式控制细胞分裂(Bai et al., 2010).Lee等(2006)报道了LNG1和LNG2基因突变体叶片短小, 两基因共同参与叶片长度的调控.GRXC9基因过表达株系叶片短小可能是通过抑制参与叶形态调控基因的表达实现.之前有报道显示, GRXC9参与SA、JA和ET等多种激素调控途径, 在植物对外界环境的响应中发挥作用, 但具体机制尚不清楚(Ndamukong et al., 2007; Zander et al., 2012; Herrera-Vásquez et al., 2015).本研究表明, GRXC9可能通过影响AN、LNG1和LNG2等基因的表达参与叶片的发育, 但此过程是否与激素SA/JA和JA/ET的信号调控网络有关, 目前尚不明确. ...
1 2008
... 谷氧还蛋白(glutaredoxin, GRX)是一类谷胱甘肽(glutathione, GSH)依赖的氧化还原酶, 最早被发现可恢复大肠杆菌硫氧还蛋白(TRX)突变体的生长发育, 且在二硫键形成过程中能循环催化核苷酸还原酶(Holmgren, 1976, 1979a, 1979b).研究表明, GRX在人类健康、菌类及植物中均发挥重要作用(Holm- gren, 1976; Lillig et al., 2008).拟南芥(Arabidopsis thaliana)中GRX家族成员共有31个(Rouhier et al., 2006).根据它们的活性位点序列可分为3类, 即CP- YC型、CGFS型和CC型.其中CC型也被称作ROXY型, 主要存在于高等植物中.拟南芥中大多数GRX (21个)为CC类型(Rouhier et al., 2008), 此外, 6个为CPYC型, 4个为CGFS型(Meyer et al., 2009). ...
1 2009
... 谷氧还蛋白(glutaredoxin, GRX)是一类谷胱甘肽(glutathione, GSH)依赖的氧化还原酶, 最早被发现可恢复大肠杆菌硫氧还蛋白(TRX)突变体的生长发育, 且在二硫键形成过程中能循环催化核苷酸还原酶(Holmgren, 1976, 1979a, 1979b).研究表明, GRX在人类健康、菌类及植物中均发挥重要作用(Holm- gren, 1976; Lillig et al., 2008).拟南芥(Arabidopsis thaliana)中GRX家族成员共有31个(Rouhier et al., 2006).根据它们的活性位点序列可分为3类, 即CP- YC型、CGFS型和CC型.其中CC型也被称作ROXY型, 主要存在于高等植物中.拟南芥中大多数GRX (21个)为CC类型(Rouhier et al., 2008), 此外, 6个为CPYC型, 4个为CGFS型(Meyer et al., 2009). ...
2 2001
... 植物器官的发育依赖于细胞的分裂、扩张及分化, 器官的最终大小由细胞数目和细胞大小决定(Mizu- kami, 2001; 李凌飞等, 2015).细胞分裂在植物器官发育过程中起重要作用(Vernoux et al., 2000).细胞周期调控因子CDKA和CycD3与一些基因(如拟南芥ANT基因)协同表达调控细胞分裂, 进而影响器官的发育(Mizukami and Fischer, 2000 ).细胞扩张与细胞分裂相比影响器官发育的方式更为复杂.植物细胞的最终大小与其倍性及核内复制相关, 但是具体分子机制尚不清楚(Kondorosi et al., 2000; Sugimoto- Shirasu and Roberts, 2003).植物一些器官(尤其是叶片)的大小和形状由极性细胞的扩张决定.例如, 拟南芥an突变体叶片较窄、厚, 原因是决定叶片宽度的细胞扩张受损(Kim et al., 2002); 拟南芥rot3突变体呈现出叶片较宽的表型, 是由于决定叶片长度的细胞扩张受损(Kim et al., 1998).此外, 细胞的伸长和扩张还受到赤霉素(GA)与油菜素内酯(BR)等多种激素的调控(Davies, 1995), 它们可能相互作用共同调控植物的器官发育和大小(Mizukami, 2001).综上, GRXC9基因可能通过抑制控制细胞扩张的基因AN以及调控叶形态的基因LNG1和LNG2的表达使叶片短小, 对拟南芥叶片的发育起负调控作用.然而, GRXC9是否也受到GA和BR等多种激素的调控, 以及是否有上游转录因子对其功能进行调节等均有待进一步研究. ... ... ), 它们可能相互作用共同调控植物的器官发育和大小(Mizukami, 2001).综上, GRXC9基因可能通过抑制控制细胞扩张的基因AN以及调控叶形态的基因LNG1和LNG2的表达使叶片短小, 对拟南芥叶片的发育起负调控作用.然而, GRXC9是否也受到GA和BR等多种激素的调控, 以及是否有上游转录因子对其功能进行调节等均有待进一步研究. ...
