Abstract Aims Litterfall is an important component of ecosystem net primary productivity, and a key link between above- and below-ground carbon processes. However, it remains unclear how biodiversity, stand factors and functional traits work together in affecting litterfall production and its temporal stability during forest succession. Methods We measured litterfall production for three years in plots from four successional stages of broad-leaf and Korean pine (Pinus koraiensis) mixed forests, at the northern limit of Korean pine forest in the Shengshan Reserve of Heilongjiang. Functional traits (leaf carbon and nitrogen contents, specific leaf area) were measured to quantify functional diversity and community weighted mean (CWM) of traits. Tree diameter and height, total basal area (TBA) and gap fraction were measured to quantify stand structure. We used hierarchy partitioning analysis and variance partitioning to evaluate the relative effects of stand factors, community level traits, and (species, functional and phylogenetic) diversity on annual litterfall production, and its temporal stability (= 1/coefficient of variation for annual litterfall production). Important findings Litterfall production was significantly lower in the early successional stage, but did no vary from the middle to late successional stages. The litterfall stability increased continuously with forest succession. The variable importance of multivariate models also suggest that, the litterfall production was mainly affected by stand factors (e.g. height, TBA, gap fraction) and functional traits (leaf carbon content), with species richness also playing a role. For stability of litterfall production, however, functional diversity was the strongest predictor, followed by stand factors (such as maximum tree diameter). The independent effect of biodiversity on litterfall production was only 0.41%, but was as much higher (33.12%) for temporal stability of litterfall, suggesting that biodiversity have an important influence on litterfall stability that is independent of stand factors and traits. There was also a strong joint effect (up to 53.8%) among the stand factors, biodiversity and functional traits, indicating that these factors collectively affect litterfall production and its stability. Our results suggest that forest succession not only increases forest productivity but also improves ecosystem stability. Therefore, protecting primary forests and promoting forest restoration are effective ways to improve ecosystem functions. Keywords:litterfall production;stability;biodiversity;stand factors;succession;functional traits;broad-leaf and Korean pine mixed forest
PDF (1203KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 孙浩哲, 王襄平, 张树斌, 吴鹏, 杨蕾. 阔叶红松林不同演替阶段凋落物产量及其稳定性的影响因素. 植物生态学报, 2021, 45(6): 594-605. DOI: 10.17521/cjpe.2020.0372 SUN Hao-Zhe, WANG Xiang-Ping, ZHANG Shu-Bin, WU Peng, YANG Lei. Abiotic and biotic modulators of litterfall production and its temporal stability during the succession of broad-leaf and Korean pine mixed forest. Chinese Journal of Plant Ecology, 2021, 45(6): 594-605. DOI: 10.17521/cjpe.2020.0372
Table 1 表1 表1黑龙江胜山保护区阔叶红松林各演替阶段样地的林分特征 Table 1Stand characteristics for plots of different succession stages of broadleaf Korean pine forest in the Shengshan Nature Reserve, Heilongjiang Province of China
演替阶段 Succession stage
平均树高 Mean tree height (m)
平均胸径 Mean DBH (cm)
最大树高 Hmax (m)
最大胸径 DBHmax (cm)
林分密度 Stem density (tree·hm-2)
林冠空隙度 Gap fraction (%)
胸高断面积 TBA (m2·hm-2)
早期 Early stage
13.3
13.2
22.6
29.8
853
32.1
14.8
中期 Middle stage
8.9
11.1
24.3
43.8
1 577
13.1
30.9
中晚期 Mid-late stage
12.0
11.9
28.9
51.8
1 723
15.9
21.4
晚期 Late stage
13.1
16.6
28.6
67.7
1 180
6.6
42.4
DBHmax, maximum diameter at breast height; Hmax, maximum tree height; TBA, total basal area.
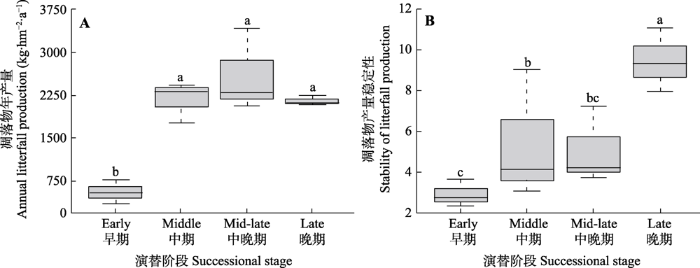
Fig. 1Difference in annual litterfall production and its temporal stability among successional stages of Korean pine forest. Different lowercase letters indicate significant differences at p < 0.05 level.
Table 2 表2 表2阔叶红松林不同演替阶段凋落物产量及其稳定性与各解释变量的决定系数(R2) Table 2Coefficients of determination (R2) for different factors in explaining annual litterfall production and its temporal stability across succesional stages of Korean pine forest
变量 Variable
凋落物产量 Annual litterfall production
凋落物产量稳定性 Stability of litterfall production
林分因子 Stand factor
最大树高 Hmax
0.57**
0.39*
最大胸径 DBHmax
0.45*
0.55**
胸高断面积 TBA
0.43*
0.57**
林冠空隙度 Gap fraction
-0.68***
-0.53**
功能性状 Functional trait
叶片碳含量 CWMLC
-0.26
0.00
叶片氮含量 CWMLN
0.05
0.05
比叶面积 CWMSLA
-0.11
-0.54**
生物多样性 Biodiversity
物种丰富度 Richness
0.65**
0.42*
物种均匀度 Evenness
0.59**
0.28
功能多样性 Rao’s Q
0.45**
0.16
系统发育多样性 PD
0.32
0.24
“-”表示负相关关系; ***, p < 0.001; **, p < 0.01; *, p < 0.05。 “-” denotes negative relationship; ***, p < 0.001; **, p < 0.01; *, p < 0.05. CWMLC and CWMLN, community weighted mean of leaf carbon and nitrogen content, respectively; CWMSLA, community weighted mean of specific leaf area; DBHmax, maximum diameter at breast height; Hmax, maximum tree height; PD, phylogenetic diversity; Rao’s Q, Rao’s quadratic entropy; TBA, total basal area.
Fig. 2Relative importance of variables retained in the models explaining litterfall production (A) and temporal stability of litterfall (B), as obtained by the hierarchical partitioning analyses. CWMLC and CWMLN, community weighted mean of leaf carbon and nitrogen content, respectively; CWMSLA, community weighted mean of specific leaf area; DBHmax, maximum diameter at breast height; Hmax, maximum tree height; PD, phylogenetic diversity; Rao’s Q, Rao’s quadratic entropy; TBA, total basal area.
为进一步反映林分因子、功能性状和多样性如何共同作用, 本研究还对BIC筛选后的多元模型进行了变异分离分析(图3)。结果表明林分因子是对凋落物产量影响最大的因素, 其总解释力为82.56% (a + d + e + g), 功能性状的总解释力为68.55% (b + d + f + g), 多样性的总解释力为64.84% (c + e + f + g)。林分因子、功能性状均有较强的独立解释力(分别为21.02%和15.36%), 而多样性对凋落物产量的独立解释力极弱(0.41%), 说明多样性主要是通过与林分因子和功能性状的协同作用来影响凋落物产量(3种因素之间的协同作用(g) = 53.8%)。林分因子、功能性状和多样性对凋落物产量稳定性的总解释力分别为59.68%、63.3%和47.67%; 不同于凋落物产量的是, 生物多样性对稳定性的独立解释力较强(33.12%), 明显大于林分因子(21.14%)和功能性状(16.79%)的独立解释力。这3种因素对稳定性同样存在很强的协同作用, 说明多样性除了自身的独立影响, 还通过和林分因子、性状的相互影响对生态系统稳定性产生重要作用。
Fig. 3Variance partitioning for the effects of stand factors, functional traits and biodiversity factors retained in the models on litterfall production (A) and temporal stability of litterfall (B). a, b, c, the independent effects by each of the three factors; d, e, f, the joint effects between two factors; g, the joint effect among three factors.
3 讨论
3.1 不同演替阶段凋落物产量及其稳定性的变化
有研究认为凋落物产量沿演替序列增加(Zhou et al., 2007), 也有****发现凋落物产量在不同演替阶段无显著差异(Huang et al., 2017)。一般而言, 在林分发育直至林冠郁闭之前每年的凋落物产量会迅速增加, 然后在相当长的一段时间内保持相对稳定(Albrektson, 1988), 本研究的结果与之一致。胜山自然保护区演替早期的林分密度、树木个体都较小, 林冠未达到郁闭导致其生产力有限, 所以凋落物产量显著低于其他演替阶段, 从演替中期至演替晚期林分高度郁闭(表1), 群落可捕获的光能大致相同, 使得3个演替阶段凋落物产量无显著差异。
AertsR, Chapin IIIFS (1999). The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns Advances in Ecological Research, 30, 1-67. [本文引用: 1]
AlbrektsonA (1988). Needle litterfall in stands of Pinus sylvestris L. in Sweden, in relation to site quality, stand age and latitude Scandinavian Journal of Forest Research, 3, 333-342. DOI:10.1080/02827588809382521URL [本文引用: 1]
AliA, LinSL, HeJK, KongFM, YuJH, JiangHS (2019). Big-sized trees overrule remaining treesʼ attributes and species richness as determinants of aboveground biomass in tropical forests Global Change Biology, 25, 2810-2824. DOI:10.1111/gcb.2019.25.issue-8URL [本文引用: 4]
AndersonKJ (2007). Temporal patterns in rates of community change during succession The American Naturalist, 169, 780-793. DOI:10.1086/516653URL [本文引用: 1]
AtkinsJW, FaheyRT, HardimanBS, GoughCM (2018). Forest canopy structural complexity and light absorption relationships at the subcontinental scale Journal of Geophysical Research, 123, 1387-1405. [本文引用: 1]
CadotteMW, Cavender-BaresJ, TilmanD, OakleyTH (2009). Using phylogenetic, functional and trait diversity to understand patterns of plant community productivity PLOS ONE, 4, e5695. DOI: 10.1371/journal.pone.0005695. URL [本文引用: 2]
CaldeiraMC, HectorA, LoreauM, PereiraJS (2005). Species richness, temporal variability and resistance of biomass production in a Mediterranean grassland Oikos, 110, 115-123. DOI:10.1111/oik.2005.110.issue-1URL [本文引用: 1]
CardinaleBJ, DuffyJE, GonzalezA, HooperDU, PerringsC, VenailP, NarwaniA, MaceGM, TilmanD, WardleDA, KinzigAP, DailyGC, LoreauM, GraceJB, LarigauderieA, SrivastavaDS, NaeemS (2012). Biodiversity loss and its impact on humanity Nature, 486, 59-67. DOI:10.1038/nature11148URL [本文引用: 1]
CavanaughKC, GosnellJS, DavisSL, AhumadaJ, BoundjaP, ClarkDB, MugerwaB, JansenPA, O’BrienTG, RoveroF, SheilD, VasquezR, AndelmanS (2014). Carbon storage in tropical forests correlates with taxonomic diversity and functional dominance on a global scale Global Ecology and Biogeography, 23, 563-573. DOI:10.1111/geb.2014.23.issue-5URL [本文引用: 2]
ClarkDA, BrownS, KicklighterDW, ChambersJQ, ThomlinsonJR, NiJ (2001). Measuring net primary production in forests: concepts and field methods Ecological Applications, 11, 356-370. DOI:10.1890/1051-0761(2001)011[0356:MNPPIF]2.0.CO;2URL [本文引用: 1]
FineganB, Peña-ClarosM, de OliveiraA, AscarrunzN, Bret-HarteMS, Carreño-RocabadoG, CasanovesF, DíazS, Eguiguren VelepuchaP, FernandezF, LiconaJC, LorenzoL, Salgado NegretB, VazM, PoorterL, CanhamC (2015). Does functional trait diversity predict above- ground biomass and productivity of tropical forests? Testing three alternative hypotheses Journal of Ecology, 103, 191-201. DOI:10.1111/1365-2745.12346URL [本文引用: 1]
ForresterDI (2014). The spatial and temporal dynamics of species interactions in mixed-species forests: from pattern to process Forest Ecology and Management, 312, 282-292. DOI:10.1016/j.foreco.2013.10.003URL [本文引用: 1]
FotisAT, MurphySJ, RicartRD, KrishnadasM, WhitacreJ, WenzelJW, QueenboroughSA, ComitaLS (2018). Above-ground biomass is driven by mass-ratio effects and stand structural attributes in a temperate deciduous forest Journal of Ecology, 106, 561-570. DOI:10.1111/1365-2745.12847URL [本文引用: 3]
GuoY, ChenHYH, MallikAU, WangB, LiD, XiangW, LiX (2019). Predominance of abiotic drivers in the relationship between species diversity and litterfall production in a tropical karst seasonal rainforest Forest Ecology and Management, 449, 117452. DOI: 10.1016/j.foreco.2019.117452. URL [本文引用: 3]
HarpoleWS, TilmanD (2007). Grassland species loss resulting from reduced niche dimension Nature, 446, 791-793. DOI:10.1038/nature05684URL [本文引用: 1]
HeJS, FangJ, WangZ, GuoD, FlynnDFB, GengZ (2006). Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China Oecologia, 149, 115-122. DOI:10.1007/s00442-006-0425-0URL [本文引用: 1]
HuangY, MaY, ZhaoK, NiklausPA, SchmidB, HeJS (2017). Positive effects of tree species diversity on litterfall quantity and quality along a secondary successional chronosequence in a subtropical forest Journal of Plant Ecology, 10, 28-35. DOI:10.1093/jpe/rtw115URL [本文引用: 3]
IsbellFI, PolleyHW, WilseyBJ (2009). Biodiversity, productivity and the temporal stability of productivity: patterns and processes Ecology Letters, 12, 443-451. DOI:10.1111/ele.2009.12.issue-5URL [本文引用: 1]
JiaBR, ZhouGS, LiuYZ, JiangYL(2016). Spatial pattern and environmental controls of annual litterfall production in natural forest ecosystems in China Scientia Sinica Vitae, 46, 1304-1311. DOI:10.1360/N052015-00319URL [本文引用: 1]
JohnsonJB, OmlandKS (2004). Model selection in ecology and evolution Trends in Ecology & Evolution, 19, 101-108. DOI:10.1016/j.tree.2003.10.013URL [本文引用: 1]
KingDA, DaviesSJ, NoorNSM (2006). Growth and mortality are related to adult tree size in a Malaysian mixed dipterocarp forest Forest Ecology and Management, 223, 152-158. DOI:10.1016/j.foreco.2005.10.066URL [本文引用: 1]
KleinD, HumpenӧderF, BauerN, DietrichJP, PoppA, BodirskyBL, BonschM, Lotze-CampenH (2014). The global economic long-term potential of modern biomass in a climate-constrained world Environmental Research Letters, 9, 074017. DOI: 10.1088/1748-9326/9/7/074017. URL [本文引用: 1]
LalibertéE, LegendreP (2010). A distance-based framework for measuring functional diversity from multiple traits Ecology, 91, 299-305. PMID:20380219 [本文引用: 1] A new framework for measuring functional diversity (FD) from multiple traits has recently been proposed. This framework was mostly limited to quantitative traits without missing values and to situations in which there are more species than traits, although the authors had suggested a way to extend their framework to other trait types. The main purpose of this note is to further develop this suggestion. We describe a highly flexible distance-based framework to measure different facets of FD in multidimensional trait space from any distance or dissimilarity measure, any number of traits, and from different trait types (i.e., quantitative, semi-quantitative, and qualitative). This new approach allows for missing trait values and the weighting of individual traits. We also present a new multidimensional FD index, called functional dispersion (FDis), which is closely related to Rao's quadratic entropy. FDis is the multivariate analogue of the weighted mean absolute deviation (MAD), in which the weights are species relative abundances. For unweighted presence-absence data, FDis can be used for a formal statistical test of differences in FD. We provide the "FD" R language package to easily implement our distance-based FD framework.
