Response of leaf litter decomposition of different tree species to nitrogen addition in a subtropical forest
Si-Lu CHEN,1,2, Jin-Song CAI4, Cheng-Fang LIN,1,2,3,*, Hao-Wei SONG1,2, Yu-Sheng YANG1,2,31School of Geographical Sciences, Fujian Normal University, Fuzhou 350007, China 2State Key Laboratory of Humid Subtropical Mountain Ecology, Fuzhou, 350007, China 3Sanming Research Station of Forest Ecosystem and Global Change, Sanming, Fujian 365000, China 4State-owned Forest Farm in the Suburbs of Sanming, Fujian Province, Sanming, Fujian 365000, China
National Natural Science Foundation of China(31770663) Natural Science Foundation of Fujian(2018J01718)
摘要 为探究不同质量凋落物对氮(N)沉降的响应, 该研究采用尼龙网袋分解法, 在亚热带福建三明格氏栲(Castanopsis kawakamii)自然保护区的米槠(Castanopsis carlesii)天然林, 选取4种本区常见的具有不同初始化学性质的树种凋落叶进行模拟N沉降(N添加)分解实验(施N水平为对照0和50 kg·hm -2·a -1)。研究结果表明: 在2年的分解期内, 对照处理的各树种凋落叶的分解速率依次为观光木(Michelia odora, 0.557 a -1)、米槠(0.440 a -1)、台湾相思(Acacia confusa, 0.357 a -1)、杉木(Cunninghamia lanceolata, 0.354 a -1); N添加处理凋落叶分解速率依次为观光木(0.447 a -1)、米槠(0.354 a -1)、杉木(0.291 a -1)、台湾相思(0.230 a -1), 除杉木凋落叶外, N添加显著降低了其他3种凋落叶分解速率。N添加不仅使4种树木凋落叶分解过程中的N释放减慢, 同时还抑制凋落叶化学组成中木质素和纤维素的降解; N添加在凋落叶分解过程中总体上提高β-葡萄糖苷酶(βG)和酸性磷酸酶活性, 对纤维素水解酶的活性影响不一致, 而降低β-N-乙酰氨基葡萄糖苷酶活性和酚氧化酶活性。凋落叶分解速率与凋落叶中的碳获取酶(βG)活性以及其化学组分中的可萃取物含量极显著正相关, 与初始碳浓度、纤维素和木质素含量极显著负相关, 与初始N含量没有显著相关性。凋落物类型和N添加的交互作用虽未影响干质量损失速率, 但对木质素和纤维素的降解具有显著效应。综上所述, 化学组分比初始N含量能更好地预测凋落叶分解速率, 而N添加主要通过抑制分解木质素的氧化酶(如PHO)来降低凋落叶分解速率。 关键词:凋落叶分解;氮沉降;酶活性;亚热带森林
Abstract Aims Litter decomposition plays a vital role in nutrient recycling of forest ecosystems. The decomposition rate of leaves can vary among tree species with different substrate quality and environmental conditions, such as the supply of exogenous nitrogen (N). However, the effects of exogenous N on leaf litter decomposition of different tree species in subtropical forests with high nitrogen deposition background remain poorly understood. Thus this study was designed to address the effect of N addition on litter decomposition of different tree species in a subtropical forest ecosystem. Methods Leaf litters of four common tree species with contrasting substrate quality were collected and decomposed in fertilized (50 kg N·hm -2·a -1) and control (0 kg N·hm -2·a -1) plots for up to two years by using the nylon bag method, in Sanming Castanopsis kawakamii nature reserve of Fujian Province. Important findings The litter decomposition rate in control plots ranked as follows: Michelia odora (0.557 a -1), Castanopsis carlesii (0.440 a -1), Acacia confusa (0.357 a -1), Cunninghamia lanceolata (0.354 a -1), while the decomposition rate in N addition plots ranked as follows: Michelia odora (0.447 a -1), Castanopsis carlesii (0.354 a -1), Cunninghamia lanceolata (0.291 a -1), Acacia confusa (0.230 a -1). Overall, N addition significantly increased the litter mass remaining of Michelia odora, Acacia confusa and Castanopsis carlesii, but not Cunninghamia lanceolata. N addition not only slowed down the release of N, but also retarded the degradation of lignin and cellulose in the decomposition process. Moreover, N addition increased the activities of β-glucosidase (βG) and acid phosphatase, had species-specific effects on the activity of cellulose hydrolase, and decreased the activity of β-N-acetylglucosaminidase and phenoloxidase (PHO). The litter mass loss rate was positively correlated with the activities of carbon acquiring enzyme (βG) in litter layer and the extractives, negatively correlated with carbon concentration, cellulose and lignin, but did not correlate significantly with the initial N concentration. Further analysis found an interactive effect of litter type and N addition on the degradation of cellulose and lignin, but not on dry mass loss. Overall, our results demonstrated that litter chemical components may be better parameters to predict the decomposition rate of leaf litters than the initial nutrient concentrations, and N addition could decrease leaf litter decomposition by inhibiting oxidase (e.g. PHO). We call for further experiments to involve more species and longer time for revealing the response of leaf litter decomposition and its extracellular enzyme activity to N addition. Keywords:leaf litter decomposition;nitrogen deposition;enzyme activities;subtropical forest
PDF (1403KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 陈思路, 蔡劲松, 林成芳, 宋豪威, 杨玉盛. 亚热带不同树种凋落叶分解对氮添加的响应. 植物生态学报, 2020, 44(3): 214-227. DOI: 10.17521/cjpe.2019.0299 CHEN Si-Lu, CAI Jin-Song, LIN Cheng-Fang, SONG Hao-Wei, YANG Yu-Sheng. Response of leaf litter decomposition of different tree species to nitrogen addition in a subtropical forest. Chinese Journal of Plant Ecology, 2020, 44(3): 214-227. DOI: 10.17521/cjpe.2019.0299
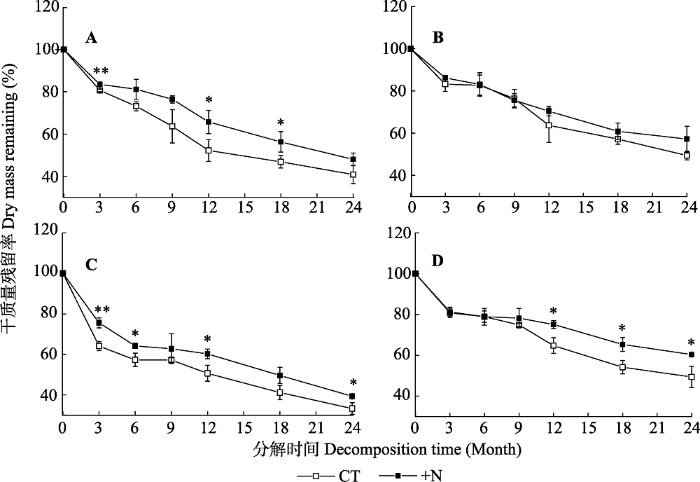
新窗口打开|下载原图ZIP|生成PPT 图1亚热带森林不同处理凋落叶分解过程中质量残留率变化(平均值±标准偏差, n = 3)。A, 米槠凋落叶。B, 杉木凋落叶。C, 观光木凋落叶。D, 台湾相思凋落叶。CT, 对照; +N, 氮添加。*, p < 0.05; **, p < 0.01。
Fig. 1Variations of litter mass remaining of different tree species during decomposition under different treatments in a subtropical forest (mean ± SD, n = 3). A, Castanopsis carlesii litter. B, Cunninghamia lanceolata litter. C, Michelia odora litter. D, Acacia confusa litter. CT, control; +N, nitrogen addition.