2000
1 2010
... 拟南芥GRX家族的很多基因已被广泛研究, 它们在植物发育、激素信号途径以及抗逆反应等多个方面发挥重要作用.GRXC7/ROXY1基因参与花瓣的发育, 其突变体表现出花瓣和花原基异常的表型(Xing et al., 2005).GRXC8/ROXY2与GRXC7功能冗余, 且其可与TGA因子互作参与花药的发育(Murmu et al., 2010).表型观察发现, roxy1和roxy2单突变体花药发育正常, roxy1/roxy2双突变体花药发育存在缺陷.Ndamukong等(2007)的研究表明, GRXC9可与TGA转录因子互作, 受水杨酸(SA)诱导, 抑制茉莉酸(JA)途径的转录因子PDF1.2的转录, 推测GRXC9可能参与激素SA/JA的信号调控网络.之后, Zander等(2012)报道GRXC9与TGA因子互作能抑制JA/ET (乙烯)诱导的信号通路; 通过GRXC9的异位表达, 他们发现其负调控JA/ET信号途径的基因表达, 但具体机制还不十分清楚.GRXS13参与抗光氧化胁迫, 过表达GRXS13的植株在MV和高光处理后损伤程度比野生型有所减弱, 说明GRXS13的表达使ROS受到限制, 其可能参与拟南芥细胞内ROS的清除和抗光氧化的调控网络(Laporte et al., 2012).GRXS14通过与CAX1的N端互作, 参与调控Ca2+的转运(Cheng and Hirschi, 2003)和氧化胁迫(Cheng et al., 2006).Zhang等(2016)报道了棉铃虫中谷氧还蛋白HaGrx、 HaGrx3和HaGrx5受不同温度胁迫及过氧化氢(H2O2)的诱导; 进一步检测发现, 敲减植株中超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)、抗坏血酸和总谷胱甘肽(total GSH)等含量升高, 说明HaGrx、HaGrx3和HaGrx5具有抗氧化防御的功能.另外, Verma等(2016)发现水稻(Oryza sativa)中谷氧还蛋白OsGrx_C7以及OsGrx_C2.1对重金属砷敏感, 转化拟南芥的过表达植株表现出对砷的抗性增强, 说明在谷氧还蛋白家族中有些基因参与砷胁迫应答反应. ...
1 2004
... 谷氧还蛋白家族基因在植物发育方面的作用已有报道, 但在植物叶片形态大小方面的研究尚少.叶片形成需要经历形态发生和组织分化两个重要过程(Bar and Ori, 2015).叶片器官形态发生可分为3个阶段: 起始阶段、叶片极性的建立和细胞扩张(Tsukaya, 2005, 2006).这3个阶段受不同基因的调控(Byrne, 2005; Barkoulas et al., 2007).AN3/AtGIF1和At- GRF5的功能缺失突变体出现窄叶表型, 该表型由叶片细胞数目变少引起(Kim and Kende, 2004; Horigu- chi et al., 2005).与野生型相比, 过表达AN3和AtGRF5的植株叶片明显变大20%-30% (Horiguchi et al., 2005).在细胞扩张阶段, ROT3和ROT4基因参与调控叶片长度, rot3突变体表现出叶片发育畸形, ROT3过表达植株表现出叶片变长, 但宽度没有变化(Tsuge et al., 1996; Kim et al., 1998).ROT4基因编码膜结合的短肽, 其突变体的表型与rot3类似, 但叶片短小是由细胞增殖减少引起(Narita et al., 2004).LNG1和LNG2也参与调控叶片长度, 其突变体叶片短小(Lee et al., 2006).还有一些基因, 如AN蛋白基因参与叶片细胞的扩张(Folkers et al., 2002); SPIK- E1参与细胞骨架的构建, 最终决定细胞的形状和组织发育(Qiu et al., 2002).AN、SPIKE1、AN3、AtGIF1和AtHB13都参与调控叶片宽度, 且突变体叶片宽度都比较小(Hanson et al., 2001; Folkers et al., 2002; Kim et al., 2002; Qiu et al., 2002; Kim and Kende, 2004). ...
