Effects of fine root decomposition on bacterial community structure of four dominated tree species in Mount Taishan, China
LU Ying, LI Kun, NI Rui-Qiang, LIANG Qiang, LI Chuan-Rong, ZHANG Cai-Hong,*Taishan Forest Ecosystem Research Station of the State Forestry Administration, Tai’an, Shandong 271018, China; and Key Labora-tory of State Forestry Administration for Silviculture of the Lower Yellow River, Tai’an, Shandong 271018, China
Supported by the National Natural Science Foundation of China.31500362 Supported by the National Natural Science Foundation of China.31570705 the Joint Special Project of Shandong Province.ZR2014CL005 the Funds of Shandong “Double Tops” Program.SYL2017XTTD03
Abstract Aims Microorganisms play a crucial role in the litter decomposition process in terrestrial ecosystems. Understanding the independent and interactive relationship between fine root decomposition and bacteria community related to substrate characteristics can help to predict the consequences of changes on ecosystem function. Therefore, the aim of this study was to identify fine roots’ influences on rhizosphere microbial structure and diversity.Methods The decomposition of root litters of four dominant tree species of Mount Taishan (Robinia pseudoacacia(RP), Quercus acutissima(QA), Pinus tabulaeformis(PT) and Pinus densiflora(PD)) was tested in a Yaoxiang Forest Farm. Using Illumina high-throughput sequencing of 16S rRNA genes, bacterial community composition was determined. Composition, diversity and relative abundance of bacteria were calculated for per fine root litter.Important findings (1) Fine root litter decomposition differed significantly among different root types. There was no difference in decomposition rate between broad-leaved species and conifer species. In all species, fine roots of RP and QA were more strongly decomposed than that of PT and PD, and these differences were significant (RP > QA > PT > PD). (2) The number of observed species, operational taxonomic units, Ace index and phylogenetic diversity in broad-leaved species were significantly lower than that in coniferous species. Bacterial community structure differed significantly among four species for root decomposition. Initial carbon (C), lignin:nitrogen (N) and C:N in fine root had a great influence on the bacterial community structure. (3) At the phylum level, a total of 4 phyla were dominant (>5% across all species). Based on the average relative abundance, the most abundant phyla were Proteobacteria, Actinomyces, Bacteroidetes and Acidobacteria. Proteobacteria’s and Acidbacteria’s abundance were significantly different among the four species. Particularly, the Proteobacteria of broad-leaved species was significantly higher than that of coniferous species. At the class level, a wide range of classes dominated. Based on the average relative abundance, the most abundance classes were Alphaproteobacteria, Betaproteobacteria, Gammaproteobacteria, unidentified-Actinobacteria and Sphingobacteriia. Alphaproteobacteria and unidentified-Actinobacteria had significant differences among the four species. (4) Pearson correlation analysis showed that the relative abundance of dominant phylum and class was affected by the initial properties of root litter, especially the Proteobacteria and Alphaproteobacteria. In addition, there was a significant positive correlation between fine root decomposition rate and relative abundance of Proteobacteria and Alphaproteobacteria. Redundancy analysis (RDA) also demonstrated that the initial properties of fine root litter (initial N, P, C:N) had significant effects on the structures of bacterial community. These results can improve understanding the links between fine root litter decomposition and functional microbial communities. Keywords:decomposition;fine root;bacterial community;diversity
PDF (1181KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 路颖, 李坤, 倪瑞强, 梁强, 李传荣, 张彩虹. 泰山4种优势造林树种细根分解对细菌群落结构的影响. 植物生态学报[J], 2018, 42(12): 1200-1210 doi:10.17521/cjpe.2018.0120 LU Ying. Effects of fine root decomposition on bacterial community structure of four dominated tree species in Mount Taishan, China. Chinese Journal of Plant Ecology[J], 2018, 42(12): 1200-1210 doi:10.17521/cjpe.2018.0120
Fig. 1Difference in decomposition rate among four litter species (mean ± SE) in Mount Taishan. PD, Pinus densiflora; PT, Pinus tabulaeformis; QA, Quercus acutissima; RP, Robinia pseudoacacia. Different lowercase letters represent significant differences among different species (p < 0.05).
