祝维1, 余立璇1, 赵德海2, 贾黎明,,1,*1 北京林业大学省部共建森林培育与保护教育部重点实验室, 北京 100083 2 Warnell School of Forestry and Natural Resources, The University of Georgia, Athens, GA 30602, USA
Architectural analysis of root systems of mature trees in sandy loam soils using the root development classification
Wei ZHU1, Li-Xuan YU1, De-Hai ZHAO2, Li-Ming JIA,,1,*1 Ministry of Education Key Laboratory of Silviculture and Conservation, Beijing Forestry University, Beijing 100083, China 2 Warnell School of Forestry and Natural Resources, The University of Georgia, Athens, GA 30602, USA
Abstract Aims Understanding the differences in root architectural strategies among the species and the differences in morphological characteristics among different root orders will facilitate our understanding root growth and development strategies, and thus provide a basis for predicting and modeling the root systems for mature trees. In this study, we analyzed the morphological characteristics and topological relationships for the root systems of two Populus tomentosa trees and one Robinia pseudoacacia tree. Methods A method combining both excavation and analysis was applied to extract and quantify root architectural characteristics of the three root systems. The morphological characteristics such as root basal diameter, root length, link length, and root number of different root orders were described using the developmental analysis method of Rose (1983), and their topological relationships were analyzed. Important findings 1) The modified topological indices qa and qb were close to 0, and the topological index TI was close to 0.5 for all three root systems, indicating their dichotomous structure. The depth and width of the systems ranged from 5.7 to 6.4 m and from 7.6 to 13.5 m, respectively. Root grafts occurred in the same species. 2) The root systems could have the seventh or eighth order roots. The basal root diameter and root length significantly decreased with increasing root order. The first order roots had 5.79-36.92 times the basal diameter and 1.45-9.11 times the length of higher order roots. With increasing root order, the root number increased, and reached a maximum value for the third order roots, and then decreased. 3) In roots of each of the first three orders, the link length varied little from the root base towards its tip, indicating that the child roots were distributed evenly on their parent roots and thus help trees absorb soil resources more efficiently. 4) The regression of basal diameters of child roots on basal diameters of their mother roots showed that the smallest slope for the first order roots (average slope 0.15) and no big difference in the slope between the second and third order roots (0.34 versus 0.35). This suggested that the first order roots developed their own diameter first for anchoring and supporting the tree, while the second and third order roots developed their child roots to facilitate nutrient uptake from the soil. 5) The regression of root length on root basal diameter suggested that the slope increased from 10.46 to 90.43 with increasing root order, which implies that the higher order roots tended to develop their length to explore resources and expand their space. Keywords:developmental classification standard;root system classification;root architecture;mature tree;sandy loam soil
PDF (1713KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 祝维, 余立璇, 赵德海, 贾黎明. 基于根系发育分级的砂壤土下成熟林木根系构型分析. 植物生态学报, 2019, 43(2): 119-130. DOI: 10.17521/cjpe.2018.0269 ZHU Wei, YU Li-Xuan, ZHAO De-Hai, JIA Li-Ming. Architectural analysis of root systems of mature trees in sandy loam soils using the root development classification. Chinese Journal of Plant Ecology, 2019, 43(2): 119-130. DOI: 10.17521/cjpe.2018.0269
研究地位于山东省高唐县国有旧城林场(36.81° N, 116.09° E), 海拔30 m, 属于黄泛冲积平原。该地属暖温带半干旱季风区域大陆性气候, 具有显著的季节变化, 光照充足, 热量丰富。年降水量544.7 mm, 降水主要集中在7-8月, 年蒸发量1 880 mm, 年平均气温12.0-14.1 ℃, 极端最高气温41.2 ℃, 极端最低气温-20.8 ℃。年日照时间达2 651.9 h, 无霜期204天, 地下水位6 m左右, 土壤质地为砂壤土(砂粒含量65%)。
在相邻的两片纯林和混交林内(表1)分别设定3个30 m × 30 m样地, 根据样地的平均树高及平均胸径选取平均标准木。在纯林内选择1株毛白杨平均标准木(基径42.9 cm, 胸径35.0 cm, 树高30.5 m), 混交林内选择相邻的1株毛白杨(基径37.2 cm, 胸径35.9 cm, 树高27.8 m)与1株刺槐(基径31.6 cm, 胸径25.1 cm, 树高20.8 m)。通过基径处的年轮数得知纯林内毛白杨的树龄为38年, 混交林内毛白杨的树龄为35年, 混交林内刺槐的树龄为34年。
Table 1 表1 表1毛白杨纯林与毛白杨刺槐混交林林地基本信息表 Table 1General information of sample plots in Populus tomentosa stand and mixtures with P. tomentosa and Robinia pseudoacacia
林型 Stand type
树种 Tree species
平均胸径 Average DBH (cm)
平均树高 Average tree height (m)
株行距 Plant & row spacing (m)
平均冠幅 Average crown width (m)
间伐措施 Thinning treatment
毛白杨纯林 Populus tomentosa stand
毛白杨 P. tomentosa
36.1
35.5
6 × 6
7.0
造林后第十年间伐除去刺槐, 以前的混交林成为现有纯林 All R. pseudoacacia trees were removed from mixed-species stand after 10 years of planting, resulting in P. tomentosa pure stand
毛白杨×刺槐混交林 Mixtures with P. tomentosa and Robinia pseudoacacia
毛白杨 P. tomentosa
37.0
26.4
3 × 6
6.6
无 None
毛白杨×刺槐混交林 Mixtures with P. tomentosa and R. pseudoacacia
Fig. 1Photos of root systems of Populus tomentosa in pure stand (A), P. tomentosa and Robinia pseudoacacia in mixed stand (B) (Lens position to the east).
