 , ,
, , 引用本文
贡献者
基金资助
接受日期:2016-03-14接受日期:2016-05-28网络出版日期:2017-07-1
-->Copyright
20172010 《植物学报》编辑部
Contributors
History
Received:Accepted:Online:
摘要:
Abstract:
Key words:
盐渍土是指受土壤中盐碱成分作用的, 包括盐土、碱土以及其它不同程度盐化和碱化的各种类型土壤的统称(董合忠, 2010)。土壤的盐渍化是农业可持续发展面临的主要环境问题。据统计, 我国盐渍化土壤面积约为3.69×107 hm2, 占现有耕地面积的25% (贾亚雄和李向林, 2008)。新疆是我国重要的棉花(Gossypium hirsutum)生产基地, 又是我国唯一的海岛棉(G. barbadense)产区, 该区域现有耕地中已有31.1%的面积受到盐渍危害(杨淑萍和危常州, 2010)。
土壤中盐(NaCl)的过量积累可导致各种农作物减产(Huseyin, 2010)。土壤中的Na盐和Ca盐对植物危害最明显。其中, 尤以Na盐的危害最为普遍。在盐胁迫下, 棉花体内Na+是主要的毒害因子(叶武威等, 2006)。
研究表明, 光合作用对盐胁迫十分敏感, 即光合速率随着盐浓度的增加和胁迫时间的延长而明显下降(孙仁国等, 2010; 刘志萍等, 2014)。盐胁迫影响CO2的扩散, 改变与光反应有关的细胞器结构与功能, 还可作用于暗反应过程, 减少同化物的转移, 最终导致光合速率降低(王智明等, 2014)。
陆地棉(G. hirsutum)和海岛棉是棉花的2个重要栽培种。陆地棉丰产性好, 而海岛棉不仅纤维品质优良, 而且是盐碱地的主要栽培作物。前人的研究表明, 棉花虽然是盐碱地的先锋作物, 但其耐盐能力十分有限, 且各生育阶段的耐盐能力不同(王俊娟等, 2011)。棉花种子萌发、出苗和幼苗阶段是对盐胁迫比较敏感的时期, 也是对棉花品种进行耐盐性筛选的关键时期。当NaCl浓度≤25 mol·L-1时, 盐胁迫对棉花种子发芽影响不显著; 而NaCl浓度≥125 mol·L-1时, 则显著抑制种子发芽; 但无论盐浓度的高低都会抑制幼苗的生长, 即幼苗根、叶、茎、根冠比和总干重均下降(文卿琳和阿曼古丽·肉孜, 2008)。研究盐胁迫下海岛棉和陆地棉幼苗生长与生理特性, 探明海岛棉和陆地棉的耐盐性差异, 对新疆棉花耐盐品种选育与高产高效栽培实践具有重要意义。
1 材料与方法1.1 供试材料实验于2015年在新疆农业大学教育部棉花工程研究中心棉花生理实验室内进行。以海岛棉(Gossypium barbadense L.)品种(新海21号和新海34号)和陆地棉 (G. hirsutum L.)品种(新陆早50号和新陆早57号)为材料(表1), 从播种期开始进行盐胁迫。供试用盐为NaCl (分析纯)。
表1
Table 1
表1
| No. | Variety name | Approval of the year | Variety type | Growing period (d) | Suitable plant region |
|---|---|---|---|---|---|
| 1 | Xinhai 21 | 2003 | Middle maturity | 141 | No wilt disease and mild disease cotton region of southern Xinjiang |
| 2 | Xinhai 34 | 2010 | Early maturity | 142 | No wilt disease or mild disease cotton region of southern Xinjiang |
| 3 | Xinluzao 57 | 2013 | Early maturity | 122 | Early mature cotton region of northern Xinjiang |
| 4 | Xinluzao 50 | 2011 | Early maturity | 126 | Early mature cotton region of northern and southern Xinjiang |
表1
供试棉花品种
Table 1
The cotton varieties tested
1.2 实验设计与幼苗培养采用盆栽法培育海岛棉和陆地棉幼苗。种子经0.1% HgCl2溶液消毒15分钟后, 置于25°C智能光照培养箱(GTOP-380B2, 浙江托普)中催芽至露白。将草炭、蛭石按1:1 (v/v)配比的复合基质于120°C高温灭菌24小时。
