 ,1,*, 马煦,1,*, 邸楠1,2, 王烨3, 刘洋1, 李广德4, 贾黎明1, 席本野1,**
,1,*, 马煦,1,*, 邸楠1,2, 王烨3, 刘洋1, 李广德4, 贾黎明1, 席本野1,**Azimuthal variation in nighttime sap flow and its mainly influence factors of Populus tomentosa
Fei-Fei ZHAO,1,*, Xu MA,1,*, Nan DI1,2, Ye WANG3, Yang LIU1, Guang-De LI4, Li-Ming JIA1, Ben-Ye XI1,**通讯作者: **benyexi@bjfu.edu.cn
编委: 段昌群
责任编辑: 李敏
收稿日期:2020-03-30接受日期:2020-07-10网络出版日期:2020-08-20
| 基金资助: |
Corresponding authors: **benyexi@bjfu.edu.cn
Received:2020-03-30Accepted:2020-07-10Online:2020-08-20
| Fund supported: |

摘要
为明确毛白杨(Populus tomentosa)不同方位夜间蒸腾量(Nt)及茎干充水量(Sr)等夜间液流活动的规律, 探究不同方位Nt和Sr的主要影响因子, 该研究使用热扩散的方法监测了宽窄行模式下栽植的毛白杨茎干不同方位夜间液流, 并用图像法区分Nt和Sr。使用自动气象站和机械式张力计监测太阳总辐射(Rs, kW·m-2)、空气温度(Ta, ℃)、空气相对湿度(RH, %)、风速(v, m·s-1)、土壤水势(ψ, kPa)等环境因子。通过比较各方位的Nt和Sr等液流活动的大小情况及其与环境因子之间的相关性得到方位间夜间液流的差异性以及各方位夜间液流的主要影响因子。结果显示: 宽行距位于东侧的样树西方位的Nt和Sr均最大, 其中西方位的Sr显著大于其他3个方位; 北方位的Nt显著小于其他3个方位; 其他方位间的Nt和Sr无显著差异; 各方位夜间茎干充水量占夜间液流量的比例(Sr/Q)无显著差异。宽行距位于西侧的样树西方位的Nt和Sr亦均最大, 其中西方位的Sr显著大于东方位和南方位; 南方位的Nt最小, 显著小于西方位和北方位, 其他方位间的Nt和Sr无显著差异; 南方位的Sr/Q显著大于其他3个方位。各方位的Nt和Sr均与水汽压亏缺(VPD)有显著的正相关关系, 部分方位Nt和Sr与Ta和RH有显著相关关系, 没有任何方位Nt和Sr与v和ψ有显著相关关系。Nt和Sr方位间的差异(NtCV、SrCV)与VPD、Ta、RH、v和ψ均无显著相关关系。此外, Sr受白天的液流活动的影响显著。综上所述, 毛白杨不同方位Nt和Sr等液流活动具有较大的差异, 且西方位是优势方位; VPD是影响各方位Nt和Sr的主要气象因子。
关键词:
Abstract
Aims To clarify the azimuthal regularity of nocturnal sap-flow activities of Populus tomentosa, which includes nocturnal transpiration (Nt) and stem water refilling (Sr), and explore the main impact factors of Nt and Sr in different orientations.
Methods The thermal dissipation method was used to monitor the nocturnal sap flow of P. tomentosa planted in wide and narrow rows patterns. The image method was used to distinguish Nt and Sr. An automatic weather station measured global solar radiation (Rs, kW·m-2), air temperature (Ta, ℃), relative humidity (RH, %), wind speed (v, m·s-1) and other environmental factors. Mechanical tensiometers measured soil water potential (ψ, kPa). The differences of nocturnal sap-flow among orientations and their main impact factors were determined by comparing the magnitudes of Nt and Sr and their correlations with the impact factors.
Important findings The results showed that, for trees on the east-wide-row, the west orientation has the largest Nt and Sr. The Sr in the west orientation was significantly larger than that in the other three orientations. In contrast, north oriented Nt was significantly smaller than that in the other three orientations. There was no significant difference in Nt and Sr among other orientations and the proportion of Sr accounted for the nighttime sap flow (Sr/Q) in all orientations. For trees on the west-wide-row, Nt and Sr in the west orientation were also the largest, and the Sr in the west orientation was significantly larger than that in the east and south. The Nt in the south orientation was the smallest and significantly smaller than that in the west and north. There was no significant difference in Nt and Sr among other orientations. The Sr/Q in the south orientation was significantly larger than that in the other three orientations. The Nt and Sr had significantly positive correlations with vapor pressure deficiency (VPD), and Nt and Sr in some orientations had significant correlations with Ta and RH, but Nt and Sr in all orientations had no significant correlation with v and ψ. The variation coefficient of Nt and Sr among the four orientations (NtCV and SrCV) had no significant correlation with VPD, Ta, RH, v and ψ. In addition, the Sr was significantly affected by the daytime sap flow. In conclusion, there were significant differences in nocturnal sap flow of P. tomentosa such as Nt and Sr, with west being the most dominant. VPD was the mainly meteorological impact factor of Nt and Sr in all orientations at night.
Keywords:
PDF (1416KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
赵飞飞, 马煦, 邸楠, 王烨, 刘洋, 李广德, 贾黎明, 席本野. 毛白杨茎干不同方位夜间液流变化规律及其主要影响因子. 植物生态学报, 2020, 44(8): 864-874. DOI: 10.17521/cjpe.2020.0089
ZHAO Fei-Fei, MA Xu, DI Nan, WANG Ye, LIU Yang, LI Guang-De, JIA Li-Ming, XI Ben-Ye.
蒸腾是植物消耗水分的主要途径, 在植物生长发育过程中起着至关重要的作用(Tsuruta et al., 2010; Zeppel et al., 2014)。早期的相关研究普遍认为夜间缺少光照、气温和水汽压亏缺较低、叶片气孔关闭, 植物不会发生蒸腾作用(Meidner & Mansfield, 1965; Ritchie et al., 1974; Benyon et al., 1999)。然而随着监测技术的进步, 越来越多的研究表明各种生态系统中的大多数植物都会出现夜间蒸腾的现象(Dawson et al., 2007; Fisher et al., 2007; Alvarado- Barrientos et al., 2013; Zeppel et al., 2014)。夜间蒸腾具有运输营养物质和氧气等生理功能(Marks & Lechowicz, 2007; Zeppel et al., 2014), 并且可以降低叶片表面温度、减少碳损失(Peraudeau et al., 2015)。除了夜间蒸腾外, 茎干充水作为植物夜间液流的另一组成部分, 同样具有重要的生理作用, 如补充植物白天蒸腾引起的水分亏缺(Wang et al., 2012), 提高第二天叶片的光合作用效率(方伟伟等, 2018), 缓解植物木质部栓塞化的产生(Carrasco et al., 2015)。
夜间蒸腾量和茎干充水量的精确估计依赖于夜间液流的精准测定以及夜间蒸腾和茎干充水的有效区分。热技术由于具有精确、不受空间限制、自动化等优点, 被广泛用于树木液流的监测与林分蒸腾的估算(Granier et al., 1996; Wilson et al., 2001; Ford et al., 2007)。在此基础上, Fisher等(2007)通过分析Pinus ponderosa、Quercus douglasii等树种夜间液流曲线的斜率, 提出了一种区分夜间蒸腾和茎干充水的方法, 即通过图像来定量计算出夜间蒸腾量和茎干充水量, 这不仅能够有效区分夜间蒸腾和茎干充水, 而且为研究植物夜间蒸腾和茎干充水的影响因素奠定了基础。
关于植物夜间蒸腾和茎干充水的发生机制和影响因子目前已有较多研究(McDonald et al., 2002; Daley & Phillips, 2006; Fisher et al., 2007; Zeppel et al., 2014), 但这些研究大多是基于单方位的液流开展。有研究表明, 植物茎干不同方位的液流存在显著差异, 而且这种方位上的差异会显著影响植株水分利用估计的准确性与可靠性, 如Tateishi等(2008)对常绿树种青冈栎(Quercus glauca)的研究发现, 仅测量一个方位的液流对蒸腾量的估计误差高达20%。Tomonori等(2012)对刺槐(Robinia pseudoacacia)和蒙栎(Quercus mongolica)的研究发现, 不同方位间液流的变异系数高达20%-45%, 忽略周向液流的差异会导致蒸腾估算量的误差达到16%-20%。也有****对银杏(Ginkgo biloba)(孙守家等, 2006), 侧柏(Platycladus orientalis)(王华田等, 2006), 日本柳杉(Cryptomeria japonica)(Tsuruta et al., 2010), 樟子松(Pinus sylvestris var. mongolica)(党宏忠等, 2020)等树种不同方位树干边材液流进行研究, 同样认为不同方位树干边材液流存在显著差异, 而且这些差异都会造成对植物茎干水分利用估计的误差。然而目前在国内外众多涉及植物方位间液流差异的研究中, 大部分都是以白天液流为例, 较少涉及夜间液流, 而夜间蒸腾和茎干充水是夜间液流的主要组成部分, 关于它们在植物茎干不同方位上的差异以及主要的影响因子目前仍不清楚, 需要进一步研究。
毛白杨(Populus tomentosa)是我国北方重要的防护与用材林树种, 因其优质、速生、丰产、抗逆性强等优点在我国华北地区广泛种植(康向阳和朱之悌, 2002)。关于毛白杨的茎干液流, 已有部分****对其进行了实验分析。李广德等(2010)的研究表明影响毛白杨不同方位液流的主导气象因子不同。刘洋等(2018)研究了宽窄行模式栽植下的毛白杨不同方位的液流, 表明不同方位液流具有显著的差异性和相关性, 方位间差异不能忽视。还有研究表明, 毛白杨夜间茎干充水量占夜间液流的比例达61%, 且夜间液流受地下水位和水汽压亏缺(VPD)等气象因子的影响(Di et al., 2019)。但是夜间毛白杨茎干不同方位的液流是否具有差异, 以及各方位夜间液流的主要影响因子是什么, 目前的了解还十分有限。基于以上问题, 本试验的研究目的是: (1)阐述毛白杨不同方位夜间液流的差异性及相关性; (2)明确不同方位间夜间蒸腾和茎干充水所占比例及其差异性; (3)探究毛白杨各方位夜间蒸腾和茎干充水的主要影响因子。
1 材料和方法
1.1 试验地概况
试验地位于山东省聊城市高唐县(36.97° N, 116.23° E, 平均海拔27 m), 该地区属于温带半湿润大陆季风气候。年平均气温13.2 ℃, 年降水量为545 mm, 年蒸发量为1 880 mm。年日照总时间达2 651.9 h, 全年无霜期204天。研究对象为栽植于2005年春季的三倍体毛白杨无性系B301人工林, 总面积3.9 hm2。林木采用宽窄行模式栽植(图1), 宽行距6 m, 窄行距2 m, 株距1 m。于2011年5-8月开展试验, 试验期间林分平均树高13.9 m, 平均胸径11.1 cm, 已基本郁闭, 无明显病虫害。林分以地下滴灌的方式进行灌溉, 滴头位于窄行中央和宽行距树60 cm的平行线上, 深20 cm, 滴头相距50 cm。当滴头附近10 cm处的土壤水势(SWP)低于-25 kPa时进行灌溉, 滴头流速为2 L·h-1。在生长季内对林分进行常规施肥, 定期用除草剂除去杂草。本试验涉及的10株样树均位于林分内部, 样树东、南、西、北4个方位的平均冠幅分别为191、164、245和152 cm。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1毛白杨宽窄行模式栽植示意图。
Fig. 1Scheme of wide and narrow row planting of Populus tomentosa.