Table 3 表3 表3毛白杨及刺槐3个根系系统拓扑指标参数 Table 3The values of the topological indices for three root systems
根系系统 Root system
a (A)
Pe
V0 (M)
b
qa
qb
TI
纯林毛白杨 Populus tomentosa in pure forest
29
8 367
793
10.551
0.02
-0.000 2
0.50
混交林毛白杨 P. tomentosa in mixed forest
61
21 322
1 456
14.644
0.03
0.004 0
0.56
混交林刺槐 Robinia pseudoacacia in mixed forest
21
3 990
465
8.581
0.02
-0.006 0
0.50
a (A), number of links in the longest unique path from the base link to an exterior link; Pe, sum of numbers of links in all possible unique paths from the base link to all exterior links; V0 (M), number of exterior links; b, average topological length b = Pe/V0; qa =$\frac{a-1-\text{ln}{{V}_{0}}/\text{ln}2}{{{V}_{0}}-1-\text{ln}{{V}_{0}}/\text{ln}2}$; qb =$\frac{b-1-\text{ln}{{V}_{0}}/\text{ln}2}{\frac{{{V}_{0}}+1}{2}-{{V}_{0}}^{-1}-ln{{V}_{0}}/\text{ln}2}$; TI = lgA/lgM (Oppelt et al., 2001). a (A), 最长通道根基到根终端连结数量; Pe, 所有通道根基到根终端通道的连结总数; V0 (M), 外部连结总和; b, 根系平均拓扑长度b = Pe/V0; qa =$\frac{a-1-\text{ln}{{V}_{0}}/\text{ln}2}{{{V}_{0}}-1-\text{ln}{{V}_{0}}/\text{ln}2}$; qb =$\frac{b-1-\text{ln}{{V}_{0}}/\text{ln}2}{\frac{{{V}_{0}}+1}{2}-{{V}_{0}}^{-1}-\text{ln}{{V}_{0}}/\text{ln}2}$; TI = lgA/lgM (Oppelt et al., 2001).
Fig. 2Variations of basal diameter, length, and number of roots for different order roots in root systems of Populus tomentosa in pure forest (A, D, G), P. tomentosa in mixed forest (B, E, H), and Robinia pseudoacacia in mixed forest (C, F, I). The individual points on the box plots represent outliers, the squares on the box plots represent the average.
Table 4 表4 表4毛白杨与刺槐根系基径、长度、数量与根系等级的拟合结果 Table 4Results of fitting regression models of Populus tomentosa and Robinia pseudoacacia—regressing root diameter, length, and numbers (y) on root orders (x), respectively
根形态指标 Root morphological characteristics
根系系统 Root system
方程 Regression model
拟合优度 R2
样本量 n
基径 Basal diameter
纯林毛白杨 P. tomentosa in pure forest
y = 0.86381 + 99.24471exp(-2.32112x)
0.484 0
940
混交林毛白杨 P. tomentosa in mixed forest
y = 0.70036 + 157.41951exp(-2.77342x)
0.414 9
1 527
混交林刺槐 R. pseudoacacia in mixed forest
y = 0.80671 + 91.76619exp(-2.63586x)
0.374 7
541
长度 Length
纯林毛白杨 P. tomentosa in pure forest
y = 67.63168 + 1098.3759exp(-3.32236x)
0.014 6
940
混交林毛白杨 P. tomentosa in mixed forest
y = 39.71891 + 1313.84526exp(-1.98679x)
0.117 2
1 527
混交林刺槐 R. pseudoacacia in mixed forest
y = 92.10741 + 477664.65953exp(-8.10278x)
0.079 5
541
数量 Numbers
纯林毛白杨 P. tomentosa in pure forest
y = 5.97738 + 396.37817exp(-0.5((x-2.93757)/0.90608)2)
0.989 5
7
混交林毛白杨 P. tomentosa in mixed forest
y = 22.41349 + 474.80796exp(-0.5((x-3.28655)/1.13184)2)
0.912 7
8
混交林刺槐 R. pseudoacacia in mixed forest
y = 5.83356 + 196.21954exp(-0.5((x-2.79311)/1.01443)2)
Fig. 3Link length variations along link sequence from the root base to the root tip in root systems of Populus tomentosa in pure forest (A), P. tomentosa in mixed forest (B), and Robinia pseudoacacia in mixed forest (C). 1-3 refer to root categories from first order to third order. The individual points on the box plots represent outliers, the squares on the box plots represent the average, and the lines connected the box plots.