Table 3 表3 表3亚热带森林不同处理凋落物分解质量残留率随时间的指数回归方程 Table 3Exponential regression equations of mass remaining rate of leaf litter with time under different treatments in a subtropical forest
树种 Species
处理 Treatment
Olson负指数方程 Olson negative exponential equation
R2
分解常数 Decay constant k (a-1)
年实际干质量 损失率 Annual observed mass loss rate (%)
年预期干质量 损失率 Annual predicted mass loss rate (%)
半分解 时间 T50% (a)
95%分解 时间 T95% (a)
米槠 Castanopsis carlesii
CT
Y = 91.26e-0.440x
0.949
0.440Ba
47.794
42.194
1.292
5.941
+N
Y = 96.35e-0.354x
0.976
0.354Bb
34.257
32.435
1.836
8.232
杉木 Cunninghamia lanceolata
CT
Y = 95.79e-0.354x
0.969
0.354Ca
36.242
30.661
1.862
8.366
+N
Y = 94.91e-0.291x
0.968
0.291BCa
29.510
28.215
2.245
10.167
观光木 Michelia odora
CT
Y = 81.65e-0.557x
0.899
0.557Aa
49.333
38.934
0.993
5.124
+N
Y = 88.03e-0.447x
0.934
0.447Ab
39.768
36.776
1.334
6.481
台湾相思 Acacia confusa
CT
Y = 91.26e-0.357x
0.964
0.357Ca
35.240
31.303
1.798
8.248
+N
Y = 96.35e-0.230x
0.908
0.230Cb
24.887
26.328
2.685
13.479
CT, control; +N, nitrogen addition. T50%, half decay rate; T95%, 95% decay rate. Different lowercase letters denote significant differences among decay constants under different treatments in the same tree species (p < 0.05); different uppercase letters denote significant differences among decay constants in different tree species under the same treatment (p < 0.05). CT, 对照; +N, 氮添加。不同小写字母代表同一树种不同处理之间的分解常数k值具有显著性差异(p < 0.05); 不同大写字母代表同一处理不同树种之间的分解常数k有显著性差异(p < 0.05)。
Table 4 表4 表4亚热带森林凋落叶分解质量损失、木质素、纤维素和氮释放的三因素重复测量方差分析 Table 4Analysis (indicated by F values from ANOVA with three factors repeated measurements) of decomposition time, litter type, nitrogen addition and their interactions on litter mass and lignin, cellulose and N release in a subtropical forest
差异来源 Different source
质量损失率 Mass loss rate
木质素损失率 Lignin loss rate
纤维素损失率 Cellulose loss rate
氮释放率 N release rate
T
356.428***
18 920.622***
8 803.023***
1 266.179***
L
60.641*
1 798.360**
4 305.721***
70.640**
N
287.734**
2 916.781***
5 176.453***
421.188**
T ′ L
4.114
1 605.655***
1 016.727***
14.430*
T ′ N
2.681
124.624**
72.162*
15.670*
L ′ N
8.713
582.266**
667.323***
7.278
T ′ L ′ N
1.038
271.636**
407.487***
2.044
T, decomposition time; L, litter type; N, nitrogen addition. T, 分解时间; L, 凋落物类型; N, 氮添加。*, p < 0.05; **, p < 0.01; ***, p < 0.001。
Fig. 5Litter enzyme activities under different treatments during decomposition in a subtropical forest (mean ± SD, n = 3). A, Castanopsis carlesii litter; B, Cunninghamia lanceolata litter; C, Michelia odora litter; D, Acacia confusa litter. CT, control; +N, nitrogen addition. AP, acid phosphatase; βG, β-1,4-glucosidase; CBH, cellobiohydrolase; NAG, β-1,4-N-acetylglucosaminidase; PEO, peroxidase; PHO, phenol oxidase. Different lowercase letters indicate significant differences among control and nitrogen addition treatments in the same enzyme (p < 0.05).