Table 2 表2 表2细根分解一年后泰山细菌多样性统计分析(平均值±标准误差, n = 3) Table 2Statistical analysis of bacterial diversity in Mount Taishan after one year of fine root decomposition (mean ± SE, n = 3)
树种 Species
物种数 NO. of observed species
覆盖率Coverage (%)
Chao1指数 Chao1 index
Ace指数 Ace index
系统发育多样性Phylogenetic diversity
Shannon-Wiener指数Shannon-Wiener index
RP
2 149 ± 71a
98.6 ± 0.1b
3 088.0 ± 140.4ab
3 062.2 ± 143.5ab
159.2 ± 4.2a
8.38 ± 0.59a
QA
1 970 ± 120a
97.7 ± 0.2a
2 824.2 ± 88.5a
2 843.8 ± 62.0a
147.8 ± 7.6a
8.14 ± 0.16a
PD
2 759 ± 25b
98.3 ± 0.2ab
3 544.7 ± 50.3c
3 530.6 ± 34.3c
198.6 ± 5.1b
8.81 ± 0.35b
PT
2 568 ± 39b
97.6 ± 0.2a
3 395.0 ± 2.2bc
3 341.9 ± 68.4bc
193.1 ± 3.2b
8.88 ± 0.18b
PD, Pinus densiflora; PT, Pinus tabulaeformis; QA, Quercus acutissima; RP, Robinia pseudoacacia. Different lowercase letters represent significant differences among different species (p < 0.05). PD, 赤松; PT, 油松; QA, 麻栎; RP, 刺槐。不同小写字母代表不同树种之间差异显著(p < 0.05)。
Table 3 表3 表3分解一年后细菌α多样性与凋落物初始化学性质之间的相关分析 Table 3Correlation analysis between bacterial α diversity and the initial properties of litter after one year of decomposition
C (%)
N (%)
P (%)
C:N
N:P
木质素 Lignin (%)
物种数 NO. Of observed species
0.884**
-0.541
0.679*
0.790*
-0.496
0.726*
覆盖率 Coverage (%)
0.331
0.437
-0.482
0.126
0.517
-0.344
Chao1指数 Chao1 index
0.858**
-0.413
0.608
0.706*
-0.377
0.642
Ace指数 Ace index
0.874**
-0.446
0.593
0.748*
-0.405
0.661*
系统发育多样性 Phylogenetic diversity
0.829*
-0.547
0.744*
0.730*
-0.515
0.749*
Shannon-Wiener指数 Shannon-Wiener index
0.552
-0.292
0.491
0.378
-0.246
0.418
The numbers in the table represent the correlation coefficient. **, p < 0.01;*, p < 0.05. 表中数字代表相关系数; **, p < 0.01; *, p < 0.05。
Fig. 2Nonmetric Multidimensional Scaling (NMDS) ordination diagram of bacterial community structure in root litter after one year of decomposition in Mount Taishan. PD, Pinus densiflora; PT, Pinus tabulaeformis; QA, Quercus acutissima; RP, Robinia pseudoacacia.
Fig. 3Redundancy analysis (RDA) based on bacterial community structure and the initial properties of fine root litter. PD, Pinus densiflora; PT, Pinus tabulaeformis; QA, Quercus acutissima; RP, Robinia pseudoacacia.
Fig. 4Differences in relative abundances of major bacterial dominant groups among the four species in Mount Taishan(mean ± SE). A, Dominant classes. B, Dominant phyla. PD, Pinus densiflora; PT, Pinus tabulaeformis; QA, Quercus acutissima; RP, Robinia pseudoacacia. Different lowercase letters indicate the significant differences in different species of the same bacterial group, while the same letter indicates no significant difference.
Table 4 表4 表4细菌优势门相对丰度与细根分解速率、凋落物初始化学性质之间的相关分析 Table 4Correlation analysis among the bacterial dominant phylum , the decomposition rate of fine roots , and the initial properties of litter
优势门 Dominant phylum
C (%)
N (%)
P (%)
木质素 Lignin (%)
C:N
N:P
分解速率 Decomposition rate
变形菌门 Proteobacteria
-0.64
0.57
0.77*
-0.63
-0.69*
0.52
0.71*
放线菌门 Actinobacteria
0.61
-0.32
-0.69*
0.48
0.50
-0.25
-0.62
拟杆菌门 Bacteroidetes
0.60
0.09
0.09
0.09
0.43
0.03
-0.36
酸杆菌门 Acidobacteria
-0.46
-0.48
-0.57
0.35
-0.16
-0.42
0.03
分解速率 Decomposition rate
-0.76**
0.74**
0.67*
-0.90**
-0.82**
0.74**
1.00
The numbers in the table represent the correlation coefficient. **, p < 0.01;*, p < 0.05 表中数字代表相关系数; **, p < 0.01; *, p < 0.05。
Table 5 表5 表5细菌优势纲相对丰度与细根分解速率、凋落物初始化学性质之间的相关分析 Table 5Correlation analysis among the decomposition rate of fine roots and bacterial dominant class and the initial properties of litter
优势纲 Dominant class
C (%)
N (%)
P (%)
木质素 Lignin (%)
C:N
N:P
分解速率 Decomposition rate
α-变形菌纲 Alphaproteobacteria
-0.33
0.79**
0.56
-0.71*
-0.73*
0.84**
0.63*
β-变形菌纲 Betaproteobacteria
-0.42
-0.18
0.09
-0.00
-0.11
-0.21
0.19
γ-变形菌纲 Gammaproteobacteria
-0.08
0.49
0.47
-0.37
-0.25
0.43
0.24
不明放线菌纲unidentified-Actinobacteria
0.84**
-0.25
-0.53
0.37
0.73*
-0.25
-0.61
鞘脂杆菌纲 Sphingobacteriia
0.20
0.40
0.49
-0.23
-0.00
0.30
0.05
The numbers in the table represent the correlation coefficient. **, p < 0.01;*, p < 0.05. 表中数字代表相关系数; **, p < 0.01; *, p < 0.05。
Fig. 5Redundancy analysis (RDA) based on dominant bacterial phylum and the initial properties of fine root litter. PD, Pinus densiflora; PT, Pinus tabulaeformis; QA, Quercus acutissima; RP, Robinia pseudoacacia.