Fig. 4Basal diameter relationships between the mother roots and the child roots. 1, refer to mother-root categories from first order. 2, refer to mother-root categories from second order. 3, refer to mother-root categories from third order. A, root system of Populus tomentosa in pure forest. B, root system of P. tomentosa in mixed forest. C, root system of Robinia pseudoacacia in mixed forest.
BakkerMR, AugustoL, AchatDL ( 2006). Fine root distribution of trees and understory in mature stands of maritime pine (Pinus pinaster) on dry and humid sites. Plant and Soil, 286, 37-51. [本文引用: 1]
BécelC, VercambreG, PagèsL ( 2012). Soil penetration resistance, a suitable soil property to account for variations in root elongation and branching Plant and Soil, 353, 169-180. [本文引用: 4]
BerntsonGM ( 1994). Modelling root architecture: Are there tradeoffs between efficiency and potential of resource acquisition? New Phytologist, 127, 483-493. [本文引用: 2]
ChangWJ, GuoDL ( 2008). Variation in root diameter among 45 common tree species in temperate, subtropical and tropical forests in China Journal of Plant Ecology (Chinese Version), 32, 1248-1257. [本文引用: 2]
DanjonF, KhuderH, StokesA ( 2013). Deep phenotyping of coarse root architecture in R. pseudoacacia reveals that tree root system plasticity is confined within its architectural model. PLOS ONE, 8, e83548. DOI: 10.1371/journal.pone.0083548. [本文引用: 1]
DanjonF, ReubensB ( 2008). Assessing and analyzing 3D architecture of woody root systems, a review of methods and applications in tree and soil stability, resource acquisition and allocation Plant and Soil, 303, 1-34. [本文引用: 3]
DanjonF, BarkerDH, DrexhageM, StokesA ( 2008). Using three-dimensional plant root architecture in models of shallow-slope stability Annals of Botany, 101, 1281-1293.
DanjonF, FourcaudT, BertD ( 2005). Root architecture and wind-firmness of mature Pinus pinaster. New Phytologist, 168, 387-400. [本文引用: 2]
DawsonTE, PateJS ( 1996). Seasonal water uptake and movement in root systems of Australian phraeatophytic plants of dimorphic root morphology: A stable isotope investigation Oecologia, 107, 13-20. [本文引用: 1]
De DeurwaerderH, Herve-FernandezP, StahlC, BurbanB, PetronelliP, HoffmanB, BonalD, BoeckxP, VerbeeckH ( 2018). Liana and tree below-ground water competition— Evidence for water resource partitioning during the dry season Tree Physiology, 38, 1071-1083. [本文引用: 1]
DiN, LiuY, MeadDJ, XieYQ, JiaLM, XiBY ( 2018). Root-system characteristics of plantation-grown Populus tomentosa adapted to seasonal fluctuation in the groundwater table. Trees, 32, 137-149. [本文引用: 2]
FanY, Miguez-MachoG, JobbagyEG, JacksonRB, Otero- CasalC ( 2017). Hydrologic regulation of plant rooting depth Proceedings of the National Academy of Sciences of the United States of America, 114, 10572-10577. [本文引用: 5]
FitterAH ( 1986). The topology and geometry of plant root systems: Influence of watering rate on root system topology in trifolium pretense Annals of Botany, 58, 91-101. [本文引用: 1]
FitterAH ( 1987). An architectural approach to the comparative ecology of plant root systems New Phytologist, 106, 61-77. [本文引用: 1]
FitterAH, SticklandTR ( 1991). Architectural analysis of plant root systems: 2. Influence of nutrient supply on architecture in contrasting plant species New Phytologist, 118, 383-389. [本文引用: 3]
G?rtnerH, WagnerB, HeinrichI, DenierC ( 2009). 3D-laser scanning: A new method to analyze coarse tree root systems Forest Snow and Landscape Research, 82, 95-106. [本文引用: 1]
GuoDL, XiaMX, WeiX, ChangWJ, LiuY, WangZQ ( 2008). Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species New Phytologist, 180, 673-683. [本文引用: 1]
KalliokoskiT, Siev?nenR, NygrenP ( 2010). Tree roots as self-similar branching structures: Axis differentiation and segment tapering in coarse roots of three boreal forest tree species Trees, 24, 219-236. [本文引用: 1]
LecompteF, PagesL, Ozier-LafontaineH ( 2005). Patterns of variability in the diameter of lateral roots in the banana root system New Phytologist, 167, 841-850. [本文引用: 1]