LaskyJR, UriarteM, BoukiliVK, EricksonDL, John KressW, ChazdonRL (2014). The relationship between tree biodiversity and biomass dynamics changes with tropical forest succession Ecology Letters, 17, 1158-1167. DOI:10.1111/ele.2014.17.issue-9URL [本文引用: 2]
LetcherSG, ChazdonRL (2009). Rapid recovery of biomass, species richness, and species composition in a forest chronosequence in northeastern Costa rica. Biotropica, 41, 608-617. [本文引用: 1]
LiangJ, CrowtherTW, PicardN, WiserS, ZhouM, AlbertiG, SchulzeED, McGuireAD, BozzatoF, PretzschH, de MiguelS, PaquetteA, HéraultB, Scherer-LorenzenM, BarrettCB, et al. (2016). Positive biodiversity-productivity relationship predominant in global forests Science, 354, aaf8957. DOI: 10.1126/science.aaf8957. [本文引用: 1]
LiangPH, WangXP, WUYL, XuK, WuP, GuoX(2016). Growth responses of broad-leaf and Korean pine mixed forests at different successional stages to climate change in the Shengshan Nature Reserve of Heilongjiang Province, China Chinese Journal of Plant Ecology, 40, 425-435. DOI:10.17521/cjpe.2015.0357URL [本文引用: 3]
LohbeckM, PoorterL, Martínez-RamosM, BongersF (2015). Biomass is the main driver of changes in ecosystem process rates during tropical forest succession Ecology, 96, 1242-1252. DOI:10.1890/14-0472.1URL [本文引用: 2]
LoreauM, de MazancourtC (2013). Biodiversity and ecosystem stability: a synthesis of underlying mechanisms Ecology Letters, 16, 106-115. DOI:10.1111/ele.2013.16.issue-s1URL [本文引用: 1]
MartinTA, BrownKJ, KučeraJ, MeinzerFC, SprugelDG, HinckleyTM (2001). Control of transpiration in a 220- year-old Abies amabilis forest. Forest Ecology and Management, 152, 211-224. [本文引用: 1]
MazzochiniGG, FonsecaCR, CostaGC, SantosRM, Oliveira- FilhoAT, GanadeG (2019). Plant phylogenetic diversity stabilizes large scale ecosystem productivity Global Ecology and Biogeography, 28, 1430-1439. DOI:10.1111/geb.12963 [本文引用: 3] Aim Global carbon cycle models do not incorporate the stabilizing effect of biodiversity on productivity despite this phenomenon has been widely described in several local scale manipulative experiments. The reason is a lack of evidence supporting the importance of biodiversity on spatial scales at which climate models are built. Here, we test the hypothesis that diversity enhances productivity stability at a large scale. Location South American dryland known as Caatinga (similar to 830,000 km(2)). Time period 2001-2010. Major taxa studied Woody plants. Methods We used the enhanced vegetation index of Caatinga vegetation remnants, from 2001 to 2010, to calculate vegetation productivity stability across years. We used occurrence records of 606 woody species from floristic surveys to derive species richness and phylogenetic diversity at similar to 5 km and similar to 55 km (0.5 degrees) resolution. Climate data were obtained from global databases. Results Plant phylogenetic diversity has a strong positive correlation with productivity stability even after controlling for several climatic variables, such as rainfall, temperature and cloudiness, at both resolutions. Species richness was not significant when climatic variables were included. Main conclusions This result expands by several orders of magnitude the spatial scale of the evidence that biodiversity strengths the resilience of key ecosystem functions. We highlight that, by incorporating plant phylogenetic diversity, regional and global climate models can generate more accurate predictions about future ecosystem functioning and services that are critical to humankind.
MoriAS, FurukawaT, SasakiT (2013). Response diversity determines the resilience of ecosystems to environmental change Biological Reviews, 88, 349-364. DOI:10.1111/brv.2013.88.issue-2URL [本文引用: 4]
MorinX, FahseL, de MazancourtC, Scherer-LorenzenM, BugmannH (2014). Temporal stability in forest productivity increases with tree diversity due to asynchrony in species dynamics Ecology Letters, 17, 1526-1535. DOI:10.1111/ele.2014.17.issue-12URL [本文引用: 2]
NeedhamJ, MerowC, ButtN, MalhiY, MarthewsTR, MorecroftM, McMahonSM (2016). Forest community response to invasive pathogens: the case of ash dieback in a British woodland Journal of Ecology, 104, 315-330. DOI:10.1111/jec.2016.104.issue-2URL [本文引用: 1]
NiinemetsÜ, HauffK, BertinN, TenhunenJD, SteinbrecherR, SeufertG (2002). Monoterpene emissions in relation to foliar photosynthetic and structural variables in Mediterranean evergreen Quercus species New Phytologist, 153, 243-256. DOI:10.1046/j.0028-646X.2001.00323.xURL [本文引用: 2]
OuyangS, XiangW, WangXP, XiaoW, ChenL, LiS, SunH, DengX, ForresterDI, ZengL, LeiP, LeiX, GouX, PengC (2019). Effects of stand age, richness and density on productivity in subtropical forests in China Journal of Ecology, 107, 2266-2277. DOI:10.1111/1365-2745.13194 [本文引用: 3] 1. Forest productivity may be determined not only by biodiversity but also by environmental factors and stand structure attributes. However, the relative importance of these factors in determining productivity is still controversial for subtropical forests. 2. Based on a large dataset from 600 permanent forest inventory plots across subtropical China, we examined the relationship between biodiversity and forest productivity and tested whether stand structural attributes (stand density in terms of trees per ha, age and tree size) and environmental factors (climate and site conditions) had larger effects on productivity. Furthermore, we quantified the relative importance of environmental factors, stand structure and diversity in determining forest productivity. 3. Diversity, together with stand structure and site conditions, regulated the variability in forest productivity. The relationship between diversity and forest productivity did not vary along environmental gradients. Stand density and age were more important modulators of forest productivity than diversity. 4. Synthesis. Diversity had significant and positive effects on productivity in species-rich subtropical forests, but the effects of stand density and age were also important. Our work highlights that while biodiversity conservation is often important, the regulation of stand structure can be even more important to maintain high productivity in subtropical forests.
PeppeDJ, RoyerDL, CariglinoB, OliverSY, NewmanS, LeightE, EnikolopovG, Fernandez-BurgosM, HerreraF, AdamsJM, CorreaE, CurranoED, EricksonJM, HinojosaLF, HogansonJW, et al. (2011). Sensitivity of leaf size and shape to climate: global patterns and paleoclimatic applications New Phytologist, 190, 724-739. DOI:10.1111/nph.2011.190.issue-3URL [本文引用: 1]
Pérez-HarguindeguyN, DíazS, GarnierE, LavorelS, PoorterH, JaureguiberryP, Bret-HarteMS, CornwellWK, CraineJM, GurvichDE, UrcelayC, VeneklaasEJ, ReichPB, PoorterL, WrightIJ, et al. (2013). New handbook for standardised measurement of plant functional traits worldwide Australian Journal of Botany, 61, 167-234. DOI:10.1071/BT12225URL [本文引用: 1]
PiaoS, FangJ, ZhouL, CiaisP, ZhuB (2006). Variations in satellite-derived phenology in China’s temperate vegetation Global Change Biology, 12, 672-685. DOI:10.1111/gcb.2006.12.issue-4URL [本文引用: 1]
PoorterH, De JongR (1999). A comparison of specific leaf area, chemical composition and leaf construction costs of field plants from 15 habitats differing in productivity New Phytologist, 143, 163-176. DOI:10.1046/j.1469-8137.1999.00428.xURL [本文引用: 1]
ScoffoniC, RawlsM, McKownA, CochardH, SackL (2011). Decline of leaf hydraulic conductance with dehydration: relationship to leaf size and venation architecture Plant Physiology, 156, 832-843. DOI:10.1104/pp.111.173856URL [本文引用: 1]
SouzaMC, RossattoDR, CookGD, FujinumaR, MenziesNW, MorellatoLPC, HabermannG (2016). Mineral nutrition and specific leaf area of plants under contrasting long-term fire frequencies: a case study in a mesic savanna in Australia Trees, 30, 329-335. DOI:10.1007/s00468-015-1285-1URL [本文引用: 2]
SterckF, MarkesteijnL, SchievingF, PoorterL (2011). Functional traits determine trade-offs and niches in a tropical forest community Proceedings of the National Academy of Sciences of the United States of America, 108, 20627-20632. DOI:10.1073/pnas.1106950108PMID:22106283 [本文引用: 1] How numerous tree species can coexist in diverse forest communities is a key question in community ecology. Whereas neutral theory assumes that species are adapted to common field conditions and coexist by chance, niche theory predicts that species are functionally different and coexist because they are specialized for different niches. We integrated biophysical principles into a mathematical plant model to determine whether and how functional plant traits and trade-offs may cause functional divergence and niche separation of tree species. We used this model to compare the carbon budget of saplings across 13 co-occurring dry-forest tree species along gradients of light and water availability. We found that species ranged in strategy, from acquisitive species with high carbon budgets at highest resource levels to more conservative species with high tolerances for both shade and drought. The crown leaf area index and nitrogen mass per leaf area drove the functional divergence along the simulated light gradient, which was consistent with observed species distributions along light gradients in the forest. Stomatal coordination to avoid low water potentials or hydraulic failure caused functional divergence along the simulated water gradient, but was not correlated to observed species distributions along the water gradient in the forest. The trait-based biophysical model thus explains how functional traits cause functional divergence across species and whether such divergence contributes to niche separation along resource gradients.
SunH, WangXP, WuP, HanW, XuK, LiangPH, LiuC, YinWL, XiaXL (2017). What causes greater deviations from predictions of metabolic scaling theory in earlier successional forests? Forest Ecology and Management, 405, 101-111. DOI:10.1016/j.foreco.2017.09.007URL [本文引用: 1]
TilmanD (2000). Causes, consequences and ethics of biodiversity Nature, 405, 208-211. DOI:10.1038/35012217URL [本文引用: 1]
TilmanD, KnopsJ, WedinD, ReichP, RitchieM, SiemannE (1997). The influence of functional diversity and composition on ecosystem processes Science, 277, 1300-1302. DOI:10.1126/science.277.5330.1300URL [本文引用: 2]
VogtKA, GrierCC, VogtDJ (1986). Production, turnover, and nutrient dynamics of above- and belowground detritus of world forests Advances in Ecological Research, 15, 303-377. [本文引用: 1]
WanSQ, NorbyRJ, LedfordJ, WeltzinJF (2007). Responses of soil respiration to elevated CO2, air warming, and changing soil water availability in a model old-field grassland Global Change Biology, 13, 2411-2424. DOI:10.1111/gcb.2007.13.issue-11URL [本文引用: 1]
WangXP, FangJY, ZhuB (2008). Forest biomass and root-shoot allocation in northeast China Forest Ecology and Management, 255, 4007-4020. DOI:10.1016/j.foreco.2008.03.055URL [本文引用: 2]
WenXS, ChenBH, ZhangSB, XuK, YeXY, NiWJ, WangXP(2019). Relationships of radial growth with climate change in larch plantations of different stand ages and species Chinese Journal of Plant Ecology, 43, 27-36. DOI:10.17521/cjpe.2018.0155URL [本文引用: 1]
WuX, WangXP, TangZY, ShenZH, ZhengCY, XiaXL, FangJY (2015). The relationship between species richness and biomass changes from boreal to subtropical forests in China Ecography, 38, 602-613. DOI:10.1111/ecog.2015.v38.i6URL [本文引用: 2]
XuK, WangXP, LiangPH, WuYL, AnHL, SunH, WuP, WuX, LiQY, GuoX, WenXS, HanW, LiuC, FanDY (2019). A new tree-ring sampling method to estimate forest productivity and its temporal variation accurately in natural forests Forest Ecology and Management, 433, 217-227. DOI:10.1016/j.foreco.2018.10.066URL [本文引用: 4]
ZanneAE, TankDC, CornwellWK, EastmanJM, SmithSA, FitzjohnRG, McGlinnDJ, O’MearaBC, MolesAT, ReichPB, RoyerDL, SoltisDE, StevensPF, WestobyM, WrightIJ, et al. (2014). Three keys to the radiation of angiosperms into freezing environments Nature, 506, 89-92. DOI:10.1038/nature12872URL [本文引用: 1]
ZhangQG, ZhangDY (2006). Species richness destabilizes ecosystem functioning in experimental aquatic microcosms Oikos, 112, 218-226. DOI:10.1111/oik.2006.112.issue-1URL [本文引用: 1]
ZhangXP, WangXP, ZhuB, ZongZJ, PengCH, FangJY(2008). Litter fall production in relation to environmental factors in northeast China’s forests Chinese Journal of Plant Ecology, 32, 1031-1040. [本文引用: 1]
ZhangY, ChenHYH (2015). Individual size inequality links forest diversity and above-ground biomass Journal of Ecology, 103, 1245-1252. DOI:10.1111/1365-2745.12425URL [本文引用: 2]
ZhengLT, ChenHYH, YanER (2019). Tree species diversity promotes litterfall productivity through crown complementarity in subtropical forests Journal of Ecology, 107, 1852-1861. DOI:10.1111/jec.2019.107.issue-4URL [本文引用: 2]
ZhouGY, GuanLL, WeiXH, ZhangDQ, ZhangQM, YanJH, WenDZ, LiuJX, LiuSG, HuangZL, KongGH, MoJM, YuQF (2007). Litterfall production along successional and altitudinal gradients of subtropical monsoon evergreen broadleaved forests in Guangdong, China Plant Ecology, 188, 77-89. DOI:10.1007/s11258-006-9149-9URL [本文引用: 1]
The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns 1 1999
... 本研究发现叶片碳含量对凋落物产量的影响较为明显(图2), 碳是构成植物叶片干物质含量的主要元素, 当植物将较多的碳用于叶片维持和防御结构上的投资时会相应减少在光合上的投资, 使植物生长变慢(He et al., 2006).本研究中叶片碳含量与凋落物产量的负相关性也支持这一点.以往研究多认为SLA、LN对生产力有显著影响(Niinemets et al., 2002), 但本研究中, 二者对凋落物产量的解释力明显低于LC.这可能说明影响凋落物产量和生长的性状存在一定差异, 因为凋落物产量只是林分生产力的一部分, 还涉及到功能性状如何影响资源分配等问题.比如, 在树叶凋落之前植物会通过将部分养分从叶中转运回体内储存, 来减弱植物对环境养分供应的依赖(Aerts & Chapin III, 1999).功能性状如何影响类似这样的资源分配过程, 还有待进一步研究. ...
