Abstract: The pollen tube growth and duration of double fertilization in Adonis amurensis were studied by paraffin section and fluorescence microscopy observation. A. amurensis has a wet column cap. At 1-2 h after pollination, the pollen grain is recognized by the stigma; at 2-4 h after pollination, the pollen grain begins germination; at 4-6 h after pollination, the pollen tube enters into the stigma. A. amurensis is porogamous. About 10 h after pollination, the sperm is released; about 30 h after pollination the egg nucleus and sperm nucleus begin fusing; at 7 d after pollination, a sperm fuses with the egg cell to form a zygote; dormant stage of the zygote is about 8 days long and at 15 d after pollination, the first mitotic division of zygote takes place. The two polar nuclei are not fused before fertilization. At 14-16 h after pollination, the sperm nucleus fuses with a polar nucleus; at 20-22 h after pollination, the sperm nucleus fuses with another polar nucleus and forms a primary endosperm nucleus. The type of double fertilization is premitotic syngamy. This study provides an understanding for the morphological change and corresponding timing of male and female cells in A. amurensis fertilization, confirms the duration of zygote dormancy, and enriches the knowledge of A. amurensis embryological information, which has great significance for A. amurensis breeding and genetic modification.
1 材料与方法1.1 材料实验材料为2010年从老山实验站引种至东北林业大学园林学院苗圃并正常生长5年的300株侧金盏花(Adonis amurensis Regel et Radde)植株。侧金盏花种植采用常规大田管理, 2014和2015年取材期间年昼夜平均温度分别为-1-11.5°C和-2-9°C, 日照条件良好。
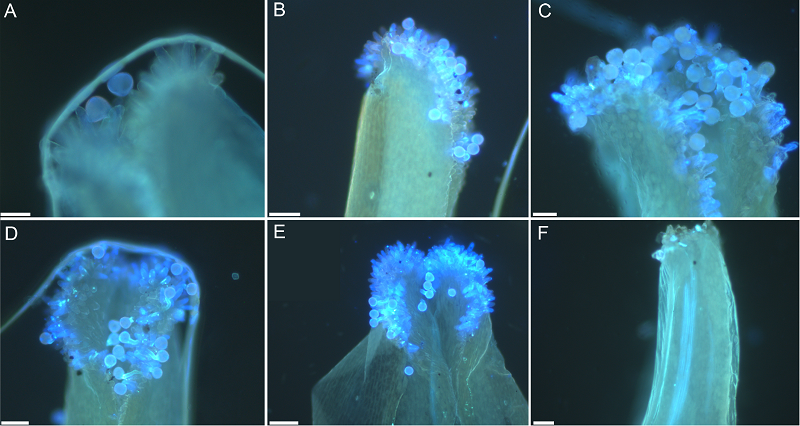
2 结果与讨论2.1 花粉萌发和花粉管生长侧金盏花花柱呈长圆柱形, 柱头具有明显的乳突状细胞, 乳突细胞在花朵开放后不久便开始分泌乳白色黏液, 进入可授期。侧金盏花的柱头为湿型。花粉和花粉管经脱色水溶性苯胺蓝染色后, 在LEICA DM2500显微镜下用荧光观察, 发现柱头上的花粉和花柱中花粉管均呈现出黄绿色荧光。人工授粉后1-2小时, 花粉粒与柱头识别(图1A, B), 此时花粉为三细胞型(图2A)。人工授粉后2-4小时, 花粉粒在柱头上萌发, 花粉管头部膨大, 2个精子进入花粉管(图2B, C), 大部分花粉粒均可萌发出花粉管(图1C, D)。虽然侧金盏花花粉粒有3条萌发沟, 但只有1条萌发沟能够萌发出花粉管。人工授粉后4-6小时, 花粉管通过乳突细胞进入柱头后继续向子房方向生长(图1E, F)。 