Abstract: Gerbera hybrida has been used as model plant to study the evolution and development of composite inflorescences. To establish an efficient protoplast isolation and transient transformation system in G. hybrida, leaves of cultivar ‘Shenzhen No. 5’ in vitro seedlings were used as materials and the effects of different concentrations of cellulase R10 and macerozyme were tested to screen a suitable preparation method. The best enzyme solution concentration consisted of cellulase R10 2.0% (w/v), macerozyme 0.3% (w/v), and 0.4 mol·L-1 mannitol and more than 2.2×107∙g-1 FW protoplasts with high activity were obtained after 4 h treatment under 25°C condition. Subcellular localization and protein-protein interaction assays were performed using these protoplasts and revealed 80% transformation rate. Collectively, this study established an efficient protoplast isolation and transient assay system that can provide an important platform to facilitate gene function studies in G. hybrida.
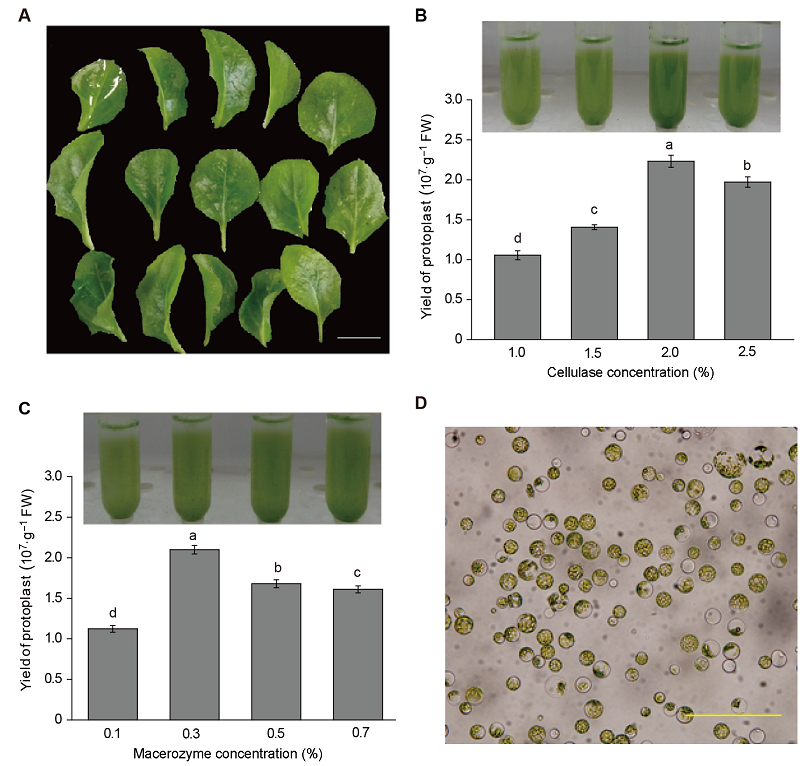
图1 非洲菊原生质体提取效果^(A) 扩繁培养2个月的S5非洲菊组培苗叶片(Bar=1 cm); (B) 在离析酶浓度为0.3%时, 不同浓度纤维素酶对非洲菊原生质体提取产量的影响; (C) 在纤维素酶浓度为2.0%时, 不同浓度离析酶对原生质体提取产量的影响; (D) 光学显微镜下原生质体形态观察, 原生质体来自2.0%纤维素酶+0.3%离析酶的提取结果(Bar=200 μm) Figure 1 Isolation efficiency of protoplast in Gerbera hybrid^(A) Leaves isolated from two-month-old in vitro seedlings (Bar=1 cm); (B) Protoplast yields under different cellulase R10 concentrations with 0.3% macerozyme; (C) Protoplast yields under different macerozyme concentrations with 2.0% cellulase R10; (D) Morphological characteristics of protoplasts, isolated from 2.0% cellulase R10 +0.3% macerozyme, observed by optical microscope (Bar=200 μm)
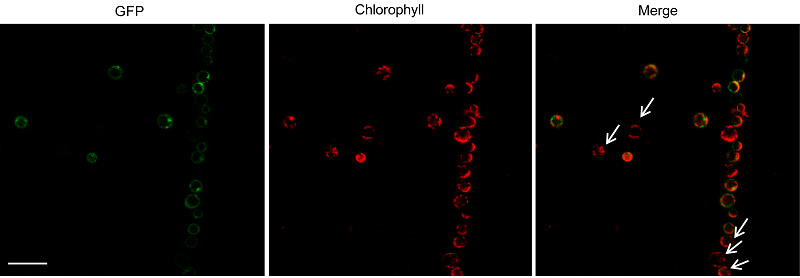
为了检测原生质体是否具有足够的活力, 将其经过处理制备成感受态后转化GFP空载体质粒。培养36小时, 在激光共聚焦扫描显微镜下观察。结果显示, 原生质体形态规则, 基本没有出现破裂的情况, 并且大部分原生质体均成功转化了GFP, 原生质体中表达GFP的区域呈现绿色荧光, 而未表达GFP的区域则不发光, 转化效率达80% (图2)。为了检测非洲菊叶肉原生质体进行亚细胞定位的情况, 将GFP和YFP空载体质粒分别转化原生质体, 经上述步骤培养和观察, 结果显示, 转化成功后, 分别在细胞质、质膜及细胞核等部位表达出GFP和YFP蛋白(图3A, B)。由此可见, 在2.0%纤维素酶+0.3%离析酶的酶组合下提取非洲菊叶片原生质体适用于蛋白的亚细胞定位研究。 图2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-511/img_2.png<b>图2</b> 非洲菊原生质体转化效率^图示激光共聚焦扫描显微镜下观察原生质体转化效率。从左到右分别为GFP荧光、叶绿体自发荧光及两者叠加图。箭头示未转化成功的原生质体。Bar=100 μm<br/><b>Figure 2</b> Transformation efficiency of protoplast in <i>Gerbera hybrid^</i>Transformation efficiency of protoplast observed by laser confocal scanning microscope. From left to right are GFP fluorescence, chloroplast autofluorescence, and a merge of above. Arrows indicate the protoplasts with unsuccessful transformation. Bar=100 μm Figure 2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-511/img_2.png<b>图2</b> 非洲菊原生质体转化效率^图示激光共聚焦扫描显微镜下观察原生质体转化效率。从左到右分别为GFP荧光、叶绿体自发荧光及两者叠加图。箭头示未转化成功的原生质体。Bar=100 μm<br/><b>Figure 2</b> Transformation efficiency of protoplast in <i>Gerbera hybrid^</i>Transformation efficiency of protoplast observed by laser confocal scanning microscope. From left to right are GFP fluorescence, chloroplast autofluorescence, and a merge of above. Arrows indicate the protoplasts with unsuccessful transformation. Bar=100 μm