2 2007
... 前人的研究表明, 拟南芥谷氧还蛋白GRXC9基因参与激素SA/JA和JA/ET的信号调控网络(Ndamu- kong et al., 2007; Zander et al., 2012; Herrera- Vásquez et al., 2015).本研究通过探讨GRX的时空表达模式、亚细胞定位及对植物生长发育的调控, 发现其具有负调控叶片大小的功能.研究结果丰富了人们对GRXC9基因功能的认识, 拓宽了谷氧还蛋白家族在植物叶片发育方面的研究价值. ... ... 前人运用基因组和表达序列数据库等手段, 对拟南芥GRX基因家族进行了研究, 发现GRX家族的很多基因参与了植物的生长发育过程(Lemaire, 2004; Rouhier et al., 2004; Xing et al., 2006).本研究表明, GRXC9基因过表达株系叶片出现短小的表型, 且叶片短小的原因是由于叶片细胞体积变小.进一步检测叶片大小相关基因的表达情况, 发现过表达株系中AN、LNG1和LNG2的表达量明显下降.Tsuge等(1996)报道了AN基因通过调控特异的极性细胞伸长控制叶的形态.Kim等(2002)的研究发现, AN不仅控制叶细胞的极性伸长, 而且还可能通过调控细胞壁形成相关基因参与叶形态的调控.同时, AN是以一种非自主的模式控制细胞分裂(Bai et al., 2010).Lee等(2006)报道了LNG1和LNG2基因突变体叶片短小, 两基因共同参与叶片长度的调控.GRXC9基因过表达株系叶片短小可能是通过抑制参与叶形态调控基因的表达实现.之前有报道显示, GRXC9参与SA、JA和ET等多种激素调控途径, 在植物对外界环境的响应中发挥作用, 但具体机制尚不清楚(Ndamukong et al., 2007; Zander et al., 2012; Herrera-Vásquez et al., 2015).本研究表明, GRXC9可能通过影响AN、LNG1和LNG2等基因的表达参与叶片的发育, 但此过程是否与激素SA/JA和JA/ET的信号调控网络有关, 目前尚不明确. ...
2 2002
... 谷氧还蛋白家族基因在植物发育方面的作用已有报道, 但在植物叶片形态大小方面的研究尚少.叶片形成需要经历形态发生和组织分化两个重要过程(Bar and Ori, 2015).叶片器官形态发生可分为3个阶段: 起始阶段、叶片极性的建立和细胞扩张(Tsukaya, 2005, 2006).这3个阶段受不同基因的调控(Byrne, 2005; Barkoulas et al., 2007).AN3/AtGIF1和At- GRF5的功能缺失突变体出现窄叶表型, 该表型由叶片细胞数目变少引起(Kim and Kende, 2004; Horigu- chi et al., 2005).与野生型相比, 过表达AN3和AtGRF5的植株叶片明显变大20%-30% (Horiguchi et al., 2005).在细胞扩张阶段, ROT3和ROT4基因参与调控叶片长度, rot3突变体表现出叶片发育畸形, ROT3过表达植株表现出叶片变长, 但宽度没有变化(Tsuge et al., 1996; Kim et al., 1998).ROT4基因编码膜结合的短肽, 其突变体的表型与rot3类似, 但叶片短小是由细胞增殖减少引起(Narita et al., 2004).LNG1和LNG2也参与调控叶片长度, 其突变体叶片短小(Lee et al., 2006).还有一些基因, 如AN蛋白基因参与叶片细胞的扩张(Folkers et al., 2002); SPIK- E1参与细胞骨架的构建, 最终决定细胞的形状和组织发育(Qiu et al., 2002).AN、SPIKE1、AN3、AtGIF1和AtHB13都参与调控叶片宽度, 且突变体叶片宽度都比较小(Hanson et al., 2001; Folkers et al., 2002; Kim et al., 2002; Qiu et al., 2002; Kim and Kende, 2004). ... ... ; Qiu et al., 2002; Kim and Kende, 2004). ...