图1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-480/img_1.png<b>图1</b> 侧金盏花花粉在柱头上萌发^(A), (B) 花粉落在柱头上(授粉后1-2小时); (C), (D) 花粉萌发产生花粉管(授粉后2-4小时); (E), (F) 花粉管进入柱头并向子房方向生长(授粉后4-6小时)。Bar=20 μm<br/><b>Figure 1</b> Pollen germinates on the stigma of <i>Adonis amurensis^</i>(A), (B) Pollen on stigmas (1-2 h after pollination); (C), (D) Pollen germination and pollen tube growth (2-4 h after pollination); (E), (F) Pollen tube enters into the stigma and grows toward the ovary (4-6 h after pollination). Bar=20 μm Figure 1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-480/img_1.png<b>图1</b> 侧金盏花花粉在柱头上萌发^(A), (B) 花粉落在柱头上(授粉后1-2小时); (C), (D) 花粉萌发产生花粉管(授粉后2-4小时); (E), (F) 花粉管进入柱头并向子房方向生长(授粉后4-6小时)。Bar=20 μm<br/><b>Figure 1</b> Pollen germinates on the stigma of <i>Adonis amurensis^</i>(A), (B) Pollen on stigmas (1-2 h after pollination); (C), (D) Pollen germination and pollen tube growth (2-4 h after pollination); (E), (F) Pollen tube enters into the stigma and grows toward the ovary (4-6 h after pollination). Bar=20 μm
图1 侧金盏花花粉在柱头上萌发^(A), (B) 花粉落在柱头上(授粉后1-2小时); (C), (D) 花粉萌发产生花粉管(授粉后2-4小时); (E), (F) 花粉管进入柱头并向子房方向生长(授粉后4-6小时)。Bar=20 μm Figure 1 Pollen germinates on the stigma of Adonis amurensis^(A), (B) Pollen on stigmas (1-2 h after pollination); (C), (D) Pollen germination and pollen tube growth (2-4 h after pollination); (E), (F) Pollen tube enters into the stigma and grows toward the ovary (4-6 h after pollination). Bar=20 μm
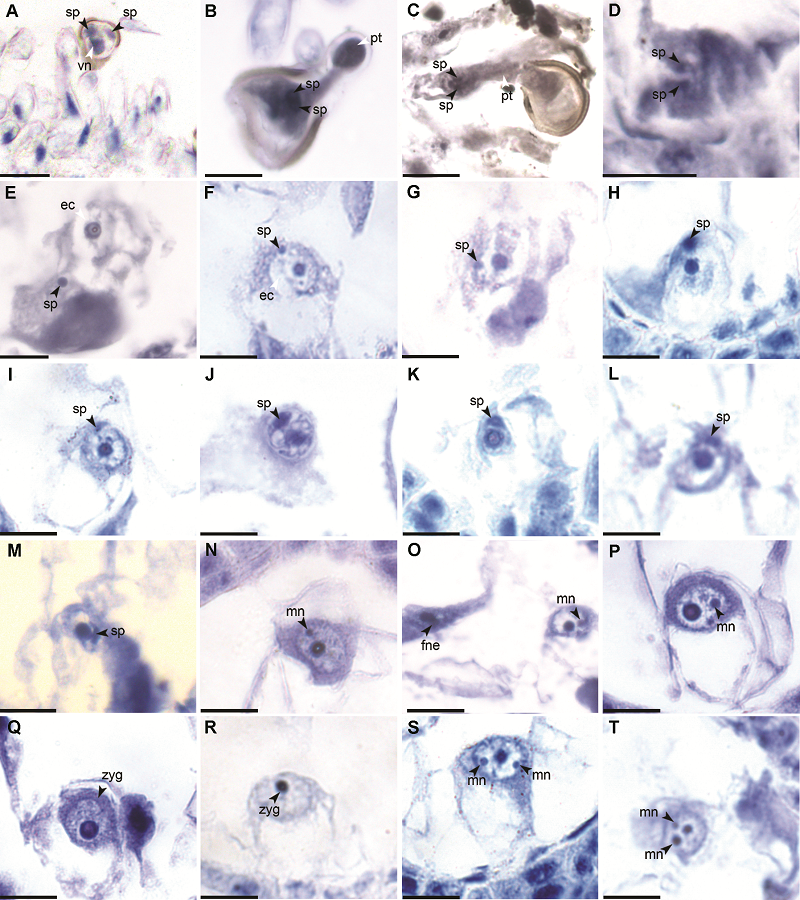
2.2 双受精过程2.2.1 卵细胞受精 侧金盏花人工授粉后10小时就有花粉管经由珠孔端进入胚囊, 1个助细胞破裂并释放花粉管的内容物与2个精子。故侧金盏花的受精模式为珠孔受精。刚释放的2个精核近似圆球形, 染色较深, 被细胞质鞘包围(图2D)。随后2个精子分开, 向卵细胞和极核方向移动(图2E), 在人工授粉后18小时的切片上可观察到精核的核膜刚刚接触到卵细胞膜(图2F), 随后精子的大部分表面紧紧地贴附在卵细胞膜上, 与卵细胞发生融合(图2G, H), 进入卵细胞细胞质内的精核渐渐向卵核的方向迁移(图2I, J), 而后精核紧紧附着于卵核的核膜之上, 形似凸透镜状, 与卵核的核膜发生融合, 在两核核膜相互接触的位置染色较深(图2K, L)。随后精核的染色质渐渐分散在卵核的一侧, 与卵核的染色质融合为一体(图2M)。接着, 卵核中出现了体积较小的雄性核仁, 小雄性核仁的体积渐渐增大, 最后与卵核核仁的体积基本相等(图2N-P)。此时, 有的胚囊中已经分布了数个胚乳游离核(图2O)。然后, 雄性核仁和卵核核仁相互靠近, 逐渐融合成1个体积较大的核仁, 形成合子(图2Q, R)。