1.2 测定项目与方法
1.2.1 环境因子本试验涉及的环境因子包括气象因子和土壤水分。气象因子利用距试验地250 m处的自动气象站(Delta-T Devices Ltd., Cambridge, UK)实时监测。测定的气象因子包括太阳总辐射(Rs, kW·m-2)、空气温度(Ta, ℃)、空气相对湿度(RH, %)、风速(v, m·s-1)和风向, 数据每10 min记录采集一次。VPD (kPa)采用Campbell和Norman (1977)的经验公式计算:
土壤水分用土壤水势(ψ, kPa)表示。在试验地布设机械式张力计来测定ψ, 位置定在距离滴头10 cm, 地表下20 cm处, 宽窄行内各布设3个。从实验日起每天7:00对张力计进行读数, 得到ψ值。
1.2.2 茎干液流
茎干液流测定采用热扩散法(Granier, 1985), 从样地中随机选择不同胸径的10株样树进行液流监测(各样树信息见表1), 分别在样木胸高(1.3 m)处, 从东、南、西、北4个方位插入探针, 同时测量树干液流。热扩散探针(TDP 30, Dynamax, Texas, USA)长30 mm, 直径1.2 mm, 探针间距40 mm。试验期间, 液流速率每30 s测定一次, 然后每10 min取平均值并存入数据采集器(Model DL2e, DelTa-T Devices, Cambridge, UK)。液流速率计算公式(Granier, 1985)如下:
Table 1
表1
表1宽窄行模式种植下毛白杨样树的信息及液流监测时期
Table 1
| 编号 Number | 胸径 Diameter at breast height (cm) | 液流测量时期 Sap flow measured date | 测量完整天数 Full measured days | 宽行距位置 Wide row position |
|---|---|---|---|---|
| T1 | 10.32 | 2011-05-16-05-19 | 4 | W |
| T2 | 10.56 | 2011-05-21-05-24 | 4 | E |
| T3 | 11.04 | 2011-05-26-05-29 | 4 | W |
| T4 | 9.80 | 2011-05-31-06-04 | 5 | E |
| T5 | 9.90 | 2011-06-06-06-12 | 7 | E |
| T6 | 10.65 | 2011-06-17-06-23 | 7 | W |
| T7 | 13.30 | 2011-06-25-07-03 | 9 | W |
| T8 | 12.72 | 2011-07-05-07-18 | 13 | W |
| T9 | 7.47 | 2011-07-19-07-29 | 11 | W |
| T10 | 8.35 | 2011-08-02-08-16 | 14 | W |
新窗口打开|下载CSV
式中, Fd为液流速率(cm·s-1); ΔT为加热探针和受热探针间的温差; ΔTm为无液流条件下探针间的温差, 为了准确计算夜间液流, ΔTm每5-10天计算一次。
为消除林木个体大小可能对树干液流速率产生的影响, 利用胸径(DBH)对液流速率进行矫正, 公式(刘洋等, 2018)如下:
式中, Fd′为校正后的液流速率(cm·s-1); $\overline {DBH}$为所有样树的平均胸径(cm); DBH为各样树的胸径(cm)。
单株样木整日的耗水量计算公式(Xi et al., 2017)为:
式中, Asf是整体的液流量(mm·d-1), As是样木的边材面积(cm2), t为一天之内液流存在的时间(s)。
1.2.3 夜间蒸腾与茎干充水的划分
将太阳总辐射(Rs)小于5.0 W·m-2的时间段定义为夜间(Daley & Phillips, 2006)。采用Fisher等(2007)的方法区分夜间蒸腾量(Nt)和茎干充水量(Sr)。根据每晚的液流情况, 采用指数函数拟合出无Nt情况下的液流变化趋势, 拟合曲线(图2虚线)下的面积为Sr, 将两个曲线之间的面积定义为Nt。
图2
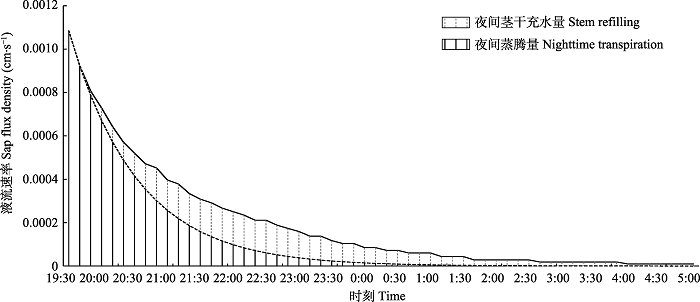 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2毛白杨夜间蒸腾和茎干充水区分方法示意图。
Fig. 2Scheme of distinguishing nighttime transpiration from stem refilling of Populus tomentosa.
1.3 数据分析
采用配对样本t检验(Paired-samples t-test)的方法分析毛白杨茎干不同方位的夜间液流的差异、相关性以及不同方位夜间蒸腾和茎干充水的差异。在所有样树中, T8样树液流监测时间较长, 且监测期内气象因子较为稳定, 因此T8样树为代表, 采用Spearman相关性检验和线性回归分析影响Nt和Sr的因子。数据由Excel 2017和SPSS 17进行处理, 图表采用Origin 9.0绘制。2 结果
2.1 气象因子变化
在研究期内, 白天和夜间的VPD、Ta、RH和v具有相似的变化趋势(图3)和显著的相关性(p < 0.01, 表2)。配对样本t检验表明4个气象因子在白天与夜间均具有显著差异(p < 0.01, 表1), 白天的VPD、Ta和v分别平均比夜间高148%、17%、59%, 而RH则比夜间低14%。研究时期内降水(P)主要集中在5月中旬、6月下旬至7月上旬以及7月下旬至8月上旬。土壤水势在-50-0 kPa内波动变化。图3
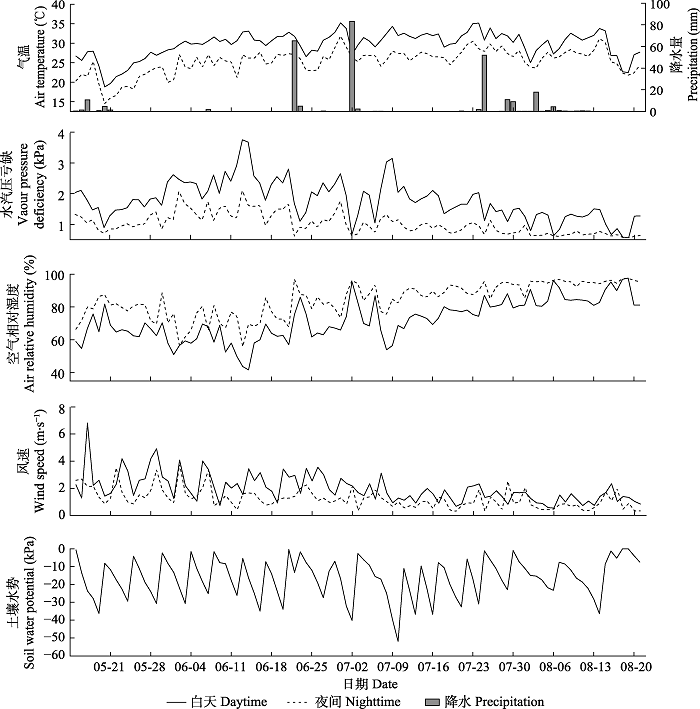 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3宽窄行模式栽植下毛白杨液流研究期内环境因子变化。
Fig. 3Variation of environmental factors during the sap flow study period of Populus tomentosa which planted in wide and narrow rows.