Table 5 表5 表5亚热带森林凋落物酶活性的双因素方差分析 Table 5Analysis (indicated by F values from ANOVA with two factors) of litter type, nitrogen addition and their interactions on litter enzyme activity in a subtropical forest
差异来源 Different source
βG
CBH
NAG
AP
PHO
PEO
L
23.755***
35.348***
80.072***
82.374***
32.250***
97.990***
N
27.609***
0.040
60.288***
7.380*
4.684*
256.223***
L × N
21.797***
29.083***
149.790***
5.044*
19.714***
15.464***
L, litter type; N, nitrogen addition. AP, acid phosphatase; βG, β-1,4-glucosidase; CBH, cellobiohydrolase; NAG, β-1,4-N-acetylglucosaminidase; PEO, peroxidase; PHO, phenol oxidase. L, 凋落物类型; N, 氮添加。AP, 酸性磷酸酶; βG, β-葡萄糖苷酶; CBH, 纤维素水解酶; NAG, β-N-乙酰氨基葡萄糖苷酶; PEO, 过氧化物酶; PHO, 酚氧化酶;。*, p < 0.05; **, p < 0.01; ***, p < 0.001。
Table 6 表6 表6亚热带森林对照样地凋落物质量损失速率与累积酶活性和初始化学性质的Pearson相关分析(n = 12, 使用平均值) Table 6Pearson correlation coefficients (r) for litter mass loss rate versus cumulative enzyme activity and initial chemistry in the control plots after 2-year decomposition in a subtropical forest (n = 12, mean value used)
AertsR (1997). Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: a triangular relationship Oikos, 79, 439-449. DOI:10.2307/3546886URL [本文引用: 1]
AllisonSD, VitousekPM (2004). Extracellular enzyme activities and carbon chemistry as drivers of tropical plant litter decomposition Biotropica, 36, 285-296. [本文引用: 2]
AponteC, GarcíaLV, Mara?ónT (2012). Tree species effect on litter decomposition and nutrient release in Mediterranean oak forests changes over time Ecosystems, 15, 1204-1218. DOI:10.1007/s10021-012-9577-4URL [本文引用: 1]
AverillC, WaringB (2018). Nitrogen limitation of decomposition and decay: How can it occur? Global Change Biology, 24, 1417-1427. DOI:10.1111/gcb.13980URL [本文引用: 1]
BergB (2000). Litter decomposition and organic matter turnover in northern forest soils Forest Ecology and Management, 133, 13-22. DOI:10.1016/S0378-1127(99)00294-7URL [本文引用: 1]
BergB, DaveyMP, de MarcoA, EmmettB, FaituriM, HobbieSE, JohanssonMB, LiuC, McClaughertyC, NorellL, RutiglianoFA, VesterdalL, de SantoAV (2010). Factors influencing limit values for pine needle litter decomposition: a synthesis for boreal and temperate pine forest systems Biogeochemistry, 100, 57-73. DOI:10.1007/s10533-009-9404-yURL [本文引用: 2]
CornwellWK, CornelissenJHC, KathrynA, EllenD, EvinerVT, OscarG, HobbieSE, BartH, HirokoK, NataliaPH (2008). Plant species traits are the predominant control on litter decomposition rates within biomes worldwide Ecology letters, 11, 1065-1071. DOI:10.1111/ele.2008.11.issue-10URL [本文引用: 1]
CotrufoMF, WallensteinMD, BootCM, DenefK, PaulE (2013). The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Global Change Biology, 19, 988-995. DOI:10.1111/gcb.12113URL [本文引用: 1]
DongLL, SunT, BergB, ZhangLL, ZhangQQ, WangZW (2019). Effects of different forms of N deposition on leaf litter decomposition and extracellular enzyme activities in a temperate grassland Soil Biology & Biochemistry, 134, 78-80. DOI:10.1016/j.soilbio.2019.03.016URL [本文引用: 3]
FangH, MoJM, PengSL, LiZA, WangH (2007). Cumulative effects of nitrogen additions on litter decomposition in three tropical forests in Southern China Plant and Soil, 297, 233-242. DOI:10.1007/s11104-007-9339-9URL [本文引用: 1]
GallowayJN, TownsendAR, ErismanJW, BekundaM, CaiZ, FreneyJR, MartinelliLA, SeitzingerSP, SuttonMA (2008). Transformation of the nitrogen cycle: recent trends, questions, and potential solutions Science, 320, 889-892. DOI:10.1126/science.1136674URL [本文引用: 3]
GuanSY, ZhangDS, ZhangZM (1986). Soil Enzyme and Its Research Method. Agricultural Press, Beijing. [本文引用: 1]
GundersenP, RasmussenL (1990). Nitrification in forest soils: effects from nitrogen deposition on soil acidification and aluminum release Reviews of environmental contamination and toxicology, 113, 1-45. [本文引用: 1]
GüsewellS, GessnerMO (2009). N:P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms Functional Ecology, 23, 211-219. DOI:10.1111/fec.2009.23.issue-1URL [本文引用: 1]
HendricksJJ, AberJD, NadelhofferKJ, HallettRD (2000). Nitrogen controls on fine root substrate quality in temperate forest ecosystems Ecosystems, 3, 57-69. DOI:10.1007/s100210000010URL [本文引用: 2]
HirobeM, SabangJ, BhattaBK, TakedaH (2004). Leaf-litter decomposition of 15 tree species in a lowland tropical rain forest in Sarawak: decomposition rates and initial litter chemistry Journal of Forest Research, 9, 341-346. DOI:10.1007/s10310-004-0087-xURL [本文引用: 2]
HirobeM, SabangJ, BhattaBK, TakedaH (2004). Leaf-litter decomposition of 15 tree species in a lowland tropical rain forest in Sarawak: decomposition rates and initial litter chemistry Journal of Forest Research, 9, 341-346. DOI:10.1007/s10310-004-0087-xURL [本文引用: 1]
HobbieSE, EddyWC, BuyarskiCR, AdairEC, OgdahlML, WeisenhornP (2012). Response of decomposing litter and its microbial community to multiple forms of nitrogen enrichment Ecological Monographs, 82, 389-405. DOI:10.1890/11-1600.1URL [本文引用: 1]
HongHB, LinCF, PengJQ, ChenYM, WeiCC, YangYS (2017). Effects of phosphorus addition on fine root decomposition and enzyme activity of Castanopsis carlesii and Cunninghamia lanceolata in subtropical forest Acta Ecologica Sinica, 37, 136-146. [本文引用: 2]