图2 非洲菊原生质体转化效率^图示激光共聚焦扫描显微镜下观察原生质体转化效率。从左到右分别为GFP荧光、叶绿体自发荧光及两者叠加图。箭头示未转化成功的原生质体。Bar=100 μm Figure 2 Transformation efficiency of protoplast in Gerbera hybrid^Transformation efficiency of protoplast observed by laser confocal scanning microscope. From left to right are GFP fluorescence, chloroplast autofluorescence, and a merge of above. Arrows indicate the protoplasts with unsuccessful transformation. Bar=100 μm
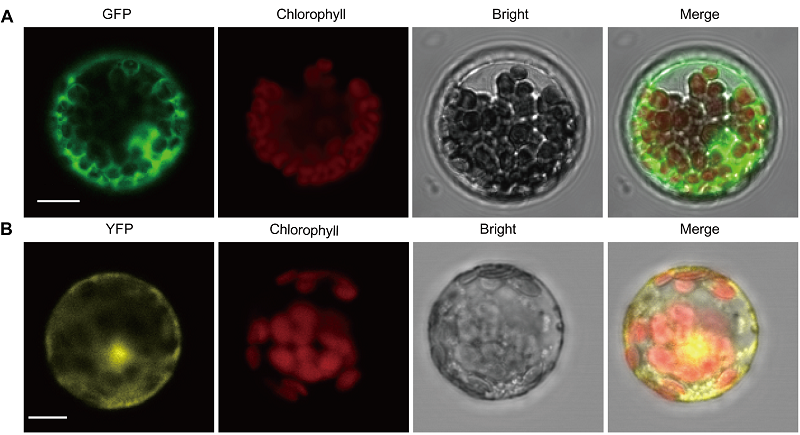
图3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-511/img_3.png<b>图3</b> GFP和YFP在非洲菊原生质体中的亚细胞定位^(A) 激光共聚焦扫描显微镜下转GFP空载体的原生质体, 从左到右分别为GFP荧光、叶绿体自发荧光、白光及叠加图; (B) 激光共聚焦扫描显微镜下转YFP空载体的原生质体, 从左到右分别为YFP荧光、叶绿体自发荧光、白光及叠加图。Bar=10 μm<br/><b>Figure 3</b> Subcellular localization of GFP and YFP in protoplasts from <i>Gerbera hybrid^</i>(A) Subcellular localization of GFP by laser confocal scanning microscope, from left to right are GFP fluorescence, chloroplast autofluorescence, bright, and a merge of above; (B) Subcellular localization of YFP by laser confocal scanning microscope, from left to right are YFP fluorescence, chloroplast autofluorescence, bright, and a merge of above. Bar=10 μm Figure 3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-511/img_3.png<b>图3</b> GFP和YFP在非洲菊原生质体中的亚细胞定位^(A) 激光共聚焦扫描显微镜下转GFP空载体的原生质体, 从左到右分别为GFP荧光、叶绿体自发荧光、白光及叠加图; (B) 激光共聚焦扫描显微镜下转YFP空载体的原生质体, 从左到右分别为YFP荧光、叶绿体自发荧光、白光及叠加图。Bar=10 μm<br/><b>Figure 3</b> Subcellular localization of GFP and YFP in protoplasts from <i>Gerbera hybrid^</i>(A) Subcellular localization of GFP by laser confocal scanning microscope, from left to right are GFP fluorescence, chloroplast autofluorescence, bright, and a merge of above; (B) Subcellular localization of YFP by laser confocal scanning microscope, from left to right are YFP fluorescence, chloroplast autofluorescence, bright, and a merge of above. Bar=10 μm
图3 GFP和YFP在非洲菊原生质体中的亚细胞定位^(A) 激光共聚焦扫描显微镜下转GFP空载体的原生质体, 从左到右分别为GFP荧光、叶绿体自发荧光、白光及叠加图; (B) 激光共聚焦扫描显微镜下转YFP空载体的原生质体, 从左到右分别为YFP荧光、叶绿体自发荧光、白光及叠加图。Bar=10 μm Figure 3 Subcellular localization of GFP and YFP in protoplasts from Gerbera hybrid^(A) Subcellular localization of GFP by laser confocal scanning microscope, from left to right are GFP fluorescence, chloroplast autofluorescence, bright, and a merge of above; (B) Subcellular localization of YFP by laser confocal scanning microscope, from left to right are YFP fluorescence, chloroplast autofluorescence, bright, and a merge of above. Bar=10 μm
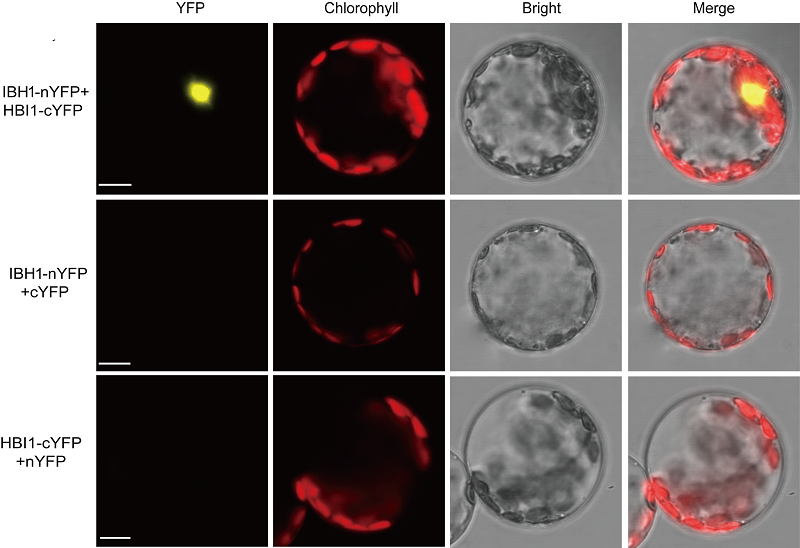