Table 2
表2
表2宽窄行模式栽植下毛白杨液流研究期内昼夜气象因子差异及相关性
Table 2
| Ta (℃) | VPD (kPa) | RH (%) | v (m·s-1) | |
|---|---|---|---|---|
| 白天-夜间 Daytime-nighttime | 4.34** | 0.77** | -12.08** | 0.73** |
| 相关系数 Correlation coefficient | 0.812** | 0.879** | 0.849** | 0.533** |
新窗口打开|下载CSV
2.2 不同方位夜间液流的差异及相关性
不论宽行距在样树东侧还是西侧, 样树不同方位夜间液流量(Q)都具有较大差异(图4), 方位间变异系数(CV)分别为29.3%、35.8%。配对样本t检验 (表3)表明, 宽行距位于东侧的样树, 其西方位的Q显著大于北方位, 而其他方位间差异不显著; 宽行距位于西侧的样树, 其西方位的Q显著大于其他3个方位, 南方位的Q显著低于西方位和北方位, 而其他方位间Q差异不显著。10株样树各方位间Q均呈现显著相关关系, 但不同样树的不同方位之间Q的相关性具有一定差异。总的来说, 宽行距位于东侧的样树的相关性较高, 其中南方位和北方位间Q的相关性最高, 西方位和北方位间Q的相关性最低。宽行距位于西侧的样树的相关性较低, 其中, 东方位和北方位间Q的相关性最高, 西方位和北方位间Q的相关性最低。图4
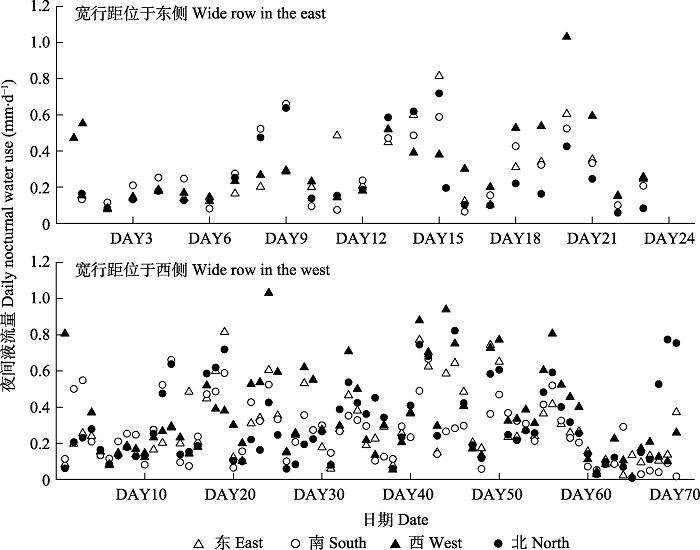 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4宽窄行模式种植下毛白杨不同方位的夜间液流量变化。DAY, 开始监测液流的天数。
Fig. 4Variation of differently azimuthal nocturnal sap flux of Populus tomentosa which planted in wide and narrow rows. DAY, the number of days to start monitoring the sap flow.
Table 3
表3
表3宽窄行模式栽植下的毛白杨不同方位夜间液流配对样本t检验结果
Table 3
| 方位 Orientation | E-S | E-W | E-N | S-W | S-N | W-N | n | |
|---|---|---|---|---|---|---|---|---|
| 宽行距位于东侧 Wide row in the east | 平均差异 Average difference | -0.005 | -0.027 | 0.021 | -0.022 | 0.026 | 0.049* | 23 |
| 相关系数 Correlation coefficient | 0.644** | 0.612** | 0.739** | 0.586** | 0.895** | 0.434* | ||
| 宽行距位于西侧 Wide row in the west | 平均差异 Average difference | 0.029 | -0.11** | -0.035 | -0.14** | -0.063* | 0.075* | 70 |
| 相关性 Correlation coefficient | 0.621** | 0.471** | 0.675** | 0.609** | 0.445** | 0.344* |
新窗口打开|下载CSV
2.3 夜间蒸腾和充水
2.3.1 不同方位夜间蒸腾量和茎干充水量的差异图5为各方位Nt和Sr以及茎干充水量占夜间液流量的比例(Sr/Q)大小。各方位Nt和Sr均具有显著差异, 方位间Sr的差异较大, 平均变异系数为45.6%, Nt的差异较小, 平均变异系数为39.2%。夜间充水量占夜间液流的比例较大, 东、南、西、北4个方位的平均占比分别为54.2%、60.7%、54.5%、57.4%。配对样本t检验结果(表4)表明, 宽行距位于东侧的样树西方位的Nt和Sr均最大, 其中西方位的Sr显著大于其他3个方位。北方位的Nt最小, 且显著小于其他3个方位, 其他方位间Nt和Sr无显著差异。各方位间的Sr/Q无显著差异。宽行距位于西侧的样树西方位的Nt和Sr亦均最大, 其中西方位的Sr显著大于东方位的和南方位的。南方位的Nt最小, 显著小于西方位的和北方位的, 其他方位间Nt和Sr无显著差异。南方位的Sr/Q显著大于其他3个方位。
图5
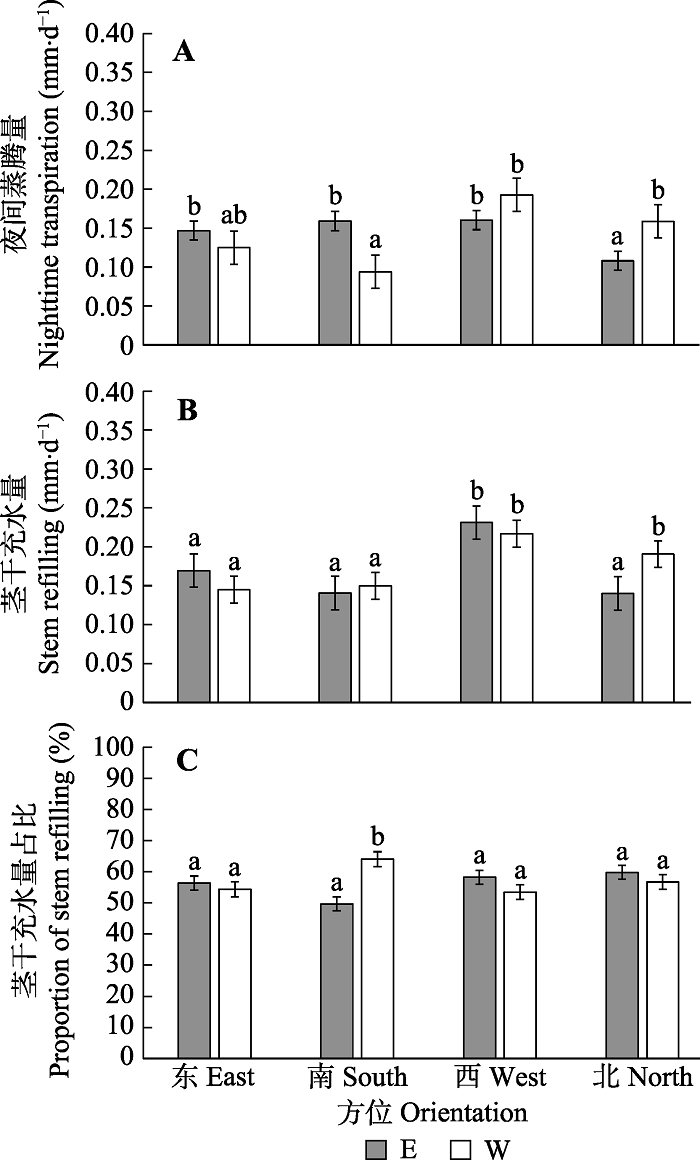 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5毛白杨不同方位夜间蒸腾、茎干充水以及充水所占比例的平均值(平均值±标准偏差)。E、W分别表示宽行位于样树的东侧和西侧。柱状图上的不同字母表示方位间的差异显著。
Fig. 5Average value (mean ± SD) of nocturnal transpiration, stem refilling, and proportion of stem refilling in different orientations of Populus tomentosa. E and W indicate wide rows are in the east and west of the sample trees, respectively. Different lowercase letters indicate significant difference in different directions.