HuS, ZhangY, ShiRJ, HanSQ, LiH, XuH (2013). Temporal variations of soil microbial biomass and enzyme activities during the secondary succession of primary broadleaved-Pinuskoraiensis forests in Changbai Mountains of Northeast China Chinese Journal of Applied Ecology, 24, 366-372. [本文引用: 1]
KeiserAD, KeiserDA, StricklandMS, BradfordMA (2014). Disentangling the mechanisms underlying functional differences among decomposer communities Journal of Ecology, 102, 603-609. DOI:10.1111/jec.2014.102.issue-3URL [本文引用: 1]
KnorrM, FreySD, CurtisPS (2005). Nitrogen additions and litter decomposition: a meta-analysis Ecology, 86, 3252-3257. DOI:10.1890/05-0150URL [本文引用: 1]
LauberCL, HamadyM, KnightR, FiererN (2009). Pyrosequencing-???based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale Applied and Environmental Microbiology, 75, 5111-5120. DOI:10.1128/AEM.00335-09URL [本文引用: 1]
LiH, WuFZ, YangWQ, XuLY, NiXY, HeJ, ChangCH (2015). Effects of snow cover on acid-soluble extractive and acid-insoluble residue during foliar litter decomposition in the alpine forest Acta Ecologica Sinica, 35, 4687-4698. [本文引用: 1]
LiXF, HanSJ, HuYL, ZhaoYT (2008). Decomposition of litter organic matter and its relations to C, N and P release in secondary conifer and broadleaf mixed forest in Changbai Mountains Chinese Journal of Applied Ecology, 19, 245-251. [本文引用: 1]
LuY, LiK, LiangQ, LiCR, ZhangCH (2019). Effects of leaf litter decomposition on bacterial community structure in the leaf litter of four dominant tree species in Mount Tai Acta Ecologica Sinica, 39, 3175-3186. [本文引用: 2]
MakitaN, FujiiS (2015). Tree species effects on microbial respiration from decomposing leaf and fine root litter Soil Biology & Biochemistry, 88, 39-47. DOI:10.1016/j.soilbio.2015.05.005URL [本文引用: 1]
MelilloJM, AberJD, MuratoreJF (1982). Nitrogen and lignin control of hardwood leaf litter decomposition dynamics Ecology, 63, 621-626. DOI:10.2307/1936780URL [本文引用: 1]
OlsonJS (1963). Energy storage and the balance of producers and decomposers in ecological systems Ecology, 44, 322-331. DOI:10.2307/1932179URL [本文引用: 2]
PalozziJE, LindoZ (2018). Are leaf litter and microbes team players? Interpreting home-field advantage decomposition dynamics Soil Biology & Biochemistry, 124, 189-198. DOI:10.1016/j.soilbio.2018.06.018URL [本文引用: 1]
RamirezKS, CraineJM, FiererN (2010). Nitrogen fertilization inhibits soil microbial respiration regardless of the form of nitrogen applied Soil Biology & Biochemistry, 42, 2336-2338. DOI:10.1016/j.soilbio.2010.08.032URL [本文引用: 1]
RyanMG, MelilloJM, RiccaA (1990). A comparison of methods for determining proximate carbon fractions of forest litter Canadian Journal of Forest Research, 20, 166-171. DOI:10.1139/x90-023URL [本文引用: 1]
Saiya-CorkKR, SinsabaughRL, ZakDR (2002). The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil Soil Biology & Biochemistry, 34, 1309-1315. DOI:10.1016/S0038-0717(02)00074-3URL [本文引用: 2]
SariyildizT, AndersonJM, KucukM (2005). Effects of tree species and topography on soil chemistry, litter quality, and decomposition in Northeast Turkey Soil Biology & Biochemistry, 37, 1695-1706. DOI:10.1016/j.soilbio.2005.02.004URL [本文引用: 2]
SinsabaughRL, CarreiroMM, RepertDA (2002). Allocation of extracellular enzymatic activity in relation to litter composition, N deposition, and mass loss Biogeochemistry, 60, 1-24. DOI:10.1023/A:1016541114786URL [本文引用: 1]
SinsabaughRL, WeilandT, LinkinsAE (1992). Enzymic and molecular analysis of microbial communities associated with lotic particulate organic matter Freshwater Biology, 28, 393-404. DOI:10.1111/fwb.1992.28.issue-3URL [本文引用: 1]
SongY, GuXR, YanHY, MaoWT, WuXL, WanYX (2014). Dynamics of microbes and enzyme activities during litter decomposition of Pinus massoniana forest in mid-?subtropical area Environmental Science, 35, 1151-1158. [本文引用: 1]
TianDS, NiuSL (2015). A global analysis of soil acidification caused by nitrogen addition Environmental Research Letters, 10, 024019. DOI: 10.1088/1748-9326/10/2/024019. DOI:10.1088/1748-9326/10/2/024019URL [本文引用: 1]
TuLH, HuHL, HuTX, ZhangJ, LuoSH, DaiHZ (2012). Response of Betula luminifera leaf litter decomposition to simulated nitrogen deposition in the Rainy Area of West China Chinese Journal of Plant Ecology, 36, 99-108. DOI:10.3724/SP.J.1258.2012.00099URL [本文引用: 4]
van HuysenTL, PerakisSS, HarmonME (2016). Decomposition drives convergence of forest litter nutrient stoichiometry following phosphorus addition Plant and Soil, 406, 1-14. DOI:10.1007/s11104-016-2857-6URL [本文引用: 1]
VitousekPM, AberJD, HowarthRW, LikensGE, MatsonPA, SchindlerDW, SchlesingerWH, TilmanDG (1997). Human alteration of the global nitrogen cycle: sources and consequences Ecological Applications, 7, 737-750. [本文引用: 1]
VivancoL, AustinAT (2011). Nitrogen addition stimulates forest litter decomposition and disrupts species interactions in Patagonia, Argentina Global Change Biology, 17, 1963-1974. DOI:10.1111/j.1365-2486.2010.02344.xURL [本文引用: 1]
WangJJ, ChengY, YangYS, LinCF, LinWS, PengJQ (2014). Responses of leaf litter decomposition and nutrient release to simulated nitrogen deposition of Castanopsis carlesii Journal of Fujian College of Forestry, 34, 113-119. [本文引用: 1]