Needle litterfall in stands of Pinus sylvestris L. in Sweden, in relation to site quality, stand age and latitude 1 1988
... 有研究认为凋落物产量沿演替序列增加(Zhou et al., 2007), 也有****发现凋落物产量在不同演替阶段无显著差异(Huang et al., 2017).一般而言, 在林分发育直至林冠郁闭之前每年的凋落物产量会迅速增加, 然后在相当长的一段时间内保持相对稳定(Albrektson, 1988), 本研究的结果与之一致.胜山自然保护区演替早期的林分密度、树木个体都较小, 林冠未达到郁闭导致其生产力有限, 所以凋落物产量显著低于其他演替阶段, 从演替中期至演替晚期林分高度郁闭(表1), 群落可捕获的光能大致相同, 使得3个演替阶段凋落物产量无显著差异. ...
Big-sized trees overrule remaining treesʼ attributes and species richness as determinants of aboveground biomass in tropical forests 4 2019
... 多年来的BEF研究表明, 除了多样性, 森林生产力还受到林分因子、群落物种组成(功能性状)、环境梯度等因素的强烈影响(Wu et al., 2015; Fotis et al., 2018; Ali et al., 2019).随着演替进展, 森林群落的多样性、林分因子、功能性状等都会发生显著的变化(Letcher & Chazdon, 2009), 而这些因素都可能影响生产力和凋落物产量, 因此森林演替系列是BEF研究的一个理想对象(Huang et al., 2017).如在林分因子中, 有研究发现较高的胸高断面积可以显著增加凋落物产量(Guo et al., 2019); 树冠的多层结构通过提高群落的光利用效率来增加凋落物产量(Zheng et al., 2019).功能性状对凋落物产量也有显著影响, 如Niinemets等(2002)发现较高的比叶面积和叶片N含量可以通过光合作用增加植物对周围环境中资源的获取能力和利用效率, 从而提高凋落物产量.关于多样性、林分因子、功能性状对生态系统功能的相对影响大小, 近期研究发现林分因子(Fotis et al., 2018; Ouyang et al., 2019)和功能性状(Garnier et al., 2004)对生物量、生产力影响明显大于多样性.对于凋落物产量是否也是如此, 目前还缺乏相关研究.关于生态系统的稳定性, 以往研究多集中于探讨多样性对其影响机制(Cleland, 2011; Mori et al., 2013).但近年来的研究表明, 森林生长在气候变化下的稳定性, 不仅受到气候变化、环境条件的影响, 林分因子(树高、密度、胸高断面积等)和物种组成也有着重要作用(温晓示等, 2019).这说明林分因子和功能性状同样可能影响森林生产力的稳定性, 但二者和生物多样性的相对重要性还少有研究, 但这对于如何进行森林经营以维持生态系统在未来气候变化下的稳定有着重要意义.本研究的另一个主要目标, 是利用凋落物产量的年际稳定性来探讨这一问题. ...
... 为研究林分因子对凋落物产量及其稳定性的影响, 在以往研究基础上, 本研究选择了4个变量, 包括: (1)最大树高(Hmax); (2)最大胸径(DBHmax); (3)胸高断面积(TBA); (4)林冠空隙度(gap fraction).近期研究表明群落中的大树对林分生物量、生产力及其波动都有重要影响, 因此本研究选择最大树高和最大胸径以反映大树的作用(Ali et al., 2019; Xu et al., 2019).胸高断面积与林分生物量高度相关(Wang et al., 2008), 也可以反映林分内的竞争强度(Zhang & Chen, 2015).由于林分密度、平均树高和平均胸径与上述几个林分因子共线性较强, 解释力也较低, 所以在最终的多元模型中没有选择这几个因子. ...
... 林分因子在早期BEF研究中未得到足够重视, 但近年越来越多的研究认为, 林分因子对生物量、生产力的作用可能大于多样性(Fotis et al., 2018; Ali et al., 2019; Ouyang et al., 2019).本研究的结果也表明, 林分因子对凋落物产量及其稳定性均有重要影响(图2).其中, 最大树高对凋落物产量影响最大, 这与近期研究发现大树对林分生产力有决定性影响的结果一致(Ali et al., 2019; Xu et al., 2019).大树往往在群落中更具竞争力, 它们可以达到较高的树冠高度并具有更高的光利用率(King et al., 2006), 因此大树可以显著提高森林的生产力(Xu et al., 2019).森林生产力是决定凋落量多少的关键因素, 生产力较低的林分凋落量自然较低.最大树高、胸高断面积都和林分生物量有密切的关系, 常被用作生物量的替代指标(Wang et al., 2008).因此, 本研究中二者对凋落物产量的重要影响(图2)也支持植被数量假说, 即生态系统过程的速率(如生产力、凋落物产量)主要决定于林分生物量(Lohbeck et al., 2015).此外, 林冠空隙度较低的群落可能具有更复杂的多层冠层结构, 不仅可更充分利用垂直生态位, 还为光合作用提供了更广泛的可吸收光谱(Atkins et al., 2018), 因此可更有效地利用光能来提高生产力. ...
... ).其中, 最大树高对凋落物产量影响最大, 这与近期研究发现大树对林分生产力有决定性影响的结果一致(Ali et al., 2019; Xu et al., 2019).大树往往在群落中更具竞争力, 它们可以达到较高的树冠高度并具有更高的光利用率(King et al., 2006), 因此大树可以显著提高森林的生产力(Xu et al., 2019).森林生产力是决定凋落量多少的关键因素, 生产力较低的林分凋落量自然较低.最大树高、胸高断面积都和林分生物量有密切的关系, 常被用作生物量的替代指标(Wang et al., 2008).因此, 本研究中二者对凋落物产量的重要影响(图2)也支持植被数量假说, 即生态系统过程的速率(如生产力、凋落物产量)主要决定于林分生物量(Lohbeck et al., 2015).此外, 林冠空隙度较低的群落可能具有更复杂的多层冠层结构, 不仅可更充分利用垂直生态位, 还为光合作用提供了更广泛的可吸收光谱(Atkins et al., 2018), 因此可更有效地利用光能来提高生产力. ...
Temporal patterns in rates of community change during succession 1 2007
Forest canopy structural complexity and light absorption relationships at the subcontinental scale 1 2018
... 林分因子在早期BEF研究中未得到足够重视, 但近年越来越多的研究认为, 林分因子对生物量、生产力的作用可能大于多样性(Fotis et al., 2018; Ali et al., 2019; Ouyang et al., 2019).本研究的结果也表明, 林分因子对凋落物产量及其稳定性均有重要影响(图2).其中, 最大树高对凋落物产量影响最大, 这与近期研究发现大树对林分生产力有决定性影响的结果一致(Ali et al., 2019; Xu et al., 2019).大树往往在群落中更具竞争力, 它们可以达到较高的树冠高度并具有更高的光利用率(King et al., 2006), 因此大树可以显著提高森林的生产力(Xu et al., 2019).森林生产力是决定凋落量多少的关键因素, 生产力较低的林分凋落量自然较低.最大树高、胸高断面积都和林分生物量有密切的关系, 常被用作生物量的替代指标(Wang et al., 2008).因此, 本研究中二者对凋落物产量的重要影响(图2)也支持植被数量假说, 即生态系统过程的速率(如生产力、凋落物产量)主要决定于林分生物量(Lohbeck et al., 2015).此外, 林冠空隙度较低的群落可能具有更复杂的多层冠层结构, 不仅可更充分利用垂直生态位, 还为光合作用提供了更广泛的可吸收光谱(Atkins et al., 2018), 因此可更有效地利用光能来提高生产力. ...
Using phylogenetic, functional and trait diversity to understand patterns of plant community productivity 2 2009
... 多年来的研究一般认为, 物种丰富的植物群落可支持较高的生物量和生产力, 并使群落的组成和功能在长期的动态变化中维持较高的稳定性.尽管多数研究认为多样性与生产力之间为正相关关系(Cavanaugh et al., 2014; Liang et al., 2016), 但也有部分研究结果发现两者之间为负相关关系或不相关(Finegan et al., 2015).在本研究中3个生物多样性指标与凋落物产量均有显著的正相关关系(表2), 其中物种多样性增加了物种数量和物种多度分布的均匀程度; 功能多样性使不同物种能够更有效地利用功能生态位、从而获取更多的资源(Harpole & Tilman, 2007); 而系统发育多样性增加了物种间进化关系和功能上的差异(Cadotte et al., 2009).多样性高的群落可以通过更有效地利用光照、养分和水等资源来提高生态系统生产力, 从而提高凋落物产量.不过, 通过消除协同变化的因子的影响, 本研究发现多样性在林分因子、功能性状之外对凋落物产量的独立作用很微弱(图3), 这说明多样性主要是通过与其他因素的共同作用来影响凋落物产量(Wu et al., 2015; Ouyang et al., 2019). ...
Measuring net primary production in forests: concepts and field methods 1 2001
... 凋落物是生态系统净初级生产力的一个重要贡献者(Clark et al., 2001), 也是联结生态系统地上和地下碳循环过程的一个关键环节, 对维持森林土壤肥力以及森林生态系统结构和功能的稳定性具有重要作用(Vogt et al., 1986).以往对凋落物产量已有较多研究, 但多集中于研究凋落物的组分、季节动态, 以及气候、林型等因素如何影响凋落物产量(张新平等, 2008; 贾丙瑞等, 2016).关于多样性对凋落物产量的影响, 则研究相对较少(Guo et al., 2019; Zheng et al., 2019).其中, 多样性如何影响凋落物产量的稳定性的研究还十分匮乏(Huang et al., 2017).近年来不少研究认为, 多样性不但能够促进群落生产力, 还可提高生态系统在环境波动下的稳定性(Cardinale et al., 2012).凋落物作为森林生产力的一个重要组分, 多样性是否对其产量及其稳定性也有显著的促进作用? 这是本文主要研究的一个问题. ...
Biodiversity and ecosystem stability 3 2011
... 多年来的BEF研究表明, 除了多样性, 森林生产力还受到林分因子、群落物种组成(功能性状)、环境梯度等因素的强烈影响(Wu et al., 2015; Fotis et al., 2018; Ali et al., 2019).随着演替进展, 森林群落的多样性、林分因子、功能性状等都会发生显著的变化(Letcher & Chazdon, 2009), 而这些因素都可能影响生产力和凋落物产量, 因此森林演替系列是BEF研究的一个理想对象(Huang et al., 2017).如在林分因子中, 有研究发现较高的胸高断面积可以显著增加凋落物产量(Guo et al., 2019); 树冠的多层结构通过提高群落的光利用效率来增加凋落物产量(Zheng et al., 2019).功能性状对凋落物产量也有显著影响, 如Niinemets等(2002)发现较高的比叶面积和叶片N含量可以通过光合作用增加植物对周围环境中资源的获取能力和利用效率, 从而提高凋落物产量.关于多样性、林分因子、功能性状对生态系统功能的相对影响大小, 近期研究发现林分因子(Fotis et al., 2018; Ouyang et al., 2019)和功能性状(Garnier et al., 2004)对生物量、生产力影响明显大于多样性.对于凋落物产量是否也是如此, 目前还缺乏相关研究.关于生态系统的稳定性, 以往研究多集中于探讨多样性对其影响机制(Cleland, 2011; Mori et al., 2013).但近年来的研究表明, 森林生长在气候变化下的稳定性, 不仅受到气候变化、环境条件的影响, 林分因子(树高、密度、胸高断面积等)和物种组成也有着重要作用(温晓示等, 2019).这说明林分因子和功能性状同样可能影响森林生产力的稳定性, 但二者和生物多样性的相对重要性还少有研究, 但这对于如何进行森林经营以维持生态系统在未来气候变化下的稳定有着重要意义.本研究的另一个主要目标, 是利用凋落物产量的年际稳定性来探讨这一问题. ...
... 不少研究发现生物多样性可提高生态系统稳定性(Caldeira et al., 2005), 但也有研究认为二者没有显著相关关系(Zhang & Zhang, 2006).本研究发现虽然物种丰富度与凋落物稳定性的相关性较高(表2), 但在考虑了其他生物和非生物因子共同作用时(图2), 功能多样性更能体现生物多样性对稳定性的作用(Mazzochini et al., 2019).相对于物种和系统发育多样性, 功能多样性可以更好地反应生态系统的功能冗余度和生态位重叠(Tilman et al., 1997).当一个生态系统中有多个具有相似生态位的物种时, 可能会产生功能冗余, 但是当环境变化而使某个物种丧失或衰退时, 这种功能冗余会使其生态位被另一个物种所补偿, 从而起到增强生态系统生产力稳定性的作用(Cleland, 2011), 即功能多样性可以通过物种间生活史策略的互补提高群落的稳定性(Mori et al., 2013).物种和系统发育多样性也进入了凋落物产量稳定性的模型, 不过影响较弱(图2).物种丰富度的解释力可能反映了物种间生长发育的不同步性对生态系统稳定性的影响(Loreau & de Mazancourt, 2013; Morin et al., 2014), 系统发育多样性的解释力则可能体现了物种间进化上的差异导致的资源获取、储存和利用等策略的不同(Souza et al., 2016), 从而提高了生态系统稳定性.综上, 本研究的结果表明, 物种、功能和系统发育多样性共同影响凋落物产量稳定性, 其中功能多样性可能起着最主要的作用.此外, 变异分离分析表明, 多样性还可通过和林分因子、性状的协同作用影响稳定性, 也支持多样性较高的演替后期林分具有较高的稳定性. ...
... 以往关于生态系统稳定性的研究多探讨其与多样性的关系(Cleland, 2011; Mori et al., 2013), 而很少考虑林分因子.本研究的结果表明, 最大胸径、TBA等林分因子对凋落物产量稳定性的影响仅次于功能多样性(图2), 并通过与多样性、性状的协同作用影响稳定性(图3).大直径的树木通常具有更高的树高和更大的冠幅, 能够调节林分水平叶面积、可利用水资源和小气候(Martin et al., 2001).Needham等(2016)的研究表明, 大树较多的森林具有较高的结构异质性, 这样的森林在应对环境波动时能更好地维持生态系统功能.Xu等(2019)对东北地区纬度梯度上不同类型森林的研究表明, 大树占据了绝大多数林分生物量和生产力, 因此林分生产力的稳定性主要决定于大树.这些结果都说明, 除了多样性之外, 林分因子(尤其是大树), 也是影响森林生产力稳定的重要因素. ...
Does functional trait diversity predict above- ground biomass and productivity of tropical forests? Testing three alternative hypotheses 1 2015
... 多年来的研究一般认为, 物种丰富的植物群落可支持较高的生物量和生产力, 并使群落的组成和功能在长期的动态变化中维持较高的稳定性.尽管多数研究认为多样性与生产力之间为正相关关系(Cavanaugh et al., 2014; Liang et al., 2016), 但也有部分研究结果发现两者之间为负相关关系或不相关(Finegan et al., 2015).在本研究中3个生物多样性指标与凋落物产量均有显著的正相关关系(表2), 其中物种多样性增加了物种数量和物种多度分布的均匀程度; 功能多样性使不同物种能够更有效地利用功能生态位、从而获取更多的资源(Harpole & Tilman, 2007); 而系统发育多样性增加了物种间进化关系和功能上的差异(Cadotte et al., 2009).多样性高的群落可以通过更有效地利用光照、养分和水等资源来提高生态系统生产力, 从而提高凋落物产量.不过, 通过消除协同变化的因子的影响, 本研究发现多样性在林分因子、功能性状之外对凋落物产量的独立作用很微弱(图3), 这说明多样性主要是通过与其他因素的共同作用来影响凋落物产量(Wu et al., 2015; Ouyang et al., 2019). ...