Table 4
表4
表4宽窄行模式种植下毛白杨不同方位夜间蒸腾量和茎干充水量配对样本t检验结果
Table 4
| E-S | E-W | E-N | S-W | S-N | W-N | ||
|---|---|---|---|---|---|---|---|
| 宽行距位于东侧 Wide row in the east | Nt | -0.012 | -0.013 | 0.039** | -0.001 | 0.051** | 0.052** |
| Sr | 0.029 | -0.062* | 0.029 | -0.091** | 0.001 | 0.091* | |
| Sr/Q | 0.067 | -0.019 | -0.034 | -0.086 | -0.100 | -0.015 | |
| 宽行距位于西侧 Wide row in the west | Nt | 0.031 | -0.068 | -0.034 | -0.099** | -0.065** | 0.034 |
| Sr | -0.005 | -0.072** | -0.046** | -0.070** | -0.041* | 0.026 | |
| Sr/Q | -0.096** | 0.009 | -0.024 | 0.110** | 0.073** | -0.033 |
新窗口打开|下载CSV
2.3.2 夜间蒸腾与茎干充水的主要影响因子
不同方位Sr和Nt与白天对应各方位液流的互相关检验结果(表5)表明, 各方位Nt与白天对应方位的液流均无显著相关关系, Sr与白天对应各方位液流均有显著正相关关系, 且北方位的相关性最低。说明白天液流对Sr有显著影响, 而对Nt无显著影响。
Table 5
表5
表5宽窄行模式栽植下毛白杨各方位夜间蒸腾量、茎干充水量与白天液流的相关系数
Table 5
| E | S | W | N | |
|---|---|---|---|---|
| Nt | 0.096 | 0.189 | -0.203 | 0.015 |
| Sr | 0.436** | 0.377** | 0.471** | 0.250* |
新窗口打开|下载CSV
Nt、Sr与环境因子的相关性检验(表6)表明, 各方位Nt和Sr均与VPD呈显著正相关关系, 除南方位的Sr外均与RH呈显著负相关关系, 仅东方位的Nt和Sr与Ta呈显著负相关关系, 而各方向Nt和Sr均未与v达到显著相关关系。Nt和Sr方位间的差异(NtCV、SrCV)与VPD、Ta、RH、v、ψ均无显著相关关系, 表明毛白杨Nt和Sr方位间的差异不受环境因子影响。图6为T8样树Nt和Sr与VPD之间的线性回归关系, 由R2可知, 各方位的Nt与VPD的线性相关关系较为显著, 说明Nt受VPD的影响较大。
Table 6
表6
表6环境因子与毛白杨各方位夜间蒸腾量和茎干充水量的相关系数
Table 6
| VPD | Ta | RH | v | ψ | |
|---|---|---|---|---|---|
| NtE | 0.590** | -0.611* | -0.814** | 0.188 | 0.042 |
| NtS | 0.760** | 0.102 | -0.613* | -0.310 | -0.354 |
| NtW | 0.577* | -0.334 | -0.602* | -0.026 | -0.178 |
| NtN | 0.678** | -0.288 | -0.623* | -0.099 | -0.212 |
| NtCV | -0.385 | -0.194 | 0.152 | 0.345 | 0.398 |
| SrE | 0.584* | -0.472* | -0.554* | -0.297 | -0.170 |
| SrS | 0.555** | 0.068 | -0.411 | -0.514 | -0.276 |
| SrW | 0.576* | -0.129 | -0.468* | -0.421 | -0.261 |
| SrN | 0.521* | -0.313 | -0.457* | -0.296 | -0.067 |
| SrCV | 0.152 | 0.159 | -0.078 | -0.321 | 0.046 |
新窗口打开|下载CSV
图6
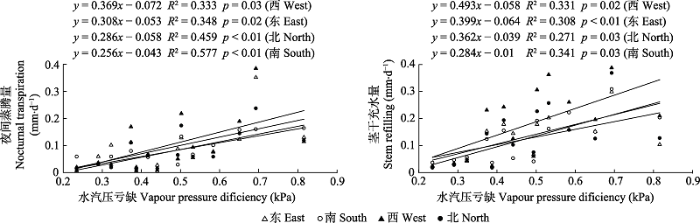 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6毛白杨各方位夜间蒸腾量和茎干充水量与水汽压亏缺的线性关系。
Fig. 6Linear relationship between nocturnal transpiration, stem refilling in different orientations of Populus tomentosa and vapour pressure deficiency.
3 讨论
3.1 各方位夜间液流量的差异及相关性
本研究中, 所有方位的夜间液流量都表现出显著的相关性(p < 0.05)(表3)。同样地, 李广德等(2010)的实验也表明, 不管是晴天、阴天、雨天, 还是全天、白天、夜间, 三倍体毛白杨东、南、西、北4个方位液流速率相互间均呈极显著的正相关关系。这说明毛白杨各方位木质部是同步进行白天以及夜间的水分运输。Waisel等(1972)发现染液在胡杨茎干中呈螺旋状或者环状上升, 直接证明了不同方位木质部中水分运输的关联。此外, 本研究中林分郁闭, 不同方位冠层所处的环境均匀且相似, 受气象因子、树形等因子同步影响, 因此也会使各方位的夜间液流量表现出相同的变化趋势与显著相关性。除了相关性, 实验结果表明, 各方位夜间液流量也存在差异, 其可能的原因主要有两点。(1)样树本身的结构造成了树干液流方位上的差异。李广德等(2010)对毛白杨周向液流的研究发现由于各方位树冠结构、树干边材面积、根系分布等的不同, 导致树干各方位的水分运输效率不同。也有****使用染液对日本柏树的边材进行染色, 通过颜色变化确定边材运输水分的面积, 并分析了树干周向液流差异与边材面积差异的关系, 得出结论, 二者显著相关(R2 = 0.49, p < 0.01), 说明树干不同方位的边材面积确实会影响不同方位的液流量(Tsuruta et al., 2010)。(2)周向液流的差异与土壤水分有关, 土壤水分的空间异质性影响着树干液流的空间异质性, 导致树干周向液流出现差异。Lu等(2000)对杧果(Mangifera indica)树的研究印证了这个观点, 他发现杧果树干周向液流的差异是土壤水分分布不均造成的, 而且在不同灌溉条件下, 各方位液流大小的排序不同。
3.2 不同方位夜间蒸腾量和茎干充水量的差异及其所占比例
本研究中, 各方位夜间蒸腾量和茎干充水量均具有较大的差异, 其中茎干充水量方位间的差异(CV = 45.6%)大于夜间蒸腾量(CV = 39.2%)。而不论夜间蒸腾量还是茎干充水量, 西方位都是优势方位。这些现象可能与环境因子以及树木茎干木质部结构有关。夜间茎干充水主要用于补充木质部中白天蒸腾耗散的水分, 与白天蒸腾量具有显著相关性(Wang et al., 2012), 而蒸腾主要受PAR和VPD的影响(Poyatos et al., 2007)。在白天, 随着太阳移动, 不同方位树冠所受的光照时间、强度不同, 因此蒸腾可能表现出较大的差异; 而在夜间无太阳辐射, 各方位蒸腾量的差异较小。木质部中的导管是水分运输的主要通道, 在长期水平上, 树木可以通过调整边材导管的直径和密度来调整水分运输(February et al., 1995)。研究表明, 在一天中, 下午的太阳辐射、VPD较上午强(马金玉等, 2007), 而西侧树冠在下午时受太阳直射, 导致西方位冠层具有较大的蒸腾量。长此以往, 西侧木质部可能会发展出水分运输能力较强的导管组织, 有可能因此导致夜间蒸腾和茎干充水的能力大于其他各方位。夜间蒸腾和茎干充水具有不同的生理意义, 因此二者占夜间液流的比例有所不同(Fisher et al., 2007)。有研究表明, 夜间茎干充水是植物夜间用水的主要方式, 约占夜间液流量的50%-70% (Fisher et al., 2007; 张婕等, 2019)。与前人的研究结果一样, 在本研究中, 夜间茎干充水量所占比例较高, 4个方位(东、西、南、北)分别是54.2%、60.7%、54.5%、57.4%, 南方位的比例显著大于其他3个方位(表4)。本实验样地位于华北地区, 昼夜温差较大(表2)。白天温度高, 蒸腾剧烈, 导致夜间水分亏缺严重, 因此夜间充水比例较大。而夜间温度降低, 导致VPD较低, 夜间蒸腾也相应减弱, 因此蒸腾量占夜间用水的比例较小。研究表明, 在不同季节、不同地点, 不同树种的夜间蒸腾量和茎干充水量所占的比例有所不同, 比如Fisher等(2007)的研究发现Quercus douglasii在冬季几乎无夜间蒸腾, 夜间液流活动表现为茎干充水, 而在夏季夜间蒸腾占夜间液流量的30%, 茎干充水占70%。Daley和Phillips (2006)发现Betula papyrifera只有夜间蒸腾而无补水。干旱地区Acacia mangium夜间茎干充水量占全天液流量的比例约为14.7%, 在较湿润的地区这个比例高达30.3% (Wang et al., 2012)。
3.3 各方位夜间蒸腾和茎干充水的影响因子
本实验结果表明, VPD为各方位夜间蒸腾与茎干充水的主要影响因子, 但其对夜间蒸腾的影响更大(表6)。植物夜间蒸腾是一项被动活动, 对气象因子, 尤其对VPD变化有较强的响应(Dawson et al., 2007), 而茎干充水是植物对蒸腾引起的水分亏缺的响应(Wang et al., 2012), 因此后者对VPD变化的响应较为迟缓。Daley和Phillips (2006)发现VPD影响着Betula papyrifera夜间气孔的开闭, 进而影响气体交换和夜间蒸腾作用的时间及强度。Chen等(2020)研究了影响不同龄级油松(Pinus tabuliformis)和元宝槭(Acer truncatum)夜间液流的气象因子, 发现夜间VPD是幼龄林的夜间液流活动的主要影响因子。而本研究的样树在实验时正处于幼龄期, 其夜间蒸腾和茎干充水等夜间液流活动对VPD的响应较为强烈。本研究还发现不同方位夜间茎干充水受到白天对应方位蒸腾量的影响显著(表5), 与Snyder (2003)结果一致。Snyder (2003)发现夜间植物茎干补水与白天液流的增加有关(R2 = 0.28, n = 522, p < 0.01), 大量的水分从地下补充到茎干中, 用来弥补白天因蒸腾而导致的水分亏缺。此外, 以前的研究也表明气象因子也能够影响树干木质部空间上的液流差异。比如, Tomonori等(2012)发现环孔木材树种树干外侧液流显著大于内侧, 呈现由外到内递减的趋势。而Poyatos等(2007)对Quercus pubescens径向液流研究发现VPD、PAR等气象因子是导致林木尤其是环孔木材树种出现径向液流出现差异的原因之一。根据管道模型理论, 单元光合器官耗水的增加会引起相应单位管道水分运输的增加。外侧树冠受气象因子的影响较大, 夜间蒸腾作用也会相应增加, 为了满足外侧树冠蒸腾耗水的需要, 其对应树干内的管道运输水分的能力较强, 而内侧树冠对应的管道水分运输能力较弱, 这是树干径向液流出现差异的主要原因(Manuel et al., 2001)。但是本实验结果有所不同, 环境因子没有造成树干夜间液流在周向空间上的的显著差异。这可能是因为夜间树干不同方位的环境条件相差不大, 对各方位液流的影响有限, 故不会造成方位间夜间液流的差异。而至于差异出现的主要原因, 今后有必要研究不同方位木质部导管组织结构差异对夜间树干周向液流差异的影响。
3.4 关于夜间液流的研究建议
本实验对毛白杨不同方位的夜间蒸腾和茎干充水等液流行为进行了研究, 分析方位间夜间蒸腾和茎干充水等夜间液流活动的差异及相关性, 以及各方位夜间蒸腾和茎干充水的影响因子, 包括气象因子和白天液流。但是还有很多问题需要进一步研究, 例如已有的实验发现地理位置(Zeppel et al., 2014), 土壤水分(Chen et al., 2014; Di et al., 2019), 树龄和树形因子(Chen et al., 2020)等会对树木的夜间液流产生影响, 但是这些影响因素对方位间夜间液流差异的影响还不清楚。方位间夜间液流量的差异对整个林分水分利用估计的影响, 以及如何避免这些误差仍有待研究。因此, 以后的研究应更多地集中在林分、生态系统等大尺度上, 以具有更广泛的适用性。4 结论
对宽窄行模式栽植下的10株样树的夜间液流进行监测分析, 发现不论宽行在样树东侧还是西侧, 样树西方位的夜间蒸腾和茎干充水等液流活动都最大, 显著或者不显著大于其他方位, 其他方位间的大小次序没有明显规律。夜间茎干充水量占夜间液流量的比例较大, 4个方位(东、南、西、北)分别为54.2%、60.7%、54.5%、57.4%。气象因子中的VPD对4个方位的夜间蒸腾和茎干充水均有显著影响, 但气象因子不是导致方位间夜间蒸腾和茎干充水出现差异的原因。白天液流对夜间蒸腾的影响不显著, 但对夜间茎干充水的影响显著。通过本实验可基本了解树木不同方位夜间液流的差异, 进一步加深对人工林水分关系的认识。致谢
感谢美国农业部林业局Mark D. Coleman教授帮助修改本文英文。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
DOI:10.1093/treephys/19.13.853URLPMID:10562402 [本文引用: 1]

Sap flow measurements showed that a well-watered four-year-old plantation of Eucalyptus grandis (Hill ex Maiden) at Wagga Wagga, New South Wales, Australia, used 0.8 mm of water between 2100 and 0500 h on the midwinter night of July 30. Sap flow ceased for 2 to 3 h after sunset before recommencing at high rates that reached a maximum of 0.3 mm per h between 0200 and 0300 h. This pattern is inconsistent with the replenishment of tissue water reserves depleted during the day. Moreover, maximum leaf conductance at night was about 20 times maximum cuticular conductance values reported in the literature, which strongly suggests that stomata were partly open and that there was substantial water loss by way of the foliage. In an 8-month period from late winter to mid-autumn, comparable rates of nighttime water use were observed on only one other occasion. However, water use at rates of 0.1 mm per h or more occurred on 24 other nights. Almost 70% of the variation in nighttime sap velocity was explained by nighttime mean vapor pressure deficit and nighttime mean wind speed. Total recorded nighttime water use of the plantation was 48 mm, or 5% of total transpiration during the 8-month study. In view of the insensitivity of heat pulse measurements at low sap flows, this value may be an underestimate of actual nighttime transpiration.