WaringBG (2013). Exploring relationships between enzyme activities and leaf litter decomposition in a wet tropical forest Soil Biology & Biochemistry, 64, 89-95. DOI:10.1016/j.soilbio.2013.04.010URL [本文引用: 6]
WiederWR, ClevelandCC, TownsendAR (2009). Controls over leaf litter decomposition in wet tropical forests Ecology, 90, 3333-3341. DOI:10.1890/08-2294.1URL [本文引用: 1]
YanJF, WangL, HuY, TsangYF, ZhangYN, WuJH, FuXH, SunY (2018). Plant litter composition selects different soil microbial structures and in turn drives different litter decomposition pattern and soil carbon sequestration capability Geoderma, 319, 194-203. DOI:10.1016/j.geoderma.2018.01.009URL [本文引用: 1]
YangL, DengCC, ChenYM, HeRL, ZhangJ, LiuY (2015). Relationships between decomposition rate of leaf litter and initial quality across the alpine timberline ecotone in Western Sichuan, China Chinese Journal of Applied Ecology, 26, 3602-3610. [本文引用: 1]
ZhangXY, WangW (2015). Control of climate and litter quality on leaf litter decomposition in different climatic zones Journal of Plant Research, 128, 791-802. DOI:10.1007/s10265-015-0743-6URL [本文引用: 1]
ZhengJQ, GuoRH, LiDS, ZhangJH, HanSJ (2017). Nitrogen addition, drought and mixture effects on litter decomposition and nitrogen immobilization in a temperate forest Plant and Soil, 416, 165-179. DOI:10.1007/s11104-017-3202-4URL [本文引用: 1]
ZhengYX, CaoJL, YangZJ, LinCF, YangYS (2018). Impacts of nitrogen deposition on soil microbial community structure in subtropical natural evergreen broad-leaved forest relative to season Acta Pedologica Sinica, 55, 1534-1544. [本文引用: 1]
ZhouSX, HuangCD, XiangYB, HanBH, XiaoYX, TangJD (2016 a). Effects of simulated nitrogen deposition on lignin and cellulose degradation of foliar litter in natural evergreen broad-leaved forest in Rainy Area of Western China Chinese Journal of Applied Ecology, 27, 1368-1374. [本文引用: 2]
ZhouSX, XiaoYX, XiangYB, HuangCD, TangJD, HanBH, LuoC (2016b). Effects of simulated nitrogen deposition on the substrate quality of foliar litter in a natural evergreen broad-leaved forest in the Rainy Area of Western China Acta Ecologica Sinica, 36, 7428-7435. [本文引用: 1]
ZhouXQ, ChenCR, WangYF, XuZH, HanHY, LiLH, WanSQ (2013). Warming and increased precipitation have differential effects on soil extracellular enzyme activities in a temperate grassland Science of The Total Environment, 444, 552-558. DOI:10.1016/j.scitotenv.2012.12.023URL [本文引用: 2]
ZhouZH, WangCK, ZhengMH, JiangLF, LuoYQ (2017). Patterns and mechanisms of responses by soil microbial communities to nitrogen addition Soil Biology & Biochemistry, 115, 433-441 DOI:10.1016/j.soilbio.2017.09.015URL [本文引用: 1]
Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: a triangular relationship 1 1997
... 微生物是凋落物的最终分解者, 凋落物本身的性质和周围土壤环境的长期相互作用影响微生物群落的结构和分解功能(Keiser et al., 2014; van Huysen et al., 2016; Palozzi & Lindo, 2018; Yan et al., 2018).近年来很多研究表明, 胞外酶活性的动态能较好地解释凋落物分解的变化(Waring, 2013; Dong et al., 2019), 如N添加改变了木质素降解酶(酚氧化酶)以及其他水解酶的活性, 从而改变了凋落物的分解速率(Saiya-Cork et al., 2002; Sinsabaugh et al., 2005; Stursova et al., 2006).测定酶活性能够直接跟踪微生物群落对凋落物性质和环境变量的功能响应(Carreiro et al., 2000), 但由于植物-凋落物-微生物交互作用具有潜在的复杂性, 酶活性会对凋落物性质和外源N输入的改变做出何种响应, 目前尚不清楚. ...
Soil acidification exerts a greater control on soil respiration than soil nitrogen availability in grasslands subjected to long-term nitrogen enrichment 1 2016
The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? 1 2013
... 由于化石燃料的燃烧和氮(N)肥的大量生产与使用, 人类活动向大气排放越来越多的含N化合物, 导致大气N沉降增加.排放到大气中的N, 从1860年的34 Tg·a-1增加到1995年的100 Tg·a-1, 预计到2050年会增加到200 Tg·a-1 (Galloway et al., 2008).N沉降可以提高土壤N有效性, 影响微生物群落结构和活性(郑裕雄等, 2018), 进而影响凋落物分解(Fang et al., 2007; Cotrufo et al., 2013), 并通过改变土壤有机质的合成与分解, 影响陆地生态系统碳(C)、N循环(Vitousek et al., 1997; Galloway et al., 2008).以往关于N沉降的研究大多集中在土壤N相对缺乏的温带和北方森林,而近年来开始关注热带、亚热带地区(Zhou et al., 2017; Yu et al., 2018).我国是全球三大严重N沉降地区之一, 尤其以东部地区最为严重(Galloway et al., 2008), 大气N沉降可能严重影响我国亚热带森林生态系统的养分循环. ...
Effects of different forms of N deposition on leaf litter decomposition and extracellular enzyme activities in a temperate grassland 3 2019
... 微生物是凋落物的最终分解者, 凋落物本身的性质和周围土壤环境的长期相互作用影响微生物群落的结构和分解功能(Keiser et al., 2014; van Huysen et al., 2016; Palozzi & Lindo, 2018; Yan et al., 2018).近年来很多研究表明, 胞外酶活性的动态能较好地解释凋落物分解的变化(Waring, 2013; Dong et al., 2019), 如N添加改变了木质素降解酶(酚氧化酶)以及其他水解酶的活性, 从而改变了凋落物的分解速率(Saiya-Cork et al., 2002; Sinsabaugh et al., 2005; Stursova et al., 2006).测定酶活性能够直接跟踪微生物群落对凋落物性质和环境变量的功能响应(Carreiro et al., 2000), 但由于植物-凋落物-微生物交互作用具有潜在的复杂性, 酶活性会对凋落物性质和外源N输入的改变做出何种响应, 目前尚不清楚. ...
Cumulative effects of nitrogen additions on litter decomposition in three tropical forests in Southern China 1 2007
... 由于化石燃料的燃烧和氮(N)肥的大量生产与使用, 人类活动向大气排放越来越多的含N化合物, 导致大气N沉降增加.排放到大气中的N, 从1860年的34 Tg·a-1增加到1995年的100 Tg·a-1, 预计到2050年会增加到200 Tg·a-1 (Galloway et al., 2008).N沉降可以提高土壤N有效性, 影响微生物群落结构和活性(郑裕雄等, 2018), 进而影响凋落物分解(Fang et al., 2007; Cotrufo et al., 2013), 并通过改变土壤有机质的合成与分解, 影响陆地生态系统碳(C)、N循环(Vitousek et al., 1997; Galloway et al., 2008).以往关于N沉降的研究大多集中在土壤N相对缺乏的温带和北方森林,而近年来开始关注热带、亚热带地区(Zhou et al., 2017; Yu et al., 2018).我国是全球三大严重N沉降地区之一, 尤其以东部地区最为严重(Galloway et al., 2008), 大气N沉降可能严重影响我国亚热带森林生态系统的养分循环. ...