土壤NaCl含量(%)设置4个水平: 0 (CK)、0.6%、0.8%和1.0%。将配制好的NaCl溶液均匀灌入草炭、蛭石复合基质内, 选择发芽整齐一致的种子播入塑料营养钵(12 cm×11 cm)中, 每盆装土310 g, 每盆播种6粒种子, 每品种每处理种18盆, 每粒种子的播种深度为2.3 cm。培养条件为30°C/22°C (昼/夜), 相对湿度(RH) 50%, 光强400-600 µmol·m-2·s-1, 光周期12小时光照/12小时黑暗。棉花出苗后24天开始取样并进行测定。
1.3 测定方法1.3.1 农艺性状
幼苗长至25天, 用直尺测定每株棉花幼苗子叶节到顶端的垂直高度、叶长和叶宽(单位均为cm), 按如下公式计算叶面积: 叶面积=叶长×叶宽×0.84。取5株幼苗进行生物量测定, 先测定鲜重, 之后将鲜样在105°C的烘箱中杀青30分钟, 于80°C下烘干至恒重并称其干重(g)。
1.3.2 生理特征
各项生理指标测定参照刘萍和李明军(2007)的方法并略有改进。幼苗长至25天, 选取长势整齐一致植株的第1片真叶, 去掉叶脉后剪碎混匀, 称取0.1 g, 用10 mL乙醇:丙酮为1:1 (v:v)的混合液暗室浸泡至组织变白(48小时), 橡皮塞封口防止提取液挥发, 用UV-1800紫外/可见分光光度计测定663 nm及645 nm处的吸光值。实验设3次重复。
幼苗生长24天后, 使用CARIS-2型便携式光合仪(PP Systems, USA)测定植株第1片真叶净光合速率(Pn)和蒸腾速率(Tr)等指标, 测定过程中光强为400 µmol·m-2·s-1, 环境温度(25±1)°C, 大气CO2浓度(Cr)变化范围为(410±10) µmol·L-1 (使用开放式气路, CO2浓度约为385 µmol·mol-1)。选取长势均匀的植株, 设5次重复。采用愈创木酚法测定过氧化物酶(POD)活性(史树德等, 2011)。采用NBT光化还原法测定超氧化物歧化酶(SOD)活性(张志安和陈展宇, 2008)。采用3%磺基水杨酸提取, 酸性茚三酮显色法测定脯氨酸(Pro)含量(蔡庆生, 2013)。采用蒽酮乙酸乙酯法测定可溶性糖(SS)含量(邹琦, 2007)。采用硫代巴比妥酸比色法测定丙二醛(MDA)含量(邹琦, 2007)。实验重复3次。
1.4 数据统计利用Excel 2010软件进行数据处理及作图。采用SPSS17.0统计学软件进行方差分析(ANOVA)和多重比较(采用LSD比较法, P<0.05)。
2 结果与讨论2.1 盐胁迫对海岛棉和陆地棉幼苗生长的影响由表2可知, 不同浓度(0.6%、0.8%和1.0%)的NaCl胁迫对海岛棉和陆地棉幼苗生长产生不同程度的抑制作用, 导致植株生长缓慢和干物质积累减少。实验结果表明, NaCl胁迫对海岛棉和陆地棉生长有比较明显的抑制作用, 且抑制效果随着盐浓度的增加而更加明显。在0.8%和1.0% NaCl胁迫下, 新海21号、新海34号和新陆早50号、新陆早57号植株的株高均下降, 与对照相比差异显著(P<0.05)。
表2
Table 2
表2
| Variety | NaCl treatment (%) | Plant height (cm) | Fresh weight (g·plant-1) | Dry weight (g·plant-1) |
|---|---|---|---|---|
| Xinhai 21 | 0 (CK) 0.6 0.8 1.0 | 16.54±0.50 a | 2.26±0.49 a | 0.40±0.05 a |
| 15.12±0.52 b | 2.07±0.12 a | 0.37±0.08 a | ||
| 12.68±1.02 c | 1.61±0.29 b | 0.27±0.04 b | ||
| 11.34±1.10 d | 1.28±0.10 b | 0.23±0.05 b | ||
| Xinhai 34 | 0 (CK) 0.6 0.8 1.0 | 14.92±0.56 a | 2.14±0.28 a | 0.32±0.05 a |
| 12.42±1.03 b | 1.61±0.06 b | 0.25±0.04 b | ||
| 9. 80±1.08 c | 1.12±0.05 c | 0.18±0.04 c | ||
| 8.48±0.73 d | 0.61±0.21 d | 0.10±0.02 d | ||
| Xinluzao 50 | 0 (CK) 0.6 0.8 1.0 | 18.78±0.73 a | 2.22±0.11 a | 0.35±0.02 a |