The spatial and temporal dynamics of species interactions in mixed-species forests: from pattern to process 1 2014
... 近半个世纪以来快速的全球气候变暖直接或间接导致了生物多样性的锐减, 改变了生态系统的结构, 严重影响了生态系统的服务功能(Tilman, 2000; Wan et al., 2007).由于环境变迁和人类活动造成的生物多样性丧失对全球生态系统带来的影响, 使得生物多样性-生态系统功能(BEF)关系一直备受关注(Lasky et al., 2014; Zhang & Chen, 2015).以往的研究一般认为生物多样性能够促进生产力等生态系统功能, 目前已经提出的两种主要假说为: 质量比假说和互补假说.其中, 质量比假说认为生态系统功能主要由优势种的组成及其功能属性决定(Grime, 1998); 互补假说认为群落生产力的增加是由于物种间的相互促进作用和生态位分化导致竞争减少, 使物种获得的资源更多, 从而增加生态系统的生产力(Tilman et al., 1997; Forrester, 2014).此外, 有关生态系统功能的影响机制, 还有其他一些假说.如有研究认为森林生态系统功能主要由植被生物量所驱动(Lohbeck et al., 2015), 即植被数量假说. ...
Above-ground biomass is driven by mass-ratio effects and stand structural attributes in a temperate deciduous forest 3 2018
... 多年来的BEF研究表明, 除了多样性, 森林生产力还受到林分因子、群落物种组成(功能性状)、环境梯度等因素的强烈影响(Wu et al., 2015; Fotis et al., 2018; Ali et al., 2019).随着演替进展, 森林群落的多样性、林分因子、功能性状等都会发生显著的变化(Letcher & Chazdon, 2009), 而这些因素都可能影响生产力和凋落物产量, 因此森林演替系列是BEF研究的一个理想对象(Huang et al., 2017).如在林分因子中, 有研究发现较高的胸高断面积可以显著增加凋落物产量(Guo et al., 2019); 树冠的多层结构通过提高群落的光利用效率来增加凋落物产量(Zheng et al., 2019).功能性状对凋落物产量也有显著影响, 如Niinemets等(2002)发现较高的比叶面积和叶片N含量可以通过光合作用增加植物对周围环境中资源的获取能力和利用效率, 从而提高凋落物产量.关于多样性、林分因子、功能性状对生态系统功能的相对影响大小, 近期研究发现林分因子(Fotis et al., 2018; Ouyang et al., 2019)和功能性状(Garnier et al., 2004)对生物量、生产力影响明显大于多样性.对于凋落物产量是否也是如此, 目前还缺乏相关研究.关于生态系统的稳定性, 以往研究多集中于探讨多样性对其影响机制(Cleland, 2011; Mori et al., 2013).但近年来的研究表明, 森林生长在气候变化下的稳定性, 不仅受到气候变化、环境条件的影响, 林分因子(树高、密度、胸高断面积等)和物种组成也有着重要作用(温晓示等, 2019).这说明林分因子和功能性状同样可能影响森林生产力的稳定性, 但二者和生物多样性的相对重要性还少有研究, 但这对于如何进行森林经营以维持生态系统在未来气候变化下的稳定有着重要意义.本研究的另一个主要目标, 是利用凋落物产量的年际稳定性来探讨这一问题. ...
... )发现较高的比叶面积和叶片N含量可以通过光合作用增加植物对周围环境中资源的获取能力和利用效率, 从而提高凋落物产量.关于多样性、林分因子、功能性状对生态系统功能的相对影响大小, 近期研究发现林分因子(Fotis et al., 2018; Ouyang et al., 2019)和功能性状(Garnier et al., 2004)对生物量、生产力影响明显大于多样性.对于凋落物产量是否也是如此, 目前还缺乏相关研究.关于生态系统的稳定性, 以往研究多集中于探讨多样性对其影响机制(Cleland, 2011; Mori et al., 2013).但近年来的研究表明, 森林生长在气候变化下的稳定性, 不仅受到气候变化、环境条件的影响, 林分因子(树高、密度、胸高断面积等)和物种组成也有着重要作用(温晓示等, 2019).这说明林分因子和功能性状同样可能影响森林生产力的稳定性, 但二者和生物多样性的相对重要性还少有研究, 但这对于如何进行森林经营以维持生态系统在未来气候变化下的稳定有着重要意义.本研究的另一个主要目标, 是利用凋落物产量的年际稳定性来探讨这一问题. ...
... 林分因子在早期BEF研究中未得到足够重视, 但近年越来越多的研究认为, 林分因子对生物量、生产力的作用可能大于多样性(Fotis et al., 2018; Ali et al., 2019; Ouyang et al., 2019).本研究的结果也表明, 林分因子对凋落物产量及其稳定性均有重要影响(图2).其中, 最大树高对凋落物产量影响最大, 这与近期研究发现大树对林分生产力有决定性影响的结果一致(Ali et al., 2019; Xu et al., 2019).大树往往在群落中更具竞争力, 它们可以达到较高的树冠高度并具有更高的光利用率(King et al., 2006), 因此大树可以显著提高森林的生产力(Xu et al., 2019).森林生产力是决定凋落量多少的关键因素, 生产力较低的林分凋落量自然较低.最大树高、胸高断面积都和林分生物量有密切的关系, 常被用作生物量的替代指标(Wang et al., 2008).因此, 本研究中二者对凋落物产量的重要影响(图2)也支持植被数量假说, 即生态系统过程的速率(如生产力、凋落物产量)主要决定于林分生物量(Lohbeck et al., 2015).此外, 林冠空隙度较低的群落可能具有更复杂的多层冠层结构, 不仅可更充分利用垂直生态位, 还为光合作用提供了更广泛的可吸收光谱(Atkins et al., 2018), 因此可更有效地利用光能来提高生产力. ...
... 多年来的BEF研究表明, 除了多样性, 森林生产力还受到林分因子、群落物种组成(功能性状)、环境梯度等因素的强烈影响(Wu et al., 2015; Fotis et al., 2018; Ali et al., 2019).随着演替进展, 森林群落的多样性、林分因子、功能性状等都会发生显著的变化(Letcher & Chazdon, 2009), 而这些因素都可能影响生产力和凋落物产量, 因此森林演替系列是BEF研究的一个理想对象(Huang et al., 2017).如在林分因子中, 有研究发现较高的胸高断面积可以显著增加凋落物产量(Guo et al., 2019); 树冠的多层结构通过提高群落的光利用效率来增加凋落物产量(Zheng et al., 2019).功能性状对凋落物产量也有显著影响, 如Niinemets等(2002)发现较高的比叶面积和叶片N含量可以通过光合作用增加植物对周围环境中资源的获取能力和利用效率, 从而提高凋落物产量.关于多样性、林分因子、功能性状对生态系统功能的相对影响大小, 近期研究发现林分因子(Fotis et al., 2018; Ouyang et al., 2019)和功能性状(Garnier et al., 2004)对生物量、生产力影响明显大于多样性.对于凋落物产量是否也是如此, 目前还缺乏相关研究.关于生态系统的稳定性, 以往研究多集中于探讨多样性对其影响机制(Cleland, 2011; Mori et al., 2013).但近年来的研究表明, 森林生长在气候变化下的稳定性, 不仅受到气候变化、环境条件的影响, 林分因子(树高、密度、胸高断面积等)和物种组成也有着重要作用(温晓示等, 2019).这说明林分因子和功能性状同样可能影响森林生产力的稳定性, 但二者和生物多样性的相对重要性还少有研究, 但这对于如何进行森林经营以维持生态系统在未来气候变化下的稳定有着重要意义.本研究的另一个主要目标, 是利用凋落物产量的年际稳定性来探讨这一问题. ...
... 叶功能性状的群落加权平均值可以用来表征群落中优势种性状与生态系统功能的关系(Garnier et al., 2004).根据各样地物种的相对胸高断面积数据, 使用R软件中“FD”包的“dbfd”函数计算每个样地叶功能性状的群落加权平均值.需要说明的是, 由于非优势种的胸高断面积占比很小, 所以本研究使用优势种性状计算群落加权平均值, 这也是多数研究采用的方法(Garnier et al., 2004; Lasky et al., 2014). ...
... ).根据各样地物种的相对胸高断面积数据, 使用R软件中“FD”包的“dbfd”函数计算每个样地叶功能性状的群落加权平均值.需要说明的是, 由于非优势种的胸高断面积占比很小, 所以本研究使用优势种性状计算群落加权平均值, 这也是多数研究采用的方法(Garnier et al., 2004; Lasky et al., 2014). ...
Benefits of plant diversity to ecosystems: immediate, filter and founder effects 2 1998
... 近半个世纪以来快速的全球气候变暖直接或间接导致了生物多样性的锐减, 改变了生态系统的结构, 严重影响了生态系统的服务功能(Tilman, 2000; Wan et al., 2007).由于环境变迁和人类活动造成的生物多样性丧失对全球生态系统带来的影响, 使得生物多样性-生态系统功能(BEF)关系一直备受关注(Lasky et al., 2014; Zhang & Chen, 2015).以往的研究一般认为生物多样性能够促进生产力等生态系统功能, 目前已经提出的两种主要假说为: 质量比假说和互补假说.其中, 质量比假说认为生态系统功能主要由优势种的组成及其功能属性决定(Grime, 1998); 互补假说认为群落生产力的增加是由于物种间的相互促进作用和生态位分化导致竞争减少, 使物种获得的资源更多, 从而增加生态系统的生产力(Tilman et al., 1997; Forrester, 2014).此外, 有关生态系统功能的影响机制, 还有其他一些假说.如有研究认为森林生态系统功能主要由植被生物量所驱动(Lohbeck et al., 2015), 即植被数量假说. ...
... 本研究结果表明凋落物产量稳定性与SLA显著负相关(表2).叶片的功能性状对气候比较敏感, 反映了植物对环境的高度适应能力和在复杂生境下的自我调控能力(Scoffoni et al., 2011), 叶片大小随着年平均气温及年降水量的降低而减小(Peppe et al., 2011).叶面积可显著影响植物的生长速度(Souza et al., 2016), 具有较低SLA的物种的生长策略为保守型, 其表现为生长速度较慢, 但更能适应环境变化且具有更高的稳定性.比如, 低SLA的物种增加了在木质素等结构物质上的投资(Poorter & De Jong, 1999), 并产生具有较小细胞和较厚细胞壁的叶片, 从而降低细胞弹性并减少叶片水势(Klein, 2014), 使它们在干旱时期蒸腾的水分更少(Sterck et al., 2011), 从而在外部环境变化时更好地保持水分平衡, 进而影响凋落物产量的稳定性.此外, 本研究结果表明, 群落水平的功能性状对凋落物产量及其稳定性都有重要影响(图2, 图3), 这也支持质量比假说(Grime, 1998), 说明除了多样性和林分因子, 优势种的功能性状对群落生产力及生态系统的稳定性也有重要影响. ...
Predominance of abiotic drivers in the relationship between species diversity and litterfall production in a tropical karst seasonal rainforest 3 2019
... 凋落物是生态系统净初级生产力的一个重要贡献者(Clark et al., 2001), 也是联结生态系统地上和地下碳循环过程的一个关键环节, 对维持森林土壤肥力以及森林生态系统结构和功能的稳定性具有重要作用(Vogt et al., 1986).以往对凋落物产量已有较多研究, 但多集中于研究凋落物的组分、季节动态, 以及气候、林型等因素如何影响凋落物产量(张新平等, 2008; 贾丙瑞等, 2016).关于多样性对凋落物产量的影响, 则研究相对较少(Guo et al., 2019; Zheng et al., 2019).其中, 多样性如何影响凋落物产量的稳定性的研究还十分匮乏(Huang et al., 2017).近年来不少研究认为, 多样性不但能够促进群落生产力, 还可提高生态系统在环境波动下的稳定性(Cardinale et al., 2012).凋落物作为森林生产力的一个重要组分, 多样性是否对其产量及其稳定性也有显著的促进作用? 这是本文主要研究的一个问题. ...
... 多年来的BEF研究表明, 除了多样性, 森林生产力还受到林分因子、群落物种组成(功能性状)、环境梯度等因素的强烈影响(Wu et al., 2015; Fotis et al., 2018; Ali et al., 2019).随着演替进展, 森林群落的多样性、林分因子、功能性状等都会发生显著的变化(Letcher & Chazdon, 2009), 而这些因素都可能影响生产力和凋落物产量, 因此森林演替系列是BEF研究的一个理想对象(Huang et al., 2017).如在林分因子中, 有研究发现较高的胸高断面积可以显著增加凋落物产量(Guo et al., 2019); 树冠的多层结构通过提高群落的光利用效率来增加凋落物产量(Zheng et al., 2019).功能性状对凋落物产量也有显著影响, 如Niinemets等(2002)发现较高的比叶面积和叶片N含量可以通过光合作用增加植物对周围环境中资源的获取能力和利用效率, 从而提高凋落物产量.关于多样性、林分因子、功能性状对生态系统功能的相对影响大小, 近期研究发现林分因子(Fotis et al., 2018; Ouyang et al., 2019)和功能性状(Garnier et al., 2004)对生物量、生产力影响明显大于多样性.对于凋落物产量是否也是如此, 目前还缺乏相关研究.关于生态系统的稳定性, 以往研究多集中于探讨多样性对其影响机制(Cleland, 2011; Mori et al., 2013).但近年来的研究表明, 森林生长在气候变化下的稳定性, 不仅受到气候变化、环境条件的影响, 林分因子(树高、密度、胸高断面积等)和物种组成也有着重要作用(温晓示等, 2019).这说明林分因子和功能性状同样可能影响森林生产力的稳定性, 但二者和生物多样性的相对重要性还少有研究, 但这对于如何进行森林经营以维持生态系统在未来气候变化下的稳定有着重要意义.本研究的另一个主要目标, 是利用凋落物产量的年际稳定性来探讨这一问题. ...
Grassland species loss resulting from reduced niche dimension 1 2007
... 多年来的研究一般认为, 物种丰富的植物群落可支持较高的生物量和生产力, 并使群落的组成和功能在长期的动态变化中维持较高的稳定性.尽管多数研究认为多样性与生产力之间为正相关关系(Cavanaugh et al., 2014; Liang et al., 2016), 但也有部分研究结果发现两者之间为负相关关系或不相关(Finegan et al., 2015).在本研究中3个生物多样性指标与凋落物产量均有显著的正相关关系(表2), 其中物种多样性增加了物种数量和物种多度分布的均匀程度; 功能多样性使不同物种能够更有效地利用功能生态位、从而获取更多的资源(Harpole & Tilman, 2007); 而系统发育多样性增加了物种间进化关系和功能上的差异(Cadotte et al., 2009).多样性高的群落可以通过更有效地利用光照、养分和水等资源来提高生态系统生产力, 从而提高凋落物产量.不过, 通过消除协同变化的因子的影响, 本研究发现多样性在林分因子、功能性状之外对凋落物产量的独立作用很微弱(图3), 这说明多样性主要是通过与其他因素的共同作用来影响凋落物产量(Wu et al., 2015; Ouyang et al., 2019). ...
Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China 1 2006
... 本研究发现叶片碳含量对凋落物产量的影响较为明显(图2), 碳是构成植物叶片干物质含量的主要元素, 当植物将较多的碳用于叶片维持和防御结构上的投资时会相应减少在光合上的投资, 使植物生长变慢(He et al., 2006).本研究中叶片碳含量与凋落物产量的负相关性也支持这一点.以往研究多认为SLA、LN对生产力有显著影响(Niinemets et al., 2002), 但本研究中, 二者对凋落物产量的解释力明显低于LC.这可能说明影响凋落物产量和生长的性状存在一定差异, 因为凋落物产量只是林分生产力的一部分, 还涉及到功能性状如何影响资源分配等问题.比如, 在树叶凋落之前植物会通过将部分养分从叶中转运回体内储存, 来减弱植物对环境养分供应的依赖(Aerts & Chapin III, 1999).功能性状如何影响类似这样的资源分配过程, 还有待进一步研究. ...
Positive effects of tree species diversity on litterfall quantity and quality along a secondary successional chronosequence in a subtropical forest 3 2017
... 凋落物是生态系统净初级生产力的一个重要贡献者(Clark et al., 2001), 也是联结生态系统地上和地下碳循环过程的一个关键环节, 对维持森林土壤肥力以及森林生态系统结构和功能的稳定性具有重要作用(Vogt et al., 1986).以往对凋落物产量已有较多研究, 但多集中于研究凋落物的组分、季节动态, 以及气候、林型等因素如何影响凋落物产量(张新平等, 2008; 贾丙瑞等, 2016).关于多样性对凋落物产量的影响, 则研究相对较少(Guo et al., 2019; Zheng et al., 2019).其中, 多样性如何影响凋落物产量的稳定性的研究还十分匮乏(Huang et al., 2017).近年来不少研究认为, 多样性不但能够促进群落生产力, 还可提高生态系统在环境波动下的稳定性(Cardinale et al., 2012).凋落物作为森林生产力的一个重要组分, 多样性是否对其产量及其稳定性也有显著的促进作用? 这是本文主要研究的一个问题. ...
... 多年来的BEF研究表明, 除了多样性, 森林生产力还受到林分因子、群落物种组成(功能性状)、环境梯度等因素的强烈影响(Wu et al., 2015; Fotis et al., 2018; Ali et al., 2019).随着演替进展, 森林群落的多样性、林分因子、功能性状等都会发生显著的变化(Letcher & Chazdon, 2009), 而这些因素都可能影响生产力和凋落物产量, 因此森林演替系列是BEF研究的一个理想对象(Huang et al., 2017).如在林分因子中, 有研究发现较高的胸高断面积可以显著增加凋落物产量(Guo et al., 2019); 树冠的多层结构通过提高群落的光利用效率来增加凋落物产量(Zheng et al., 2019).功能性状对凋落物产量也有显著影响, 如Niinemets等(2002)发现较高的比叶面积和叶片N含量可以通过光合作用增加植物对周围环境中资源的获取能力和利用效率, 从而提高凋落物产量.关于多样性、林分因子、功能性状对生态系统功能的相对影响大小, 近期研究发现林分因子(Fotis et al., 2018; Ouyang et al., 2019)和功能性状(Garnier et al., 2004)对生物量、生产力影响明显大于多样性.对于凋落物产量是否也是如此, 目前还缺乏相关研究.关于生态系统的稳定性, 以往研究多集中于探讨多样性对其影响机制(Cleland, 2011; Mori et al., 2013).但近年来的研究表明, 森林生长在气候变化下的稳定性, 不仅受到气候变化、环境条件的影响, 林分因子(树高、密度、胸高断面积等)和物种组成也有着重要作用(温晓示等, 2019).这说明林分因子和功能性状同样可能影响森林生产力的稳定性, 但二者和生物多样性的相对重要性还少有研究, 但这对于如何进行森林经营以维持生态系统在未来气候变化下的稳定有着重要意义.本研究的另一个主要目标, 是利用凋落物产量的年际稳定性来探讨这一问题. ...
... 有研究认为凋落物产量沿演替序列增加(Zhou et al., 2007), 也有****发现凋落物产量在不同演替阶段无显著差异(Huang et al., 2017).一般而言, 在林分发育直至林冠郁闭之前每年的凋落物产量会迅速增加, 然后在相当长的一段时间内保持相对稳定(Albrektson, 1988), 本研究的结果与之一致.胜山自然保护区演替早期的林分密度、树木个体都较小, 林冠未达到郁闭导致其生产力有限, 所以凋落物产量显著低于其他演替阶段, 从演替中期至演替晚期林分高度郁闭(表1), 群落可捕获的光能大致相同, 使得3个演替阶段凋落物产量无显著差异. ...
Biodiversity, productivity and the temporal stability of productivity: patterns and processes 1 2009
Growth and mortality are related to adult tree size in a Malaysian mixed dipterocarp forest 1 2006
... 林分因子在早期BEF研究中未得到足够重视, 但近年越来越多的研究认为, 林分因子对生物量、生产力的作用可能大于多样性(Fotis et al., 2018; Ali et al., 2019; Ouyang et al., 2019).本研究的结果也表明, 林分因子对凋落物产量及其稳定性均有重要影响(图2).其中, 最大树高对凋落物产量影响最大, 这与近期研究发现大树对林分生产力有决定性影响的结果一致(Ali et al., 2019; Xu et al., 2019).大树往往在群落中更具竞争力, 它们可以达到较高的树冠高度并具有更高的光利用率(King et al., 2006), 因此大树可以显著提高森林的生产力(Xu et al., 2019).森林生产力是决定凋落量多少的关键因素, 生产力较低的林分凋落量自然较低.最大树高、胸高断面积都和林分生物量有密切的关系, 常被用作生物量的替代指标(Wang et al., 2008).因此, 本研究中二者对凋落物产量的重要影响(图2)也支持植被数量假说, 即生态系统过程的速率(如生产力、凋落物产量)主要决定于林分生物量(Lohbeck et al., 2015).此外, 林冠空隙度较低的群落可能具有更复杂的多层冠层结构, 不仅可更充分利用垂直生态位, 还为光合作用提供了更广泛的可吸收光谱(Atkins et al., 2018), 因此可更有效地利用光能来提高生产力. ...
The global economic long-term potential of modern biomass in a climate-constrained world 1 2014
... 本研究结果表明凋落物产量稳定性与SLA显著负相关(表2).叶片的功能性状对气候比较敏感, 反映了植物对环境的高度适应能力和在复杂生境下的自我调控能力(Scoffoni et al., 2011), 叶片大小随着年平均气温及年降水量的降低而减小(Peppe et al., 2011).叶面积可显著影响植物的生长速度(Souza et al., 2016), 具有较低SLA的物种的生长策略为保守型, 其表现为生长速度较慢, 但更能适应环境变化且具有更高的稳定性.比如, 低SLA的物种增加了在木质素等结构物质上的投资(Poorter & De Jong, 1999), 并产生具有较小细胞和较厚细胞壁的叶片, 从而降低细胞弹性并减少叶片水势(Klein, 2014), 使它们在干旱时期蒸腾的水分更少(Sterck et al., 2011), 从而在外部环境变化时更好地保持水分平衡, 进而影响凋落物产量的稳定性.此外, 本研究结果表明, 群落水平的功能性状对凋落物产量及其稳定性都有重要影响(图2, 图3), 这也支持质量比假说(Grime, 1998), 说明除了多样性和林分因子, 优势种的功能性状对群落生产力及生态系统的稳定性也有重要影响. ...
A distance-based framework for measuring functional diversity from multiple traits 1 2010
The relationship between tree biodiversity and biomass dynamics changes with tropical forest succession 2 2014
... 近半个世纪以来快速的全球气候变暖直接或间接导致了生物多样性的锐减, 改变了生态系统的结构, 严重影响了生态系统的服务功能(Tilman, 2000; Wan et al., 2007).由于环境变迁和人类活动造成的生物多样性丧失对全球生态系统带来的影响, 使得生物多样性-生态系统功能(BEF)关系一直备受关注(Lasky et al., 2014; Zhang & Chen, 2015).以往的研究一般认为生物多样性能够促进生产力等生态系统功能, 目前已经提出的两种主要假说为: 质量比假说和互补假说.其中, 质量比假说认为生态系统功能主要由优势种的组成及其功能属性决定(Grime, 1998); 互补假说认为群落生产力的增加是由于物种间的相互促进作用和生态位分化导致竞争减少, 使物种获得的资源更多, 从而增加生态系统的生产力(Tilman et al., 1997; Forrester, 2014).此外, 有关生态系统功能的影响机制, 还有其他一些假说.如有研究认为森林生态系统功能主要由植被生物量所驱动(Lohbeck et al., 2015), 即植被数量假说. ...
... 叶功能性状的群落加权平均值可以用来表征群落中优势种性状与生态系统功能的关系(Garnier et al., 2004).根据各样地物种的相对胸高断面积数据, 使用R软件中“FD”包的“dbfd”函数计算每个样地叶功能性状的群落加权平均值.需要说明的是, 由于非优势种的胸高断面积占比很小, 所以本研究使用优势种性状计算群落加权平均值, 这也是多数研究采用的方法(Garnier et al., 2004; Lasky et al., 2014). ...
Rapid recovery of biomass, species richness, and species composition in a forest chronosequence in northeastern 1 2009
... 多年来的BEF研究表明, 除了多样性, 森林生产力还受到林分因子、群落物种组成(功能性状)、环境梯度等因素的强烈影响(Wu et al., 2015; Fotis et al., 2018; Ali et al., 2019).随着演替进展, 森林群落的多样性、林分因子、功能性状等都会发生显著的变化(Letcher & Chazdon, 2009), 而这些因素都可能影响生产力和凋落物产量, 因此森林演替系列是BEF研究的一个理想对象(Huang et al., 2017).如在林分因子中, 有研究发现较高的胸高断面积可以显著增加凋落物产量(Guo et al., 2019); 树冠的多层结构通过提高群落的光利用效率来增加凋落物产量(Zheng et al., 2019).功能性状对凋落物产量也有显著影响, 如Niinemets等(2002)发现较高的比叶面积和叶片N含量可以通过光合作用增加植物对周围环境中资源的获取能力和利用效率, 从而提高凋落物产量.关于多样性、林分因子、功能性状对生态系统功能的相对影响大小, 近期研究发现林分因子(Fotis et al., 2018; Ouyang et al., 2019)和功能性状(Garnier et al., 2004)对生物量、生产力影响明显大于多样性.对于凋落物产量是否也是如此, 目前还缺乏相关研究.关于生态系统的稳定性, 以往研究多集中于探讨多样性对其影响机制(Cleland, 2011; Mori et al., 2013).但近年来的研究表明, 森林生长在气候变化下的稳定性, 不仅受到气候变化、环境条件的影响, 林分因子(树高、密度、胸高断面积等)和物种组成也有着重要作用(温晓示等, 2019).这说明林分因子和功能性状同样可能影响森林生产力的稳定性, 但二者和生物多样性的相对重要性还少有研究, 但这对于如何进行森林经营以维持生态系统在未来气候变化下的稳定有着重要意义.本研究的另一个主要目标, 是利用凋落物产量的年际稳定性来探讨这一问题. ...
Positive biodiversity-productivity relationship predominant in global forests 1 2016
... 多年来的研究一般认为, 物种丰富的植物群落可支持较高的生物量和生产力, 并使群落的组成和功能在长期的动态变化中维持较高的稳定性.尽管多数研究认为多样性与生产力之间为正相关关系(Cavanaugh et al., 2014; Liang et al., 2016), 但也有部分研究结果发现两者之间为负相关关系或不相关(Finegan et al., 2015).在本研究中3个生物多样性指标与凋落物产量均有显著的正相关关系(表2), 其中物种多样性增加了物种数量和物种多度分布的均匀程度; 功能多样性使不同物种能够更有效地利用功能生态位、从而获取更多的资源(Harpole & Tilman, 2007); 而系统发育多样性增加了物种间进化关系和功能上的差异(Cadotte et al., 2009).多样性高的群落可以通过更有效地利用光照、养分和水等资源来提高生态系统生产力, 从而提高凋落物产量.不过, 通过消除协同变化的因子的影响, 本研究发现多样性在林分因子、功能性状之外对凋落物产量的独立作用很微弱(图3), 这说明多样性主要是通过与其他因素的共同作用来影响凋落物产量(Wu et al., 2015; Ouyang et al., 2019). ...
Biomass is the main driver of changes in ecosystem process rates during tropical forest succession 2 2015
... 近半个世纪以来快速的全球气候变暖直接或间接导致了生物多样性的锐减, 改变了生态系统的结构, 严重影响了生态系统的服务功能(Tilman, 2000; Wan et al., 2007).由于环境变迁和人类活动造成的生物多样性丧失对全球生态系统带来的影响, 使得生物多样性-生态系统功能(BEF)关系一直备受关注(Lasky et al., 2014; Zhang & Chen, 2015).以往的研究一般认为生物多样性能够促进生产力等生态系统功能, 目前已经提出的两种主要假说为: 质量比假说和互补假说.其中, 质量比假说认为生态系统功能主要由优势种的组成及其功能属性决定(Grime, 1998); 互补假说认为群落生产力的增加是由于物种间的相互促进作用和生态位分化导致竞争减少, 使物种获得的资源更多, 从而增加生态系统的生产力(Tilman et al., 1997; Forrester, 2014).此外, 有关生态系统功能的影响机制, 还有其他一些假说.如有研究认为森林生态系统功能主要由植被生物量所驱动(Lohbeck et al., 2015), 即植被数量假说. ...
... 林分因子在早期BEF研究中未得到足够重视, 但近年越来越多的研究认为, 林分因子对生物量、生产力的作用可能大于多样性(Fotis et al., 2018; Ali et al., 2019; Ouyang et al., 2019).本研究的结果也表明, 林分因子对凋落物产量及其稳定性均有重要影响(图2).其中, 最大树高对凋落物产量影响最大, 这与近期研究发现大树对林分生产力有决定性影响的结果一致(Ali et al., 2019; Xu et al., 2019).大树往往在群落中更具竞争力, 它们可以达到较高的树冠高度并具有更高的光利用率(King et al., 2006), 因此大树可以显著提高森林的生产力(Xu et al., 2019).森林生产力是决定凋落量多少的关键因素, 生产力较低的林分凋落量自然较低.最大树高、胸高断面积都和林分生物量有密切的关系, 常被用作生物量的替代指标(Wang et al., 2008).因此, 本研究中二者对凋落物产量的重要影响(图2)也支持植被数量假说, 即生态系统过程的速率(如生产力、凋落物产量)主要决定于林分生物量(Lohbeck et al., 2015).此外, 林冠空隙度较低的群落可能具有更复杂的多层冠层结构, 不仅可更充分利用垂直生态位, 还为光合作用提供了更广泛的可吸收光谱(Atkins et al., 2018), 因此可更有效地利用光能来提高生产力. ...