NagelKA, KastenholzB, JahnkeS, Van DusschotenD, AachT, MuhlichM, TruhnD, ScharrH, TerjungS, WalterA, SchurrU ( 2009). Temperature responses of roots: Impact on growth, root system architecture and implications for phenotyping Functional Plant Biology, 36, 947-959. [本文引用: 1]
OppeltAL, KurthW, GodboldDL ( 2001). Topology, scaling relations and Leonardo’s rule in root systems from African tree species Tree Physiology, 21, 117-128. [本文引用: 3]
Ozier-LafontaineH, SillonJF ( 1999). Fractal analysis of the root architecture of Gliricidia sepium for the spatial prediction of root branching, size and mass: Model development and evaluation in agroforestry. Plant and Soil, 209, 167-179. [本文引用: 5]
PagèsL, DoussanC, VercambreG ( 2000). An introduction on below-ground environment and resource acquisition, with special reference on trees. Simulation models should include plant structure and function Annals of Forest Science, 57, 513-520. [本文引用: 1]
PagèsL, XieJ, SerraV ( 2013). Potential and actual root growth variations in root systems: Modeling them with a two-step stochastic approach Plant and Soil, 373, 723-735. [本文引用: 1]
PagèsL ( 2014). Branching patterns of root systems: Quantitative analysis of the diversity among dicotyledonous species Annals of Botany, 114, 591-598. [本文引用: 1]
PateJS, JeschkeWD, AylwardMJ ( 1995). Hydraulic architecture and xylem structure of the dimorphic root systems of South-West Australian species of Proteaceae Journal of Experimental Botany, 46, 907-915. [本文引用: 1]
PregitzerKS, KubiskeME, YuCK, HendrickRL ( 1997). Relationships among root branch order, carbon, and nitrogen in four temperate species Oecologia, 111, 302-308. [本文引用: 1]
PregitzerKS, DeforestJL, BurtonAJ, AllenMF, HendrickRRL ( 2002). Fine root architecture of nine North American trees Ecological Monographs, 72, 293-309. [本文引用: 2]
RoseDA ( 1983). The description of the growth of root systems Plant and Soil, 75, 405-415. [本文引用: 6]
Salahuddin, RewaldB, RazaqM, LixueY, LiJ, KhanF, JieZ ( 2018). Root order-based traits of Manchurian walnut & larch and their plasticity under interspecific competition Scientific Reports, 8, 9815. DOI: 10.1038/s41598-?018-?27832-0. [本文引用: 1]
SchenkHJ, JacksonRB ( 2002 a). The global biogeography of roots Ecological Monographs, 72, 311-328. [本文引用: 1]
SchenkHJ, JacksonRB ( 2002 b). Rooting depths, lateral spreads, and below-ground/above-ground allometries of plants in water-limited ecosystems Journal of Ecology, 90, 480-494. [本文引用: 1]
SchenkHJ, JacksonRB ( 2005). Mapping the global distribution of deep roots in relation to climate and soil characteristics Geoderma, 126, 129-140. [本文引用: 2]
ShafrothPB, StrombergJC, PattenDT ( 2000). Woody riparian vegetation response to different alluvial water table regimes Western North American Naturalist, 60, 66-76. [本文引用: 1]
StrahlerAN ( 1957). Quantitative analysis of watershed geomorphology Eos Transactions American Geophysical Union, 38, 913-920.
TardieuF, PellerinS ( 1991). Influence of soil temperature during root appearance on the trajectory of nodal roots of field grown maize Plant and Soil, 131, 207-214. [本文引用: 1]
Valdes-RodriguezOA, Sanchez-SanchezO, Perez-VazquezA, CaplanJS, DanjonF (2013). Jatropha curcas L. root structure and growth in diverse soils The Scientific World Journal, 2013,827295. DOI: 10.1155/2013/827295. [本文引用: 1]
VercambreG, PagesL, DoussanC, HabibR ( 2003). Architectural analysis and synthesis of the plum tree root system in an orchard using a quantitative modelling approach Plant and Soil, 251, 1-11. [本文引用: 1]
VennetierM, ZanettiC, MeriauxP, MaryB ( 2015). Tree root architecture: New insights from a comprehensive study on dikes Plant and Soil, 387, 81-101. [本文引用: 1]
WagnerB, Ga?rtnerH, SantiniS, IngensandH ( 2011). Cross- sectional interpolation of annual rings within a 3D root model Dendrochronologia, 29, 201-210. [本文引用: 1]
WuQ ( 2016). Analyzing and Modeling of Root Architecture of Individual Maize Plants Based on High-accuracy Field Data and Its Applications PhD dissertation, China Agricultural University, Beijing. [本文引用: 4]
WuQ, PagèsL, WuJ ( 2016). Relationships between root diameter, root length and root branching along lateral roots in adult, field-grown maize Annals of Botany, 117, 379.