| 14.90±0.29 b | 1.56±0.33 b | 0.24±0.06 b | ||
| 11.22±1.51 c | 0.99±0.24 c | 0.17±0.05 c | ||
| 9.62±0.98 d | 0.29±0.09 d | 0.09±0.07 d | ||
| Xinluzao 57 | 0 (CK) 0.6 0.8 1.0 | 16.86±1.00 a | 2.24±0.11 a | 0.37±0.06 a |
| 13.52±0.58 b | 1.68±0.10 b | 0.26±0.05 b | ||
| 10.62±0.95 c | 1.17±0.15 c | 0.19±0.03 c | ||
| 9.26±0.58 d | 0.52±0.38 d | 0.12±0.05 c |
表2
盐胁迫对海岛棉和陆地棉幼苗生长的影响
Table 2
Effects of salt stress on plant growth of Sea Island cotton and Upland cotton seedlings
从单株鲜重和干重的变化来看, 不同浓度的盐胁迫下, 海岛棉和陆地棉鲜重和干重均受到影响。在0.6% NaCl胁迫下, 新海21号的鲜重和干重受盐胁迫的影响较轻, 与对照(CK)相比差异不显著, 说明此浓度的盐胁迫对新海21号幼苗干物质的积累影响不大; 但随着盐浓度的增加, 干物质积累受到明显影响。而在0.6% NaCl胁迫下, 其余3个供试品种的鲜重和干重均降低, 且与对照相比差异显著。在高盐胁迫(1.0%NaCl)下, 新海21号、新海34号、新陆早50号与新陆早57号的鲜重分别比对照下降43.36%、71.50%、86.94%和76.79%。
2.2 盐胁迫对海岛棉和陆地棉幼苗叶面积和叶绿素含量的影响叶片是植物进行光合作用的重要器官。从表3可以看出, 盐胁迫使海岛棉和陆地棉叶面积显著减少, 且随盐浓度的增加抑制作用增强。0.6% NaCl胁迫下, 4个供试品种的叶面积受胁迫的影响相比其它浓度盐处理较轻, 但下降幅度与对照(CK)相比差异显著(P< 0.05)。高浓度盐胁迫下, 2个棉花栽培种的叶面积受到明显抑制, 出现叶片小、萎蔫、发黄和增厚等现象。新陆早50号和新陆早57号对高浓度盐胁迫较敏感, 其第1片真叶展开受到严重抑制, 生长发育迟缓, 与对照相比长时间处在子叶期。以上结果表明NaCl胁迫对陆地棉叶面积的抑制作用强于海岛棉。
表3
Table 3
表3
| Variety | NaCl treatment (%) | Leaf area per plant (cm2·plant-1) | Chlorophyll-a content (mg·g-1 FW) | Chlorophyll-b content (mg·g-1 FW) | Total chlorophyll content (mg·g-1 FW) | |
|---|---|---|---|---|---|---|
| Xinhai 21 | 0 (CK) 0.6 0.8 1.0 | 25.15±3.73 a | 1.57±0.11 a | 0.43±0.06 a | 2.00±0.17 a | |
| 19.54±3.38 b | 1.27±0.24 ab | 0.35±0.07 a | 1.62±0.31 ab | |||
| 17.17±3.20 bd | 0.81±0.60 ab | 0.29±0.19 a | 1.10±0.45 bc | |||
| 13.83±1.31 d | 0.59±0.68 b | 0.18±0.08 ab | 0.77±0.69 c | |||
| Xinhai 34 | 0 (CK) 0.6 0.8 1.0 | 24.98±2.03 a | 2.49±0.52 a | 0.82±0.24 a | 3.31±0.75 a | |
| 18.39±4.024 b | 1.82±0.56 ab | 0.57±0.08 b | 2.39±0.57 ab | |||
| 12.14±1.67 c | 1.63±0.80 bc | 0.44±0.08 bc | 2.07±0.88 b | |||
| 8.00±0.94 d | 0.70±0.04 c | 0.27±0.06 c | 0.97±0.06 c | |||
| Xinluzao 50 | 0 (CK) 0.6 0.8 1.0 | 14.38±1.68 a | 2.06±0.01 a | 0.78±0.03 a | 2.84±0.03 a | |
| 11.30±1.90 b | 1.48±0.08 b | 0.53±0.14 b | 1.98±0.21 b | |||