Biodiversity and ecosystem stability: a synthesis of underlying mechanisms 1 2013
... 不少研究发现生物多样性可提高生态系统稳定性(Caldeira et al., 2005), 但也有研究认为二者没有显著相关关系(Zhang & Zhang, 2006).本研究发现虽然物种丰富度与凋落物稳定性的相关性较高(表2), 但在考虑了其他生物和非生物因子共同作用时(图2), 功能多样性更能体现生物多样性对稳定性的作用(Mazzochini et al., 2019).相对于物种和系统发育多样性, 功能多样性可以更好地反应生态系统的功能冗余度和生态位重叠(Tilman et al., 1997).当一个生态系统中有多个具有相似生态位的物种时, 可能会产生功能冗余, 但是当环境变化而使某个物种丧失或衰退时, 这种功能冗余会使其生态位被另一个物种所补偿, 从而起到增强生态系统生产力稳定性的作用(Cleland, 2011), 即功能多样性可以通过物种间生活史策略的互补提高群落的稳定性(Mori et al., 2013).物种和系统发育多样性也进入了凋落物产量稳定性的模型, 不过影响较弱(图2).物种丰富度的解释力可能反映了物种间生长发育的不同步性对生态系统稳定性的影响(Loreau & de Mazancourt, 2013; Morin et al., 2014), 系统发育多样性的解释力则可能体现了物种间进化上的差异导致的资源获取、储存和利用等策略的不同(Souza et al., 2016), 从而提高了生态系统稳定性.综上, 本研究的结果表明, 物种、功能和系统发育多样性共同影响凋落物产量稳定性, 其中功能多样性可能起着最主要的作用.此外, 变异分离分析表明, 多样性还可通过和林分因子、性状的协同作用影响稳定性, 也支持多样性较高的演替后期林分具有较高的稳定性. ...
Control of transpiration in a 220- year-old 1 2001
... 以往关于生态系统稳定性的研究多探讨其与多样性的关系(Cleland, 2011; Mori et al., 2013), 而很少考虑林分因子.本研究的结果表明, 最大胸径、TBA等林分因子对凋落物产量稳定性的影响仅次于功能多样性(图2), 并通过与多样性、性状的协同作用影响稳定性(图3).大直径的树木通常具有更高的树高和更大的冠幅, 能够调节林分水平叶面积、可利用水资源和小气候(Martin et al., 2001).Needham等(2016)的研究表明, 大树较多的森林具有较高的结构异质性, 这样的森林在应对环境波动时能更好地维持生态系统功能.Xu等(2019)对东北地区纬度梯度上不同类型森林的研究表明, 大树占据了绝大多数林分生物量和生产力, 因此林分生产力的稳定性主要决定于大树.这些结果都说明, 除了多样性之外, 林分因子(尤其是大树), 也是影响森林生产力稳定的重要因素. ...
Plant phylogenetic diversity stabilizes large scale ecosystem productivity 3 2019
... 稳定性表征群落抵抗干扰、维持结构与功能保持原状的能力, 是衡量生态系统功能和生态系统健康程度的一个重要方面(Mazzochini et al., 2019).保险假说是多样性可促进生态系统稳定性的主要假说之一, 该假说认为由于物种对环境变化的异步响应, 使得不同物种在时间生态位上也可实现功能互补, 从而增强生态系统在环境波动、干扰下的稳定性(Morin et al., 2014; Mazzochini et al., 2019).此外, 多样性还可通过空间上的生态位互补提高生产力, 也可降低生态系统功能在环境变化时的波动性, 从而提高生态系统的稳定性(Naeem & Li, 1997; Mori et al., 2013). ...
... ; Mazzochini et al., 2019).此外, 多样性还可通过空间上的生态位互补提高生产力, 也可降低生态系统功能在环境变化时的波动性, 从而提高生态系统的稳定性(Naeem & Li, 1997; Mori et al., 2013). ...
... 不少研究发现生物多样性可提高生态系统稳定性(Caldeira et al., 2005), 但也有研究认为二者没有显著相关关系(Zhang & Zhang, 2006).本研究发现虽然物种丰富度与凋落物稳定性的相关性较高(表2), 但在考虑了其他生物和非生物因子共同作用时(图2), 功能多样性更能体现生物多样性对稳定性的作用(Mazzochini et al., 2019).相对于物种和系统发育多样性, 功能多样性可以更好地反应生态系统的功能冗余度和生态位重叠(Tilman et al., 1997).当一个生态系统中有多个具有相似生态位的物种时, 可能会产生功能冗余, 但是当环境变化而使某个物种丧失或衰退时, 这种功能冗余会使其生态位被另一个物种所补偿, 从而起到增强生态系统生产力稳定性的作用(Cleland, 2011), 即功能多样性可以通过物种间生活史策略的互补提高群落的稳定性(Mori et al., 2013).物种和系统发育多样性也进入了凋落物产量稳定性的模型, 不过影响较弱(图2).物种丰富度的解释力可能反映了物种间生长发育的不同步性对生态系统稳定性的影响(Loreau & de Mazancourt, 2013; Morin et al., 2014), 系统发育多样性的解释力则可能体现了物种间进化上的差异导致的资源获取、储存和利用等策略的不同(Souza et al., 2016), 从而提高了生态系统稳定性.综上, 本研究的结果表明, 物种、功能和系统发育多样性共同影响凋落物产量稳定性, 其中功能多样性可能起着最主要的作用.此外, 变异分离分析表明, 多样性还可通过和林分因子、性状的协同作用影响稳定性, 也支持多样性较高的演替后期林分具有较高的稳定性. ...
Response diversity determines the resilience of ecosystems to environmental change 4 2013
... 稳定性表征群落抵抗干扰、维持结构与功能保持原状的能力, 是衡量生态系统功能和生态系统健康程度的一个重要方面(Mazzochini et al., 2019).保险假说是多样性可促进生态系统稳定性的主要假说之一, 该假说认为由于物种对环境变化的异步响应, 使得不同物种在时间生态位上也可实现功能互补, 从而增强生态系统在环境波动、干扰下的稳定性(Morin et al., 2014; Mazzochini et al., 2019).此外, 多样性还可通过空间上的生态位互补提高生产力, 也可降低生态系统功能在环境变化时的波动性, 从而提高生态系统的稳定性(Naeem & Li, 1997; Mori et al., 2013). ...
... 多年来的BEF研究表明, 除了多样性, 森林生产力还受到林分因子、群落物种组成(功能性状)、环境梯度等因素的强烈影响(Wu et al., 2015; Fotis et al., 2018; Ali et al., 2019).随着演替进展, 森林群落的多样性、林分因子、功能性状等都会发生显著的变化(Letcher & Chazdon, 2009), 而这些因素都可能影响生产力和凋落物产量, 因此森林演替系列是BEF研究的一个理想对象(Huang et al., 2017).如在林分因子中, 有研究发现较高的胸高断面积可以显著增加凋落物产量(Guo et al., 2019); 树冠的多层结构通过提高群落的光利用效率来增加凋落物产量(Zheng et al., 2019).功能性状对凋落物产量也有显著影响, 如Niinemets等(2002)发现较高的比叶面积和叶片N含量可以通过光合作用增加植物对周围环境中资源的获取能力和利用效率, 从而提高凋落物产量.关于多样性、林分因子、功能性状对生态系统功能的相对影响大小, 近期研究发现林分因子(Fotis et al., 2018; Ouyang et al., 2019)和功能性状(Garnier et al., 2004)对生物量、生产力影响明显大于多样性.对于凋落物产量是否也是如此, 目前还缺乏相关研究.关于生态系统的稳定性, 以往研究多集中于探讨多样性对其影响机制(Cleland, 2011; Mori et al., 2013).但近年来的研究表明, 森林生长在气候变化下的稳定性, 不仅受到气候变化、环境条件的影响, 林分因子(树高、密度、胸高断面积等)和物种组成也有着重要作用(温晓示等, 2019).这说明林分因子和功能性状同样可能影响森林生产力的稳定性, 但二者和生物多样性的相对重要性还少有研究, 但这对于如何进行森林经营以维持生态系统在未来气候变化下的稳定有着重要意义.本研究的另一个主要目标, 是利用凋落物产量的年际稳定性来探讨这一问题. ...
... 不少研究发现生物多样性可提高生态系统稳定性(Caldeira et al., 2005), 但也有研究认为二者没有显著相关关系(Zhang & Zhang, 2006).本研究发现虽然物种丰富度与凋落物稳定性的相关性较高(表2), 但在考虑了其他生物和非生物因子共同作用时(图2), 功能多样性更能体现生物多样性对稳定性的作用(Mazzochini et al., 2019).相对于物种和系统发育多样性, 功能多样性可以更好地反应生态系统的功能冗余度和生态位重叠(Tilman et al., 1997).当一个生态系统中有多个具有相似生态位的物种时, 可能会产生功能冗余, 但是当环境变化而使某个物种丧失或衰退时, 这种功能冗余会使其生态位被另一个物种所补偿, 从而起到增强生态系统生产力稳定性的作用(Cleland, 2011), 即功能多样性可以通过物种间生活史策略的互补提高群落的稳定性(Mori et al., 2013).物种和系统发育多样性也进入了凋落物产量稳定性的模型, 不过影响较弱(图2).物种丰富度的解释力可能反映了物种间生长发育的不同步性对生态系统稳定性的影响(Loreau & de Mazancourt, 2013; Morin et al., 2014), 系统发育多样性的解释力则可能体现了物种间进化上的差异导致的资源获取、储存和利用等策略的不同(Souza et al., 2016), 从而提高了生态系统稳定性.综上, 本研究的结果表明, 物种、功能和系统发育多样性共同影响凋落物产量稳定性, 其中功能多样性可能起着最主要的作用.此外, 变异分离分析表明, 多样性还可通过和林分因子、性状的协同作用影响稳定性, 也支持多样性较高的演替后期林分具有较高的稳定性. ...
... 以往关于生态系统稳定性的研究多探讨其与多样性的关系(Cleland, 2011; Mori et al., 2013), 而很少考虑林分因子.本研究的结果表明, 最大胸径、TBA等林分因子对凋落物产量稳定性的影响仅次于功能多样性(图2), 并通过与多样性、性状的协同作用影响稳定性(图3).大直径的树木通常具有更高的树高和更大的冠幅, 能够调节林分水平叶面积、可利用水资源和小气候(Martin et al., 2001).Needham等(2016)的研究表明, 大树较多的森林具有较高的结构异质性, 这样的森林在应对环境波动时能更好地维持生态系统功能.Xu等(2019)对东北地区纬度梯度上不同类型森林的研究表明, 大树占据了绝大多数林分生物量和生产力, 因此林分生产力的稳定性主要决定于大树.这些结果都说明, 除了多样性之外, 林分因子(尤其是大树), 也是影响森林生产力稳定的重要因素. ...
Temporal stability in forest productivity increases with tree diversity due to asynchrony in species dynamics 2 2014
... 稳定性表征群落抵抗干扰、维持结构与功能保持原状的能力, 是衡量生态系统功能和生态系统健康程度的一个重要方面(Mazzochini et al., 2019).保险假说是多样性可促进生态系统稳定性的主要假说之一, 该假说认为由于物种对环境变化的异步响应, 使得不同物种在时间生态位上也可实现功能互补, 从而增强生态系统在环境波动、干扰下的稳定性(Morin et al., 2014; Mazzochini et al., 2019).此外, 多样性还可通过空间上的生态位互补提高生产力, 也可降低生态系统功能在环境变化时的波动性, 从而提高生态系统的稳定性(Naeem & Li, 1997; Mori et al., 2013). ...
... 不少研究发现生物多样性可提高生态系统稳定性(Caldeira et al., 2005), 但也有研究认为二者没有显著相关关系(Zhang & Zhang, 2006).本研究发现虽然物种丰富度与凋落物稳定性的相关性较高(表2), 但在考虑了其他生物和非生物因子共同作用时(图2), 功能多样性更能体现生物多样性对稳定性的作用(Mazzochini et al., 2019).相对于物种和系统发育多样性, 功能多样性可以更好地反应生态系统的功能冗余度和生态位重叠(Tilman et al., 1997).当一个生态系统中有多个具有相似生态位的物种时, 可能会产生功能冗余, 但是当环境变化而使某个物种丧失或衰退时, 这种功能冗余会使其生态位被另一个物种所补偿, 从而起到增强生态系统生产力稳定性的作用(Cleland, 2011), 即功能多样性可以通过物种间生活史策略的互补提高群落的稳定性(Mori et al., 2013).物种和系统发育多样性也进入了凋落物产量稳定性的模型, 不过影响较弱(图2).物种丰富度的解释力可能反映了物种间生长发育的不同步性对生态系统稳定性的影响(Loreau & de Mazancourt, 2013; Morin et al., 2014), 系统发育多样性的解释力则可能体现了物种间进化上的差异导致的资源获取、储存和利用等策略的不同(Souza et al., 2016), 从而提高了生态系统稳定性.综上, 本研究的结果表明, 物种、功能和系统发育多样性共同影响凋落物产量稳定性, 其中功能多样性可能起着最主要的作用.此外, 变异分离分析表明, 多样性还可通过和林分因子、性状的协同作用影响稳定性, 也支持多样性较高的演替后期林分具有较高的稳定性. ...
... 稳定性表征群落抵抗干扰、维持结构与功能保持原状的能力, 是衡量生态系统功能和生态系统健康程度的一个重要方面(Mazzochini et al., 2019).保险假说是多样性可促进生态系统稳定性的主要假说之一, 该假说认为由于物种对环境变化的异步响应, 使得不同物种在时间生态位上也可实现功能互补, 从而增强生态系统在环境波动、干扰下的稳定性(Morin et al., 2014; Mazzochini et al., 2019).此外, 多样性还可通过空间上的生态位互补提高生产力, 也可降低生态系统功能在环境变化时的波动性, 从而提高生态系统的稳定性(Naeem & Li, 1997; Mori et al., 2013). ...
Forest community response to invasive pathogens: the case of ash dieback in a British woodland 1 2016
... 以往关于生态系统稳定性的研究多探讨其与多样性的关系(Cleland, 2011; Mori et al., 2013), 而很少考虑林分因子.本研究的结果表明, 最大胸径、TBA等林分因子对凋落物产量稳定性的影响仅次于功能多样性(图2), 并通过与多样性、性状的协同作用影响稳定性(图3).大直径的树木通常具有更高的树高和更大的冠幅, 能够调节林分水平叶面积、可利用水资源和小气候(Martin et al., 2001).Needham等(2016)的研究表明, 大树较多的森林具有较高的结构异质性, 这样的森林在应对环境波动时能更好地维持生态系统功能.Xu等(2019)对东北地区纬度梯度上不同类型森林的研究表明, 大树占据了绝大多数林分生物量和生产力, 因此林分生产力的稳定性主要决定于大树.这些结果都说明, 除了多样性之外, 林分因子(尤其是大树), 也是影响森林生产力稳定的重要因素. ...
Monoterpene emissions in relation to foliar photosynthetic and structural variables in Mediterranean evergreen Quercus species 2 2002
... 多年来的BEF研究表明, 除了多样性, 森林生产力还受到林分因子、群落物种组成(功能性状)、环境梯度等因素的强烈影响(Wu et al., 2015; Fotis et al., 2018; Ali et al., 2019).随着演替进展, 森林群落的多样性、林分因子、功能性状等都会发生显著的变化(Letcher & Chazdon, 2009), 而这些因素都可能影响生产力和凋落物产量, 因此森林演替系列是BEF研究的一个理想对象(Huang et al., 2017).如在林分因子中, 有研究发现较高的胸高断面积可以显著增加凋落物产量(Guo et al., 2019); 树冠的多层结构通过提高群落的光利用效率来增加凋落物产量(Zheng et al., 2019).功能性状对凋落物产量也有显著影响, 如Niinemets等(2002)发现较高的比叶面积和叶片N含量可以通过光合作用增加植物对周围环境中资源的获取能力和利用效率, 从而提高凋落物产量.关于多样性、林分因子、功能性状对生态系统功能的相对影响大小, 近期研究发现林分因子(Fotis et al., 2018; Ouyang et al., 2019)和功能性状(Garnier et al., 2004)对生物量、生产力影响明显大于多样性.对于凋落物产量是否也是如此, 目前还缺乏相关研究.关于生态系统的稳定性, 以往研究多集中于探讨多样性对其影响机制(Cleland, 2011; Mori et al., 2013).但近年来的研究表明, 森林生长在气候变化下的稳定性, 不仅受到气候变化、环境条件的影响, 林分因子(树高、密度、胸高断面积等)和物种组成也有着重要作用(温晓示等, 2019).这说明林分因子和功能性状同样可能影响森林生产力的稳定性, 但二者和生物多样性的相对重要性还少有研究, 但这对于如何进行森林经营以维持生态系统在未来气候变化下的稳定有着重要意义.本研究的另一个主要目标, 是利用凋落物产量的年际稳定性来探讨这一问题. ...