1 2006
... 谷氧还蛋白(glutaredoxin, GRX)是一类谷胱甘肽(glutathione, GSH)依赖的氧化还原酶, 最早被发现可恢复大肠杆菌硫氧还蛋白(TRX)突变体的生长发育, 且在二硫键形成过程中能循环催化核苷酸还原酶(Holmgren, 1976, 1979a, 1979b).研究表明, GRX在人类健康、菌类及植物中均发挥重要作用(Holm- gren, 1976; Lillig et al., 2008).拟南芥(Arabidopsis thaliana)中GRX家族成员共有31个(Rouhier et al., 2006).根据它们的活性位点序列可分为3类, 即CP- YC型、CGFS型和CC型.其中CC型也被称作ROXY型, 主要存在于高等植物中.拟南芥中大多数GRX (21个)为CC类型(Rouhier et al., 2008), 此外, 6个为CPYC型, 4个为CGFS型(Meyer et al., 2009). ...
1 2004
... 前人运用基因组和表达序列数据库等手段, 对拟南芥GRX基因家族进行了研究, 发现GRX家族的很多基因参与了植物的生长发育过程(Lemaire, 2004; Rouhier et al., 2004; Xing et al., 2006).本研究表明, GRXC9基因过表达株系叶片出现短小的表型, 且叶片短小的原因是由于叶片细胞体积变小.进一步检测叶片大小相关基因的表达情况, 发现过表达株系中AN、LNG1和LNG2的表达量明显下降.Tsuge等(1996)报道了AN基因通过调控特异的极性细胞伸长控制叶的形态.Kim等(2002)的研究发现, AN不仅控制叶细胞的极性伸长, 而且还可能通过调控细胞壁形成相关基因参与叶形态的调控.同时, AN是以一种非自主的模式控制细胞分裂(Bai et al., 2010).Lee等(2006)报道了LNG1和LNG2基因突变体叶片短小, 两基因共同参与叶片长度的调控.GRXC9基因过表达株系叶片短小可能是通过抑制参与叶形态调控基因的表达实现.之前有报道显示, GRXC9参与SA、JA和ET等多种激素调控途径, 在植物对外界环境的响应中发挥作用, 但具体机制尚不清楚(Ndamukong et al., 2007; Zander et al., 2012; Herrera-Vásquez et al., 2015).本研究表明, GRXC9可能通过影响AN、LNG1和LNG2等基因的表达参与叶片的发育, 但此过程是否与激素SA/JA和JA/ET的信号调控网络有关, 目前尚不明确. ...
1 2008
... 谷氧还蛋白(glutaredoxin, GRX)是一类谷胱甘肽(glutathione, GSH)依赖的氧化还原酶, 最早被发现可恢复大肠杆菌硫氧还蛋白(TRX)突变体的生长发育, 且在二硫键形成过程中能循环催化核苷酸还原酶(Holmgren, 1976, 1979a, 1979b).研究表明, GRX在人类健康、菌类及植物中均发挥重要作用(Holm- gren, 1976; Lillig et al., 2008).拟南芥(Arabidopsis thaliana)中GRX家族成员共有31个(Rouhier et al., 2006).根据它们的活性位点序列可分为3类, 即CP- YC型、CGFS型和CC型.其中CC型也被称作ROXY型, 主要存在于高等植物中.拟南芥中大多数GRX (21个)为CC类型(Rouhier et al., 2008), 此外, 6个为CPYC型, 4个为CGFS型(Meyer et al., 2009). ...