XuGQ, LiY ( 2008). Rooting depth and leaf hydraulic conductance in the xeric tree Haloxyolon ammodendron growing at sites of contrasting soil texture. Functional Plant Biology, 35, 1234-1242. [本文引用: 1]
YanXL, LiaoH, GeZY, LuoXW ( 2000). Root architectural characteristics and phosphorus acquisition efficiency in plants Chinese Bulletin of Botany, 17, 511-519. [本文引用: 3]
YangM, DéfossezP, DanjonF, FourcaudT ( 2018). Analyzing key factors of roots and soil contributing to tree anchorage of Pinus species. Trees, 32, 703-712. [本文引用: 1]
YuP, HochholdingerF, LiC ( 2015). Root-type-specific plasticity in response to localized high nitrate supply in maize (Zea mays). Annals of Botany, 116, 751-762. [本文引用: 1]
ZhaoYY, LuZH, XiaJB, LiuJT ( 2015). Root architecture and adaptive strategy of 3 shrubs in Shell Bay in Yellow River Delta Acta Ecologica Sinica , 35, 1688-1695. [本文引用: 1]
Deep phenotyping of coarse root architecture in R. pseudoacacia reveals that tree root system plasticity is confined within its architectural model. 1 2013
... 对根系构型的精确评估需要挖掘根部甚至整个根系统, 这是昂贵且耗时的(Smit et al., 2000; Danjon & Reubens, 2008).这就是在根系构型的详细研究中研究个体数量较少的原因(Vercambre et al., 2003; Danjon et al., 2005, 2013; Wagner et al., 2011; Valdes-Rodriguez et al., 2013), 而对成熟林木的根系研究相对更少.有****们使用三维仪器和软件来深入探究成熟林木尤其是相当大的根系的结构与功能(Danjon & Reubens, 2008), 比如G?rtner等(2009)提出用三维激光扫描(TLS)技术来为成熟云杉(Picea abies)林木根系建立三维模型, 为根系锚固研究和根系年轮研究奠定基础; Danjon等(2005)使用3D数字化设备对海岸松(Pinus pinaster)成熟林木的根系结构和抗风力进行探究; 还有****用吊车获取了10个不同树种的106个成熟林木根系系统, 揭示根系构型中物种、环境、根大小和根类型之间的关系(Vennetier et al., 2015); Osawa等(2010)在西伯利亚多年冻土区通过挖掘法获取了不同林龄(幼龄林、成熟林和过熟林)的落叶松(Larix gmelinii)树根, 研究了完整根系的空间结构、根系分布与冠层分布的关系以及根系结构与土壤因子的关系.成熟林木的完整根系系统中各级根系间的几何形态和拓扑结构对根系结构-功能的研究具有重要意义, 但是因为工作复杂且繁琐, 至今没有研究深入和详细地探 索过. ...
Assessing and analyzing 3D architecture of woody root systems, a review of methods and applications in tree and soil stability, resource acquisition and allocation 3 2008
Seasonal water uptake and movement in root systems of Australian phraeatophytic plants of dimorphic root morphology: A stable isotope investigation 1 1996
... 深根和广泛侧根的同时发育也表明砂壤土上的成熟林木具有二态型根系(Pate et al., 1995; Dawson & Pate, 1996; Fan et al., 2017)以适应这种季节性水分限制环境. ...
Liana and tree below-ground water competition— Evidence for water resource partitioning during the dry season 1 2018
... 植物根系分布深度受树种(Fan et al., 2017), 降水(Schenk & Jackson, 2002a, 2005), 土壤厚度和土壤质地、地形(Schenk & Jackson, 2005; Xu & Li, 2008; Fan et al., 2017), 地下水位(Fan et al., 2017; Di et al., 2018)等影响.本研究中根系的扎根深度最远达到6.4 m (研究地的地下水位6 m左右), 而根系系统主体分布在1.5 m以上的土壤层.造成这种现象的原因可能是根系为适应季节性的降水, 提高了湿润季的吸水效率, 使根系在浅土层集中分布(De Deurwaerder et al., 2018), 同时湿润季节较深的降雨入渗会诱导深土层根系的增殖, 促进形成相对较深的根系分布, 在这个过程中, 砂壤土的质地还可以调节水的向下渗透和向上毛细提升, 共同塑造根系的深层扎根(Fan et al., 2017), 形成的深层根系可以利用地下水来缓解旱季的缺水(Di et al., 2018). ...