3 讨论
研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007)。4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2)。张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果。一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989)。其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长。叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2)。细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989)。另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017)。ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致。这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1)。分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018)。阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1)。细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017)。阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016)。这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4)。分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%。前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011)。变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017)。阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B)。P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003)。N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009)。分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致。放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015)。放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致。放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的。但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4)。拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004)。前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致。可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化。酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011)。酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4)。这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989)。其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因。在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05)。张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致。刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5)。研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006)。这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因。γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快。β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变。鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用。Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群。我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同。该研究采用PCR-DGGE, 而我们采用高通量测序技术。
AdamSW, ZacchaeusGC, CindyML ( 2013). Contrasting rRNA gene abundance patterns for aquatic fungi and bacteria in response to leaf-litter chemistry Freshwater Science, 32, 663-672. DOI:10.1899/12-122.1URL [本文引用: 1] Few investigators have examined simultaneous bacterial and fungal responses to leaf-litter chemistry in fresh water. We tested the hypothesis that bacteria would be more abundant on labile litter with lower concentrations of defensive compounds, whereas fungi would be more abundant on recalcitrant litter. We used quantitative-polymerase chain reaction (qPCR) to measure the abundance of bacterial 16S and fungal 18S ribosomal ribonucleic acid (rRNA) genes and found that these groups responded differently to leaf chemistry. Bacterial 16S rRNA genes were 4X more abundant on labile than on recalcitrant litter. In contrast, fungal 18S rRNA genes were 8X more abundant on recalcitrant than on labile litter. Peak bacterial gene abundances on day 6 were related to leaf-litter % bound condensed tannin (r(2) = 0.38), and peak fungal gene abundances on day 14 were related to % soluble condensed tannin (r(2) = 0.49), % bound condensed tannin (r(2) = 0.34), and % lignin (r(2) = 0.33). Leaf-litter C:N ratios were not associated with microbial gene abundance. The ratio of fungal 18S: bacterial 16S genes also increased along the 1st axis in a principal components analysis of phytochemical variables. The early peak in bacterial rRNA gene abundance may indicate the role of bacteria in the early decomposition of leaf litter. rRNA gene abundance patterns demonstrate that bacteria and fungi have different patterns of growth and productivity in response to leaf-litter chemistry.
BarretM, MorrisseyJP, O’GaraF ( 2011). Functional genomics analysis of plant growth-promoting rhizobacterial traits involved in rhizosphere competence Biology and Fertility of Soils, 47, 729-743. DOI:10.1007/s00374-011-0605-xURL [本文引用: 1] In soil, some specific bacterial populations, called plant growth-promoting rhizobacteria are able to promote plant growth and/or reduce the incidence of soil-borne diseases. Rhizosphere competence is an important prerequisite for the efficacy of these biocontrol strains. Therefore, over decades, multiple approaches have been combined to understand the molecular basis of bacterial traits involved in rhizosphere competence. This review addresses the bacterial genes expressed during bacterial lant interactions in the rhizosphere of different plant species. The distribution of these key genes in natural populations of rhizobacteria is also discussed.
ChapmanSK, KochGW ( 2007). What type of diversity yields synergy during mixed litter decomposition in a natural forest ecosystem? Plant and Soil, 299, 153-162. DOI:10.1007/s11104-007-9372-8URL [本文引用: 3] Investigating the relationship of biodiversity and ecosystem function in natural forests allows incorporation of established feedbacks between long-lived plants and soil processes. We studied forested stands in northern Arizona that vary in dominant species richness across small areas. We examined the effects of natural variation in dominant tree biodiversity on ecosystem parameters, particularly litter decomposition. We determined not only whether plant species decompose in mixture as predicted by their individual decomposition rates but also: (1) how particular species affect the decomposition rate of each other in mixture; and (2) whether litter decomposes more rapidly at its site of origin; i.e. is there a “home field advantage” to decomposition? Over a 2-year period, litter mixtures of functionally similar tree species decomposed more rapidly than expected from rates of the individual species alone. Mixtures of conifer species litter decomposed up to 50% faster than expected, with individual conifer members of those mixtures decomposing up to 85% faster than expected. In contrast, more functionally diverse mixtures of litter, which included a deciduous species, did not show synergistic effects during decomposition. We found no significant “home-field advantage” to decomposition. Our study is the first to demonstrate that litter mixtures from more closely related plant species give rise to the most synergistic effects of biodiversity on litter dynamics, indicating that more taxonomically and functionally diverse plant assemblages do not always drive greater emergent effects on ecosystem function.