卵细胞的受精过程到此结束。由于精卵核的融合早于合子的有丝分裂, 所以侧金盏花受精作用的类型属于有丝分裂前配子融合型(胡适宜和朱瀓, 1979)。在整个卵细胞受精过程中, 可观察到在卵细胞内有3个大小不等的核仁(图2S, T), 这是由于多个花粉管进入胚囊, 并释放多个精子到胚囊中, 使2个精子先后与卵细胞融合(多精入卵为偶发现象, 出现概率约为1/1000)。 图2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-480/img_2.png<b>图2</b> 侧金盏花卵细胞受精^(A) 花粉落在柱头上; (B), (C) 花粉萌发, 花粉管头部膨大; (D) 退化助细胞中有2个精子; (E) 精子向卵细胞方向移动; (F) 精核核膜贴在卵细胞膜上; (G), (H) 精子与卵细胞膜融合; (I), (J) 精核逐渐移向卵核; (K), (L) 精核以凸透镜状贴在卵核核膜上; (M) 精核中的染色质分散到卵核中, 精核和卵核融为一体; (N)-(P) 卵核里出现雄性核仁并增大; (Q), (R) 合子; (S), (T) 合子中有2个雄性核仁。ec: 卵细胞; fne: 游离胚乳核; mn: 雄性核仁; pt: 花粉管; sp: 精核; zyg: 合子; vn: 营养核。Bar=10 μm<br/><b>Figure 2</b> Egg fertilization of <i>Adonis amurensis^</i>(A) The pollen falls onto the stigma; (B), (C) Pollen germinates, and the head of pollen tube expands; (D) Degenerated synergid contains 2 sperm cells; (E) Sperm moves to the egg cell; (F) The sperm nucleus membrane attaches to the egg cell membrane; (G), (H) Fusion of sperm and egg cell membrane; (I), (J) Sperm nucleus gradually moves to the egg nucleus; (K), (L) Sperm nucleus attaches to the egg nucleus membrane like convex lens; (M) Chromatin in the sperm nucleus is dispersed into the egg nucleus, sperm nucleus fuses with egg nucleus; (N)-(P) A small male nucleolus appears and enlarges in the egg nucleus; (Q), (R) Zygote; (S), (T) There are two small male nucleolus in zygote. ec: Egg cell; fne: Free nuclear endosperm; mn: Male nucleolus; pt: Pollen tube; sp: Sperm nucleus; zyg: Zygote; vn: Vegetative nucleus. Bar=10 μm Figure 2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-480/img_2.png<b>图2</b> 侧金盏花卵细胞受精^(A) 花粉落在柱头上; (B), (C) 花粉萌发, 花粉管头部膨大; (D) 退化助细胞中有2个精子; (E) 精子向卵细胞方向移动; (F) 精核核膜贴在卵细胞膜上; (G), (H) 精子与卵细胞膜融合; (I), (J) 精核逐渐移向卵核; (K), (L) 精核以凸透镜状贴在卵核核膜上; (M) 精核中的染色质分散到卵核中, 精核和卵核融为一体; (N)-(P) 卵核里出现雄性核仁并增大; (Q), (R) 合子; (S), (T) 合子中有2个雄性核仁。ec: 卵细胞; fne: 游离胚乳核; mn: 雄性核仁; pt: 花粉管; sp: 精核; zyg: 合子; vn: 营养核。