1 2003
... 植物器官的发育依赖于细胞的分裂、扩张及分化, 器官的最终大小由细胞数目和细胞大小决定(Mizu- kami, 2001; 李凌飞等, 2015).细胞分裂在植物器官发育过程中起重要作用(Vernoux et al., 2000).细胞周期调控因子CDKA和CycD3与一些基因(如拟南芥ANT基因)协同表达调控细胞分裂, 进而影响器官的发育(Mizukami and Fischer, 2000 ).细胞扩张与细胞分裂相比影响器官发育的方式更为复杂.植物细胞的最终大小与其倍性及核内复制相关, 但是具体分子机制尚不清楚(Kondorosi et al., 2000; Sugimoto- Shirasu and Roberts, 2003).植物一些器官(尤其是叶片)的大小和形状由极性细胞的扩张决定.例如, 拟南芥an突变体叶片较窄、厚, 原因是决定叶片宽度的细胞扩张受损(Kim et al., 2002); 拟南芥rot3突变体呈现出叶片较宽的表型, 是由于决定叶片长度的细胞扩张受损(Kim et al., 1998).此外, 细胞的伸长和扩张还受到赤霉素(GA)与油菜素内酯(BR)等多种激素的调控(Davies, 1995), 它们可能相互作用共同调控植物的器官发育和大小(Mizukami, 2001).综上, GRXC9基因可能通过抑制控制细胞扩张的基因AN以及调控叶形态的基因LNG1和LNG2的表达使叶片短小, 对拟南芥叶片的发育起负调控作用.然而, GRXC9是否也受到GA和BR等多种激素的调控, 以及是否有上游转录因子对其功能进行调节等均有待进一步研究. ...
2 1996
... 谷氧还蛋白家族基因在植物发育方面的作用已有报道, 但在植物叶片形态大小方面的研究尚少.叶片形成需要经历形态发生和组织分化两个重要过程(Bar and Ori, 2015).叶片器官形态发生可分为3个阶段: 起始阶段、叶片极性的建立和细胞扩张(Tsukaya, 2005, 2006).这3个阶段受不同基因的调控(Byrne, 2005; Barkoulas et al., 2007).AN3/AtGIF1和At- GRF5的功能缺失突变体出现窄叶表型, 该表型由叶片细胞数目变少引起(Kim and Kende, 2004; Horigu- chi et al., 2005).与野生型相比, 过表达AN3和AtGRF5的植株叶片明显变大20%-30% (Horiguchi et al., 2005).在细胞扩张阶段, ROT3和ROT4基因参与调控叶片长度, rot3突变体表现出叶片发育畸形, ROT3过表达植株表现出叶片变长, 但宽度没有变化(Tsuge et al., 1996; Kim et al., 1998).ROT4基因编码膜结合的短肽, 其突变体的表型与rot3类似, 但叶片短小是由细胞增殖减少引起(Narita et al., 2004).LNG1和LNG2也参与调控叶片长度, 其突变体叶片短小(Lee et al., 2006).还有一些基因, 如AN蛋白基因参与叶片细胞的扩张(Folkers et al., 2002); SPIK- E1参与细胞骨架的构建, 最终决定细胞的形状和组织发育(Qiu et al., 2002).AN、SPIKE1、AN3、AtGIF1和AtHB13都参与调控叶片宽度, 且突变体叶片宽度都比较小(Hanson et al., 2001; Folkers et al., 2002; Kim et al., 2002; Qiu et al., 2002; Kim and Kende, 2004). ... ... 前人运用基因组和表达序列数据库等手段, 对拟南芥GRX基因家族进行了研究, 发现GRX家族的很多基因参与了植物的生长发育过程(Lemaire, 2004; Rouhier et al., 2004; Xing et al., 2006).本研究表明, GRXC9基因过表达株系叶片出现短小的表型, 且叶片短小的原因是由于叶片细胞体积变小.进一步检测叶片大小相关基因的表达情况, 发现过表达株系中AN、LNG1和LNG2的表达量明显下降.Tsuge等(1996)报道了AN基因通过调控特异的极性细胞伸长控制叶的形态.Kim等(2002)的研究发现, AN不仅控制叶细胞的极性伸长, 而且还可能通过调控细胞壁形成相关基因参与叶形态的调控.同时, AN是以一种非自主的模式控制细胞分裂(Bai et al., 2010).Lee等(2006)报道了LNG1和LNG2基因突变体叶片短小, 两基因共同参与叶片长度的调控.GRXC9基因过表达株系叶片短小可能是通过抑制参与叶形态调控基因的表达实现.之前有报道显示, GRXC9参与SA、JA和ET等多种激素调控途径, 在植物对外界环境的响应中发挥作用, 但具体机制尚不清楚(Ndamukong et al., 2007; Zander et al., 2012; Herrera-Vásquez et al., 2015).本研究表明, GRXC9可能通过影响AN、LNG1和LNG2等基因的表达参与叶片的发育, 但此过程是否与激素SA/JA和JA/ET的信号调控网络有关, 目前尚不明确. ...