2.2 非洲菊叶片原生质体的共转化利用原生质体进行蛋白相互作用研究是常见的策略。为了验证非洲菊原生质体具有该潜力, 选取拟南芥中能够相互作用的2个转录因子IBH1和HBI1 (Bai et al., 2012; Fan et al., 2014; Malinovsky et al., 2014)探究上述原生质体提取体系能否应用于遗传共转化实验。利用IBH1-nYFP和HBI1-cYFP共转化非洲菊原生质体, 通过原生质体荧光定位, 能直接观察到IBH1和HBI1互作所形成的复合体在原生质体中的定位情况。结果显示, 与IBH1-nYFP+cYFP和HBI1-cYFP+nYFP阴性对照相比, 在共转化IBH1-nYFP和HBI1-cYFP质粒的细胞核中能观察到强烈的YFP荧光信号(图4)。 图4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-511/img_4.png<b>图4</b> 用双分子荧光互补技术(BiFC)分析IBH1和HBI1蛋白互作^IBH1-nYFP与HBI1-cYFP分别代表IBH1和HBI1连接在YFP的N端片段和C端片段上, IBH1-nYFP+cYFP和HBI1-cYFP+nYFP为阴性对照。Bar=10 μm<br/><b>Figure 4</b> Analysis of interaction of IBH1 and HBI1 by bimolecular fluorescence complementation (BiFC)^IBH1-nYFP and HBI1-cYFP represent IBH1 and HBI1 connected to the N-terminal fragment and the C-terminal fragment of YFP, respectively. IBH1-nYFP+cYFP and HBI1-cYFP+nYFP were used as negative control. Bar=10 μm Figure 4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-511/img_4.png<b>图4</b> 用双分子荧光互补技术(BiFC)分析IBH1和HBI1蛋白互作^IBH1-nYFP与HBI1-cYFP分别代表IBH1和HBI1连接在YFP的N端片段和C端片段上, IBH1-nYFP+cYFP和HBI1-cYFP+nYFP为阴性对照。Bar=10 μm<br/><b>Figure 4</b> Analysis of interaction of IBH1 and HBI1 by bimolecular fluorescence complementation (BiFC)^IBH1-nYFP and HBI1-cYFP represent IBH1 and HBI1 connected to the N-terminal fragment and the C-terminal fragment of YFP, respectively. IBH1-nYFP+cYFP and HBI1-cYFP+nYFP were used as negative control. Bar=10 μm
图4 用双分子荧光互补技术(BiFC)分析IBH1和HBI1蛋白互作^IBH1-nYFP与HBI1-cYFP分别代表IBH1和HBI1连接在YFP的N端片段和C端片段上, IBH1-nYFP+cYFP和HBI1-cYFP+nYFP为阴性对照。Bar=10 μm Figure 4 Analysis of interaction of IBH1 and HBI1 by bimolecular fluorescence complementation (BiFC)^IBH1-nYFP and HBI1-cYFP represent IBH1 and HBI1 connected to the N-terminal fragment and the C-terminal fragment of YFP, respectively. IBH1-nYFP+cYFP and HBI1-cYFP+nYFP were used as negative control. Bar=10 μm
AnCl, SawadaA, KawaguchiY, FukusakiE, KobayashiA (2005). Transient RNAi induction against endogenous genes in Arabidopsis protoplasts using in vitro prepared double-stranded RNA.Biosci Biotechnol Biochem 69, 415-418. [本文引用: 1]
[15]
BaiMY, FanM, OhE, WangZY (2012). A triple helix-loop- helix/basic helix-loop-helix cascade controls cell elonga- tion downstream of multiple hormonal and environmental signaling pathways in Arabidopsis.Plant Cell 24, 4917-4929. [本文引用: 1]
[16]
BroholmSK, TahtiharjuS, LaitinenRA, AlbertVA, TeeriTH, ElomaaP (2008). A TCP domain transcription factor controls flower type specification along the radial axis of theGerbera(Asteraceae) inflorescence. Proc Natl Acad Sci USA 105, 9117-9122. [本文引用: 2]
[17]
CaoH, GuoS, XuY, JiangK, JonesAM, ChongK (2011). Reduced expression of a gene encoding a golgi localized monosaccharide transporter (OsGMST1) confers hypersensitivity to salt in rice (Oryza sativa). J Exp Bot 62, 4595-4604.