URLPMID:11662225 [本文引用: 1]
DOI:10.1093/treephys/tpu087URLPMID:25428825 [本文引用: 1]
Wood biophysical properties and the dynamics of water storage discharge and refilling were studied in the trunk of canopy tree species with diverse life history and functional traits in subtropical forests of northeast Argentina. Multiple techniques assessing capacitance and storage capacity were used simultaneously to improve our understanding of the functional significance of internal water sources in trunks of large trees. Sapwood capacitances of 10 tree species were characterized using pressure-volume relationships of sapwood samples obtained from the trunk. Frequency domain reflectometry was used to continuously monitor the volumetric water content in the main stems. Simultaneous sap flow measurements on branches and at the base of the tree trunk, as well as diurnal variations in trunk contraction and expansion, were used as additional measures of stem water storage use and refilling dynamics. All evidence indicates that tree trunk internal water storage contributes from 6 to 28% of the daily water budget of large trees depending on the species. The contribution of stored water in stems of trees to total daily transpiration was greater for deciduous species, which exhibited higher capacitance and lower sapwood density. A linear relationship across species was observed between wood density and growth rates with the higher wood density species (mostly evergreen) associated with lower growth rates and the lower wood density species (mostly deciduous) associated with higher growth rates. The large sapwood capacitance in deciduous species may help to avoid catastrophic embolism in xylem conduits. This may be a low-cost adaptation to avoid water deficits during peak water use at midday and under temporary drought periods and will contribute to higher growth rates in deciduous tree species compared with evergreen ones. Large capacitance appears to have a central role in the rapid growth patterns of deciduous species facilitating rapid canopy access as these species are less shade tolerant than evergreen species.
[本文引用: 1]
[本文引用: 2]
DOI:10.1093/treephys/26.4.411URLPMID:16414920 [本文引用: 4]
Transpiration is generally assumed to be insignificant at night when stomata close in response to the lack of photosynthetically active radiation. However, there is increasing evidence that the stomata of some species remain open at night, which would allow for nighttime transpiration if there were a sufficient environmental driving force. We examined nighttime water use in co-occurring species in a mixed deciduous stand at Harvard Forest, MA, using whole-tree and leaf-level measurements. Diurnal whole-tree water use was monitored continuously with Granier-style sap flux sensors in paper birch (Betula papyrifera Marsh.), red oak (Quercus rubra L.) and red maple (Acer rubrum L.). An analysis was conducted in which nighttime water flux could be partitioned between refilling of internal water stores and transpiration. Substantial nighttime sap flux was observed in all species and much of this flux was attributed to the refilling of depleted water stores. However, in paper birch, nighttime sap flux frequently exceeded recharge estimates. Over 10% of the total daily sap flux during the growing season was due to transpiration at night in paper birch. Nighttime sap flux was over 8% of the total daily flux in red oak and 2% in red maple; however, this flux was mainly associated with recharge. On nights with elevated vapor pressure deficit, sap flux continued through the night in paper birch, whereas it reached zero during the night in red oak and red maple. Measurements of leaf-level gas exchange on a night with elevated vapor pressure deficit showed stomatal conductance dropping by only 25% in paper birch, while approaching zero in red oak and red maple. The study highlighted differences in ecophysiological controls on sap flux exerted by co-occurring species. Paper birch is a fast-growing, shade-intolerant species with an earlier successional status than red oak and red maple. Risking water loss through nighttime transpiration may provide paper birch with an ecological advantage by enabling the species to maximize photosynthesis and support rapid growth. Nighttime transpiration may also be a mechanism for delivering oxygen to respiring cells in the deep sapwood of paper birch.
[本文引用: 1]
[本文引用: 1]
DOI:10.1093/treephys/27.4.561URLPMID:17241998 [本文引用: 2]
It is commonly assumed that transpiration does not occur at night because leaf stomata are closed in the dark. We tested this assumption across a diversity of ecosystems and woody plant species by various methods to explore the circumstances when this assumption is false. Our primary goals were: (1) to evaluate the nature and magnitude of nighttime transpiration, E(n), or stomatal conductance, g(n); and (2) to seek potential generalizations about where and when it occurs. Sap-flow, porometry and stable isotope tracer measurements were made on 18 tree and eight shrub species from seven ecosystem types. Coupled with environmental data, our findings revealed that most of these species transpired at night. For some species and circumstances, nighttime leaf water loss constituted a significant fraction of total daily water use. Our evidence shows that E(n) or g(n) can occur in all but one shrub species across the systems we investigated. However, under conditions of high nighttime evaporative demand or low soil water availability, stomata were closed and E(n) or g(n) approached zero in eleven tree and seven shrub species. When soil water was available, E(n) or g(n) was measurable in these same species demonstrating plasticity for E(n) or g(n). We detected E(n) or g(n) in both trees and shrubs, and values were highest in plants from sites with higher soil water contents and in plants from ecosystems that were less prone to atmospheric or soil water deficits. Irrespective of plant or ecosystem type, many species showed E(n) or g(n) when soil water deficits were slight or non-existent, or immediately after rainfall events that followed a period of soil water deficit. The strongest relationship was between E(n) or g(n) and warm, low humidity and (or) windy (> 0.8 m s(-1)) nights when the vapor pressure deficit remained high (> 0.2 kPa in wet sites, > 0.7 kPa in dry sites). Why E(n) or g(n) occurs likely varies with species and ecosystem type; however, our data support four plausible explanations: (1) it may facilitate carbon fixation earlier in the day because stomata are already open; (2) it may enhance nutrient supply to distal parts of the crown when these nutrients are most available (in wet soils) and transport is rapid; (3) it may allow for the delivery of dissolved O(2) via the parenchyma to woody tissue sinks; or (4) it may occur simply because of leaky cuticles in older leaves or when stomata cannot close fully because of obstructions from stomatal (waxy) plugs, leaf endophytes or asymmetrical guard cells (all non-adaptive reasons). We discuss the methodological, ecophysiological, and theoretical implications of the occurrence of E(n) or g(n) for investigations at a variety of scales.