1 2005
... 谷氧还蛋白家族基因在植物发育方面的作用已有报道, 但在植物叶片形态大小方面的研究尚少.叶片形成需要经历形态发生和组织分化两个重要过程(Bar and Ori, 2015).叶片器官形态发生可分为3个阶段: 起始阶段、叶片极性的建立和细胞扩张(Tsukaya, 2005, 2006).这3个阶段受不同基因的调控(Byrne, 2005; Barkoulas et al., 2007).AN3/AtGIF1和At- GRF5的功能缺失突变体出现窄叶表型, 该表型由叶片细胞数目变少引起(Kim and Kende, 2004; Horigu- chi et al., 2005).与野生型相比, 过表达AN3和AtGRF5的植株叶片明显变大20%-30% (Horiguchi et al., 2005).在细胞扩张阶段, ROT3和ROT4基因参与调控叶片长度, rot3突变体表现出叶片发育畸形, ROT3过表达植株表现出叶片变长, 但宽度没有变化(Tsuge et al., 1996; Kim et al., 1998).ROT4基因编码膜结合的短肽, 其突变体的表型与rot3类似, 但叶片短小是由细胞增殖减少引起(Narita et al., 2004).LNG1和LNG2也参与调控叶片长度, 其突变体叶片短小(Lee et al., 2006).还有一些基因, 如AN蛋白基因参与叶片细胞的扩张(Folkers et al., 2002); SPIK- E1参与细胞骨架的构建, 最终决定细胞的形状和组织发育(Qiu et al., 2002).AN、SPIKE1、AN3、AtGIF1和AtHB13都参与调控叶片宽度, 且突变体叶片宽度都比较小(Hanson et al., 2001; Folkers et al., 2002; Kim et al., 2002; Qiu et al., 2002; Kim and Kende, 2004). ...
1 2006
... 谷氧还蛋白家族基因在植物发育方面的作用已有报道, 但在植物叶片形态大小方面的研究尚少.叶片形成需要经历形态发生和组织分化两个重要过程(Bar and Ori, 2015).叶片器官形态发生可分为3个阶段: 起始阶段、叶片极性的建立和细胞扩张(Tsukaya, 2005, 2006).这3个阶段受不同基因的调控(Byrne, 2005; Barkoulas et al., 2007).AN3/AtGIF1和At- GRF5的功能缺失突变体出现窄叶表型, 该表型由叶片细胞数目变少引起(Kim and Kende, 2004; Horigu- chi et al., 2005).与野生型相比, 过表达AN3和AtGRF5的植株叶片明显变大20%-30% (Horiguchi et al., 2005).在细胞扩张阶段, ROT3和ROT4基因参与调控叶片长度, rot3突变体表现出叶片发育畸形, ROT3过表达植株表现出叶片变长, 但宽度没有变化(Tsuge et al., 1996; Kim et al., 1998).ROT4基因编码膜结合的短肽, 其突变体的表型与rot3类似, 但叶片短小是由细胞增殖减少引起(Narita et al., 2004).LNG1和LNG2也参与调控叶片长度, 其突变体叶片短小(Lee et al., 2006).还有一些基因, 如AN蛋白基因参与叶片细胞的扩张(Folkers et al., 2002); SPIK- E1参与细胞骨架的构建, 最终决定细胞的形状和组织发育(Qiu et al., 2002).AN、SPIKE1、AN3、AtGIF1和AtHB13都参与调控叶片宽度, 且突变体叶片宽度都比较小(Hanson et al., 2001; Folkers et al., 2002; Kim et al., 2002; Qiu et al., 2002; Kim and Kende, 2004). ...