Root-system characteristics of plantation-grown Populus tomentosa adapted to seasonal fluctuation in the groundwater table. 2 2018
... 植物根系分布深度受树种(Fan et al., 2017), 降水(Schenk & Jackson, 2002a, 2005), 土壤厚度和土壤质地、地形(Schenk & Jackson, 2005; Xu & Li, 2008; Fan et al., 2017), 地下水位(Fan et al., 2017; Di et al., 2018)等影响.本研究中根系的扎根深度最远达到6.4 m (研究地的地下水位6 m左右), 而根系系统主体分布在1.5 m以上的土壤层.造成这种现象的原因可能是根系为适应季节性的降水, 提高了湿润季的吸水效率, 使根系在浅土层集中分布(De Deurwaerder et al., 2018), 同时湿润季节较深的降雨入渗会诱导深土层根系的增殖, 促进形成相对较深的根系分布, 在这个过程中, 砂壤土的质地还可以调节水的向下渗透和向上毛细提升, 共同塑造根系的深层扎根(Fan et al., 2017), 形成的深层根系可以利用地下水来缓解旱季的缺水(Di et al., 2018). ...
... ), 形成的深层根系可以利用地下水来缓解旱季的缺水(Di et al., 2018). ...
Hydrologic regulation of plant rooting depth 5 2017
... 植物根系分布深度受树种(Fan et al., 2017), 降水(Schenk & Jackson, 2002a, 2005), 土壤厚度和土壤质地、地形(Schenk & Jackson, 2005; Xu & Li, 2008; Fan et al., 2017), 地下水位(Fan et al., 2017; Di et al., 2018)等影响.本研究中根系的扎根深度最远达到6.4 m (研究地的地下水位6 m左右), 而根系系统主体分布在1.5 m以上的土壤层.造成这种现象的原因可能是根系为适应季节性的降水, 提高了湿润季的吸水效率, 使根系在浅土层集中分布(De Deurwaerder et al., 2018), 同时湿润季节较深的降雨入渗会诱导深土层根系的增殖, 促进形成相对较深的根系分布, 在这个过程中, 砂壤土的质地还可以调节水的向下渗透和向上毛细提升, 共同塑造根系的深层扎根(Fan et al., 2017), 形成的深层根系可以利用地下水来缓解旱季的缺水(Di et al., 2018). ...
... ; Fan et al., 2017), 地下水位(Fan et al., 2017; Di et al., 2018)等影响.本研究中根系的扎根深度最远达到6.4 m (研究地的地下水位6 m左右), 而根系系统主体分布在1.5 m以上的土壤层.造成这种现象的原因可能是根系为适应季节性的降水, 提高了湿润季的吸水效率, 使根系在浅土层集中分布(De Deurwaerder et al., 2018), 同时湿润季节较深的降雨入渗会诱导深土层根系的增殖, 促进形成相对较深的根系分布, 在这个过程中, 砂壤土的质地还可以调节水的向下渗透和向上毛细提升, 共同塑造根系的深层扎根(Fan et al., 2017), 形成的深层根系可以利用地下水来缓解旱季的缺水(Di et al., 2018). ...
... ), 地下水位(Fan et al., 2017; Di et al., 2018)等影响.本研究中根系的扎根深度最远达到6.4 m (研究地的地下水位6 m左右), 而根系系统主体分布在1.5 m以上的土壤层.造成这种现象的原因可能是根系为适应季节性的降水, 提高了湿润季的吸水效率, 使根系在浅土层集中分布(De Deurwaerder et al., 2018), 同时湿润季节较深的降雨入渗会诱导深土层根系的增殖, 促进形成相对较深的根系分布, 在这个过程中, 砂壤土的质地还可以调节水的向下渗透和向上毛细提升, 共同塑造根系的深层扎根(Fan et al., 2017), 形成的深层根系可以利用地下水来缓解旱季的缺水(Di et al., 2018). ...
... ), 同时湿润季节较深的降雨入渗会诱导深土层根系的增殖, 促进形成相对较深的根系分布, 在这个过程中, 砂壤土的质地还可以调节水的向下渗透和向上毛细提升, 共同塑造根系的深层扎根(Fan et al., 2017), 形成的深层根系可以利用地下水来缓解旱季的缺水(Di et al., 2018). ...
... 深根和广泛侧根的同时发育也表明砂壤土上的成熟林木具有二态型根系(Pate et al., 1995; Dawson & Pate, 1996; Fan et al., 2017)以适应这种季节性水分限制环境. ...