| 4.25±1.36 c | 0.89±0.56 c | 0.32±0.15 c | 1.21±0.50 c | |||
| 2.34±1.29 c | / | / | / | |||
| Xinluzao 57 | 0 (CK) 0.6 0.8 1.0 | 16.42±2.81 a | 1.98±0.06 a | 0.84±0.08 a | 2.82±0.02 a | |
| 11.77±1.91 b | 1.44±0.06 b | 0.58±0.09 b | 2.02±0.06 b | |||
| 6.55±0.79 c | 0.91±0.40 c | 0.35±0.05 c | 1.26±0.37 c | |||
| 2.98±1.07 d | / | / | / |
表3
盐胁迫对海岛棉和陆地棉单株叶面积和叶绿素含量的影响
Table 3
Effects of salt stress on leaf area per plant and chlorophyll content of Sea Island cotton and Upland cotton seedlings
叶绿素是与光合作用有关的重要色素, 在逆境条件下, 光合功能的降低常与叶绿素含量的变化有关。本研究表明, 盐胁迫下, 海岛棉和陆地棉叶片叶绿素含量与各自对照相比发生了明显变化, 且随着土壤盐浓度的增加, 4个供试品种叶绿素含量呈不同程度的下降趋势(表3)。由表3可知, 0.6% NaCl胁迫对新海21号、新海34号和新陆早50号、新陆早57号叶片叶绿素含量的影响相对较轻, 而在0.8%和1.0%盐胁迫下, 4个品种的叶绿素含量则显著下降。新陆早50号和新陆早57号在高盐(1.0% NaCl)胁迫后, 植株死亡(叶绿素含量、光合气体交换参数及抗氧化酶活性等数据未能采集)。
2.3 盐胁迫对海岛棉和陆地棉幼苗叶片净光合速率和蒸腾速率的影响从表4可以看出, 新陆早50号对照(CK)叶片净光合速率略高于海岛棉品种新海34号和新海21号。经NaCl胁迫之后, 4个供试品种的净光合速率(Pn)显著下降, 新陆早57号和新陆早50号的降幅大于新海21号和新海34号。在0.8% NaCl胁迫下, 新海21号、新海34号、新陆早57号和新陆早50号与对照相比分别下降47.35%、50.56%、67.09%和73.94%。在1.0% NaCl胁迫下, 新陆早50号和新陆早57号幼苗叶片受盐胁迫影响较严重, 叶片较小, 无法测定Pn; 而新海21号和新海34号在此盐胁迫条件下, Pn与CK相比仅分别下降61.15%和75.62%。由此可见, 与新陆早57号和新陆早50号相比, 新海21号和新海34号幼苗叶片在1.0% NaCl胁迫下仍能维持较高的净光合速率, 说明盐胁迫对2种海岛棉幼苗碳同化能力的影响较小。
表4
Table 4
表4
| Variety | NaCl treatment (%) | Net photosynthe- tic rate (μmol CO2∙m-2∙s-1) | Transpiration rate (mmol H2O∙m-2∙s-1) |
|---|---|---|---|
| Xinhai 21 | 0 (CK) 0.6 0.8 1.0 | 9.52±1.32 a 7.80±1.13 b 5.24±1.08 c 3.66±1.50 c | 2.94±0.54 a 2.81±1.33 a 2.00±1.02 ab 1.14±0.70 b |
| Xinhai 34 | 0 (CK) 0.6 0.8 1.0 | 8.86±1.59 a 6.94±1.37 b 3.98±0.45 c 2.16±0.98 d | 3.25±0.23 a 2.96±0.41 a 2.01±0.57 b 0.84±0.38 c |
| Xinluzao 50 | 0 (CK) 0.6 0.8 1.0 | 10.36±1.90 a 6.88±1.65 b 3.54±0.51 c / | 4.09±0.57 a 3.18±0.50 a 1.30±0.81 a / |
| Xinluzao 57 | 0 (CK) 0.6 0.8 1.0 | 9.48±2.21 a 6.64±1.93 b 4.02±1.79 c / | 3.30±0.81 a 2.56±1.22 a 1.16±0.50 b / |
表4
盐胁迫对海岛棉和陆地棉叶片净光合速率(Pn)和蒸腾速率(Tr)的影响
Table 4
Effects of salt stress on net photosynthetic rate (Pn) and transpiration rate (Tr) of Sea Island cotton and Upland cotton seedlings