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1093/treephys/27.4.597URLPMID:17242001 [本文引用: 7]
At the leaf scale, it is a long-held assumption that stomata close at night in the absence of light, causing transpiration to decrease to zero. Energy balance models and evapotranspiration equations often rely on net radiation as an upper bound, and some models reduce evapotranspiration to zero at night when there is no solar radiation. Emerging research is showing, however, that transpiration can occur throughout the night in a variety of vegetation types and biomes. At the ecosystem scale, eddy covariance measurements have provided extensive data on latent heat flux for a multitude of ecosystem types globally. Nighttime eddy covariance measurements, however, are generally unreliable because of low turbulence. If significant nighttime water loss occurs, eddy flux towers may be missing key information on latent heat flux. We installed and measured rates of sap flow by the heat ratio method (Burgess et al. 2001) at two AmeriFlux (part of FLUXNET) sites in California. The heat ratio method allows measurement and quantification of low rates of sap flow, including negative rates (i.e., hydraulic lift). We measured sap flow in five Pinus ponderosa Dougl. ex Laws. trees and three Arctostaphylos manzanita Parry and two Ceanothus cordulatus A. Kellog shrubs in the Sierra Nevada Mountains, and in five Quercus douglasii Hook and Arn. trees at an oak savanna in the Central Valley of California. Nocturnal sap flow was observed in all species, and significant nighttime water loss was observed in both species of trees. Vapor pressure deficit and air temperature were both well correlated with nighttime transpiration; the influence of wind speed on nighttime transpiration was insignificant at both sites. We distinguished between storage-tissue refilling and water loss based on data from Year 2005, and calculated the percentage by which nighttime transpiration was underestimated by eddy covariance measurements at both sites.
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
URLPMID:12651518 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1093/treephys/21.6.409URLPMID:11282581 [本文引用: 1]
Within-tree variation in sap flow density (SFD) was measured in two isolated evergreen oak (Quercus ilex L.) trees growing in an oak savannah (dehesa) in southwest Spain. Sap flow was estimated by the constant heating method. Three sensors were installed in the trunk of each tree in three orientations: northeast (NE), northwest (NW) and south (S). Sap flow density was monitored continuously from May 18 to September 27, 1993. Daily values of SFD ranged between 500 and 4500 mm3 mm-2 day-1. There were significant differences in SFD between orientations; SFD was higher in the NE and NW orientations than in the S orientation. These differences were noted on both a daily and seasonal time scale, and were less pronounced on cloudy days and at the end of the drought period, when SFD was relatively low. Our results support the idea that branches of trees can be viewed as a collection of small independent plants.
DOI:10.1093/treephys/27.4.577URLPMID:17241999 [本文引用: 1]
Contrary to the conventional theory of optimal stomatal control, there is substantial transpiration at night in many tree species, but the functional significance of this phenomenon remains uncertain. To investigate the possible roles of nocturnal transpiration, we compared and contrasted the correlations of both nocturnal and diurnal sap flow with a range of traits in 21 temperate deciduous tree species. These traits included soil water affinity, shade tolerance, cold hardiness, nitrogen concentration of tissues, minimum transpiration rate of excised leaves, growth rate, photosynthetic capacity, stomatal length and density, and the water potential and relative water content of leaves at the wilting point. Nocturnal sap flow was higher in species with higher leaf nitrogen concentrations, higher rates of extension growth and lower shade tolerances. Diurnal sap flow was higher in species with higher leaf nitrogen concentrations and photosynthetic capacities on a leaf area basis. Because leaf metabolism and dark respiration, in particular, are strongly related to leaf nitrogen concentration, our findings suggest that nocturnal transpiration functions to sustain carbohydrate export and other processes driven by dark respiration, and that this function is most important in fast- growing shade-intolerant tree species.
DOI:10.1071/FP02007URLPMID:32689563 [本文引用: 1]
[本文引用: 1]
DOI:10.1093/jxb/erv193URLPMID:25954047 [本文引用: 1]
Global warming causes night temperature (NT) to increase faster than day temperature in the tropics. According to crop growth models, respiration incurs a loss of 40-60% of photosynthate. The thermal sensitivity of night respiration (R(n)) will thus reduce biomass. Instantaneous and acclimated effects of NT on R(n) of leaves and seedlings of two rice cultivars having a variable level of carbohydrates, induced by exposure to different light intensity on the previous day, were investigated. Experiments were conducted in a greenhouse and growth chambers, with R(n) measured on the youngest fully expanded leaves or whole seedlings. Dry weight-based R(n) was 2.6-fold greater for seedlings than for leaves. Leaf R(n) was linearly related to starch (positive intercept) and soluble sugar concentration (zero intercept). Increased NT caused higher R(n) at a given carbohydrate concentration. The change of R(n) at NT increasing from 21 degrees C to 31 degrees C was 2.4-fold for the instantaneous response but 1.2- to 1.7-fold after acclimation. The maintenance component of R(n) (R(m)'), estimated by assimilate starvation, averaged 28% in seedlings and 34% in leaves, with no significant thermal effect on this ratio. The acclimated effect of increased NT on R(m)' across experiments was 1.5-fold for a 10 degrees C increase in NT. No cultivar differences were observed in R(n) or R(m)' responses. The results suggest that the commonly used Q10=2 rule overestimates thermal response of respiration, and R(n) largely depends on assimilate resources.
DOI:10.1093/treephys/27.4.537URLPMID:17241996 [本文引用: 2]
Radial variation in sap flux density across the sapwood was assessed by the heat field deformation method in several trees of Quercus pubescens Wild., a ring-porous species. Sapwood depths were delimited by identifying the point of zero flow in radial patterns of sap flow, yielding tree sapwood areas that were 1.5-2 times larger than assumed based on visual examinations of wood cores. The patterns of sap flow varied both among trees and diurnally. Rates of sap flow were higher close to the cambium, although there was a significant contribution from the inner sapwood, which was greater (up to 60% of total flow) during the early morning and late in the day. Accordingly, the normalized difference between outer and inner sapwood flow was stable during the middle of the day, but showed a general decline in the afternoon. The distribution of sap flux density across the sapwood allowed us to derive correction coefficients for single-point heat dissipation sap flow measurements. We used daytime-averaged coefficients that depended on the particular shape of the radial profile and ranged between 0.45 and 1.28. Stand transpiration calculated using the new method of estimating sapwood areas and the radial correction coefficients was similar to (Year 2003), or about 25% higher than (Year 2004), previous uncorrected values, and was 20-30% of reference evapotranspiration. We demonstrated how inaccuracies in determining sapwood depths and mean sap flux density across the sapwood of ring-porous species could affect tree and stand transpiration estimates.
DOI:10.1016/0002-1571(74)90018-1URL [本文引用: 1]
DOI:10.1093/jxb/erg082URLPMID:12554729 [本文引用: 2]
Significant night-time stomatal conductance and transpiration were found for 11 out of 17 species with a range of life histories (herbaceous annual, perennial grass, shrub, tree), photosynthetic pathways (C(3), C(4)), and habitats in the western United States. Across species and habitats, higher night-time conductance and transpiration were associated with higher daytime values. The prevalence, mechanisms and ecological implications of substantial night-time water loss deserve further investigation.