The topology and geometry of plant root systems: Influence of watering rate on root system topology in trifolium pretense 1 1986
3D-laser scanning: A new method to analyze coarse tree root systems 1 2009
... 对根系构型的精确评估需要挖掘根部甚至整个根系统, 这是昂贵且耗时的(Smit et al., 2000; Danjon & Reubens, 2008).这就是在根系构型的详细研究中研究个体数量较少的原因(Vercambre et al., 2003; Danjon et al., 2005, 2013; Wagner et al., 2011; Valdes-Rodriguez et al., 2013), 而对成熟林木的根系研究相对更少.有****们使用三维仪器和软件来深入探究成熟林木尤其是相当大的根系的结构与功能(Danjon & Reubens, 2008), 比如G?rtner等(2009)提出用三维激光扫描(TLS)技术来为成熟云杉(Picea abies)林木根系建立三维模型, 为根系锚固研究和根系年轮研究奠定基础; Danjon等(2005)使用3D数字化设备对海岸松(Pinus pinaster)成熟林木的根系结构和抗风力进行探究; 还有****用吊车获取了10个不同树种的106个成熟林木根系系统, 揭示根系构型中物种、环境、根大小和根类型之间的关系(Vennetier et al., 2015); Osawa等(2010)在西伯利亚多年冻土区通过挖掘法获取了不同林龄(幼龄林、成熟林和过熟林)的落叶松(Larix gmelinii)树根, 研究了完整根系的空间结构、根系分布与冠层分布的关系以及根系结构与土壤因子的关系.成熟林木的完整根系系统中各级根系间的几何形态和拓扑结构对根系结构-功能的研究具有重要意义, 但是因为工作复杂且繁琐, 至今没有研究深入和详细地探 索过. ...
Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species 1 2008
... a (A), number of links in the longest unique path from the base link to an exterior link; Pe, sum of numbers of links in all possible unique paths from the base link to all exterior links; V0 (M), number of exterior links; b, average topological length b = Pe/V0; qa =$\frac{a-1-\text{ln}{{V}_{0}}/\text{ln}2}{{{V}_{0}}-1-\text{ln}{{V}_{0}}/\text{ln}2}$; qb =$\frac{b-1-\text{ln}{{V}_{0}}/\text{ln}2}{\frac{{{V}_{0}}+1}{2}-{{V}_{0}}^{-1}-ln{{V}_{0}}/\text{ln}2}$; TI = lgA/lgM (Oppelt et al., 2001). ...
... a (A), 最长通道根基到根终端连结数量; Pe, 所有通道根基到根终端通道的连结总数; V0 (M), 外部连结总和; b, 根系平均拓扑长度b = Pe/V0; qa =$\frac{a-1-\text{ln}{{V}_{0}}/\text{ln}2}{{{V}_{0}}-1-\text{ln}{{V}_{0}}/\text{ln}2}$; qb =$\frac{b-1-\text{ln}{{V}_{0}}/\text{ln}2}{\frac{{{V}_{0}}+1}{2}-{{V}_{0}}^{-1}-\text{ln}{{V}_{0}}/\text{ln}2}$; TI = lgA/lgM (Oppelt et al., 2001). ...
... 对根系构型的精确评估需要挖掘根部甚至整个根系统, 这是昂贵且耗时的(Smit et al., 2000; Danjon & Reubens, 2008).这就是在根系构型的详细研究中研究个体数量较少的原因(Vercambre et al., 2003; Danjon et al., 2005, 2013; Wagner et al., 2011; Valdes-Rodriguez et al., 2013), 而对成熟林木的根系研究相对更少.有****们使用三维仪器和软件来深入探究成熟林木尤其是相当大的根系的结构与功能(Danjon & Reubens, 2008), 比如G?rtner等(2009)提出用三维激光扫描(TLS)技术来为成熟云杉(Picea abies)林木根系建立三维模型, 为根系锚固研究和根系年轮研究奠定基础; Danjon等(2005)使用3D数字化设备对海岸松(Pinus pinaster)成熟林木的根系结构和抗风力进行探究; 还有****用吊车获取了10个不同树种的106个成熟林木根系系统, 揭示根系构型中物种、环境、根大小和根类型之间的关系(Vennetier et al., 2015); Osawa等(2010)在西伯利亚多年冻土区通过挖掘法获取了不同林龄(幼龄林、成熟林和过熟林)的落叶松(Larix gmelinii)树根, 研究了完整根系的空间结构、根系分布与冠层分布的关系以及根系结构与土壤因子的关系.成熟林木的完整根系系统中各级根系间的几何形态和拓扑结构对根系结构-功能的研究具有重要意义, 但是因为工作复杂且繁琐, 至今没有研究深入和详细地探 索过. ...
Fractal analysis of the root architecture of Gliricidia sepium for the spatial prediction of root branching, size and mass: Model development and evaluation in agroforestry. 5 1999
An introduction on below-ground environment and resource acquisition, with special reference on trees. Simulation models should include plant structure and function 1 2000
... A method combining both excavation and analysis was applied to extract and quantify root architectural characteristics of the three root systems. The morphological characteristics such as root basal diameter, root length, link length, and root number of different root orders were described using the developmental analysis method of Rose (1983), and their topological relationships were analyzed. ...