蒸腾作用是植物对水分和矿物质吸收和运输的主要动力。在盐胁迫下, 4个供试品种幼苗叶片蒸腾速率(Tr)均下降。Tr大幅度降低表明盐胁迫抑制了棉花幼苗叶片对水分及矿物质的吸收和运输, 从而影响正常的光合作用。在盐胁迫下, 新海21号、新海34号和新陆早50号、新陆早57号的Tr均呈不同程度的下降, 但新海21号和新海34号在所有浓度的盐胁迫下, 均保持比新陆早57号和新陆早50号更高的Tr。
2.4 盐胁迫对海岛棉和陆地棉幼苗叶片过氧化物酶和超氧化物歧化酶活性的影响由图1A可以看出, 新陆早57号和新陆早50号对照 (CK)叶片POD活性高于新海21号和新海34号对照。在盐胁迫下, 4个供试品种叶片POD活性随着土壤盐浓度的增加呈先升高后降低的趋势, 上升和降低与对照相比均达到显著水平(P<0.05)。新海34号、新陆早57号和新陆早50号在0.6% NaCl胁迫下叶片POD活性达到最大值, 与对照相比分别提高了47.50%、29.27%和23.44%。当NaCl浓度为1.0%时, 新海34号、新陆早50号和新陆早57号由于受胁迫较严重, 叶片生长不良(没有正常的叶片), 因此未能采集数据。而新海21号随着盐浓度的增加其叶片POD活性一直呈上升趋势, 即在0.8% NaCl胁迫下叶片POD活性达到最大值, 比对照增高了70.54%。但盐浓度达到最高时其POD活性开始下降, 但仍高于对照。
图1
Figure 1

下载原图ZIP
生成PPT
图1
盐胁迫对海岛棉和陆地棉幼苗叶片过氧化物酶(A)和超氧化物歧化酶(B)活性的影响
Figure 1
Effects of salt stress on peroxidase (A) and super oxide dismutase (B) activity of Sea Island cotton and Upland cotton seedlings
从图1B可以看出, 经盐胁迫后, 随土壤盐浓度的增加, 4个供试品种SOD活性均呈先升高后降低的趋势, 与POD活性变化规律一致。0.6% NaCl胁迫下, 4个供试品种叶片SOD活性增加, 均高于对照(CK), 且差异达显著水平(P<0.05)。而在0.8% NaCl胁迫下, 新海34号、新陆早57号和新陆早50号的幼苗叶片SOD活性开始下降, 与对照相比分别下降24.63%、44.21%和46.38%, 新海21号SOD活性则与CK相当; 高盐(1.0% NaCl)胁迫时, 新海21号叶片SOD活性迅速下降。
总体上看, 在0.6%-0.8%盐胁迫环境中, 新海21号和新海34号叶片SOD活性上升幅度要大于新陆早57号和新陆早50号; 且在0.8%-1.0% NaCl胁迫下, 海岛棉品种SOD活性的下降幅度低于新陆早57号和新陆早50号, 表明新海21号和新海34号具有更加敏感的SOD活性调节能力, 能快速提高SOD活性, 消除超氧阴离子(O2∙-), 降低活性氧对植物的伤害。
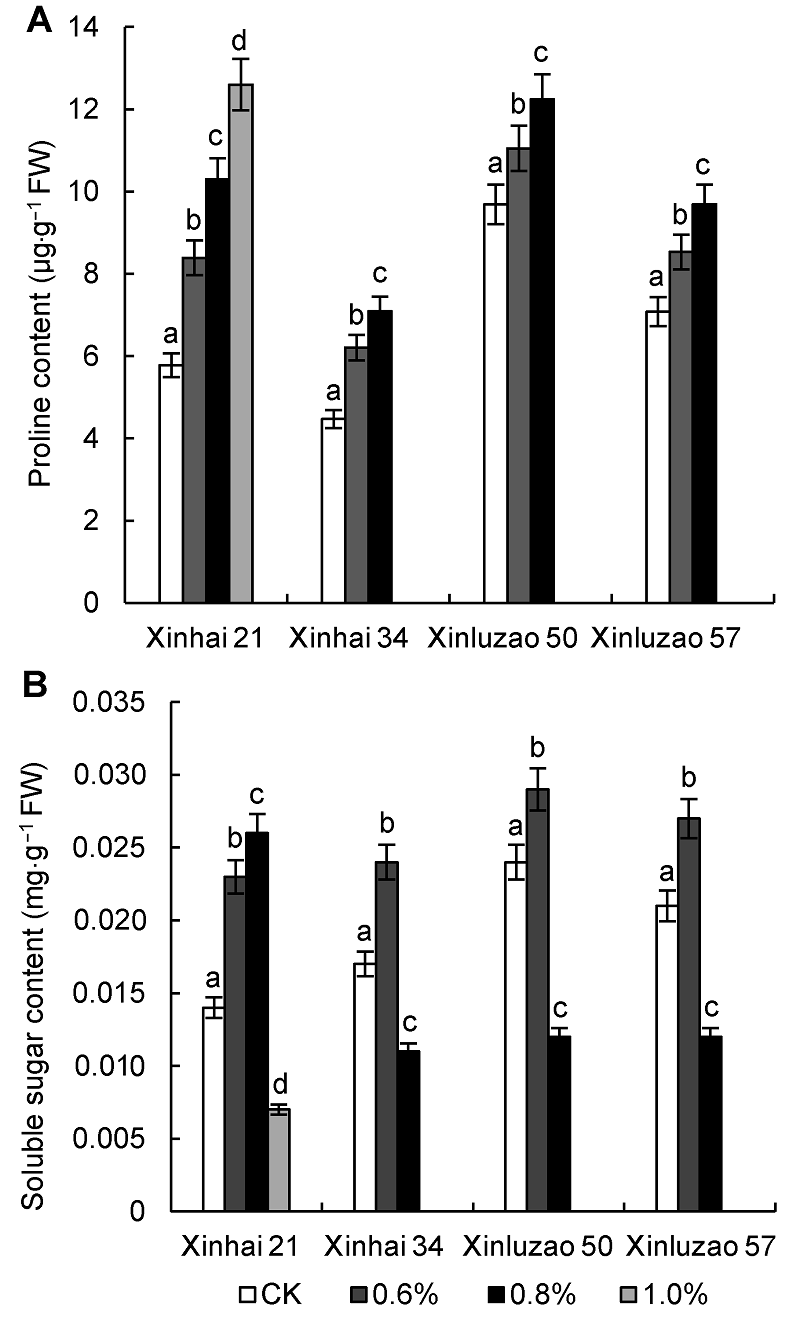
2.5 盐胁迫对海岛棉和陆地棉幼苗叶片渗透调节物质含量的影响由图2A可知, 在各浓度的盐胁迫条件下, 海岛棉和陆地棉幼苗叶片脯氨酸(Pro)含量总体呈上升趋势。新海21号、新海34号、新陆早57号和新陆早50号在0.8%盐胁迫下分别比相应对照提高了78.20%、58.61%、36.86%和26.32%。不同品种间, 新海21号和新海34号在各浓度盐胁迫下均保持比新陆早57号和新陆早50号更高的Pro含量, 有利于维持细胞的渗透调节能力, 阻止水分丧失, 说明海岛棉品种具有较强的抗盐
能力。
图2
Figure 2
下载原图ZIP
生成PPT
图2
盐胁迫对海岛棉和陆地棉幼苗叶片脯氨酸(A)和可溶性糖(B)含量的影响
Figure 2
Effects of salt stress on proline (A) and soluble sugar (B) content of Sea Island cotton and Upland cotton seedlings
由图2B可知, 盐胁迫导致4个供试品种幼苗叶片可溶性糖(SS)含量上升, 但在不同浓度盐胁迫下, SS含量变化趋势不同。新海34号、新陆早57号和新陆早50号在0.6%和0.8% NaCl胁迫下, 叶片SS含量均呈先升后降趋势, 且在0.6% NaCl胁迫时达到最高值, 与CK相比SS含量分别上升了41.18%、28.57%和20.83%, 新海21号则同比上升了64.29%; 而新海21号在高盐(1.0% NaCl)胁迫时SS含量迅速下降。总体看来, 新海21号和新海34号SS含量要高于新陆早57号和新陆早50号, 表明这2个品种对盐土环境有更强的抗性。