Bar=10 μm<br/><b>Figure 2</b> Egg fertilization of <i>Adonis amurensis^</i>(A) The pollen falls onto the stigma; (B), (C) Pollen germinates, and the head of pollen tube expands; (D) Degenerated synergid contains 2 sperm cells; (E) Sperm moves to the egg cell; (F) The sperm nucleus membrane attaches to the egg cell membrane; (G), (H) Fusion of sperm and egg cell membrane; (I), (J) Sperm nucleus gradually moves to the egg nucleus; (K), (L) Sperm nucleus attaches to the egg nucleus membrane like convex lens; (M) Chromatin in the sperm nucleus is dispersed into the egg nucleus, sperm nucleus fuses with egg nucleus; (N)-(P) A small male nucleolus appears and enlarges in the egg nucleus; (Q), (R) Zygote; (S), (T) There are two small male nucleolus in zygote. ec: Egg cell; fne: Free nuclear endosperm; mn: Male nucleolus; pt: Pollen tube; sp: Sperm nucleus; zyg: Zygote; vn: Vegetative nucleus. Bar=10 μm
图2 侧金盏花卵细胞受精^(A) 花粉落在柱头上; (B), (C) 花粉萌发, 花粉管头部膨大; (D) 退化助细胞中有2个精子; (E) 精子向卵细胞方向移动; (F) 精核核膜贴在卵细胞膜上; (G), (H) 精子与卵细胞膜融合; (I), (J) 精核逐渐移向卵核; (K), (L) 精核以凸透镜状贴在卵核核膜上; (M) 精核中的染色质分散到卵核中, 精核和卵核融为一体; (N)-(P) 卵核里出现雄性核仁并增大; (Q), (R) 合子; (S), (T) 合子中有2个雄性核仁。ec: 卵细胞; fne: 游离胚乳核; mn: 雄性核仁; pt: 花粉管; sp: 精核; zyg: 合子; vn: 营养核。Bar=10 μm Figure 2 Egg fertilization of Adonis amurensis^(A) The pollen falls onto the stigma; (B), (C) Pollen germinates, and the head of pollen tube expands; (D) Degenerated synergid contains 2 sperm cells; (E) Sperm moves to the egg cell; (F) The sperm nucleus membrane attaches to the egg cell membrane; (G), (H) Fusion of sperm and egg cell membrane; (I), (J) Sperm nucleus gradually moves to the egg nucleus; (K), (L) Sperm nucleus attaches to the egg nucleus membrane like convex lens; (M) Chromatin in the sperm nucleus is dispersed into the egg nucleus, sperm nucleus fuses with egg nucleus; (N)-(P) A small male nucleolus appears and enlarges in the egg nucleus; (Q), (R) Zygote; (S), (T) There are two small male nucleolus in zygote. ec: Egg cell; fne: Free nuclear endosperm; mn: Male nucleolus; pt: Pollen tube; sp: Sperm nucleus; zyg: Zygote; vn: Vegetative nucleus. Bar=10 μm