DOI:10.11707/j.1001-7488.20060505URL [本文引用: 1]
Ginkgo biloba. The results indicated that sap flow velocity was significantly different at different heights, depths and directions of the trunk. Sap flow velocity of upper position at the trunk was more than that of the middle and lower position. But cumulative flux was not significantly different at upper, middle and lower section. Sap flow velocity at 10 mm was the most and that at 20 mm was the least, but sap flow velocity at 5 mm and 15 mm was similar and took the second place among four depths. It also showed that sap flow velocity of the south was the most and that of the west was the second place among the different directions. And an Automatic Weather Station of HOBO was synchronously applied to measure meteorological parameters, which were used to analyze the relationship with changes of trunk sap flow velocity. The results indicated that change of sap flow velocity was a single-peak curve in fine day and multi-peak curve in cloudy and rainy day. In addition, Stepwise regression analyses revealed that PAR, temperature and wind speed were the main environmental factors affecting sap flow velocity. The efficient means to reduce water transpiration of the trees were tried to find through investigating the effect of techniques for inhibiting transpiration including pruning-leaf, overshadowing, spraying of antitranspirants. And the results indicated that spraying of antitranspirants, pruning-leaf and overshadowing could significantly reduce transpiration but the effect of pruning-leaf and overshadowing was far better than that of spraying of antitranspirants.]]>
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 3]
[本文引用: 1]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1093/treephys/tpu089URLPMID:25413023 [本文引用: 5]
Total daily water use is a key factor influencing the growth of many terrestrial plants, and reflects both day-time and nocturnal water fluxes. However, while nocturnal sap flow (En) and stomatal conductance (gs,n) have been reported across a range of species, ecosystems and microclimatic conditions, the regulation of these fluxes remains poorly understood. Here, we present a framework describing the role of abiotic and biotic factors in regulating En and gs,n highlighting recent developments in this field. Across ecosystems, En and gs,n generally increased with increasing soil water content and vapor pressure deficit, but the interactive effects of these factors and the potential roles of wind speed and other abiotic factors remain unclear. On average, gs,n and En are higher in broad-leaved compared with needle-leaved plants, in C3 compared with C4 plants, and in tropical compared with temperate species. We discuss the impacts of leaf age, elevated [CO2] and refilling of capacitance on night-time water loss, and how nocturnal gs,n may be included in vegetation models. Younger leaves may have higher gs,n than older leaves. Embolism refilling and recharge of capacitance may affect sap flow such that total plant water loss at night may be less than estimated solely from En measurements. Our estimates of gs,n for typical plant functional types, based on the published literature, suggest that nocturnal water loss may be a significant fraction (10-25%) of total daily water loss. Counter-intuitively, elevated [CO2] may increase nocturnal water loss. Assumptions in process-based ecophysiological models and dynamic global vegetation models that gs is zero when solar radiation is zero are likely to be incorrect. Consequently, failure to adequately consider nocturnal water loss may lead to substantial under-estimation of total plant water use and inaccurate estimation of ecosystem level water balance.
[本文引用: 1]
[本文引用: 1]
Is nighttime transpiration enhanced after fog events?
1
2013
... 蒸腾是植物消耗水分的主要途径, 在植物生长发育过程中起着至关重要的作用(
Nighttime water use in an irrigated Eucalyptus grandis plantation
1
1999
... 蒸腾是植物消耗水分的主要途径, 在植物生长发育过程中起着至关重要的作用(
An Introduction to Environmental Biophysics. Springer-Verlag
1
1977
... 本试验涉及的环境因子包括气象因子和土壤水分.气象因子利用距试验地250 m处的自动气象站(Delta-T Devices Ltd., Cambridge, UK)实时监测.测定的气象因子包括太阳总辐射(Rs, kW·m-2)、空气温度(Ta, ℃)、空气相对湿度(RH, %)、风速(v, m·s-1)和风向, 数据每10 min记录采集一次.VPD (kPa)采用
Water storage dynamics in the main stem of subtropical tree species differing in wood density, growth rate and life history traits
1
2015
... 蒸腾是植物消耗水分的主要途径, 在植物生长发育过程中起着至关重要的作用(
Soil water affects transpiration response to rainfall and vapor pressure deficit in poplar plantation
1
2014
... 本实验对毛白杨不同方位的夜间蒸腾和茎干充水等液流行为进行了研究, 分析方位间夜间蒸腾和茎干充水等夜间液流活动的差异及相关性, 以及各方位夜间蒸腾和茎干充水的影响因子, 包括气象因子和白天液流.但是还有很多问题需要进一步研究, 例如已有的实验发现地理位置(
Biophysical controls on nocturnal sap flow in plantation forests in a semi-arid region of northern China
2
2020
... 本实验结果表明, VPD为各方位夜间蒸腾与茎干充水的主要影响因子, 但其对夜间蒸腾的影响更大(
... 本实验对毛白杨不同方位的夜间蒸腾和茎干充水等液流行为进行了研究, 分析方位间夜间蒸腾和茎干充水等夜间液流活动的差异及相关性, 以及各方位夜间蒸腾和茎干充水的影响因子, 包括气象因子和白天液流.但是还有很多问题需要进一步研究, 例如已有的实验发现地理位置(
Interspecific variation in nighttime transpiration and stomatal conductance in a mixed New England deciduous forest
4
2006
... 关于植物夜间蒸腾和茎干充水的发生机制和影响因子目前已有较多研究(
... 将太阳总辐射(Rs)小于5.0 W·m-2的时间段定义为夜间(
... 夜间蒸腾和茎干充水具有不同的生理意义, 因此二者占夜间液流的比例有所不同(
... 本实验结果表明, VPD为各方位夜间蒸腾与茎干充水的主要影响因子, 但其对夜间蒸腾的影响更大(
沙地樟子松边材液流速率的方位差异特征
1
2020
... 关于植物夜间蒸腾和茎干充水的发生机制和影响因子目前已有较多研究(
沙地樟子松边材液流速率的方位差异特征
1
2020
... 关于植物夜间蒸腾和茎干充水的发生机制和影响因子目前已有较多研究(
Nighttime transpiration in woody plants from contrasting ecosystems
2
2007
... 蒸腾是植物消耗水分的主要途径, 在植物生长发育过程中起着至关重要的作用(
... 本实验结果表明, VPD为各方位夜间蒸腾与茎干充水的主要影响因子, 但其对夜间蒸腾的影响更大(
Diurnal and nocturnal transpiration behaviors and their responses to groundwater-table fluctuations and meteorological factors of Populus tomentosa in the North China Plain
2
2019
... 毛白杨(Populus tomentosa)是我国北方重要的防护与用材林树种, 因其优质、速生、丰产、抗逆性强等优点在我国华北地区广泛种植(
... 本实验对毛白杨不同方位的夜间蒸腾和茎干充水等液流行为进行了研究, 分析方位间夜间蒸腾和茎干充水等夜间液流活动的差异及相关性, 以及各方位夜间蒸腾和茎干充水的影响因子, 包括气象因子和白天液流.但是还有很多问题需要进一步研究, 例如已有的实验发现地理位置(
植物夜间液流的发生、生理意义及影响因素研究进展
1
2018
... 蒸腾是植物消耗水分的主要途径, 在植物生长发育过程中起着至关重要的作用(
植物夜间液流的发生、生理意义及影响因素研究进展
1
2018
... 蒸腾是植物消耗水分的主要途径, 在植物生长发育过程中起着至关重要的作用(
Relationships between water availability and selected vessel characteristics in Eucalyptus grandis and two hybrids
1
1995
... 本研究中, 各方位夜间蒸腾量和茎干充水量均具有较大的差异, 其中茎干充水量方位间的差异(CV = 45.6%)大于夜间蒸腾量(CV = 39.2%).而不论夜间蒸腾量还是茎干充水量, 西方位都是优势方位.这些现象可能与环境因子以及树木茎干木质部结构有关.夜间茎干充水主要用于补充木质部中白天蒸腾耗散的水分, 与白天蒸腾量具有显著相关性(
What the towers don’t see at night: nocturnal sap ?ow in trees and shrubs at two AmeriFlux sites in California
7
2007
... 蒸腾是植物消耗水分的主要途径, 在植物生长发育过程中起着至关重要的作用(
... 夜间蒸腾量和茎干充水量的精确估计依赖于夜间液流的精准测定以及夜间蒸腾和茎干充水的有效区分.热技术由于具有精确、不受空间限制、自动化等优点, 被广泛用于树木液流的监测与林分蒸腾的估算(
... 关于植物夜间蒸腾和茎干充水的发生机制和影响因子目前已有较多研究(
... 将太阳总辐射(Rs)小于5.0 W·m-2的时间段定义为夜间(
... 夜间蒸腾和茎干充水具有不同的生理意义, 因此二者占夜间液流的比例有所不同(
... ).有研究表明, 夜间茎干充水是植物夜间用水的主要方式, 约占夜间液流量的50%-70% (
... 较低, 夜间蒸腾也相应减弱, 因此蒸腾量占夜间用水的比例较小.研究表明, 在不同季节、不同地点, 不同树种的夜间蒸腾量和茎干充水量所占的比例有所不同, 比如
A comparison of sap flux-based evapotranspiration estimates with catchment-scale water balance
1
2007
... 夜间蒸腾量和茎干充水量的精确估计依赖于夜间液流的精准测定以及夜间蒸腾和茎干充水的有效区分.热技术由于具有精确、不受空间限制、自动化等优点, 被广泛用于树木液流的监测与林分蒸腾的估算(
A new method of sap flow measurement in tree stems
2
1985
... 茎干液流测定采用热扩散法(
... ), 分别在样木胸高(1.3 m)处, 从东、南、西、北4个方位插入探针, 同时测量树干液流.热扩散探针(TDP 30, Dynamax, Texas, USA)长30 mm, 直径1.2 mm, 探针间距40 mm.试验期间, 液流速率每30 s测定一次, 然后每10 min取平均值并存入数据采集器(Model DL2e, DelTa-T Devices, Cambridge, UK).液流速率计算公式(
Transpiration of trees and forest stands: short and long term monitoring using sapflow methods
1
1996
... 夜间蒸腾量和茎干充水量的精确估计依赖于夜间液流的精准测定以及夜间蒸腾和茎干充水的有效区分.热技术由于具有精确、不受空间限制、自动化等优点, 被广泛用于树木液流的监测与林分蒸腾的估算(
三倍体毛白杨在我国纸浆生产中的地位与作用
1
2002
... 毛白杨(Populus tomentosa)是我国北方重要的防护与用材林树种, 因其优质、速生、丰产、抗逆性强等优点在我国华北地区广泛种植(
三倍体毛白杨在我国纸浆生产中的地位与作用
1
2002
... 毛白杨(Populus tomentosa)是我国北方重要的防护与用材林树种, 因其优质、速生、丰产、抗逆性强等优点在我国华北地区广泛种植(
三倍体毛白杨不同方位树干边材液流特性研究
3
2010
... 毛白杨(Populus tomentosa)是我国北方重要的防护与用材林树种, 因其优质、速生、丰产、抗逆性强等优点在我国华北地区广泛种植(
... 本研究中, 所有方位的夜间液流量都表现出显著的相关性(p < 0.05)(
... 除了相关性, 实验结果表明, 各方位夜间液流量也存在差异, 其可能的原因主要有两点.(1)样树本身的结构造成了树干液流方位上的差异.