[18]
DavidH, LaigneauC, DavidA (1989). Growth and soluble proteins of cell cultures derived from explants and protoplasts ofPinus pinaster cotyledons. Tree Physiol 5, 497-506. [本文引用: 1]
[19]
ElomaaP, UimariA, MehtoM, AlbertVA, LaitinenRA, TeeriTH (2003). Activation of anthocyanin biosynthesis in Gerbera hybrida(Asteraceae) suggests conserved protein-protein and protein-promoter interactions between the anciently diverged monocots and eudicots.Plant Physiol 133, 1831-1842.
[20]
FanM, BaiMY, KimJG, WangT, OhE, ChenL, ParkCH, SonSH, KimSK, MudgettMB, WangZY (2014). The bHLH transcription factor HBI1 mediates the trade-off between growth and pathogen-associated molecular pat- tern-triggered immunity in Arabidopsis.Plant Cell 26, 828-841. [本文引用: 1]
[21]
FontesN, SilvaR, VignaultC, LecourieuxF, GerósH, DelrotS (2010). Purification and functional characterization of protoplasts and intact vacuoles from grape cells.BMC Res Notes 3, 19. [本文引用: 1]
[22]
GuoJ, Morrell-FalveyJL, LabbéJL, MucheroW, KalluriUC, TuskanGA, ChenJG (2012). Highly efficient isolation of Populus mesophyll protoplasts and its application in transient expression assays.PLoS One 7, e44908. [本文引用: 1]
[23]
JonesAM, ChattopadhyayA, ShuklaM, ZońJ, SaxenaPK (2012). Inhibition of phenylpropanoid biosynthesis increases cell wall digestibility, protoplast isolation, and facilitates sustained cell division in American elm (Ulmus americana).BMC Plant Biol 12, 1-11. [本文引用: 1]
[24]
LaitinenRA, PollanenE, TeeriTH, ElomaaP, KotilainenM (2007). Transcriptional analysis of petal organogenesis in Gerbera hybrida.Planta 226, 347-360. [本文引用: 1]
[25]
LiLF, ZhangWB, ZhangLL, LiN, PengJZ, WangYQ, ZhongCM, YangYP, SunSL, LiangS, WangXJ (2015). Transcriptomic insights into antagonistic effects of gibbe- rellin and abscisic acid on petal growth in Gerbera hybrida.Front Plant Sci 6, 168. [本文引用: 1]
LungSC, SmithMD, ChuongSD (2015). Isolation of chloroplasts from plant protoplasts.Cold Spring Harb Pro- toc 2015, 895-899. [本文引用: 1]
[28]
MalinovskyFG, BatouxM, SchwessingerB, YounJH, StransfeldL, WinJ, KimSK, ZipfelC (2014). Antago- nistic regulation of growth and immunity by the Arabidopsis basic helix-loop-helix transcription factor HOMOLOG OF BRA- SSINOSTEROID ENHANCED EXPRESSION2 INTERACTING WITH INCREASED LEAF INCLINATION1 BINDING bHLH1.Plant Physiol 164, 1443-1455. [本文引用: 1]
[29]
MazareiM, Al-AhmadH, RudisMR, Jr StewartCN (2008). Protoplast isolation and transient gene expression in switchgrass,Panicum virgatum L. Biotech J 3, 354-359. [本文引用: 1]
[30]
SheenJ (2001). Signal transduction in maize and Arabidopsis mesophyll protoplasts.Plant Physiol 127, 1466-1475. [本文引用: 1]
[31]
TahtiharjuS, RijpkemaAS, VetterliA, AlbertVA, TeeriTH, ElomaaP (2012). Evolution and diversification of the CYC/TB1 gene family in Asteraceae—a comparative study in Gerbera(Mutisieae) and sunflower (Heliantheae). Mol Biol Evol 29, 1155-1166. [本文引用: 1]
[32]
TangZ, FanX, LiQ, FengH, MillerAJ, ShenQ, XuG (2012). Knockdown of a rice stelar nitrate transporter alters long-distance translocation but not root influx.Plant Phy- siol 160, 2052-2063.