ChiginevaNI, AleksandrovaAV, TiunovAV ( 2009). The addition of labile carbon alters litter fungal communities and decreases litter decomposition rates Applied Soil Ecology, 42, 264-270. DOI:10.1016/j.apsoil.2009.05.001URL [本文引用: 1] Many field and laboratory studies have demonstrated that the addition of easily available carbon can increase rates of decomposition of plant litter or humified soil organic matter, though opposite trends are also common. In boreal forests, a large part of labile carbon enters soil via tree roots and ectomycorrhizal (EM) network. The influence of increased C availability and the presence of EM roots on litter decomposition was studied in a long-term field experiment in a 50-year-old Picea abies plantation. Litterbags containing litter of three species ( Populus tremula, Quercus robur, or P. abies) were buried to the depth of ca. 2.5 cm. The experiment was set up in a full-factorial design with carbon availability and presence of roots as factors. Carbon was added as an aqueous sucrose solution (50 g C m 2) every second week during snow-free period. Spruce roots around and underneath experimental plots were cut once a month. Subsets of litterbags were analysed after 12, 21 and 24 months. The serial dilution technique was used to assess the fungal community composition. The addition of labile carbon led to a strong decrease in litter decomposition rates, associated with consistent shifts in the composition of saprotrophic microfungal communities, and to an increase in the overall density of culturable fungal species ( r-strategists), but not of specialized cellulolytic microfungi ( K-strategists), isolated on Hutchinson agar. Sucrose addition did not affect the number of species isolated but the dominance structure was shifted toward the domination of the 2 4 most abundant species. The presence of EM roots significantly reduced moisture content in decomposing litter but had only weak and inconsistent effect on the decomposition rates and on the community composition of saprotrophic microfungi; these effects did not depend on the level of carbon availability.
DingXJ, JingRY, HuangYL, ChenBJ, MaFY ( 2017). Bacterial structure and diversity in rhizosphere and non-?rhizosphere of Robinia pseudoacacia in the Yellow River Delta. Acta Pedologica Sinica, 54, 1293-1302. DOI:10.11766/trxb201703230510URL [本文引用: 3] 为精确分析黄河三角洲刺槐根际与非根际土壤细菌群落定殖情况,本研究采用高通量测序方法对刺槐根际与非根际土壤细菌结构及多样性进行了研究.研究表明,根际土壤细菌共有36门214属,非根际土壤细菌共有33门153属.变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、酸杆菌门(Acidobacteria)细菌丰度超过15%,是根际与非根际土壤中的优势菌落.根际与非根际土壤中酸杆菌门、硝化螺旋菌门(Nitrospirae)丰度差异显著.根际土壤中红游动菌属(Rhodoplanes)、溶杆菌属(Lysobacter)、热单胞菌属(Thermomonas)、链霉菌属(Streptomyces)及非根际土壤中红游动菌属、溶杆菌属、链霉菌属、Kaistobacter细菌丰度超过4%.根际土壤中固氮菌丰度显著高于非根际土壤,解磷、解钾细菌丰度差异不显著.根际与非根际土壤细菌Chao丰富度分别为2054、2376,差异显著.根际与非根际土壤细菌之间的权重(Weighted Unifrac)距离在0.12~0.25之间.综上所述,黄河三角洲刺槐根际与非根际土壤细菌结构具有一定差异,多样性差异显著. [ 丁新景, 敬如岩, 黄雅丽, 陈博杰, 马风云 ( 2017). 黄河三角洲刺槐根际与非根际细菌结构及多样性 土壤学报, 54, 1293-1302.] DOI:10.11766/trxb201703230510URL [本文引用: 3] 为精确分析黄河三角洲刺槐根际与非根际土壤细菌群落定殖情况,本研究采用高通量测序方法对刺槐根际与非根际土壤细菌结构及多样性进行了研究.研究表明,根际土壤细菌共有36门214属,非根际土壤细菌共有33门153属.变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、酸杆菌门(Acidobacteria)细菌丰度超过15%,是根际与非根际土壤中的优势菌落.根际与非根际土壤中酸杆菌门、硝化螺旋菌门(Nitrospirae)丰度差异显著.根际土壤中红游动菌属(Rhodoplanes)、溶杆菌属(Lysobacter)、热单胞菌属(Thermomonas)、链霉菌属(Streptomyces)及非根际土壤中红游动菌属、溶杆菌属、链霉菌属、Kaistobacter细菌丰度超过4%.根际土壤中固氮菌丰度显著高于非根际土壤,解磷、解钾细菌丰度差异不显著.根际与非根际土壤细菌Chao丰富度分别为2054、2376,差异显著.根际与非根际土壤细菌之间的权重(Weighted Unifrac)距离在0.12~0.25之间.综上所述,黄河三角洲刺槐根际与非根际土壤细菌结构具有一定差异,多样性差异显著.