... 本研究发现叶片碳含量对凋落物产量的影响较为明显(图2), 碳是构成植物叶片干物质含量的主要元素, 当植物将较多的碳用于叶片维持和防御结构上的投资时会相应减少在光合上的投资, 使植物生长变慢(He et al., 2006).本研究中叶片碳含量与凋落物产量的负相关性也支持这一点.以往研究多认为SLA、LN对生产力有显著影响(Niinemets et al., 2002), 但本研究中, 二者对凋落物产量的解释力明显低于LC.这可能说明影响凋落物产量和生长的性状存在一定差异, 因为凋落物产量只是林分生产力的一部分, 还涉及到功能性状如何影响资源分配等问题.比如, 在树叶凋落之前植物会通过将部分养分从叶中转运回体内储存, 来减弱植物对环境养分供应的依赖(Aerts & Chapin III, 1999).功能性状如何影响类似这样的资源分配过程, 还有待进一步研究. ...
Effects of stand age, richness and density on productivity in subtropical forests in China 3 2019
... 多年来的BEF研究表明, 除了多样性, 森林生产力还受到林分因子、群落物种组成(功能性状)、环境梯度等因素的强烈影响(Wu et al., 2015; Fotis et al., 2018; Ali et al., 2019).随着演替进展, 森林群落的多样性、林分因子、功能性状等都会发生显著的变化(Letcher & Chazdon, 2009), 而这些因素都可能影响生产力和凋落物产量, 因此森林演替系列是BEF研究的一个理想对象(Huang et al., 2017).如在林分因子中, 有研究发现较高的胸高断面积可以显著增加凋落物产量(Guo et al., 2019); 树冠的多层结构通过提高群落的光利用效率来增加凋落物产量(Zheng et al., 2019).功能性状对凋落物产量也有显著影响, 如Niinemets等(2002)发现较高的比叶面积和叶片N含量可以通过光合作用增加植物对周围环境中资源的获取能力和利用效率, 从而提高凋落物产量.关于多样性、林分因子、功能性状对生态系统功能的相对影响大小, 近期研究发现林分因子(Fotis et al., 2018; Ouyang et al., 2019)和功能性状(Garnier et al., 2004)对生物量、生产力影响明显大于多样性.对于凋落物产量是否也是如此, 目前还缺乏相关研究.关于生态系统的稳定性, 以往研究多集中于探讨多样性对其影响机制(Cleland, 2011; Mori et al., 2013).但近年来的研究表明, 森林生长在气候变化下的稳定性, 不仅受到气候变化、环境条件的影响, 林分因子(树高、密度、胸高断面积等)和物种组成也有着重要作用(温晓示等, 2019).这说明林分因子和功能性状同样可能影响森林生产力的稳定性, 但二者和生物多样性的相对重要性还少有研究, 但这对于如何进行森林经营以维持生态系统在未来气候变化下的稳定有着重要意义.本研究的另一个主要目标, 是利用凋落物产量的年际稳定性来探讨这一问题. ...
... 多年来的研究一般认为, 物种丰富的植物群落可支持较高的生物量和生产力, 并使群落的组成和功能在长期的动态变化中维持较高的稳定性.尽管多数研究认为多样性与生产力之间为正相关关系(Cavanaugh et al., 2014; Liang et al., 2016), 但也有部分研究结果发现两者之间为负相关关系或不相关(Finegan et al., 2015).在本研究中3个生物多样性指标与凋落物产量均有显著的正相关关系(表2), 其中物种多样性增加了物种数量和物种多度分布的均匀程度; 功能多样性使不同物种能够更有效地利用功能生态位、从而获取更多的资源(Harpole & Tilman, 2007); 而系统发育多样性增加了物种间进化关系和功能上的差异(Cadotte et al., 2009).多样性高的群落可以通过更有效地利用光照、养分和水等资源来提高生态系统生产力, 从而提高凋落物产量.不过, 通过消除协同变化的因子的影响, 本研究发现多样性在林分因子、功能性状之外对凋落物产量的独立作用很微弱(图3), 这说明多样性主要是通过与其他因素的共同作用来影响凋落物产量(Wu et al., 2015; Ouyang et al., 2019). ...
... 林分因子在早期BEF研究中未得到足够重视, 但近年越来越多的研究认为, 林分因子对生物量、生产力的作用可能大于多样性(Fotis et al., 2018; Ali et al., 2019; Ouyang et al., 2019).本研究的结果也表明, 林分因子对凋落物产量及其稳定性均有重要影响(图2).其中, 最大树高对凋落物产量影响最大, 这与近期研究发现大树对林分生产力有决定性影响的结果一致(Ali et al., 2019; Xu et al., 2019).大树往往在群落中更具竞争力, 它们可以达到较高的树冠高度并具有更高的光利用率(King et al., 2006), 因此大树可以显著提高森林的生产力(Xu et al., 2019).森林生产力是决定凋落量多少的关键因素, 生产力较低的林分凋落量自然较低.最大树高、胸高断面积都和林分生物量有密切的关系, 常被用作生物量的替代指标(Wang et al., 2008).因此, 本研究中二者对凋落物产量的重要影响(图2)也支持植被数量假说, 即生态系统过程的速率(如生产力、凋落物产量)主要决定于林分生物量(Lohbeck et al., 2015).此外, 林冠空隙度较低的群落可能具有更复杂的多层冠层结构, 不仅可更充分利用垂直生态位, 还为光合作用提供了更广泛的可吸收光谱(Atkins et al., 2018), 因此可更有效地利用光能来提高生产力. ...
Sensitivity of leaf size and shape to climate: global patterns and paleoclimatic applications 1 2011
... 本研究结果表明凋落物产量稳定性与SLA显著负相关(表2).叶片的功能性状对气候比较敏感, 反映了植物对环境的高度适应能力和在复杂生境下的自我调控能力(Scoffoni et al., 2011), 叶片大小随着年平均气温及年降水量的降低而减小(Peppe et al., 2011).叶面积可显著影响植物的生长速度(Souza et al., 2016), 具有较低SLA的物种的生长策略为保守型, 其表现为生长速度较慢, 但更能适应环境变化且具有更高的稳定性.比如, 低SLA的物种增加了在木质素等结构物质上的投资(Poorter & De Jong, 1999), 并产生具有较小细胞和较厚细胞壁的叶片, 从而降低细胞弹性并减少叶片水势(Klein, 2014), 使它们在干旱时期蒸腾的水分更少(Sterck et al., 2011), 从而在外部环境变化时更好地保持水分平衡, 进而影响凋落物产量的稳定性.此外, 本研究结果表明, 群落水平的功能性状对凋落物产量及其稳定性都有重要影响(图2, 图3), 这也支持质量比假说(Grime, 1998), 说明除了多样性和林分因子, 优势种的功能性状对群落生产力及生态系统的稳定性也有重要影响. ...
New handbook for standardised measurement of plant functional traits worldwide 1 2013
... 为了研究群落水平功能性状对凋落物产量及其稳定性的影响, 本研究选取了各个样地的优势树种对其功能性状进行了测定, 所选优势种的胸高断面积之和占各样地总胸高断面积的85%以上.测定的功能性状包括: 比叶面积(SLA)、叶片碳含量(LC)和叶片氮含量(LN).在样地附近, 对每个优势树种随机选取健康成熟、长势均匀的5株平均木, 在树冠外侧的阳面, 剪下10片健康无损、暴露在日光下的叶片.采下的新鲜叶片当天用CanoScan LiDE 100扫描仪扫描叶片, 利用ImageJ软件计算全版叶面积, 之后将叶片在烘箱中70 ℃烘干至恒质量, 测定叶片干质量, 用叶面积除以叶干质量获得比叶面积.采用C/N元素分析仪(2400 II CHNS/O Element Analyzer, Perkin-Elmer, Boston, USA)测定样品全C和全N含量.所有性状的取样、测定步骤都按照国际通用技术规范执行(Pérez-Harguindeguy et al., 2013). ...
Variations in satellite-derived phenology in China’s temperate vegetation 1 2006
Responses of soil respiration to elevated CO2, air warming, and changing soil water availability in a model old-field grassland 1 2007
... 近半个世纪以来快速的全球气候变暖直接或间接导致了生物多样性的锐减, 改变了生态系统的结构, 严重影响了生态系统的服务功能(Tilman, 2000; Wan et al., 2007).由于环境变迁和人类活动造成的生物多样性丧失对全球生态系统带来的影响, 使得生物多样性-生态系统功能(BEF)关系一直备受关注(Lasky et al., 2014; Zhang & Chen, 2015).以往的研究一般认为生物多样性能够促进生产力等生态系统功能, 目前已经提出的两种主要假说为: 质量比假说和互补假说.其中, 质量比假说认为生态系统功能主要由优势种的组成及其功能属性决定(Grime, 1998); 互补假说认为群落生产力的增加是由于物种间的相互促进作用和生态位分化导致竞争减少, 使物种获得的资源更多, 从而增加生态系统的生产力(Tilman et al., 1997; Forrester, 2014).此外, 有关生态系统功能的影响机制, 还有其他一些假说.如有研究认为森林生态系统功能主要由植被生物量所驱动(Lohbeck et al., 2015), 即植被数量假说. ...
Forest biomass and root-shoot allocation in northeast China 2 2008
... 为研究林分因子对凋落物产量及其稳定性的影响, 在以往研究基础上, 本研究选择了4个变量, 包括: (1)最大树高(Hmax); (2)最大胸径(DBHmax); (3)胸高断面积(TBA); (4)林冠空隙度(gap fraction).近期研究表明群落中的大树对林分生物量、生产力及其波动都有重要影响, 因此本研究选择最大树高和最大胸径以反映大树的作用(Ali et al., 2019; Xu et al., 2019).胸高断面积与林分生物量高度相关(Wang et al., 2008), 也可以反映林分内的竞争强度(Zhang & Chen, 2015).由于林分密度、平均树高和平均胸径与上述几个林分因子共线性较强, 解释力也较低, 所以在最终的多元模型中没有选择这几个因子. ...
... 林分因子在早期BEF研究中未得到足够重视, 但近年越来越多的研究认为, 林分因子对生物量、生产力的作用可能大于多样性(Fotis et al., 2018; Ali et al., 2019; Ouyang et al., 2019).本研究的结果也表明, 林分因子对凋落物产量及其稳定性均有重要影响(图2).其中, 最大树高对凋落物产量影响最大, 这与近期研究发现大树对林分生产力有决定性影响的结果一致(Ali et al., 2019; Xu et al., 2019).大树往往在群落中更具竞争力, 它们可以达到较高的树冠高度并具有更高的光利用率(King et al., 2006), 因此大树可以显著提高森林的生产力(Xu et al., 2019).森林生产力是决定凋落量多少的关键因素, 生产力较低的林分凋落量自然较低.最大树高、胸高断面积都和林分生物量有密切的关系, 常被用作生物量的替代指标(Wang et al., 2008).因此, 本研究中二者对凋落物产量的重要影响(图2)也支持植被数量假说, 即生态系统过程的速率(如生产力、凋落物产量)主要决定于林分生物量(Lohbeck et al., 2015).此外, 林冠空隙度较低的群落可能具有更复杂的多层冠层结构, 不仅可更充分利用垂直生态位, 还为光合作用提供了更广泛的可吸收光谱(Atkins et al., 2018), 因此可更有效地利用光能来提高生产力. ...
不同林龄、树种落叶松人工林径向生长与气候变化的关系 1 2019
... 多年来的BEF研究表明, 除了多样性, 森林生产力还受到林分因子、群落物种组成(功能性状)、环境梯度等因素的强烈影响(Wu et al., 2015; Fotis et al., 2018; Ali et al., 2019).随着演替进展, 森林群落的多样性、林分因子、功能性状等都会发生显著的变化(Letcher & Chazdon, 2009), 而这些因素都可能影响生产力和凋落物产量, 因此森林演替系列是BEF研究的一个理想对象(Huang et al., 2017).如在林分因子中, 有研究发现较高的胸高断面积可以显著增加凋落物产量(Guo et al., 2019); 树冠的多层结构通过提高群落的光利用效率来增加凋落物产量(Zheng et al., 2019).功能性状对凋落物产量也有显著影响, 如Niinemets等(2002)发现较高的比叶面积和叶片N含量可以通过光合作用增加植物对周围环境中资源的获取能力和利用效率, 从而提高凋落物产量.关于多样性、林分因子、功能性状对生态系统功能的相对影响大小, 近期研究发现林分因子(Fotis et al., 2018; Ouyang et al., 2019)和功能性状(Garnier et al., 2004)对生物量、生产力影响明显大于多样性.对于凋落物产量是否也是如此, 目前还缺乏相关研究.关于生态系统的稳定性, 以往研究多集中于探讨多样性对其影响机制(Cleland, 2011; Mori et al., 2013).但近年来的研究表明, 森林生长在气候变化下的稳定性, 不仅受到气候变化、环境条件的影响, 林分因子(树高、密度、胸高断面积等)和物种组成也有着重要作用(温晓示等, 2019).这说明林分因子和功能性状同样可能影响森林生产力的稳定性, 但二者和生物多样性的相对重要性还少有研究, 但这对于如何进行森林经营以维持生态系统在未来气候变化下的稳定有着重要意义.本研究的另一个主要目标, 是利用凋落物产量的年际稳定性来探讨这一问题. ...
不同林龄、树种落叶松人工林径向生长与气候变化的关系 1 2019
... 多年来的BEF研究表明, 除了多样性, 森林生产力还受到林分因子、群落物种组成(功能性状)、环境梯度等因素的强烈影响(Wu et al., 2015; Fotis et al., 2018; Ali et al., 2019).随着演替进展, 森林群落的多样性、林分因子、功能性状等都会发生显著的变化(Letcher & Chazdon, 2009), 而这些因素都可能影响生产力和凋落物产量, 因此森林演替系列是BEF研究的一个理想对象(Huang et al., 2017).如在林分因子中, 有研究发现较高的胸高断面积可以显著增加凋落物产量(Guo et al., 2019); 树冠的多层结构通过提高群落的光利用效率来增加凋落物产量(Zheng et al., 2019).功能性状对凋落物产量也有显著影响, 如Niinemets等(2002)发现较高的比叶面积和叶片N含量可以通过光合作用增加植物对周围环境中资源的获取能力和利用效率, 从而提高凋落物产量.关于多样性、林分因子、功能性状对生态系统功能的相对影响大小, 近期研究发现林分因子(Fotis et al., 2018; Ouyang et al., 2019)和功能性状(Garnier et al., 2004)对生物量、生产力影响明显大于多样性.对于凋落物产量是否也是如此, 目前还缺乏相关研究.关于生态系统的稳定性, 以往研究多集中于探讨多样性对其影响机制(Cleland, 2011; Mori et al., 2013).但近年来的研究表明, 森林生长在气候变化下的稳定性, 不仅受到气候变化、环境条件的影响, 林分因子(树高、密度、胸高断面积等)和物种组成也有着重要作用(温晓示等, 2019).这说明林分因子和功能性状同样可能影响森林生产力的稳定性, 但二者和生物多样性的相对重要性还少有研究, 但这对于如何进行森林经营以维持生态系统在未来气候变化下的稳定有着重要意义.本研究的另一个主要目标, 是利用凋落物产量的年际稳定性来探讨这一问题. ...