[33]
TeeriTH, ElomaaP, KotilainenM, AlbertVA (2006). Mining plant diversity:Gerbera as a model system for plant developmental and biosynthetic research. Bioessays 28, 756-767. [本文引用: 1]
[34]
YooSD, ChoYH, SheenJ (2007). Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis.Nat Protoc 2, 1565-1572. [本文引用: 5]
... 植物原生质体(protoplast)是指通过质壁分离能够分开的那部分仅由细胞膜包围的具有活力的细胞物质(周波等, 2008; 于晓玲等, 2009).细胞壁对植物细胞具有支撑和保护作用, 其主要成分是果胶质、纤维素、半纤维素与少量蛋白质(刘敏和顾志敏, 2014).原生质体能摄入DNA、质粒、病毒、细胞器等外源物质(An et al., 2005), 又可用于研究蛋白定位, 还可以验证蛋白之间的互作, 检测其互作强弱, 定位互作蛋白的位点, 从而为推断蛋白质的生物学功能研究提供必要的基础信息, 因此在细胞工程和基因工程中应用广泛(刘敏和顾志敏, 2014).在模式植物拟南芥(Arab- idopsis thaliana) (Lung et al., 2015)及烟草(Nicotia- na tobacum) (Jones et al., 2012)中, 原生质提取及转化效果较好, 并得到广泛应用.而在非模式植物如葡萄(Vitis vinifera) (Fontes et al., 2010; 舒小娟等, 2015)、柳枝黍(Panicum virgatum) (Mazarei et al., 2008)、杨树(Populus tremula × alba) (Guo et al., 2012)、松树(Pinus pinaster) (David et al., 1989)及紫锥菊(Echinacea augustifolia) (Liqing et al., 2005; 朱蠡庆等, 2005)中也有相关报道. ... ... 目前原生质体在基因工程及分子生物学研究中都已得到普遍应用(Sheen, 2001).采用GFP和YFP荧光标记, 通过激光共聚焦显微镜检测, 可以简单方便地确定目标蛋白在细胞中的定位(周波等, 2008).Cao等(2011)采用目的基因融合GFP的方法, 可清晰地观察到单糖转运蛋白OsGMST1位于高尔基体上.Tang等(2012)利用原生质体技术定位水稻(Oryza sativa)硝酸盐转运蛋白OsNRT2.3a, 发现其主要位于细胞膜上.现有的原生质体转化方法主要有PEG-Ca2+介导转化、电转化及显微注射3种(Yoo et al., 2007).我们利用PEG-Ca2+介导转化法将提取得到的原生质体进行GFP和YFP空载体转化, 转化效率高达80% (图2), 且瞬时表达出蛋白(图3).一般认为, 转化效率在50%以上可以用于常规的分子机理研究(Yoo et al., 2007), 因此, 本实验建立的提取及转化系统可用于研究蛋白质的亚细胞定位. ...
紫锥菊愈伤组织原生质体分离方法初探 1 2005
... 植物原生质体(protoplast)是指通过质壁分离能够分开的那部分仅由细胞膜包围的具有活力的细胞物质(周波等, 2008; 于晓玲等, 2009).细胞壁对植物细胞具有支撑和保护作用, 其主要成分是果胶质、纤维素、半纤维素与少量蛋白质(刘敏和顾志敏, 2014).原生质体能摄入DNA、质粒、病毒、细胞器等外源物质(An et al., 2005), 又可用于研究蛋白定位, 还可以验证蛋白之间的互作, 检测其互作强弱, 定位互作蛋白的位点, 从而为推断蛋白质的生物学功能研究提供必要的基础信息, 因此在细胞工程和基因工程中应用广泛(刘敏和顾志敏, 2014).在模式植物拟南芥(Arab- idopsis thaliana) (Lung et al., 2015)及烟草(Nicotia- na tobacum) (Jones et al., 2012)中, 原生质提取及转化效果较好, 并得到广泛应用.而在非模式植物如葡萄(Vitis vinifera) (Fontes et al., 2010; 舒小娟等, 2015)、柳枝黍(Panicum virgatum) (Mazarei et al., 2008)、杨树(Populus tremula × alba) (Guo et al., 2012)、松树(Pinus pinaster) (David et al., 1989)及紫锥菊(Echinacea augustifolia) (Liqing et al., 2005; 朱蠡庆等, 2005)中也有相关报道. ...
1 2005
... 植物原生质体(protoplast)是指通过质壁分离能够分开的那部分仅由细胞膜包围的具有活力的细胞物质(周波等, 2008; 于晓玲等, 2009).细胞壁对植物细胞具有支撑和保护作用, 其主要成分是果胶质、纤维素、半纤维素与少量蛋白质(刘敏和顾志敏, 2014).原生质体能摄入DNA、质粒、病毒、细胞器等外源物质(An et al., 2005), 又可用于研究蛋白定位, 还可以验证蛋白之间的互作, 检测其互作强弱, 定位互作蛋白的位点, 从而为推断蛋白质的生物学功能研究提供必要的基础信息, 因此在细胞工程和基因工程中应用广泛(刘敏和顾志敏, 2014).在模式植物拟南芥(Arab- idopsis thaliana) (Lung et al., 2015)及烟草(Nicotia- na tobacum) (Jones et al., 2012)中, 原生质提取及转化效果较好, 并得到广泛应用.而在非模式植物如葡萄(Vitis vinifera) (Fontes et al., 2010; 舒小娟等, 2015)、柳枝黍(Panicum virgatum) (Mazarei et al., 2008)、杨树(Populus tremula × alba) (Guo et al., 2012)、松树(Pinus pinaster) (David et al., 1989)及紫锥菊(Echinacea augustifolia) (Liqing et al., 2005; 朱蠡庆等, 2005)中也有相关报道. ...
1 2012
... 利用原生质体进行蛋白相互作用研究是常见的策略.为了验证非洲菊原生质体具有该潜力, 选取拟南芥中能够相互作用的2个转录因子IBH1和HBI1 (Bai et al., 2012; Fan et al., 2014; Malinovsky et al., 2014)探究上述原生质体提取体系能否应用于遗传共转化实验.利用IBH1-nYFP和HBI1-cYFP共转化非洲菊原生质体, 通过原生质体荧光定位, 能直接观察到IBH1和HBI1互作所形成的复合体在原生质体中的定位情况.结果显示, 与IBH1-nYFP+cYFP和HBI1-cYFP+nYFP阴性对照相比, 在共转化IBH1-nYFP和HBI1-cYFP质粒的细胞核中能观察到强烈的YFP荧光信号(图4). ...