Transformation of the nitrogen cycle: recent trends, questions, and potential solutions 3 2008
... 由于化石燃料的燃烧和氮(N)肥的大量生产与使用, 人类活动向大气排放越来越多的含N化合物, 导致大气N沉降增加.排放到大气中的N, 从1860年的34 Tg·a-1增加到1995年的100 Tg·a-1, 预计到2050年会增加到200 Tg·a-1 (Galloway et al., 2008).N沉降可以提高土壤N有效性, 影响微生物群落结构和活性(郑裕雄等, 2018), 进而影响凋落物分解(Fang et al., 2007; Cotrufo et al., 2013), 并通过改变土壤有机质的合成与分解, 影响陆地生态系统碳(C)、N循环(Vitousek et al., 1997; Galloway et al., 2008).以往关于N沉降的研究大多集中在土壤N相对缺乏的温带和北方森林,而近年来开始关注热带、亚热带地区(Zhou et al., 2017; Yu et al., 2018).我国是全球三大严重N沉降地区之一, 尤其以东部地区最为严重(Galloway et al., 2008), 大气N沉降可能严重影响我国亚热带森林生态系统的养分循环. ...
... ; Galloway et al., 2008).以往关于N沉降的研究大多集中在土壤N相对缺乏的温带和北方森林,而近年来开始关注热带、亚热带地区(Zhou et al., 2017; Yu et al., 2018).我国是全球三大严重N沉降地区之一, 尤其以东部地区最为严重(Galloway et al., 2008), 大气N沉降可能严重影响我国亚热带森林生态系统的养分循环. ...
... ).我国是全球三大严重N沉降地区之一, 尤其以东部地区最为严重(Galloway et al., 2008), 大气N沉降可能严重影响我国亚热带森林生态系统的养分循环. ...
Disentangling the mechanisms underlying functional differences among decomposer communities 1 2014
... 微生物是凋落物的最终分解者, 凋落物本身的性质和周围土壤环境的长期相互作用影响微生物群落的结构和分解功能(Keiser et al., 2014; van Huysen et al., 2016; Palozzi & Lindo, 2018; Yan et al., 2018).近年来很多研究表明, 胞外酶活性的动态能较好地解释凋落物分解的变化(Waring, 2013; Dong et al., 2019), 如N添加改变了木质素降解酶(酚氧化酶)以及其他水解酶的活性, 从而改变了凋落物的分解速率(Saiya-Cork et al., 2002; Sinsabaugh et al., 2005; Stursova et al., 2006).测定酶活性能够直接跟踪微生物群落对凋落物性质和环境变量的功能响应(Carreiro et al., 2000), 但由于植物-凋落物-微生物交互作用具有潜在的复杂性, 酶活性会对凋落物性质和外源N输入的改变做出何种响应, 目前尚不清楚. ...
Nitrogen additions and litter decomposition: a meta-analysis 1 2005
Are leaf litter and microbes team players? Interpreting home-field advantage decomposition dynamics 1 2018
... 微生物是凋落物的最终分解者, 凋落物本身的性质和周围土壤环境的长期相互作用影响微生物群落的结构和分解功能(Keiser et al., 2014; van Huysen et al., 2016; Palozzi & Lindo, 2018; Yan et al., 2018).近年来很多研究表明, 胞外酶活性的动态能较好地解释凋落物分解的变化(Waring, 2013; Dong et al., 2019), 如N添加改变了木质素降解酶(酚氧化酶)以及其他水解酶的活性, 从而改变了凋落物的分解速率(Saiya-Cork et al., 2002; Sinsabaugh et al., 2005; Stursova et al., 2006).测定酶活性能够直接跟踪微生物群落对凋落物性质和环境变量的功能响应(Carreiro et al., 2000), 但由于植物-凋落物-微生物交互作用具有潜在的复杂性, 酶活性会对凋落物性质和外源N输入的改变做出何种响应, 目前尚不清楚. ...
Nitrogen fertilization inhibits soil microbial respiration regardless of the form of nitrogen applied 1 2010
A comparison of methods for determining proximate carbon fractions of forest litter 1 1990
... 将烘干后凋落物样品粉碎后用碳氮元素分析仪(Vario ELIII, Elementar, Hanau, Germany)测定凋落叶的C、N含量.凋落叶P含量采用HClO4-H2SO4消煮法, 取上清液后使用连续流动分析仪(San++, Skalar, Breda, Netherlands)测定.用近似系统分析法测定纤维素和木质素含量(Ryan et al., 1990; Hendricks et al., 2000). ...
The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil 2 2002
... 微生物是凋落物的最终分解者, 凋落物本身的性质和周围土壤环境的长期相互作用影响微生物群落的结构和分解功能(Keiser et al., 2014; van Huysen et al., 2016; Palozzi & Lindo, 2018; Yan et al., 2018).近年来很多研究表明, 胞外酶活性的动态能较好地解释凋落物分解的变化(Waring, 2013; Dong et al., 2019), 如N添加改变了木质素降解酶(酚氧化酶)以及其他水解酶的活性, 从而改变了凋落物的分解速率(Saiya-Cork et al., 2002; Sinsabaugh et al., 2005; Stursova et al., 2006).测定酶活性能够直接跟踪微生物群落对凋落物性质和环境变量的功能响应(Carreiro et al., 2000), 但由于植物-凋落物-微生物交互作用具有潜在的复杂性, 酶活性会对凋落物性质和外源N输入的改变做出何种响应, 目前尚不清楚. ...
Extracellular enzyme activities and soil organic matter dynamics for northern hardwood forests receiving simulated nitrogen deposition 1 2005
... 微生物是凋落物的最终分解者, 凋落物本身的性质和周围土壤环境的长期相互作用影响微生物群落的结构和分解功能(Keiser et al., 2014; van Huysen et al., 2016; Palozzi & Lindo, 2018; Yan et al., 2018).近年来很多研究表明, 胞外酶活性的动态能较好地解释凋落物分解的变化(Waring, 2013; Dong et al., 2019), 如N添加改变了木质素降解酶(酚氧化酶)以及其他水解酶的活性, 从而改变了凋落物的分解速率(Saiya-Cork et al., 2002; Sinsabaugh et al., 2005; Stursova et al., 2006).测定酶活性能够直接跟踪微生物群落对凋落物性质和环境变量的功能响应(Carreiro et al., 2000), 但由于植物-凋落物-微生物交互作用具有潜在的复杂性, 酶活性会对凋落物性质和外源N输入的改变做出何种响应, 目前尚不清楚. ...