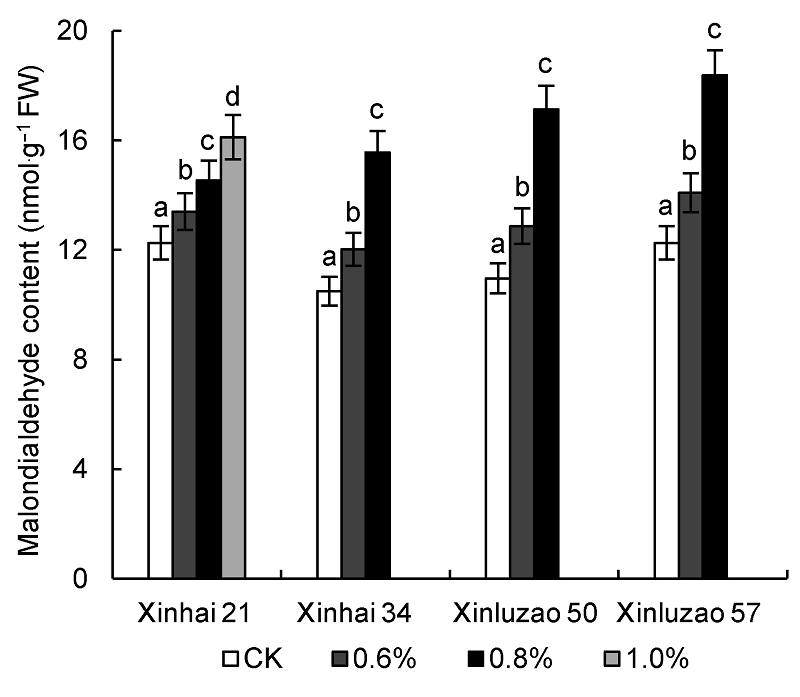
2.6 盐胁迫对海岛棉和陆地棉幼苗叶片丙二醛含量的影响丙二醛(MDA)是植物在逆境胁迫下组织或器官膜脂发生过氧化的最终分解产物。由图3可知, 盐胁迫导致海岛棉和陆地棉幼苗叶片MDA含量总体呈上升趋势。在0.8% NaCl胁迫下, 新海21号、新海34号、新陆早57号和新陆早50号与相应CK相比分别上升了18.60%、48.33%、49.92%和56.39%。
图3
Figure 3
下载原图ZIP
生成PPT
图3
盐胁迫对海岛棉和陆地棉幼苗叶片丙二醛(MDA)含量的影响
Figure 3
Effects of salt stress on malondialdehyde (MDA) content of Sea Island cotton and Upland cotton seedlings
2.7 讨论2.7.1 盐胁迫对棉花幼苗生长的影响
在自然条件下, 植物的生长发育经常遭受各种逆境(干旱、盐碱、高温和低温等)的影响, 使其生长发育受到抑制, 甚至导致植株死亡。有研究表明, 盐胁迫可能通过离子毒害和渗透胁迫抑制幼苗生长, 无论盐浓度的高低, 盐胁迫都会对棉花幼苗生长产生抑制作用, 且随着土壤盐浓度的增加, 抑制作用更加明显(谢德意等, 2000)。本研究表明, 盐胁迫显著抑制海岛棉与陆地棉幼苗生长。随着NaCl浓度的增加, 4个供试品种的株高、单株叶面积、鲜重与干重均下降, 尤其是在高盐胁迫下, 海岛棉和陆地棉幼苗第1片真叶展开和正常生长受到严重抑制, 且2个棉花栽培种的叶片状况差异明显, 即高浓度的盐胁迫使新陆早50号和新陆早57号幼苗第1片真叶展开时间延长, 与对照相比幼苗较长时间处在子叶期, 表现出叶片小、萎蔫、发黄和增厚等现象。研究表明, 高盐胁迫下只有Cl-主要分布在叶片中, 且叶片中Cl-的积累量与大豆(Glycine max)品种之间的耐盐性相关(罗庆云等, 2001)。本研究中, 海岛棉与陆地棉幼苗叶片在0.8%- 1.0% NaCl胁迫下生长不良, 且品种之间耐盐性差异较大, 可能是高浓度的盐在棉花叶片中分布, 影响叶片生长及其正常的生理生化反应所致。
2.7.2 盐胁迫对棉花幼苗叶片叶绿素与净光合速率的影响
叶绿素是与光合作用有关的重要色素。盐胁迫使叶绿素含量下降、叶绿素合成受阻、叶绿体超微结构和精细结构的形成受阻, 以及导致叶绿体超微结构发生变异(张景云和吴凤芝, 2009)。本研究显示, 盐胁迫导致海岛棉和陆地棉叶片叶绿素含量下降。高浓度(0.8%和1.0%)的盐胁迫下, 新海21号、新海34号和新陆早50号、新陆早57号幼苗叶片叶绿素(Chl)含量均显著下降。这可能是叶片经过高浓度盐胁迫后, 活性氧自由基产生和清除的平衡被打破, 叶片各项生理功能紊乱, 叶绿体色素生物合成酶功能受抑制, 降解速度远大于合成速度, 从而导致叶绿素含量迅速下降。
光合作用是复杂的光化学反应, 极易受到逆境因素的影响。光合速率(photosynthetic rate)是衡量植物光合作用受内外因素影响的常用指标。有研究表明, 盐胁迫影响植物幼苗叶片光合作用(王鑫等, 2010; 徐晨等, 2013)。本研究表明, 海岛棉(新海21号和新海34号)和陆地棉(新陆早50号和新陆早57号)幼苗叶片净光合速率(Pn)显著降低, 在高浓度盐胁迫下, 新海21号和新海34号Pn降低幅度比新陆早50号和新陆早57号低。
2.7.3 盐胁迫对棉花幼苗叶片抗氧化酶活性的影响
植物体内的抗氧化酶(POD和SOD等)可清除过多的活性氧, 保证细胞的正常功能和植株的正常生长。本研究表明, NaCl胁迫下海岛棉和陆地棉幼苗叶片中的丙二醛含量均呈上升趋势, 并且在高盐胁迫下, MDA积累更加显著, 尤其是新陆早50号和新陆早57号幼苗叶片MDA含量明显高于新海21号和新海34号, 这可能是由于新陆早50号和新陆早57号在高浓度盐胁迫时, 保护系统产生的调节能力低于海岛棉所致。海岛棉和陆地棉幼苗叶片POD和SOD活性随着土壤盐浓度的升高而增加, 以清除棉花体内多余的自由基, 保护细胞膜的正常功能, 保证机体的正常生长, 但超过一定盐浓度后, POD和SOD活性均降低, 这与马丽等(2008)和何林池等(2014)的研究结果一致。
2.7.4 盐胁迫对棉花幼苗叶片渗透调节物质含量的影响