2 2008
... 在进化上有明显优势的菊科植物是23 000多种开花植物中最大的分支(Broholm et al., 2008).非洲菊(Gerbera hybrida)为菊科大丁草属多年生常绿草本植物, 可作为研究菊科植物器官生长发育调控的模式植物(Teeri et al., 2006; Laitinen et al., 2007; Broholm et al., 2008; Li et al., 2015).与其它模式植物(如拟南芥或烟草)相比, 非洲菊遗传背景较为复杂、基因数量多、遗传转化困难, 其分子机制研究更难, 而原生质体提取及瞬时转化则为克服这一障碍开辟了一条可能的新途径(舒小娟等, 2015).近年来, 随着对非洲菊的深入研究, 许多基因功能的揭示需要回到植株器官或细胞中, 其中包括蛋白亚细胞定位和蛋白间相互作用.目前, 非洲菊原生质体研究进展缓慢, 大多处于分离阶段, 且效率非常低, 也并未用于遗传转化研究(李丹等, 2009).因此, 建立高效的非洲菊原生质体提取及瞬时转化体系十分必要.本研究借鉴前人经验, 以非洲菊组培苗叶片为材料, 利用不同浓度纤维素酶和离析酶配比对非洲菊原生质体提取效率的影响进行了探索, 并在此基础上建立瞬时转化体系. ... ... ; Broholm et al., 2008; Li et al., 2015).与其它模式植物(如拟南芥或烟草)相比, 非洲菊遗传背景较为复杂、基因数量多、遗传转化困难, 其分子机制研究更难, 而原生质体提取及瞬时转化则为克服这一障碍开辟了一条可能的新途径(舒小娟等, 2015).近年来, 随着对非洲菊的深入研究, 许多基因功能的揭示需要回到植株器官或细胞中, 其中包括蛋白亚细胞定位和蛋白间相互作用.目前, 非洲菊原生质体研究进展缓慢, 大多处于分离阶段, 且效率非常低, 也并未用于遗传转化研究(李丹等, 2009).因此, 建立高效的非洲菊原生质体提取及瞬时转化体系十分必要.本研究借鉴前人经验, 以非洲菊组培苗叶片为材料, 利用不同浓度纤维素酶和离析酶配比对非洲菊原生质体提取效率的影响进行了探索, 并在此基础上建立瞬时转化体系. ...
2011
1 1989
... 植物原生质体(protoplast)是指通过质壁分离能够分开的那部分仅由细胞膜包围的具有活力的细胞物质(周波等, 2008; 于晓玲等, 2009).细胞壁对植物细胞具有支撑和保护作用, 其主要成分是果胶质、纤维素、半纤维素与少量蛋白质(刘敏和顾志敏, 2014).原生质体能摄入DNA、质粒、病毒、细胞器等外源物质(An et al., 2005), 又可用于研究蛋白定位, 还可以验证蛋白之间的互作, 检测其互作强弱, 定位互作蛋白的位点, 从而为推断蛋白质的生物学功能研究提供必要的基础信息, 因此在细胞工程和基因工程中应用广泛(刘敏和顾志敏, 2014).在模式植物拟南芥(Arab- idopsis thaliana) (Lung et al., 2015)及烟草(Nicotia- na tobacum) (Jones et al., 2012)中, 原生质提取及转化效果较好, 并得到广泛应用.而在非模式植物如葡萄(Vitis vinifera) (Fontes et al., 2010; 舒小娟等, 2015)、柳枝黍(Panicum virgatum) (Mazarei et al., 2008)、杨树(Populus tremula × alba) (Guo et al., 2012)、松树(Pinus pinaster) (David et al., 1989)及紫锥菊(Echinacea augustifolia) (Liqing et al., 2005; 朱蠡庆等, 2005)中也有相关报道. ...
2003
1 2014
... 利用原生质体进行蛋白相互作用研究是常见的策略.为了验证非洲菊原生质体具有该潜力, 选取拟南芥中能够相互作用的2个转录因子IBH1和HBI1 (Bai et al., 2012; Fan et al., 2014; Malinovsky et al., 2014)探究上述原生质体提取体系能否应用于遗传共转化实验.利用IBH1-nYFP和HBI1-cYFP共转化非洲菊原生质体, 通过原生质体荧光定位, 能直接观察到IBH1和HBI1互作所形成的复合体在原生质体中的定位情况.结果显示, 与IBH1-nYFP+cYFP和HBI1-cYFP+nYFP阴性对照相比, 在共转化IBH1-nYFP和HBI1-cYFP质粒的细胞核中能观察到强烈的YFP荧光信号(图4). ...
1 2010
... 植物原生质体(protoplast)是指通过质壁分离能够分开的那部分仅由细胞膜包围的具有活力的细胞物质(周波等, 2008; 于晓玲等, 2009).细胞壁对植物细胞具有支撑和保护作用, 其主要成分是果胶质、纤维素、半纤维素与少量蛋白质(刘敏和顾志敏, 2014).原生质体能摄入DNA、质粒、病毒、细胞器等外源物质(An et al., 2005), 又可用于研究蛋白定位, 还可以验证蛋白之间的互作, 检测其互作强弱, 定位互作蛋白的位点, 从而为推断蛋白质的生物学功能研究提供必要的基础信息, 因此在细胞工程和基因工程中应用广泛(刘敏和顾志敏, 2014).在模式植物拟南芥(Arab- idopsis thaliana) (Lung et al., 2015)及烟草(Nicotia- na tobacum) (Jones et al., 2012)中, 原生质提取及转化效果较好, 并得到广泛应用.而在非模式植物如葡萄(Vitis vinifera) (Fontes et al., 2010; 舒小娟等, 2015)、柳枝黍(Panicum virgatum) (Mazarei et al., 2008)、杨树(Populus tremula × alba) (Guo et al., 2012)、松树(Pinus pinaster) (David et al., 1989)及紫锥菊(Echinacea augustifolia) (Liqing et al., 2005; 朱蠡庆等, 2005)中也有相关报道. ...