The relationship between species richness and biomass changes from boreal to subtropical forests in China 2 2015
... 多年来的BEF研究表明, 除了多样性, 森林生产力还受到林分因子、群落物种组成(功能性状)、环境梯度等因素的强烈影响(Wu et al., 2015; Fotis et al., 2018; Ali et al., 2019).随着演替进展, 森林群落的多样性、林分因子、功能性状等都会发生显著的变化(Letcher & Chazdon, 2009), 而这些因素都可能影响生产力和凋落物产量, 因此森林演替系列是BEF研究的一个理想对象(Huang et al., 2017).如在林分因子中, 有研究发现较高的胸高断面积可以显著增加凋落物产量(Guo et al., 2019); 树冠的多层结构通过提高群落的光利用效率来增加凋落物产量(Zheng et al., 2019).功能性状对凋落物产量也有显著影响, 如Niinemets等(2002)发现较高的比叶面积和叶片N含量可以通过光合作用增加植物对周围环境中资源的获取能力和利用效率, 从而提高凋落物产量.关于多样性、林分因子、功能性状对生态系统功能的相对影响大小, 近期研究发现林分因子(Fotis et al., 2018; Ouyang et al., 2019)和功能性状(Garnier et al., 2004)对生物量、生产力影响明显大于多样性.对于凋落物产量是否也是如此, 目前还缺乏相关研究.关于生态系统的稳定性, 以往研究多集中于探讨多样性对其影响机制(Cleland, 2011; Mori et al., 2013).但近年来的研究表明, 森林生长在气候变化下的稳定性, 不仅受到气候变化、环境条件的影响, 林分因子(树高、密度、胸高断面积等)和物种组成也有着重要作用(温晓示等, 2019).这说明林分因子和功能性状同样可能影响森林生产力的稳定性, 但二者和生物多样性的相对重要性还少有研究, 但这对于如何进行森林经营以维持生态系统在未来气候变化下的稳定有着重要意义.本研究的另一个主要目标, 是利用凋落物产量的年际稳定性来探讨这一问题. ...
... 多年来的研究一般认为, 物种丰富的植物群落可支持较高的生物量和生产力, 并使群落的组成和功能在长期的动态变化中维持较高的稳定性.尽管多数研究认为多样性与生产力之间为正相关关系(Cavanaugh et al., 2014; Liang et al., 2016), 但也有部分研究结果发现两者之间为负相关关系或不相关(Finegan et al., 2015).在本研究中3个生物多样性指标与凋落物产量均有显著的正相关关系(表2), 其中物种多样性增加了物种数量和物种多度分布的均匀程度; 功能多样性使不同物种能够更有效地利用功能生态位、从而获取更多的资源(Harpole & Tilman, 2007); 而系统发育多样性增加了物种间进化关系和功能上的差异(Cadotte et al., 2009).多样性高的群落可以通过更有效地利用光照、养分和水等资源来提高生态系统生产力, 从而提高凋落物产量.不过, 通过消除协同变化的因子的影响, 本研究发现多样性在林分因子、功能性状之外对凋落物产量的独立作用很微弱(图3), 这说明多样性主要是通过与其他因素的共同作用来影响凋落物产量(Wu et al., 2015; Ouyang et al., 2019). ...
A new tree-ring sampling method to estimate forest productivity and its temporal variation accurately in natural forests 4 2019
... 为研究林分因子对凋落物产量及其稳定性的影响, 在以往研究基础上, 本研究选择了4个变量, 包括: (1)最大树高(Hmax); (2)最大胸径(DBHmax); (3)胸高断面积(TBA); (4)林冠空隙度(gap fraction).近期研究表明群落中的大树对林分生物量、生产力及其波动都有重要影响, 因此本研究选择最大树高和最大胸径以反映大树的作用(Ali et al., 2019; Xu et al., 2019).胸高断面积与林分生物量高度相关(Wang et al., 2008), 也可以反映林分内的竞争强度(Zhang & Chen, 2015).由于林分密度、平均树高和平均胸径与上述几个林分因子共线性较强, 解释力也较低, 所以在最终的多元模型中没有选择这几个因子. ...
... 林分因子在早期BEF研究中未得到足够重视, 但近年越来越多的研究认为, 林分因子对生物量、生产力的作用可能大于多样性(Fotis et al., 2018; Ali et al., 2019; Ouyang et al., 2019).本研究的结果也表明, 林分因子对凋落物产量及其稳定性均有重要影响(图2).其中, 最大树高对凋落物产量影响最大, 这与近期研究发现大树对林分生产力有决定性影响的结果一致(Ali et al., 2019; Xu et al., 2019).大树往往在群落中更具竞争力, 它们可以达到较高的树冠高度并具有更高的光利用率(King et al., 2006), 因此大树可以显著提高森林的生产力(Xu et al., 2019).森林生产力是决定凋落量多少的关键因素, 生产力较低的林分凋落量自然较低.最大树高、胸高断面积都和林分生物量有密切的关系, 常被用作生物量的替代指标(Wang et al., 2008).因此, 本研究中二者对凋落物产量的重要影响(图2)也支持植被数量假说, 即生态系统过程的速率(如生产力、凋落物产量)主要决定于林分生物量(Lohbeck et al., 2015).此外, 林冠空隙度较低的群落可能具有更复杂的多层冠层结构, 不仅可更充分利用垂直生态位, 还为光合作用提供了更广泛的可吸收光谱(Atkins et al., 2018), 因此可更有效地利用光能来提高生产力. ...
... ), 因此大树可以显著提高森林的生产力(Xu et al., 2019).森林生产力是决定凋落量多少的关键因素, 生产力较低的林分凋落量自然较低.最大树高、胸高断面积都和林分生物量有密切的关系, 常被用作生物量的替代指标(Wang et al., 2008).因此, 本研究中二者对凋落物产量的重要影响(图2)也支持植被数量假说, 即生态系统过程的速率(如生产力、凋落物产量)主要决定于林分生物量(Lohbeck et al., 2015).此外, 林冠空隙度较低的群落可能具有更复杂的多层冠层结构, 不仅可更充分利用垂直生态位, 还为光合作用提供了更广泛的可吸收光谱(Atkins et al., 2018), 因此可更有效地利用光能来提高生产力. ...
... 以往关于生态系统稳定性的研究多探讨其与多样性的关系(Cleland, 2011; Mori et al., 2013), 而很少考虑林分因子.本研究的结果表明, 最大胸径、TBA等林分因子对凋落物产量稳定性的影响仅次于功能多样性(图2), 并通过与多样性、性状的协同作用影响稳定性(图3).大直径的树木通常具有更高的树高和更大的冠幅, 能够调节林分水平叶面积、可利用水资源和小气候(Martin et al., 2001).Needham等(2016)的研究表明, 大树较多的森林具有较高的结构异质性, 这样的森林在应对环境波动时能更好地维持生态系统功能.Xu等(2019)对东北地区纬度梯度上不同类型森林的研究表明, 大树占据了绝大多数林分生物量和生产力, 因此林分生产力的稳定性主要决定于大树.这些结果都说明, 除了多样性之外, 林分因子(尤其是大树), 也是影响森林生产力稳定的重要因素. ...
Three keys to the radiation of angiosperms into freezing environments 1 2014
... 近半个世纪以来快速的全球气候变暖直接或间接导致了生物多样性的锐减, 改变了生态系统的结构, 严重影响了生态系统的服务功能(Tilman, 2000; Wan et al., 2007).由于环境变迁和人类活动造成的生物多样性丧失对全球生态系统带来的影响, 使得生物多样性-生态系统功能(BEF)关系一直备受关注(Lasky et al., 2014; Zhang & Chen, 2015).以往的研究一般认为生物多样性能够促进生产力等生态系统功能, 目前已经提出的两种主要假说为: 质量比假说和互补假说.其中, 质量比假说认为生态系统功能主要由优势种的组成及其功能属性决定(Grime, 1998); 互补假说认为群落生产力的增加是由于物种间的相互促进作用和生态位分化导致竞争减少, 使物种获得的资源更多, 从而增加生态系统的生产力(Tilman et al., 1997; Forrester, 2014).此外, 有关生态系统功能的影响机制, 还有其他一些假说.如有研究认为森林生态系统功能主要由植被生物量所驱动(Lohbeck et al., 2015), 即植被数量假说. ...
... 为研究林分因子对凋落物产量及其稳定性的影响, 在以往研究基础上, 本研究选择了4个变量, 包括: (1)最大树高(Hmax); (2)最大胸径(DBHmax); (3)胸高断面积(TBA); (4)林冠空隙度(gap fraction).近期研究表明群落中的大树对林分生物量、生产力及其波动都有重要影响, 因此本研究选择最大树高和最大胸径以反映大树的作用(Ali et al., 2019; Xu et al., 2019).胸高断面积与林分生物量高度相关(Wang et al., 2008), 也可以反映林分内的竞争强度(Zhang & Chen, 2015).由于林分密度、平均树高和平均胸径与上述几个林分因子共线性较强, 解释力也较低, 所以在最终的多元模型中没有选择这几个因子. ...
Tree species diversity promotes litterfall productivity through crown complementarity in subtropical forests 2 2019
... 凋落物是生态系统净初级生产力的一个重要贡献者(Clark et al., 2001), 也是联结生态系统地上和地下碳循环过程的一个关键环节, 对维持森林土壤肥力以及森林生态系统结构和功能的稳定性具有重要作用(Vogt et al., 1986).以往对凋落物产量已有较多研究, 但多集中于研究凋落物的组分、季节动态, 以及气候、林型等因素如何影响凋落物产量(张新平等, 2008; 贾丙瑞等, 2016).关于多样性对凋落物产量的影响, 则研究相对较少(Guo et al., 2019; Zheng et al., 2019).其中, 多样性如何影响凋落物产量的稳定性的研究还十分匮乏(Huang et al., 2017).近年来不少研究认为, 多样性不但能够促进群落生产力, 还可提高生态系统在环境波动下的稳定性(Cardinale et al., 2012).凋落物作为森林生产力的一个重要组分, 多样性是否对其产量及其稳定性也有显著的促进作用? 这是本文主要研究的一个问题. ...
... 多年来的BEF研究表明, 除了多样性, 森林生产力还受到林分因子、群落物种组成(功能性状)、环境梯度等因素的强烈影响(Wu et al., 2015; Fotis et al., 2018; Ali et al., 2019).随着演替进展, 森林群落的多样性、林分因子、功能性状等都会发生显著的变化(Letcher & Chazdon, 2009), 而这些因素都可能影响生产力和凋落物产量, 因此森林演替系列是BEF研究的一个理想对象(Huang et al., 2017).如在林分因子中, 有研究发现较高的胸高断面积可以显著增加凋落物产量(Guo et al., 2019); 树冠的多层结构通过提高群落的光利用效率来增加凋落物产量(Zheng et al., 2019).功能性状对凋落物产量也有显著影响, 如Niinemets等(2002)发现较高的比叶面积和叶片N含量可以通过光合作用增加植物对周围环境中资源的获取能力和利用效率, 从而提高凋落物产量.关于多样性、林分因子、功能性状对生态系统功能的相对影响大小, 近期研究发现林分因子(Fotis et al., 2018; Ouyang et al., 2019)和功能性状(Garnier et al., 2004)对生物量、生产力影响明显大于多样性.对于凋落物产量是否也是如此, 目前还缺乏相关研究.关于生态系统的稳定性, 以往研究多集中于探讨多样性对其影响机制(Cleland, 2011; Mori et al., 2013).但近年来的研究表明, 森林生长在气候变化下的稳定性, 不仅受到气候变化、环境条件的影响, 林分因子(树高、密度、胸高断面积等)和物种组成也有着重要作用(温晓示等, 2019).这说明林分因子和功能性状同样可能影响森林生产力的稳定性, 但二者和生物多样性的相对重要性还少有研究, 但这对于如何进行森林经营以维持生态系统在未来气候变化下的稳定有着重要意义.本研究的另一个主要目标, 是利用凋落物产量的年际稳定性来探讨这一问题. ...
Litterfall production along successional and altitudinal gradients of subtropical monsoon evergreen broadleaved forests in Guangdong, China 1 2007
... 有研究认为凋落物产量沿演替序列增加(Zhou et al., 2007), 也有****发现凋落物产量在不同演替阶段无显著差异(Huang et al., 2017).一般而言, 在林分发育直至林冠郁闭之前每年的凋落物产量会迅速增加, 然后在相当长的一段时间内保持相对稳定(Albrektson, 1988), 本研究的结果与之一致.胜山自然保护区演替早期的林分密度、树木个体都较小, 林冠未达到郁闭导致其生产力有限, 所以凋落物产量显著低于其他演替阶段, 从演替中期至演替晚期林分高度郁闭(表1), 群落可捕获的光能大致相同, 使得3个演替阶段凋落物产量无显著差异. ...
 ,
, ,*, 张树斌, 吴鹏, 杨蕾北京林业大学生态与自然保护学院, 北京 100083
,*, 张树斌, 吴鹏, 杨蕾北京林业大学生态与自然保护学院, 北京 100083

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT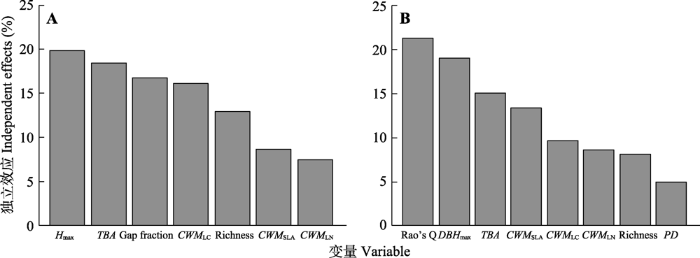 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT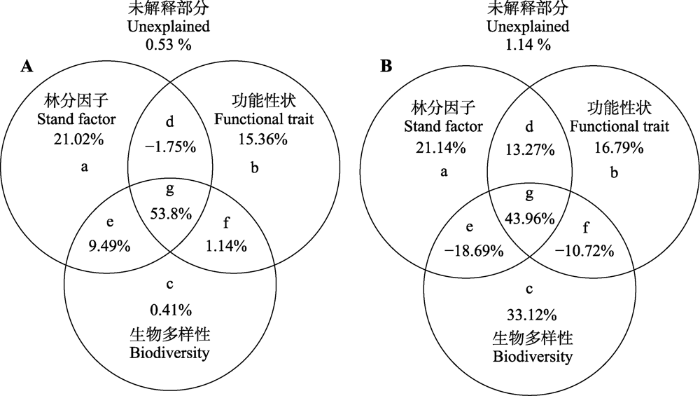 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}