EichorstSA, KuskeCR, SchmidtTM ( 2011). Influence of plant polymers on the distribution and cultivation of bacteria in the phylum Acidobacteria Applied and Environmental Microbiology, 77, 586-596. DOI:10.1128/AEM.01080-10URLPMID:3020536 [本文引用: 1] Members of the phylum Acidobacteria are among the most abundant bacteria in soil. Although they have been characterized as versatile heterotrophs, it is unclear if the types and availability of organic resources influence their distribution in soil. The potential for organic resources to select for different acidobacteria was assessed using molecular and cultivation-based approaches with agricultural and managed grassland soils in Michigan. The distribution of acidobacteria varied with the carbon content of soil: the proportion of subdivision 4 sequences was highest in agricultural soils (ca. 41%) that contained less carbon than grassland soils, where the proportions of subdivision 1, 3, 4, and 6 sequences were similar. Either readily oxidizable carbon or plant polymers were used as the sole carbon and energy source to isolate heterotrophic bacteria from these soils. Plant polymers increased the diversity of acidobacteria cultivated but decreased the total number of heterotrophs recovered compared to readily oxidizable carbon. Two phylogenetically novel Acidobacteria strains isolated on the plant polymer medium were characterized. Strains KBS 83 (subdivision 1) and KBS 96 (subdivision 3) are moderate acidophiles with pH optima of 5.0 and 6.0, respectively. Both strains grew slowly ( = 0.01 h
FiererN, BradfordMA, JacksonRB ( 2007). Toward an ecological classification of soil bacteria Ecology, 88, 1354-1364. DOI:10.1890/05-1839URL [本文引用: 1]
GessnerMO, ChauvetE, DobsonM ( 1999). A perspective on leaf litter breakdown in streams Oikos, 85, 377-384. DOI:10.2307/3546505URL [本文引用: 1] Leaf litter breakdown, a critical ecosystem level process in streams and other aquatic environments, has been conceptualized using models borrowed from terrestrial systems. We argue that current views of the process in freshwaters need to be conceptually improved. Specifically, we think the idea that breakdown proceeds in three distinct temporal stages (leaching, conditioning, fragmentation) has been overemphasized. Leaching, the massive loss of soluble leaf components within 24 h after immersion, is generally considered to constitute a well-defined first stage. Recent evidence suggests, however, that the initial solute losses are largely an effect of the unnatural drying procedures to which experimental leaves are normally subjected. Fresh leaf litter does lose solutes when immersed, but gradually throughout the breakdown process rather than instantly upon wetting. Conditioning, the second breakdown stage, describes the enhancement of leaf palatability for detritivores by microbial colonization, and is thus ultimately targeted towards a group of organisms (which contribute to litter degradation) rather than addressing the breakdown process per se. Furthermore, conditioning implies a key role for detritivorous invertebrates and underrates the established direct degradative activity of microbial decomposers. If, thus, leaching and conditioning are not generally useful operators to describe portions of the litter breakdown process in freshwaters, the traditional concept, which emphasises leaching, conditioning and fragmentation as three sequential stages, loses much of its appeal. Consequently, we propose a new conceptual model, in which the coincidence and interplay of various subprocesses of litter breakdown is more strongly recognized. In this model, we propose to view the process in terms of the products of litter breakdown-as a complement to the usual perspective which focuses on litter mass loss. Six primary breakdown products are considered: bacterial, fungal and shredder biomass; dissolved organic matter; fine-particulate organic matter; and inorganic mineralization products such as CO2, NH4+ and PO43-. We present a scheme illustrating the hypothesized formation of these products throughout breakdown. However, to improve understanding of the process, application of the proposed conceptual framework in experimental work is necessary.
GrierCC ( 1981). Biomass distribution and above- and below-?ground production in young and mature Abies amabilis zone ecosystems of the Washington Cascades. Canadian Journal of Forest Research, 11, 155-167. [本文引用: 1]
GuiH, PurahongW, HydeKD, XuJC, MortimerPE ( 2017). The arbuscular mycorrhizal fungus Funneliformis mosseae alters bacterial communities in subtropical forest soils during litter decomposition Frontiers in Microbiology, 8, 1-11. DOI:10.3389/fmicb.2017.01120URLPMID:5476864 [本文引用: 3] Bacterial communities and arbuscular mycorrhizal fungi (AMF) co-occur in the soil, however, the interaction between these two groups during litter decomposition remains largely unexplored. In order to investigate the effect of AMF on soil bacterial communities, we designed dual compartment microcosms, where AMF (Funneliformis mosseae) was allowed access (AM) to, or excluded (NM) from, a compartment containing forest soil and litterbags. Soil samples from this compartment were analyzed at 0, 90, 120, 150, and 180 days. For each sample, Illumina sequencing was used to assess any changes in the soil bacterial communities. We found that most of the obtained operational taxonomic units (OTUs) from both treatments belonged to the phylum of Proteobacteria, Acidobacteria, and Actinobacteria. The community composition of bacteria at phylum and class levels was slightly influenced by both time and AMF. In addition, time and AMF significantly affected bacterial genera (e.g.,Candidatus Solibacter, Dyella, Phenylobacterium) involved in litter decomposition. Opposite to the bacterial community composition, we found that overall soil bacterial OTU richness and diversity are relatively stable and were not significantly influenced by either time or AMF inoculation. OTU richness at phylum and class levels also showed consistent results with overall bacterial OTU richness. Our study provides new insight into the influence of AMF on soil bacterial communities at the genus level.