1 2012
... 植物原生质体(protoplast)是指通过质壁分离能够分开的那部分仅由细胞膜包围的具有活力的细胞物质(周波等, 2008; 于晓玲等, 2009).细胞壁对植物细胞具有支撑和保护作用, 其主要成分是果胶质、纤维素、半纤维素与少量蛋白质(刘敏和顾志敏, 2014).原生质体能摄入DNA、质粒、病毒、细胞器等外源物质(An et al., 2005), 又可用于研究蛋白定位, 还可以验证蛋白之间的互作, 检测其互作强弱, 定位互作蛋白的位点, 从而为推断蛋白质的生物学功能研究提供必要的基础信息, 因此在细胞工程和基因工程中应用广泛(刘敏和顾志敏, 2014).在模式植物拟南芥(Arab- idopsis thaliana) (Lung et al., 2015)及烟草(Nicotia- na tobacum) (Jones et al., 2012)中, 原生质提取及转化效果较好, 并得到广泛应用.而在非模式植物如葡萄(Vitis vinifera) (Fontes et al., 2010; 舒小娟等, 2015)、柳枝黍(Panicum virgatum) (Mazarei et al., 2008)、杨树(Populus tremula × alba) (Guo et al., 2012)、松树(Pinus pinaster) (David et al., 1989)及紫锥菊(Echinacea augustifolia) (Liqing et al., 2005; 朱蠡庆等, 2005)中也有相关报道. ...
1 2012
... 植物原生质体(protoplast)是指通过质壁分离能够分开的那部分仅由细胞膜包围的具有活力的细胞物质(周波等, 2008; 于晓玲等, 2009).细胞壁对植物细胞具有支撑和保护作用, 其主要成分是果胶质、纤维素、半纤维素与少量蛋白质(刘敏和顾志敏, 2014).原生质体能摄入DNA、质粒、病毒、细胞器等外源物质(An et al., 2005), 又可用于研究蛋白定位, 还可以验证蛋白之间的互作, 检测其互作强弱, 定位互作蛋白的位点, 从而为推断蛋白质的生物学功能研究提供必要的基础信息, 因此在细胞工程和基因工程中应用广泛(刘敏和顾志敏, 2014).在模式植物拟南芥(Arab- idopsis thaliana) (Lung et al., 2015)及烟草(Nicotia- na tobacum) (Jones et al., 2012)中, 原生质提取及转化效果较好, 并得到广泛应用.而在非模式植物如葡萄(Vitis vinifera) (Fontes et al., 2010; 舒小娟等, 2015)、柳枝黍(Panicum virgatum) (Mazarei et al., 2008)、杨树(Populus tremula × alba) (Guo et al., 2012)、松树(Pinus pinaster) (David et al., 1989)及紫锥菊(Echinacea augustifolia) (Liqing et al., 2005; 朱蠡庆等, 2005)中也有相关报道. ...
1 2007
... 在进化上有明显优势的菊科植物是23 000多种开花植物中最大的分支(Broholm et al., 2008).非洲菊(Gerbera hybrida)为菊科大丁草属多年生常绿草本植物, 可作为研究菊科植物器官生长发育调控的模式植物(Teeri et al., 2006; Laitinen et al., 2007; Broholm et al., 2008; Li et al., 2015).与其它模式植物(如拟南芥或烟草)相比, 非洲菊遗传背景较为复杂、基因数量多、遗传转化困难, 其分子机制研究更难, 而原生质体提取及瞬时转化则为克服这一障碍开辟了一条可能的新途径(舒小娟等, 2015).近年来, 随着对非洲菊的深入研究, 许多基因功能的揭示需要回到植株器官或细胞中, 其中包括蛋白亚细胞定位和蛋白间相互作用.目前, 非洲菊原生质体研究进展缓慢, 大多处于分离阶段, 且效率非常低, 也并未用于遗传转化研究(李丹等, 2009).因此, 建立高效的非洲菊原生质体提取及瞬时转化体系十分必要.本研究借鉴前人经验, 以非洲菊组培苗叶片为材料, 利用不同浓度纤维素酶和离析酶配比对非洲菊原生质体提取效率的影响进行了探索, 并在此基础上建立瞬时转化体系. ...
1 2015
... 在进化上有明显优势的菊科植物是23 000多种开花植物中最大的分支(Broholm et al., 2008).非洲菊(Gerbera hybrida)为菊科大丁草属多年生常绿草本植物, 可作为研究菊科植物器官生长发育调控的模式植物(Teeri et al., 2006; Laitinen et al., 2007; Broholm et al., 2008; Li et al., 2015).与其它模式植物(如拟南芥或烟草)相比, 非洲菊遗传背景较为复杂、基因数量多、遗传转化困难, 其分子机制研究更难, 而原生质体提取及瞬时转化则为克服这一障碍开辟了一条可能的新途径(舒小娟等, 2015).近年来, 随着对非洲菊的深入研究, 许多基因功能的揭示需要回到植株器官或细胞中, 其中包括蛋白亚细胞定位和蛋白间相互作用.目前, 非洲菊原生质体研究进展缓慢, 大多处于分离阶段, 且效率非常低, 也并未用于遗传转化研究(李丹等, 2009).因此, 建立高效的非洲菊原生质体提取及瞬时转化体系十分必要.本研究借鉴前人经验, 以非洲菊组培苗叶片为材料, 利用不同浓度纤维素酶和离析酶配比对非洲菊原生质体提取效率的影响进行了探索, 并在此基础上建立瞬时转化体系. ...