Enzymic and molecular analysis of microbial communities associated with lotic particulate organic matter 1 1992
Microbial responses to long-term N deposition in a semiarid grassland 1 2006
... 微生物是凋落物的最终分解者, 凋落物本身的性质和周围土壤环境的长期相互作用影响微生物群落的结构和分解功能(Keiser et al., 2014; van Huysen et al., 2016; Palozzi & Lindo, 2018; Yan et al., 2018).近年来很多研究表明, 胞外酶活性的动态能较好地解释凋落物分解的变化(Waring, 2013; Dong et al., 2019), 如N添加改变了木质素降解酶(酚氧化酶)以及其他水解酶的活性, 从而改变了凋落物的分解速率(Saiya-Cork et al., 2002; Sinsabaugh et al., 2005; Stursova et al., 2006).测定酶活性能够直接跟踪微生物群落对凋落物性质和环境变量的功能响应(Carreiro et al., 2000), 但由于植物-凋落物-微生物交互作用具有潜在的复杂性, 酶活性会对凋落物性质和外源N输入的改变做出何种响应, 目前尚不清楚. ...
Decomposition drives convergence of forest litter nutrient stoichiometry following phosphorus addition 1 2016
... 微生物是凋落物的最终分解者, 凋落物本身的性质和周围土壤环境的长期相互作用影响微生物群落的结构和分解功能(Keiser et al., 2014; van Huysen et al., 2016; Palozzi & Lindo, 2018; Yan et al., 2018).近年来很多研究表明, 胞外酶活性的动态能较好地解释凋落物分解的变化(Waring, 2013; Dong et al., 2019), 如N添加改变了木质素降解酶(酚氧化酶)以及其他水解酶的活性, 从而改变了凋落物的分解速率(Saiya-Cork et al., 2002; Sinsabaugh et al., 2005; Stursova et al., 2006).测定酶活性能够直接跟踪微生物群落对凋落物性质和环境变量的功能响应(Carreiro et al., 2000), 但由于植物-凋落物-微生物交互作用具有潜在的复杂性, 酶活性会对凋落物性质和外源N输入的改变做出何种响应, 目前尚不清楚. ...
Human alteration of the global nitrogen cycle: sources and consequences 1 1997
... 由于化石燃料的燃烧和氮(N)肥的大量生产与使用, 人类活动向大气排放越来越多的含N化合物, 导致大气N沉降增加.排放到大气中的N, 从1860年的34 Tg·a-1增加到1995年的100 Tg·a-1, 预计到2050年会增加到200 Tg·a-1 (Galloway et al., 2008).N沉降可以提高土壤N有效性, 影响微生物群落结构和活性(郑裕雄等, 2018), 进而影响凋落物分解(Fang et al., 2007; Cotrufo et al., 2013), 并通过改变土壤有机质的合成与分解, 影响陆地生态系统碳(C)、N循环(Vitousek et al., 1997; Galloway et al., 2008).以往关于N沉降的研究大多集中在土壤N相对缺乏的温带和北方森林,而近年来开始关注热带、亚热带地区(Zhou et al., 2017; Yu et al., 2018).我国是全球三大严重N沉降地区之一, 尤其以东部地区最为严重(Galloway et al., 2008), 大气N沉降可能严重影响我国亚热带森林生态系统的养分循环. ...
Nitrogen addition stimulates forest litter decomposition and disrupts species interactions in Patagonia, Argentina 1 2011
... 试验地设在福建省三明格氏栲(Castanopsis kawakamii)自然保护区(26.17o N, 117.45o E)内, 该研究区平均海拔300 m, 地形以低山丘陵为主.属亚热带季风气候, 多年平均气温19.1 ℃, 年降水量1 749 mm.土壤主要为花岗岩或砂岩发育的酸性红壤, 土层厚度一般超过1 m.实验区为近200年无人干扰的米槠天然林, 样地海拔为315 m, 坡向NE 25°, 坡度35°, 土壤为黑云母花岗岩发育的红壤.优势种为米槠, 其他树种有木荷(Schima superba)、格氏栲、马尾松(Pinus massoniana)、虎皮楠(Daphnipnhyllum oldhamii)、桂北木姜子(Litsea subcoriacea)、杜英(Elaeocarpus decipiens)、毛冬青(Ilex pubescens)等.群落分层明显, 可划分为乔木层、灌木层和草本层3个层次.林下地被层较厚, 并且散布有枯立木、倒木和死树枝杆等, 枯枝落叶厚度为5-8 cm (林伟盛等, 2013).林分详细概况参见表1 (王家骏等, 2014). ...
米槠叶凋落物分解及养分释放对模拟N沉降的响应 1 2014
... 试验地设在福建省三明格氏栲(Castanopsis kawakamii)自然保护区(26.17o N, 117.45o E)内, 该研究区平均海拔300 m, 地形以低山丘陵为主.属亚热带季风气候, 多年平均气温19.1 ℃, 年降水量1 749 mm.土壤主要为花岗岩或砂岩发育的酸性红壤, 土层厚度一般超过1 m.实验区为近200年无人干扰的米槠天然林, 样地海拔为315 m, 坡向NE 25°, 坡度35°, 土壤为黑云母花岗岩发育的红壤.优势种为米槠, 其他树种有木荷(Schima superba)、格氏栲、马尾松(Pinus massoniana)、虎皮楠(Daphnipnhyllum oldhamii)、桂北木姜子(Litsea subcoriacea)、杜英(Elaeocarpus decipiens)、毛冬青(Ilex pubescens)等.群落分层明显, 可划分为乔木层、灌木层和草本层3个层次.林下地被层较厚, 并且散布有枯立木、倒木和死树枝杆等, 枯枝落叶厚度为5-8 cm (林伟盛等, 2013).林分详细概况参见表1 (王家骏等, 2014). ...
Exploring relationships between enzyme activities and leaf litter decomposition in a wet tropical forest 6 2013
... 微生物是凋落物的最终分解者, 凋落物本身的性质和周围土壤环境的长期相互作用影响微生物群落的结构和分解功能(Keiser et al., 2014; van Huysen et al., 2016; Palozzi & Lindo, 2018; Yan et al., 2018).近年来很多研究表明, 胞外酶活性的动态能较好地解释凋落物分解的变化(Waring, 2013; Dong et al., 2019), 如N添加改变了木质素降解酶(酚氧化酶)以及其他水解酶的活性, 从而改变了凋落物的分解速率(Saiya-Cork et al., 2002; Sinsabaugh et al., 2005; Stursova et al., 2006).测定酶活性能够直接跟踪微生物群落对凋落物性质和环境变量的功能响应(Carreiro et al., 2000), 但由于植物-凋落物-微生物交互作用具有潜在的复杂性, 酶活性会对凋落物性质和外源N输入的改变做出何种响应, 目前尚不清楚. ...