 ,
, ,1,*
,1,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT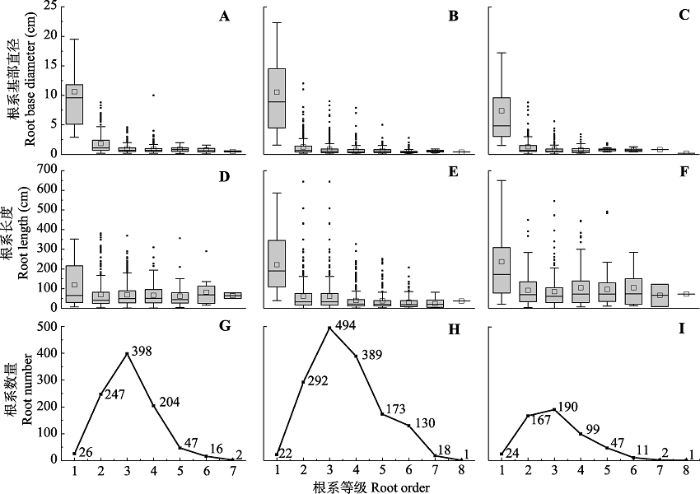 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT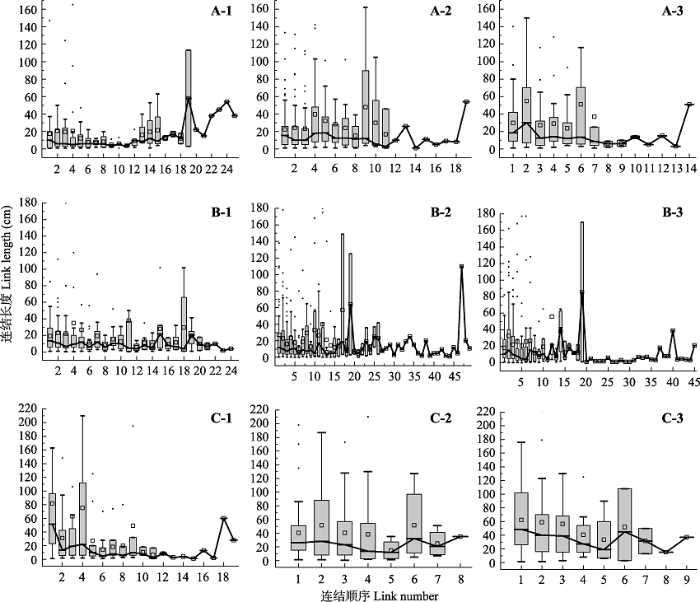 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT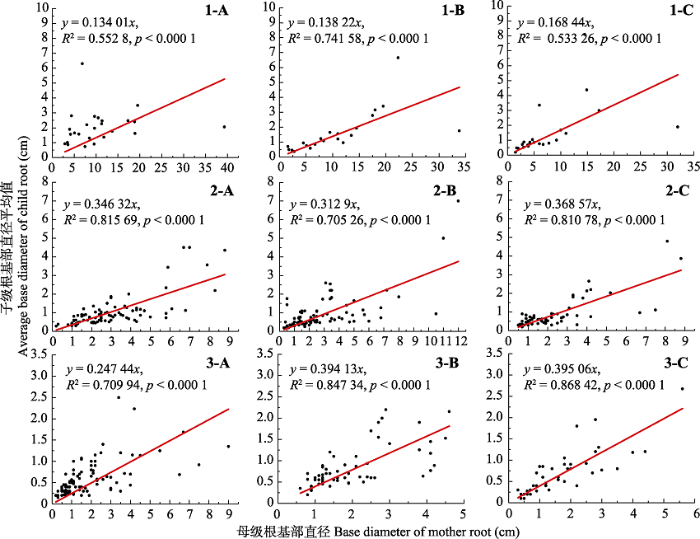 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT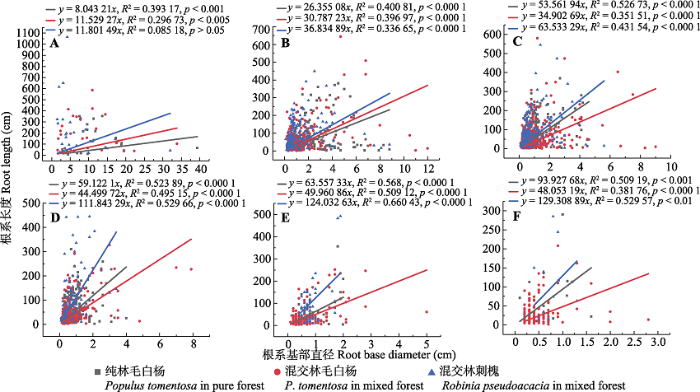 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}