1 2005
... 植物原生质体(protoplast)是指通过质壁分离能够分开的那部分仅由细胞膜包围的具有活力的细胞物质(周波等, 2008; 于晓玲等, 2009).细胞壁对植物细胞具有支撑和保护作用, 其主要成分是果胶质、纤维素、半纤维素与少量蛋白质(刘敏和顾志敏, 2014).原生质体能摄入DNA、质粒、病毒、细胞器等外源物质(An et al., 2005), 又可用于研究蛋白定位, 还可以验证蛋白之间的互作, 检测其互作强弱, 定位互作蛋白的位点, 从而为推断蛋白质的生物学功能研究提供必要的基础信息, 因此在细胞工程和基因工程中应用广泛(刘敏和顾志敏, 2014).在模式植物拟南芥(Arab- idopsis thaliana) (Lung et al., 2015)及烟草(Nicotia- na tobacum) (Jones et al., 2012)中, 原生质提取及转化效果较好, 并得到广泛应用.而在非模式植物如葡萄(Vitis vinifera) (Fontes et al., 2010; 舒小娟等, 2015)、柳枝黍(Panicum virgatum) (Mazarei et al., 2008)、杨树(Populus tremula × alba) (Guo et al., 2012)、松树(Pinus pinaster) (David et al., 1989)及紫锥菊(Echinacea augustifolia) (Liqing et al., 2005; 朱蠡庆等, 2005)中也有相关报道. ...
1 2015
... 植物原生质体(protoplast)是指通过质壁分离能够分开的那部分仅由细胞膜包围的具有活力的细胞物质(周波等, 2008; 于晓玲等, 2009).细胞壁对植物细胞具有支撑和保护作用, 其主要成分是果胶质、纤维素、半纤维素与少量蛋白质(刘敏和顾志敏, 2014).原生质体能摄入DNA、质粒、病毒、细胞器等外源物质(An et al., 2005), 又可用于研究蛋白定位, 还可以验证蛋白之间的互作, 检测其互作强弱, 定位互作蛋白的位点, 从而为推断蛋白质的生物学功能研究提供必要的基础信息, 因此在细胞工程和基因工程中应用广泛(刘敏和顾志敏, 2014).在模式植物拟南芥(Arab- idopsis thaliana) (Lung et al., 2015)及烟草(Nicotia- na tobacum) (Jones et al., 2012)中, 原生质提取及转化效果较好, 并得到广泛应用.而在非模式植物如葡萄(Vitis vinifera) (Fontes et al., 2010; 舒小娟等, 2015)、柳枝黍(Panicum virgatum) (Mazarei et al., 2008)、杨树(Populus tremula × alba) (Guo et al., 2012)、松树(Pinus pinaster) (David et al., 1989)及紫锥菊(Echinacea augustifolia) (Liqing et al., 2005; 朱蠡庆等, 2005)中也有相关报道. ...
1 2014
... 利用原生质体进行蛋白相互作用研究是常见的策略.为了验证非洲菊原生质体具有该潜力, 选取拟南芥中能够相互作用的2个转录因子IBH1和HBI1 (Bai et al., 2012; Fan et al., 2014; Malinovsky et al., 2014)探究上述原生质体提取体系能否应用于遗传共转化实验.利用IBH1-nYFP和HBI1-cYFP共转化非洲菊原生质体, 通过原生质体荧光定位, 能直接观察到IBH1和HBI1互作所形成的复合体在原生质体中的定位情况.结果显示, 与IBH1-nYFP+cYFP和HBI1-cYFP+nYFP阴性对照相比, 在共转化IBH1-nYFP和HBI1-cYFP质粒的细胞核中能观察到强烈的YFP荧光信号(图4). ...
1 2008
... 植物原生质体(protoplast)是指通过质壁分离能够分开的那部分仅由细胞膜包围的具有活力的细胞物质(周波等, 2008; 于晓玲等, 2009).细胞壁对植物细胞具有支撑和保护作用, 其主要成分是果胶质、纤维素、半纤维素与少量蛋白质(刘敏和顾志敏, 2014).原生质体能摄入DNA、质粒、病毒、细胞器等外源物质(An et al., 2005), 又可用于研究蛋白定位, 还可以验证蛋白之间的互作, 检测其互作强弱, 定位互作蛋白的位点, 从而为推断蛋白质的生物学功能研究提供必要的基础信息, 因此在细胞工程和基因工程中应用广泛(刘敏和顾志敏, 2014).在模式植物拟南芥(Arab- idopsis thaliana) (Lung et al., 2015)及烟草(Nicotia- na tobacum) (Jones et al., 2012)中, 原生质提取及转化效果较好, 并得到广泛应用.而在非模式植物如葡萄(Vitis vinifera) (Fontes et al., 2010; 舒小娟等, 2015)、柳枝黍(Panicum virgatum) (Mazarei et al., 2008)、杨树(Populus tremula × alba) (Guo et al., 2012)、松树(Pinus pinaster) (David et al., 1989)及紫锥菊(Echinacea augustifolia) (Liqing et al., 2005; 朱蠡庆等, 2005)中也有相关报道. ...
 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}