Plant litter composition selects different soil microbial structures and in turn drives different litter decomposition pattern and soil carbon sequestration capability 1 2018
... 微生物是凋落物的最终分解者, 凋落物本身的性质和周围土壤环境的长期相互作用影响微生物群落的结构和分解功能(Keiser et al., 2014; van Huysen et al., 2016; Palozzi & Lindo, 2018; Yan et al., 2018).近年来很多研究表明, 胞外酶活性的动态能较好地解释凋落物分解的变化(Waring, 2013; Dong et al., 2019), 如N添加改变了木质素降解酶(酚氧化酶)以及其他水解酶的活性, 从而改变了凋落物的分解速率(Saiya-Cork et al., 2002; Sinsabaugh et al., 2005; Stursova et al., 2006).测定酶活性能够直接跟踪微生物群落对凋落物性质和环境变量的功能响应(Carreiro et al., 2000), 但由于植物-凋落物-微生物交互作用具有潜在的复杂性, 酶活性会对凋落物性质和外源N输入的改变做出何种响应, 目前尚不清楚. ...
 ,1,2, 蔡劲松4, 林成芳
,1,2, 蔡劲松4, 林成芳 ,1,2,3,*, 宋豪威1,2, 杨玉盛1,2,3
,1,2,3,*, 宋豪威1,2, 杨玉盛1,2,3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT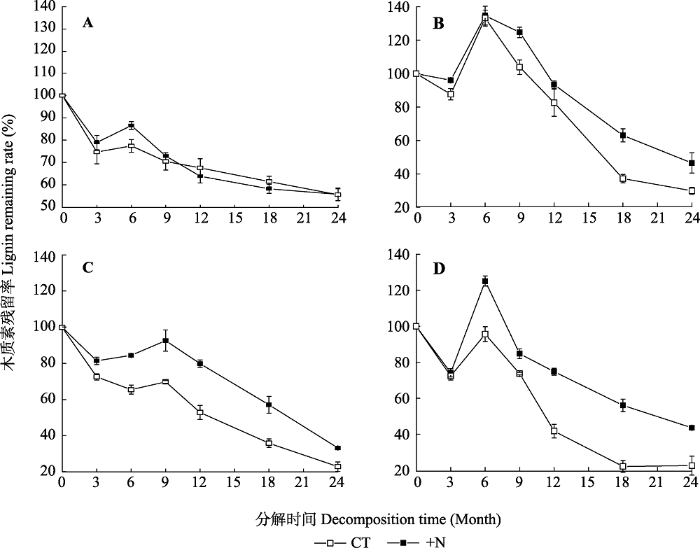 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT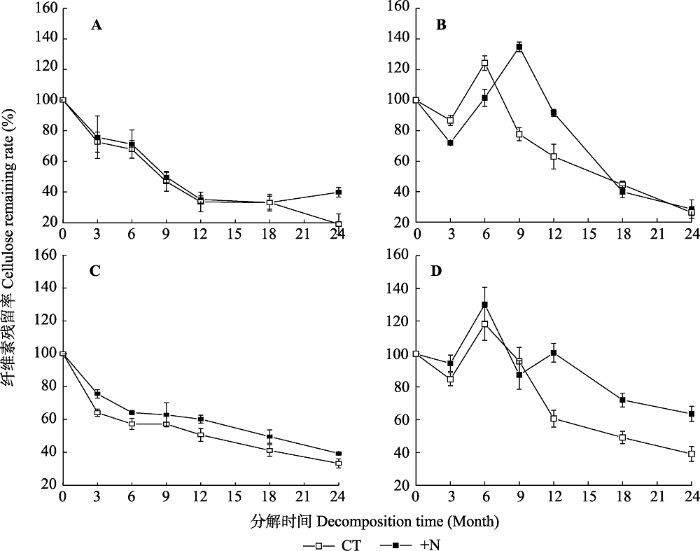 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT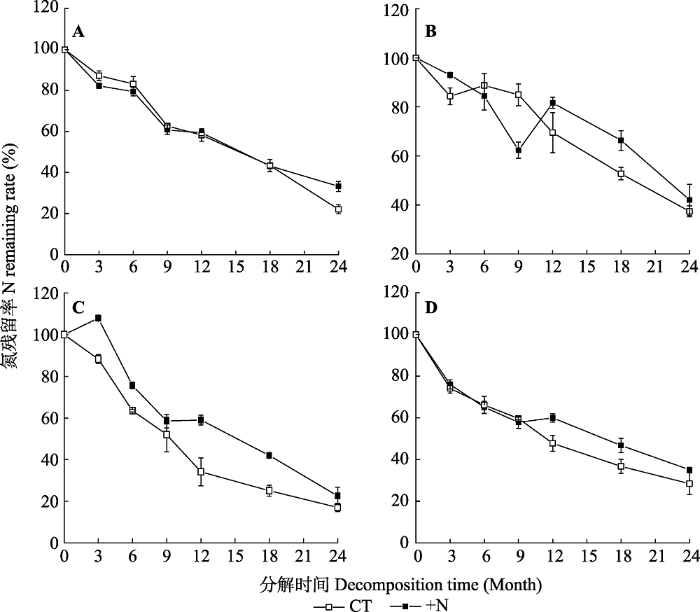 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT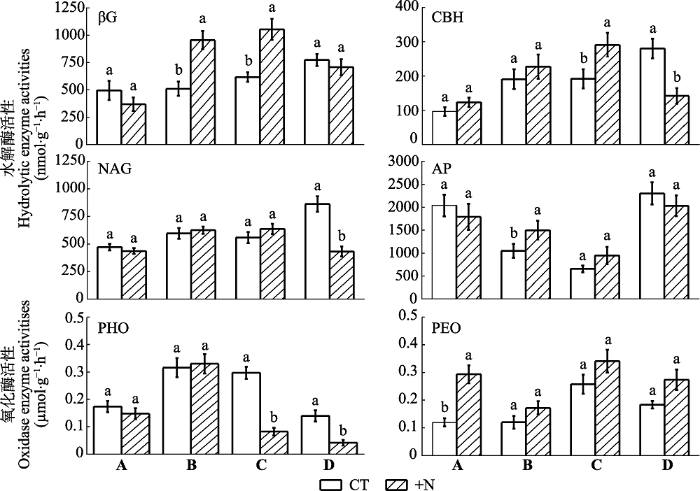 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}