HultmanJ, WaldropMP, MackelprangR, DavidMM, McFarlandJ, BlazewiczSJ, HardenJ, TuretskyMR, McGuireAD, ShahMB, VerBerkmoesNC, LeeLH, MavrommatisK, JanssonJK . ( 2015). Multi-omics of permafrost, active layer and thermokarst bog soil microbiomes Nature, 521, 208-212. DOI:10.1038/nature14238URLPMID:25739499 [本文引用: 1] Over 20% of Earth's terrestrial surface is underlain by permafrost with vast stores of carbon that, once thawed, may represent the largest future transfer of carbon from the biosphere to the atmosphere. This process is largely dependent on microbial responses, but we know little about microbial activity in intact, let alone in thawing, permafrost. Molecular approaches have recently revealed the identities and functional gene composition of microorganisms in some permafrost soils and a rapid shift in functional gene composition during short-term thaw experiments. However, the fate of permafrost carbon depends on climatic, hydrological and microbial responses to thaw at decadal scales. Here we use the combination of several molecular 'omics' approaches to determine the phylogenetic composition of the microbial communities, including several draft genomes of novel species, their functional potential and activity in soils representing different states of thaw: intact permafrost, seasonally thawed active layer and thermokarst bog. The multi-omics strategy reveals a good correlation of process rates to omics data for dominant processes, such as methanogenesis in the bog, as well as novel survival strategies for potentially active microbes in permafrost.
JonesRT, RobesonMS, LauberCL, HamadyM, KnightR, FiererN ( 2009). A comprehensive survey of soil acidobacterial diversity using pyrosequencing and clone library analyses The ISME Journal, 3, 442-453. DOI:10.1038/ismej.2008.127URLPMID:19129864 [本文引用: 2] The ISME Journal: Multidisciplinary Journal of Microbial Ecology is the official Journal of the International Society for Microbial Ecology, publishing high-quality, original research papers, short communications, commentary articles and reviews in the rapidly expanding and diverse discipline of microbial ecology.
LydellC, DowellL, SikaroodiM, GillevetP , Emerson ( 2004). A population survey of members of the phylum Bacteroidetes isolated from salt marsh sediments along the east coast of the United States Microbial Ecology, 48, 263-273. DOI:10.1007/s00248-003-1068-xURLPMID:15107955 [本文引用: 1] The population diversity of cultured isolates of the phylum Bacteroidetes was investigated from salt-marsh sediments. A total of 44 isolates that belonged to this phylum were isolated either from high-dilution plates or from end-dilution most-probable-number (MPN) tubes. The majority of the isolates came from Virginia, with others isolated from salt marshes in Delaware and North Carolina. All the isolates were aerobic Gram-negative, catalase positive small rods that formed uniform colonies; most had either yellow or orange pigmentation. Riboprinting of 40 isolates revealed they were genotypically diverse, consisting of 33 different riboprint patterns; there were four riboprint groups with two or more members. The isolates could be divided into 23 different fatty acid methyl ester (FAME) profiles at the species level with 14 of the profiles being unique to single isolates. One group of 10 isolates was closely related, suggesting this group may be well adapted for life in salt marshes. Thirteen of the isolates were selected for sequencing of the small-subunit ribosomal RNA gene representing a diverse group of isolates that fell within the classes Sphingobacteria and Flavobacteria . Only one of the isolates was >97% similar at the 16S rDNA to a described species of Cytophaga marinoflava ; the other isolates were 94 to 96.5% related to undescribed isolates mostly within the class Flavobacteria . There was good concordance between the FAME dendrogram and a phylogenetic tree based on comparison of 16S sequences. There were no obvious temporal or spatial distribution patterns to the isolates, suggesting that this group of bacteria is inherently diverse.
McLarenJR, TurkingtonR ( 2010). Plant functional group identity differentially affects leaf and root decomposition Global Change Biology, 16, 3075-3084. DOI:10.1111/j.1365-2486.2009.02151.xURL [本文引用: 2] Losses of species and changes in the composition of plant communities are likely to influence numerous ecosystem functions. Changes in the plant-soil interactions that control decomposition, in particular, could alter carbon and nutrient cycling in soils and further alter other ecosystem functions. The effects of plant communities on decomposition may depend both on the type of tissue being decomposed and also on the different stages of the decomposition process. We used an experimental design where single plant functional groups were removed from a northern grassland to examine the role of plant identity in determining both short-term and long-term above- and belowground decomposition rates. Plant removals were conducted across fertilization and fungicide treatments to examine environmental context-dependency of functional group identity effects on decomposition. There were significant effects of plant functional group identity on aboveground decomposition, with the loss of grasses and forbs slowing decomposition, whereas the effects on belowground decomposition were rare and transient. Effects of plant identity on decomposition were consistent in both short- and long-term decomposition studies indicating that the influences of identity on the decomposition environment remained consistent throughout the different stages of the decomposition process. Both fertilizer and fungicide treatments affected overall decomposition rate, but there were few interactions between these treatments and plant removals. Although current species loss is likely to be happening in concert with environmental changes, the role a species plays in determining ecosystem functions such as decomposition may not be context-dependent in these northern environments, and this may provide greater predictive power in determining the effects of species loss with changing environments. Further, as plant identity shows significant effects on litter decomposition rates, the effects of current and predicted future biodiversity losses may depend specifically on which species are lost.