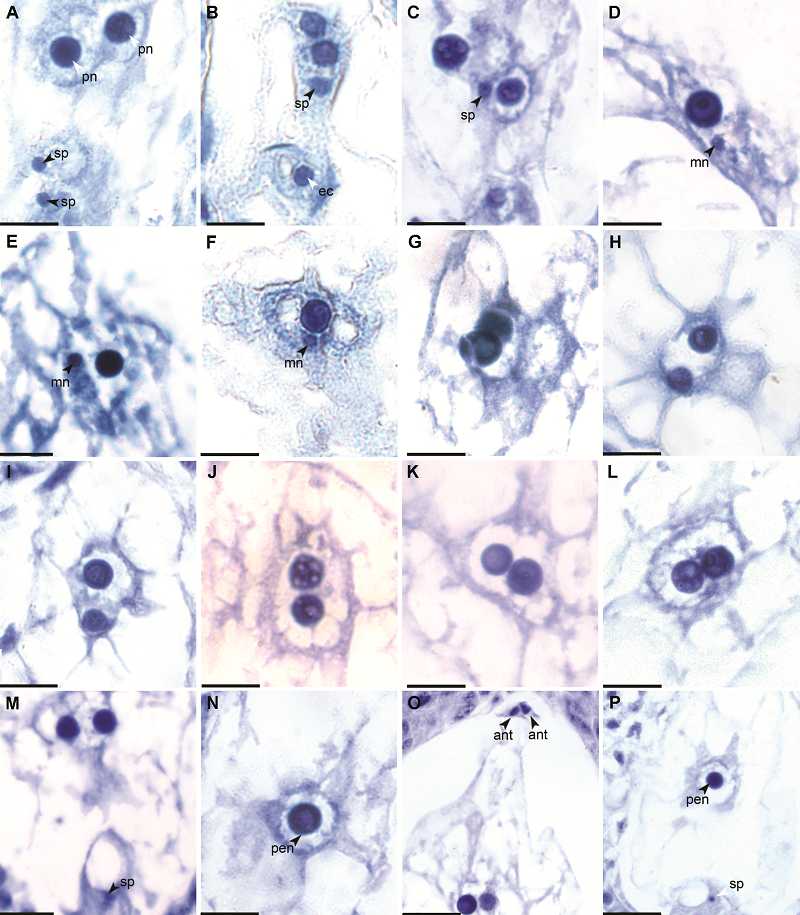
2.2.2 极核受精 释放后的2个精子分别移至卵细胞和2个极核的附近, 其中1个精子游向极核的速度要快于另1个精子游向卵细胞的速度(图3A)。精核与极核的融合速度更快(图3M, P)。在侧金盏花中, 受精前2个极核紧紧贴在一起并不发生融合, 另1个精核移至2个极核附近, 贴附于距离卵细胞较近的1个极核之上(图3B, C), 两者的核膜相互融合之后, 极核内出现小雄性核仁(图3D)。雄性核仁的体积逐渐由小变大(图3E, F), 雄性核仁和极核的核仁融合成1个大的核仁, 形成受精极核(图3G)。随后, 受精极核与另外1个极核发生融合。首先两者间的核膜逐渐消失, 核质相互融合(图3H, I, M), 然后2个极核的核仁慢慢靠近, 融合(图3J-L), 最后形成1个初生胚乳核, 内含1个体积较大的核仁(图3N, P)。初生胚乳核位于胚囊的中央。在极核受精过程中, 反足细胞逐渐退化(图3O)。至此, 侧金盏花的双受精过程完成。 图3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-480/img_3.png<b>图3</b> 侧金盏花极核受精^(A) 2个精子分别移向极核和卵核; (B), (C) 精核紧紧贴附于极核之上; (D)-(F) 极核内出现雄性核仁且其体积逐渐增大; (G) 雄性核仁与极核核仁融合; (H), (I) 2个极核间的核膜消失, 核质融为一体; (J)-(L) 2个核仁慢慢靠近、融合; (M) 2个极核融合时, 卵细胞还未开始受精; (N) 初生胚乳核; (O) 2个极核融合过程中, 反足细胞退化; (P) 初生胚乳核形成时, 卵核还未开始受精。ec: 卵细胞; mn: 雄性核仁; pn: 极核; pen: 初生胚乳核; ant: 反足细胞; sp: 精核。Bar=10 μm<br/><b>Figure 3</b> Polar nuclei fertilization of <i>Adonis amurensis^</i>(A) Two sperm cells move to the polar nucleus and the egg nucleus, respectively; (B), (C) Sperm nucleus attaches on polar nucleus; (D)-(F) Male nucleolus appears and enlarges in polar nucleus; (G) The fusion of the male nucleus and the polar nucleus; (H), (I) The nuclear membrane of 2 polar nuclei disappear and the karyoplasm fuses; (J)-(L) Two nucleoli gradually come near and fuse; (M) When the two polar nuclei merge, the egg cell is still unfertilized; (N) Primary endosperm nucleus; (O) Antipodal cells degenerate when two polar nuclei fusing; (P) When the primary endosperm nucleus forms, the egg cell is still unfertilized. ec: Egg cell; mn: Male nucleolus; pn: Polar nucleus; pen: Primary endosperm nucleus; ant: Antipodal cell; sp: Sperm nucleus. Bar=10 μm Figure 3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-480/img_3.png<b>图3</b> 侧金盏花极核受精^(A) 2个精子分别移向极核和卵核; (B), (C) 精核紧紧贴附于极核之上; (D)-(F) 极核内出现雄性核仁且其体积逐渐增大; (G) 雄性核仁与极核核仁融合; (H), (I) 2个极核间的核膜消失, 核质融为一体; (J)-(L) 2个核仁慢慢靠近、融合; (M) 2个极核融合时, 卵细胞还未开始受精; (N) 初生胚乳核; (O) 2个极核融合过程中, 反足细胞退化; (P) 初生胚乳核形成时, 卵核还未开始受精。