三倍体毛白杨不同方位树干边材液流特性研究
3
2010
... 毛白杨(Populus tomentosa)是我国北方重要的防护与用材林树种, 因其优质、速生、丰产、抗逆性强等优点在我国华北地区广泛种植(
... 本研究中, 所有方位的夜间液流量都表现出显著的相关性(p < 0.05)(
... 除了相关性, 实验结果表明, 各方位夜间液流量也存在差异, 其可能的原因主要有两点.(1)样树本身的结构造成了树干液流方位上的差异.
宽窄行栽植下毛白杨不同方位树干液流的差异
1
2018
... 为消除林木个体大小可能对树干液流速率产生的影响, 利用胸径(DBH)对液流速率进行矫正, 公式(
宽窄行栽植下毛白杨不同方位树干液流的差异
1
2018
... 为消除林木个体大小可能对树干液流速率产生的影响, 利用胸径(DBH)对液流速率进行矫正, 公式(
Spatial variations in xylem sap flux density in the trunk of orchard-grown, mature mango trees under changing soil water conditions
1
2000
... 除了相关性, 实验结果表明, 各方位夜间液流量也存在差异, 其可能的原因主要有两点.(1)样树本身的结构造成了树干液流方位上的差异.
基于试验观测的光合有效辐射特征分析
1
2007
... 本研究中, 各方位夜间蒸腾量和茎干充水量均具有较大的差异, 其中茎干充水量方位间的差异(CV = 45.6%)大于夜间蒸腾量(CV = 39.2%).而不论夜间蒸腾量还是茎干充水量, 西方位都是优势方位.这些现象可能与环境因子以及树木茎干木质部结构有关.夜间茎干充水主要用于补充木质部中白天蒸腾耗散的水分, 与白天蒸腾量具有显著相关性(
基于试验观测的光合有效辐射特征分析
1
2007
... 本研究中, 各方位夜间蒸腾量和茎干充水量均具有较大的差异, 其中茎干充水量方位间的差异(CV = 45.6%)大于夜间蒸腾量(CV = 39.2%).而不论夜间蒸腾量还是茎干充水量, 西方位都是优势方位.这些现象可能与环境因子以及树木茎干木质部结构有关.夜间茎干充水主要用于补充木质部中白天蒸腾耗散的水分, 与白天蒸腾量具有显著相关性(
Within-tree variation in transpiration in isolated evergreen oak trees: evidence in support of the pipe model theory
1
2001
... 此外, 以前的研究也表明气象因子也能够影响树干木质部空间上的液流差异.比如,
The ecological and functional correlates of nocturnal transpiration
1
2007
... 蒸腾是植物消耗水分的主要途径, 在植物生长发育过程中起着至关重要的作用(
Can decreased transpiration limit plant nitrogen acquisition in elevated CO2?
1
2002
... 关于植物夜间蒸腾和茎干充水的发生机制和影响因子目前已有较多研究(
Stomatal responses to illumination
1
1965
... 蒸腾是植物消耗水分的主要途径, 在植物生长发育过程中起着至关重要的作用(
Effect of carbohydrates and night temperature on night respiration in rice
1
2015
... 蒸腾是植物消耗水分的主要途径, 在植物生长发育过程中起着至关重要的作用(
Variation in the radial patterns of sap flux density in pubescent oak (Quercus pubescens) and its implications for tree and stand transpiration measurements
2
2007
... 本研究中, 各方位夜间蒸腾量和茎干充水量均具有较大的差异, 其中茎干充水量方位间的差异(CV = 45.6%)大于夜间蒸腾量(CV = 39.2%).而不论夜间蒸腾量还是茎干充水量, 西方位都是优势方位.这些现象可能与环境因子以及树木茎干木质部结构有关.夜间茎干充水主要用于补充木质部中白天蒸腾耗散的水分, 与白天蒸腾量具有显著相关性(
... 此外, 以前的研究也表明气象因子也能够影响树干木质部空间上的液流差异.比如,
Atmospheric and soil water in?uences on the plant water balance
1
1974
... 蒸腾是植物消耗水分的主要途径, 在植物生长发育过程中起着至关重要的作用(
Night-time conductance in C3 and C4 species: Do plants lose water at night?
2
2003
... 本实验结果表明, VPD为各方位夜间蒸腾与茎干充水的主要影响因子, 但其对夜间蒸腾的影响更大(
... 结果一致.
银杏树干茎流变化及其对抑制蒸腾措施的响应
1
2006
... 关于植物夜间蒸腾和茎干充水的发生机制和影响因子目前已有较多研究(
银杏树干茎流变化及其对抑制蒸腾措施的响应
1
2006
... 关于植物夜间蒸腾和茎干充水的发生机制和影响因子目前已有较多研究(
Spatial variations in xylem sap flux density in evergreen oak trees with radial-porous wood: comparisons with anatomical observations
1
2008
... 关于植物夜间蒸腾和茎干充水的发生机制和影响因子目前已有较多研究(
Spatial variation in sap flow velocity in semiarid region trees: its impact on stand-scale transpiration estimates
2
2012
... 关于植物夜间蒸腾和茎干充水的发生机制和影响因子目前已有较多研究(
... 此外, 以前的研究也表明气象因子也能够影响树干木质部空间上的液流差异.比如,
Azimuthal variations of sap flux density within Japanese cypress xylem trunks and their effects on tree transpiration estimates
3
2010
... 蒸腾是植物消耗水分的主要途径, 在植物生长发育过程中起着至关重要的作用(
... 关于植物夜间蒸腾和茎干充水的发生机制和影响因子目前已有较多研究(
... 除了相关性, 实验结果表明, 各方位夜间液流量也存在差异, 其可能的原因主要有两点.(1)样树本身的结构造成了树干液流方位上的差异.
Patterns of water movement in trees and shrubs
1
1972
... 本研究中, 所有方位的夜间液流量都表现出显著的相关性(p < 0.05)(
Nighttime sap flow of Acacia mangium and its implications for nighttime transpiration and stem water storage
4
2012
... 蒸腾是植物消耗水分的主要途径, 在植物生长发育过程中起着至关重要的作用(
... 本研究中, 各方位夜间蒸腾量和茎干充水量均具有较大的差异, 其中茎干充水量方位间的差异(CV = 45.6%)大于夜间蒸腾量(CV = 39.2%).而不论夜间蒸腾量还是茎干充水量, 西方位都是优势方位.这些现象可能与环境因子以及树木茎干木质部结构有关.夜间茎干充水主要用于补充木质部中白天蒸腾耗散的水分, 与白天蒸腾量具有显著相关性(
... 夜间蒸腾和茎干充水具有不同的生理意义, 因此二者占夜间液流的比例有所不同(
... 本实验结果表明, VPD为各方位夜间蒸腾与茎干充水的主要影响因子, 但其对夜间蒸腾的影响更大(
侧柏树干边材液流的空间变化规律及其相关因子
1
2006
... 关于植物夜间蒸腾和茎干充水的发生机制和影响因子目前已有较多研究(
侧柏树干边材液流的空间变化规律及其相关因子
1
2006
... 关于植物夜间蒸腾和茎干充水的发生机制和影响因子目前已有较多研究(
A comparison of methods for determining forest evapotranspiration and its components: sap-flow, soil water budget, eddy covariance and catchment water balance
1
2001
... 夜间蒸腾量和茎干充水量的精确估计依赖于夜间液流的精准测定以及夜间蒸腾和茎干充水的有效区分.热技术由于具有精确、不受空间限制、自动化等优点, 被广泛用于树木液流的监测与林分蒸腾的估算(
Modeling stand water use response to soil water availability and groundwater level for a mature Populus tomentosa plantation located on the North China Plain
1
2017
... 单株样木整日的耗水量计算公式(
Consequences of nocturnal water loss: a synthesis of regulating factors and implications for capacitance, embolism and use in models
5
2014
... 蒸腾是植物消耗水分的主要途径, 在植物生长发育过程中起着至关重要的作用(
... ;
... ;
... 关于植物夜间蒸腾和茎干充水的发生机制和影响因子目前已有较多研究(
... 本实验对毛白杨不同方位的夜间蒸腾和茎干充水等液流行为进行了研究, 分析方位间夜间蒸腾和茎干充水等夜间液流活动的差异及相关性, 以及各方位夜间蒸腾和茎干充水的影响因子, 包括气象因子和白天液流.但是还有很多问题需要进一步研究, 例如已有的实验发现地理位置(
北京山区元宝枫夜间液流活动特征及影响因素
1
2019
... 夜间蒸腾和茎干充水具有不同的生理意义, 因此二者占夜间液流的比例有所不同(
北京山区元宝枫夜间液流活动特征及影响因素
1
2019
... 夜间蒸腾和茎干充水具有不同的生理意义, 因此二者占夜间液流的比例有所不同(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}