脯氨酸是高等植物体内重要的有机小分子渗透调节物质, 具有很高的水溶性。在NaCl胁迫下, 海岛棉叶片脯氨酸含量显著增加(杨淑萍等, 2010)。可溶性糖是一种渗透调节物质, 是合成有机溶质的碳架和能量来源。在盐胁迫初期植物叶片中可溶性糖含量增加, 但随着盐浓度的增加和时间的延长其含量却降低, 可能是由于呼吸作用的增强和光合作用的衰竭所致(张海燕和赵可夫, 1998)。有研究表明, 构树(Brousson- etia kazinoki)幼苗可溶性糖含量在盐胁迫之前高, 盐胁迫之后变化不明显(杨帆等, 2008)。在适宜的NaCl条件下, 棉花叶片可溶性糖含量增加, 而在高盐浓度下叶片可溶性糖含量下降(张子学等, 2004)。本研究表明, 新海21号、新海34号、新陆早57号和新陆早50号在0.6%盐逆境中具有更灵敏和高效的渗透调节响应机制, 可有效抵御盐逆境。但高盐胁迫下, 4个供试品种Pro和SS积累较少, 尤其是新陆早50号和新陆早57号。
3 结论盐胁迫显著抑制海岛棉和陆地棉植株的生长, 但不同盐浓度胁迫下海岛棉和陆地棉受抑制的程度不同, 虽然在0.6% NaCl胁迫条件下, 海岛棉和陆地棉都表现出较好的耐盐性, 但0.8%和1.0%高盐胁迫对海岛棉和陆地棉幼苗都有很大的影响。陆地棉(新陆早57号和新陆早50号)对较高浓度NaCl胁迫较敏感, 其幼苗生长受到的抑制作用比海岛棉更大, 幼苗第1片真叶展开和正常生长受到严重抑制。海岛棉和陆地棉对NaCl胁迫的响应差异可能与其本身的基因型和盐浓度有关。棉花苗期叶片生长状况可以作为筛选耐盐性棉花品种的形态指标。
参考文献
文献选项
原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
1
... 幼苗生长24天后, 使用CARIS-2型便携式光合仪(PP Systems, USA)测定植株第1片真叶净光合速率(Pn)和蒸腾速率(Tr)等指标, 测定过程中光强为400 µmol·m-2·s-1, 环境温度(25±1)°C, 大气CO2浓度(Cr)变化范围为(410±10) µmol·L-1 (使用开放式气路, CO2浓度约为385 µmol·mol-1).选取长势均匀的植株, 设5次重复.采用愈创木酚法测定过氧化物酶(POD)活性(
1
... 盐渍土是指受土壤中盐碱成分作用的, 包括盐土、碱土以及其它不同程度盐化和碱化的各种类型土壤的统称(
耐盐差异性不同棉花品种的抗氧化酶活性及SNP/InDel分析
2014
披碱草属野生种质资源苗期耐盐性评价及相关生理机制研究
1
2008
... 盐渍土是指受土壤中盐碱成分作用的, 包括盐土、碱土以及其它不同程度盐化和碱化的各种类型土壤的统称(
1
...
NaCl胁迫对大麦幼苗生长及光合特性的影响
1
2014
... 研究表明, 光合作用对盐胁迫十分敏感, 即光合速率随着盐浓度的增加和胁迫时间的延长而明显下降(
大豆苗期耐盐性鉴定指标的检验
1
2001
... 在自然条件下, 植物的生长发育经常遭受各种逆境(干旱、盐碱、高温和低温等)的影响, 使其生长发育受到抑制, 甚至导致植株死亡.有研究表明, 盐胁迫可能通过离子毒害和渗透胁迫抑制幼苗生长, 无论盐浓度的高低, 盐胁迫都会对棉花幼苗生长产生抑制作用, 且随着土壤盐浓度的增加, 抑制作用更加明显(
NaCl胁迫对棉花幼苗生理特性的影响
1
2008
... 植物体内的抗氧化酶(POD和SOD等)可清除过多的活性氧, 保证细胞的正常功能和植株的正常生长.本研究表明, NaCl胁迫下海岛棉和陆地棉幼苗叶片中的丙二醛含量均呈上升趋势, 并且在高盐胁迫下, MDA积累更加显著, 尤其是新陆早50号和新陆早57号幼苗叶片MDA含量明显高于新海21号和新海34号, 这可能是由于新陆早50号和新陆早57号在高浓度盐胁迫时, 保护系统产生的调节能力低于海岛棉所致.海岛棉和陆地棉幼苗叶片POD和SOD活性随着土壤盐浓度的升高而增加, 以清除棉花体内多余的自由基, 保护细胞膜的正常功能, 保证机体的正常生长, 但超过一定盐浓度后, POD和SOD活性均降低,
1
... 幼苗生长24天后, 使用CARIS-2型便携式光合仪(PP Systems, USA)测定植株第1片真叶净光合速率(Pn)和蒸腾速率(Tr)等指标, 测定过程中光强为400 µmol·m-2·s-1, 环境温度(25±1)°C, 大气CO2浓度(Cr)变化范围为(410±10) µmol·L-1 (使用开放式气路, CO2浓度约为385 µmol·mol-1).选取长势均匀的植株, 设5次重复.采用愈创木酚法测定过氧化物酶(POD)活性(
盐胁迫对燕麦地上干物质积累及灌浆期光合特性的影响
1
2010
... 研究表明, 光合作用对盐胁迫十分敏感, 即光合速率随着盐浓度的增加和胁迫时间的延长而明显下降(
陆地棉萌发至三叶期不同生育阶段耐盐特性
1
2011