ec: 卵细胞; mn: 雄性核仁; pn: 极核; pen: 初生胚乳核; ant: 反足细胞; sp: 精核。Bar=10 μm<br/><b>Figure 3</b> Polar nuclei fertilization of <i>Adonis amurensis^</i>(A) Two sperm cells move to the polar nucleus and the egg nucleus, respectively; (B), (C) Sperm nucleus attaches on polar nucleus; (D)-(F) Male nucleolus appears and enlarges in polar nucleus; (G) The fusion of the male nucleus and the polar nucleus; (H), (I) The nuclear membrane of 2 polar nuclei disappear and the karyoplasm fuses; (J)-(L) Two nucleoli gradually come near and fuse; (M) When the two polar nuclei merge, the egg cell is still unfertilized; (N) Primary endosperm nucleus; (O) Antipodal cells degenerate when two polar nuclei fusing; (P) When the primary endosperm nucleus forms, the egg cell is still unfertilized. ec: Egg cell; mn: Male nucleolus; pn: Polar nucleus; pen: Primary endosperm nucleus; ant: Antipodal cell; sp: Sperm nucleus. Bar=10 μm
图3 侧金盏花极核受精^(A) 2个精子分别移向极核和卵核; (B), (C) 精核紧紧贴附于极核之上; (D)-(F) 极核内出现雄性核仁且其体积逐渐增大; (G) 雄性核仁与极核核仁融合; (H), (I) 2个极核间的核膜消失, 核质融为一体; (J)-(L) 2个核仁慢慢靠近、融合; (M) 2个极核融合时, 卵细胞还未开始受精; (N) 初生胚乳核; (O) 2个极核融合过程中, 反足细胞退化; (P) 初生胚乳核形成时, 卵核还未开始受精。ec: 卵细胞; mn: 雄性核仁; pn: 极核; pen: 初生胚乳核; ant: 反足细胞; sp: 精核。Bar=10 μm Figure 3 Polar nuclei fertilization of Adonis amurensis^(A) Two sperm cells move to the polar nucleus and the egg nucleus, respectively; (B), (C) Sperm nucleus attaches on polar nucleus; (D)-(F) Male nucleolus appears and enlarges in polar nucleus; (G) The fusion of the male nucleus and the polar nucleus; (H), (I) The nuclear membrane of 2 polar nuclei disappear and the karyoplasm fuses; (J)-(L) Two nucleoli gradually come near and fuse; (M) When the two polar nuclei merge, the egg cell is still unfertilized; (N) Primary endosperm nucleus; (O) Antipodal cells degenerate when two polar nuclei fusing; (P) When the primary endosperm nucleus forms, the egg cell is still unfertilized. ec: Egg cell; mn: Male nucleolus; pn: Polar nucleus; pen: Primary endosperm nucleus; ant: Antipodal cell; sp: Sperm nucleus. Bar=10 μm