MeentemeyerV ( 1978). Macroclimate and lignin control of litter decomposition rates Ecology, 59, 465-472. DOI:10.2307/1936576URL [本文引用: 1] In order to develop a general model of litter decomposition rates suitable for the prediction of regional variations in decay rates, and to determine the relative control by macroclimate and litter quality on decomposition rates, data were selected from 5 locations ranging in climate from subpolar to warm temperate. Actual evapotranspiration (AET) was selected as an index of the climatic (energy and moisture) forcing function of the specialized decomposers which is superior to temperature and precipitation. Lignin concentration was selected as an index of litter quality and may be treated as a mediator of climatically (AET) regulated decay rates. In a stepwise, multiple linear correlation-regression, using AET, lignin concentration (%) and AET/lignin concentration (interaction), AET alone accounted for 51% of the variance in observed decay rates, AET/lignin concentration (interaction) added 19% and lignin concentration added 2% of the total (72%) variance accounted. Simple correlation of the five locations between lignin concentration and decomposition rate ranged from r = .32 to r = .95, however, the regression lines for each of the 5 locations indicated that these slopes progressively declined with AET. Moreover, the slope decline was not parallel, indicating a climatically variable control by lignin concentration on decay rates. In low-AET (not arid) climates, litter with high and low lignin, will decay at more nearly similar rates, but as the AET environment increases, the difference in decay rates becomes progressively greater than the increase in AET alone would seem to warrant. A general model of the interaction control by AET and lignin concentration on decomposition rates was formulated which overcomes the restraints of the multiple regression model. At the scale of subpolar to warm-temperate climates, the climate as indicated by AET is several orders of magnitude more important as a predictor of decay rate than is litter quality. This importance is evident in spite of the fact that the data on lignin concentration used in this analysis had a 12-fold range while the AET values had a 2.3-fold range.
SauvadetM, ChauvatM, CluzeauD, MaronPA, VillenaveC, BertrandI ( 2016). The dynamics of soil micro-food web structure and functions vary according to litter quality Soil Biology & Biochemistry, 95, 262-274. DOI:10.1016/j.soilbio.2016.01.003URL [本文引用: 1] Highlights 61 Micro-food web interactions decreased with increasing litter recalcitrance. 61 Interactions networks revealed microbial but not faunal links. 61 Biota strategies for C acquisition differed according to the initial litter quality. 61 Enzymatic production was not explained by the microbial structure. 61 L. terrestris changed enzyme dynamics without altering the micro-food web structure. Abstract Anthropogenic pressures on agricultural soils are known to alter the properties of soil food webs, which may affect ecosystem functions and the capacity to deliver services. Cropping systems fuel belowground biota by supplying litter, but litter quality varies from year to year. In this study, we hypothesized that (i) the structure and function of the soil biota may respond strongly to the quality of crop residue, and (ii) this response may vary with the complexity of the food web. To test this hypothesis, a 3-month incubation experiment was performed using soil columns filled with a silty loam agricultural soil (Estrées-Mons, Northern France), and plant litter was incorporated into the 0–5-cm layer. The quality of the litter was modulated by using either high quality (maize leaves) or low quality materials (maize roots), and two levels of biotic complexity were tested by introducing or not introducing an ecosystem engineer (Lumbricus terrestris, anecic earthworm) into the columns. We then investigated litter decomposition and enzymatic activities, as well as the biomass and diversity of the bacterial, fungal and nematode communities. The C mineralization and enzymatic profiles varied according to litter quality, and the most obvious differences were observed in the hydrolytic enzymes and phenol oxidase. Micro-food web interactions and structure were mainly impacted by litter type and the time since litter incorporation, and as expected, we observed a temporal succession of cellulolytic organisms followed by lignolytic organisms. This progression was mirrored by the nematode community; the ratio of fungal feeders/bacterial feeders increased between 35 and 91 days. Initial litter quality primarily impacted bacterial and fungal community structure, and the development of the bacterial channel was greater with the addition of leaf litter. This was related to an increase in specific, favored phyla, Bacteroidetes and Proteobacteria, whereas Actinobacteria increased more with the addition of root litter. However, we could not statistically link the changes in the structure of the microbial communities over time with those in enzyme activity, which led us to assume that other factors, such as top-down regulation, are more important than microbial community structure in the determination of the enzyme secretions in the soil. Finally, the addition of earthworms did not significantly impact litter decomposition and had a low impact on micro-food web structure and interactions. However, the presence of L. terrestris did modify the enzymatic secretions, probably through its endemic microbial population, as well as the stimulation of microbial activity through predation.