... 陆地棉(G. hirsutum)和海岛棉是棉花的2个重要栽培种.陆地棉丰产性好, 而海岛棉不仅纤维品质优良, 而且是盐碱地的主要栽培作物.前人的研究表明, 棉花虽然是盐碱地的先锋作物, 但其耐盐能力十分有限, 且各生育阶段的耐盐能力不同(
盐胁迫下高粱新生叶片结构和光合特性的系统调控
1
2010
... 光合作用是复杂的光化学反应, 极易受到逆境因素的影响.光合速率(photosynthetic rate)是衡量植物光合作用受内外因素影响的常用指标.有研究表明, 盐胁迫影响植物幼苗叶片光合作用(
植物耐盐生理生化指标研究进展
1
2014
... 研究表明, 光合作用对盐胁迫十分敏感, 即光合速率随着盐浓度的增加和胁迫时间的延长而明显下降(
盐胁迫对海岛棉种子萌发的影响
1
2008
... 陆地棉(G. hirsutum)和海岛棉是棉花的2个重要栽培种.陆地棉丰产性好, 而海岛棉不仅纤维品质优良, 而且是盐碱地的主要栽培作物.前人的研究表明, 棉花虽然是盐碱地的先锋作物, 但其耐盐能力十分有限, 且各生育阶段的耐盐能力不同(
盐胁迫对棉花种子萌发及幼苗生长的影响
1
2000
... 在自然条件下, 植物的生长发育经常遭受各种逆境(干旱、盐碱、高温和低温等)的影响, 使其生长发育受到抑制, 甚至导致植株死亡.有研究表明, 盐胁迫可能通过离子毒害和渗透胁迫抑制幼苗生长, 无论盐浓度的高低, 盐胁迫都会对棉花幼苗生长产生抑制作用, 且随着土壤盐浓度的增加, 抑制作用更加明显(
盐胁迫对不同水稻品种光合特性和生理生化特性的影响
1
2013
... 光合作用是复杂的光化学反应, 极易受到逆境因素的影响.光合速率(photosynthetic rate)是衡量植物光合作用受内外因素影响的常用指标.有研究表明, 盐胁迫影响植物幼苗叶片光合作用(
土壤盐胁迫对构树幼苗生理特性的影响
1
2008
... 脯氨酸是高等植物体内重要的有机小分子渗透调节物质, 具有很高的水溶性.在NaCl胁迫下, 海岛棉叶片脯氨酸含量显著增加(
盐胁迫对海岛棉不同基因型幼苗生长及生理生态特征的影响
2
2010
... 盐渍土是指受土壤中盐碱成分作用的, 包括盐土、碱土以及其它不同程度盐化和碱化的各种类型土壤的统称(
... 脯氨酸是高等植物体内重要的有机小分子渗透调节物质, 具有很高的水溶性.在NaCl胁迫下, 海岛棉叶片脯氨酸含量显著增加(
海岛棉不同基因型幼苗对盐胁迫的生理响应
2010
盐胁迫下棉花体内Na+的积累、分配及耐盐机制研究
1
2006
... 土壤中盐(NaCl)的过量积累可导致各种农作物减产(
盐分和水分胁迫对盐地碱蓬幼苗渗透调节效应的研究
1
1998
... 脯氨酸是高等植物体内重要的有机小分子渗透调节物质, 具有很高的水溶性.在NaCl胁迫下, 海岛棉叶片脯氨酸含量显著增加(
盐胁迫对黄瓜不同耐盐品种叶绿素含量和叶绿体超微结构的影响
1
2009
... 叶绿素是与光合作用有关的重要色素.盐胁迫使叶绿素含量下降、叶绿素合成受阻、叶绿体超微结构和精细结构的形成受阻, 以及导致叶绿体超微结构发生变异(
1
... 幼苗生长24天后, 使用CARIS-2型便携式光合仪(PP Systems, USA)测定植株第1片真叶净光合速率(Pn)和蒸腾速率(Tr)等指标, 测定过程中光强为400 µmol·m-2·s-1, 环境温度(25±1)°C, 大气CO2浓度(Cr)变化范围为(410±10) µmol·L-1 (使用开放式气路, CO2浓度约为385 µmol·mol-1).选取长势均匀的植株, 设5次重复.采用愈创木酚法测定过氧化物酶(POD)活性(
不同棉花品种NaCl胁迫的生理表现及其耐盐性筛选指标分析
1
2004
... 脯氨酸是高等植物体内重要的有机小分子渗透调节物质, 具有很高的水溶性.在NaCl胁迫下, 海岛棉叶片脯氨酸含量显著增加(
2
... 幼苗生长24天后, 使用CARIS-2型便携式光合仪(PP Systems, USA)测定植株第1片真叶净光合速率(Pn)和蒸腾速率(Tr)等指标, 测定过程中光强为400 µmol·m-2·s-1, 环境温度(25±1)°C, 大气CO2浓度(Cr)变化范围为(410±10) µmol·L-1 (使用开放式气路, CO2浓度约为385 µmol·mol-1).选取长势均匀的植株, 设5次重复.采用愈创木酚法测定过氧化物酶(POD)活性(
... ).采用硫代巴比妥酸比色法测定丙二醛(MDA)含量(
1
2010
... 土壤中盐(NaCl)的过量积累可导致各种农作物减产(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}