2.2.3 双受精过程经历时间 侧金盏花从传粉到完成受精所经历的时间是从人工授粉开始, 到雌雄配子融合完成为止。栽培环境以及气候条件也影响经历时间的长短, 尤其是生长温度对其影响最大。因此, 本实验记录的双受精过程中各阶段所需要的时间(表1)仅为近似值。从表1可以看出, 精核移向极核的速度要快于精核移向卵核的速度, 且精卵核融合速度慢于精核与极核的融合。确定传粉至完成受精所需要的时间, 对于其遗传育种研究十分重要。 表1 Table 1 表1 表1 侧金盏花双受精作用时间表 Table 1 The duration of double fertilization in Adonis amurensis
After pollination
Egg fertilization
After pollination
Polar nuclei fertilization
1-2 h
Pollen recognition
1-2 h
Pollen recognition
2-4 h
Pollen germination
2-4 h
Pollen germination
4-6 h
Pollen tube enters into stigma
4-6 h
Pollen tube enter into stigma
10 h
Sperm enters into synergid
10 h
Sperm enters into synergid
10-20 h
Sperm moves to egg cell
10-12 h
Sperm moves to polar nucleus
20 h
The sperm nucleus attaches to the egg cell membrane
12-14 h
The sperm nuclei attache to the polar nucleus membrane
30 h
Egg nucleus fuses with sperm nucleus
14-16 h
Egg nucleus fuses with sperm nucleus
72 h
Male nucleolus appears in the egg nucleus
16-18 h
Male nucleolus appears in the polar nucleus
7 d
Zygote formed
18-20 h
Fertilized polar nucleus formed
15 d
The division of zygote
20-22 h
The endosperm nucleus formed
表1 侧金盏花双受精作用时间表 Table 1 The duration of double fertilization in Adonis amurensis
CunninghamFX, GanttE (2005). A study in scarlet: enzymes of ketocarotenoid biosynthesis in the flowers ofAdonis aestivalis. Plant J 41, 478-492. [本文引用: 1]
[18]
FuY, YuanM, HuangBQ, YangHY, ZeeSY, O’BrienTP (2000). Change in actin organization in the living egg apparatus of Torenia fournieri during fertilization. Sex Plant Rep 12, 315-322. [本文引用: 1]
[19]
HeynCC, PazyB (1989). The annual species of Adonis (Ranunculaceae)—a polyploid complex. Plant Syst Evol 168, 181-193. [本文引用: 1]
[20]
ZhangZ, RussellSD (1999). Sperm cell surface characte- ristics of Plumbago zeylanica L. in relation to transport in the embryo sac. Planta 208, 539-544. [本文引用: 1]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}