1 2016
... 拟南芥GRX家族的很多基因已被广泛研究, 它们在植物发育、激素信号途径以及抗逆反应等多个方面发挥重要作用.GRXC7/ROXY1基因参与花瓣的发育, 其突变体表现出花瓣和花原基异常的表型(Xing et al., 2005).GRXC8/ROXY2与GRXC7功能冗余, 且其可与TGA因子互作参与花药的发育(Murmu et al., 2010).表型观察发现, roxy1和roxy2单突变体花药发育正常, roxy1/roxy2双突变体花药发育存在缺陷.Ndamukong等(2007)的研究表明, GRXC9可与TGA转录因子互作, 受水杨酸(SA)诱导, 抑制茉莉酸(JA)途径的转录因子PDF1.2的转录, 推测GRXC9可能参与激素SA/JA的信号调控网络.之后, Zander等(2012)报道GRXC9与TGA因子互作能抑制JA/ET (乙烯)诱导的信号通路; 通过GRXC9的异位表达, 他们发现其负调控JA/ET信号途径的基因表达, 但具体机制还不十分清楚.GRXS13参与抗光氧化胁迫, 过表达GRXS13的植株在MV和高光处理后损伤程度比野生型有所减弱, 说明GRXS13的表达使ROS受到限制, 其可能参与拟南芥细胞内ROS的清除和抗光氧化的调控网络(Laporte et al., 2012).GRXS14通过与CAX1的N端互作, 参与调控Ca2+的转运(Cheng and Hirschi, 2003)和氧化胁迫(Cheng et al., 2006).Zhang等(2016)报道了棉铃虫中谷氧还蛋白HaGrx、 HaGrx3和HaGrx5受不同温度胁迫及过氧化氢(H2O2)的诱导; 进一步检测发现, 敲减植株中超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)、抗坏血酸和总谷胱甘肽(total GSH)等含量升高, 说明HaGrx、HaGrx3和HaGrx5具有抗氧化防御的功能.另外, Verma等(2016)发现水稻(Oryza sativa)中谷氧还蛋白OsGrx_C7以及OsGrx_C2.1对重金属砷敏感, 转化拟南芥的过表达植株表现出对砷的抗性增强, 说明在谷氧还蛋白家族中有些基因参与砷胁迫应答反应. ...
1 2000
... 植物器官的发育依赖于细胞的分裂、扩张及分化, 器官的最终大小由细胞数目和细胞大小决定(Mizu- kami, 2001; 李凌飞等, 2015).细胞分裂在植物器官发育过程中起重要作用(Vernoux et al., 2000).细胞周期调控因子CDKA和CycD3与一些基因(如拟南芥ANT基因)协同表达调控细胞分裂, 进而影响器官的发育(Mizukami and Fischer, 2000 ).细胞扩张与细胞分裂相比影响器官发育的方式更为复杂.植物细胞的最终大小与其倍性及核内复制相关, 但是具体分子机制尚不清楚(Kondorosi et al., 2000; Sugimoto- Shirasu and Roberts, 2003).植物一些器官(尤其是叶片)的大小和形状由极性细胞的扩张决定.例如, 拟南芥an突变体叶片较窄、厚, 原因是决定叶片宽度的细胞扩张受损(Kim et al., 2002); 拟南芥rot3突变体呈现出叶片较宽的表型, 是由于决定叶片长度的细胞扩张受损(Kim et al., 1998).此外, 细胞的伸长和扩张还受到赤霉素(GA)与油菜素内酯(BR)等多种激素的调控(Davies, 1995), 它们可能相互作用共同调控植物的器官发育和大小(Mizukami, 2001).综上, GRXC9基因可能通过抑制控制细胞扩张的基因AN以及调控叶形态的基因LNG1和LNG2的表达使叶片短小, 对拟南芥叶片的发育起负调控作用.然而, GRXC9是否也受到GA和BR等多种激素的调控, 以及是否有上游转录因子对其功能进行调节等均有待进一步研究. ...
1 2006
... 前人运用基因组和表达序列数据库等手段, 对拟南芥GRX基因家族进行了研究, 发现GRX家族的很多基因参与了植物的生长发育过程(Lemaire, 2004; Rouhier et al., 2004; Xing et al., 2006).本研究表明, GRXC9基因过表达株系叶片出现短小的表型, 且叶片短小的原因是由于叶片细胞体积变小.进一步检测叶片大小相关基因的表达情况, 发现过表达株系中AN、LNG1和LNG2的表达量明显下降.Tsuge等(1996)报道了AN基因通过调控特异的极性细胞伸长控制叶的形态.Kim等(2002)的研究发现, AN不仅控制叶细胞的极性伸长, 而且还可能通过调控细胞壁形成相关基因参与叶形态的调控.同时, AN是以一种非自主的模式控制细胞分裂(Bai et al., 2010).Lee等(2006)报道了LNG1和LNG2基因突变体叶片短小, 两基因共同参与叶片长度的调控.GRXC9基因过表达株系叶片短小可能是通过抑制参与叶形态调控基因的表达实现.之前有报道显示, GRXC9参与SA、JA和ET等多种激素调控途径, 在植物对外界环境的响应中发挥作用, 但具体机制尚不清楚(Ndamukong et al., 2007; Zander et al., 2012; Herrera-Vásquez et al., 2015).本研究表明, GRXC9可能通过影响AN、LNG1和LNG2等基因的表达参与叶片的发育, 但此过程是否与激素SA/JA和JA/ET的信号调控网络有关, 目前尚不明确. ...
2 2005
... 拟南芥GRX家族的很多基因已被广泛研究, 它们在植物发育、激素信号途径以及抗逆反应等多个方面发挥重要作用.GRXC7/ROXY1基因参与花瓣的发育, 其突变体表现出花瓣和花原基异常的表型(Xing et al., 2005).GRXC8/ROXY2与GRXC7功能冗余, 且其可与TGA因子互作参与花药的发育(Murmu et al., 2010).表型观察发现, roxy1和roxy2单突变体花药发育正常, roxy1/roxy2双突变体花药发育存在缺陷.Ndamukong等(2007)的研究表明, GRXC9可与TGA转录因子互作, 受水杨酸(SA)诱导, 抑制茉莉酸(JA)途径的转录因子PDF1.2的转录, 推测GRXC9可能参与激素SA/JA的信号调控网络.之后, Zander等(2012)报道GRXC9与TGA因子互作能抑制JA/ET (乙烯)诱导的信号通路; 通过GRXC9的异位表达, 他们发现其负调控JA/ET信号途径的基因表达, 但具体机制还不十分清楚.GRXS13参与抗光氧化胁迫, 过表达GRXS13的植株在MV和高光处理后损伤程度比野生型有所减弱, 说明GRXS13的表达使ROS受到限制, 其可能参与拟南芥细胞内ROS的清除和抗光氧化的调控网络(Laporte et al., 2012).GRXS14通过与CAX1的N端互作, 参与调控Ca2+的转运(Cheng and Hirschi, 2003)和氧化胁迫(Cheng et al., 2006).Zhang等(2016)报道了棉铃虫中谷氧还蛋白HaGrx、 HaGrx3和HaGrx5受不同温度胁迫及过氧化氢(H2O2)的诱导; 进一步检测发现, 敲减植株中超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)、抗坏血酸和总谷胱甘肽(total GSH)等含量升高, 说明HaGrx、HaGrx3和HaGrx5具有抗氧化防御的功能.另外, Verma等(2016)发现水稻(Oryza sativa)中谷氧还蛋白OsGrx_C7以及OsGrx_C2.1对重金属砷敏感, 转化拟南芥的过表达植株表现出对砷的抗性增强, 说明在谷氧还蛋白家族中有些基因参与砷胁迫应答反应. ... ... 基因精确的时空表达是其正常行使功能的基础与前提, 基因在不同时间空间的表达差异暗示了其功能的不同.因此, 研究GRXC9基因的时空表达, 将为探明该基因的功能提供重要线索.目前, 对GRX基因家族表达部位的研究较少.已有的研究表明, CC型的RO- XY1 (GRXC7)基因在花序、根以及果荚中强烈表达, 而在茎叶中不表达(Xing et al., 2005). ROXY2 (GR- XC8)基因在花原基和分生组织中表达较少, 而在花药发育时期表达量最高, 这与其参与雄蕊发育的功能相对应(Xing and Zachgo, 2008).GRXS14是组成型表达基因, 其在叶片、维管束、根、茎和花中均有表达, 但在叶片中的表达量最高(Cheng et al., 2006).通过RT-PCR分析及构建GUS启动子融合表达载体两种手段, 我们研究了GRXC9的时空表达模式, 结果发现GRXC9基因的表达不具有组织特异性, 其在植物根、茎、叶、花和角果中均有表达, 且在花中的表达主要是花丝和花药部位, 暗示GRXC9基因可能参与雄蕊的生长发育. ...


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}