ShenYF, WangN, ChengRM, XiaoWF, YangS, GuoY, LeiL, ZengLX, WangXR ( 2017). Characteristics of fine roots of Pinus massoniana in the Three Gorges Reservoir Area, China Forests, 8(6), 1-13. [本文引用: 1]
SoaresRA, RoeschLFW, ZanattaG, De Oliveira CamargoFA, PassagliaLMP ( 2006). Occurrence and distribution of nitrogen fixing bacterial community associated with oat ( Avena sativa) assessed by molecular and microbiological techniques. Applied Soil Ecology, 33, 221-234. DOI:10.1016/j.apsoil.2006.01.001URL [本文引用: 1] Molecular and microbiological techniques were used to evaluate the nitrogen fixing bacterial community associated with oat ( Avena sativa) in different soil management systems of Rio Grande do Sul State. The conventional microbiological approach made use of semi-solid nitrogen-free and solid selective media to isolate some of these nitrogen fixing bacteria. For the molecular approach, a PCR-RFLP strategy was adopted to amplify a segment of the nifH gene, which encodes nitrogenase reductase, from DNA samples extracted from rhizosphere soils, roots and leaves of oat. The amplified nifH fragments were cleaved with TaqI and HaeIII endonucleases and the products analyzed in polyacrylamide gels. One sample was analyzed in more detail. From this site rhizosphere soil and leaf samples PCR fragments were cloned into pUC18, generating 55 nifH positive clones, from which 19 different RFLP patterns were obtained. These patterns were compared with RFLP patterns generated from DNA obtained from pure strains or by theoretical digestion data. To identify and confirm the RFLP patterns obtained, at least one clone of each was sequenced and the nucleotide sequences were compared with the GenBank nucleotide database. As the partial nifH sequences analyzed had shown homologies with several known and unknown diazotrophs, we attempted to identify the diazotrophs through the positioning of the clones in the NifH partial sequence-derived tree. Through the molecular biological analysis we were able to identify a reasonable occurrence of diazotrophs in the rhizosphere soil and leaf samples analyzed, 9 and 13 different genera, respectively. Of these, two genera were particularly abundant: Azospirillum and Herbaspirillum. These two genera were also the two most abundant genera isolated by conventional microbiological techniques. Both approaches indicated that the nitrogen-fixing communities were different in soils under cultivation and permanent pasture from these under cultivation without live stock. Our results showed that while the molecular approach gives us a general view of the variability of nitrogen fixers in the community, the conventional methodology allows us to isolate those bacteria and to select some of them that have showed the highest nitrogen fixation capacity and auxin production.
SunH, WangQX, LiuN, LiL, ZhangCG, LiuZB, ZhangYY ( 2017). Effects of different leaf litters on the physicochemical properties and bacterial communities in Panax ginseng-growing soil. Applied Soil Ecology, 111, 17-24.
TaylorBR, ParkinsonD, ParsonsWFJ ( 1989). Nitrogen and lignin content as predictors of litter decay rates: A microcosm test Ecology, 70, 97-104. DOI:10.2307/1938416URL [本文引用: 5]
TuomiM, ThumT, JarvinenH, FronzekS, BergB, HarmonM, TrofymowJA, SevantoS, LiskiJ ( 2009). Leaf litter decomposition—Estimates of global variability based on Yasso07 model Ecological Modeling, 220, 3362-3371. DOI:10.1016/j.ecolmodel.2009.05.016URL [本文引用: 1] http://linkinghub.elsevier.com/retrieve/pii/S030438000900386X
UrbanováM, ?najdrJ, BaldrianP ( 2015). Composition of fungal and bacterial communities in forest litter and soil is largely determined by dominant trees Soil Biology & Biochemistry, 84, 53-64. DOI:10.1016/j.soilbio.2015.02.011URL [本文引用: 1] http://linkinghub.elsevier.com/retrieve/pii/S0038071715000565
VětrovskyT, BaldrianP ( 2015). An in-depth analysis of actinobacterial communities shows their high diversity in grassland soils along a gradient of mixed heavy metal contamination Biology and Fertility of Soils, 51, 827-837. DOI:10.1007/s00374-015-1029-9URL [本文引用: 1] Several previous studies indicated that Actinobacteria may be enriched in soils with elevated content of heavy metals. In this study, we have developed a method for the in-depth analysis of actinobacterial communities in soil through phylum-targeted high-throughput sequencing and used it to address this question and examine the community composition in grassland soils along a gradient of heavy metal contamination (Cu, Zn, Cd, Pb). The use of the 16Sact111r primer specific for Actinobacteria resulted in a dataset obtained by pyrosequencing where over 98 % of the sequences belonged to Actinobacteria. The diversity within the Actinobacterial community was not affected by the heavy metals, but the contamination was the most important factor affecting community composition. The most significant changes in community composition were due to the content of Cu and Pb, while the effects of Zn and Cd were relatively minor. For the most abundant actinobacterial taxa, the abundance of taxa identified as members of the genera Acidothermus, Streptomyces, Pseudonocardia, Janibacter and Microlunatus increased with increasing metal content, while those belonging to Jatrophihabitans and Actinoallomurus decreased. The genus Ilumatobacter contained operational taxonomic units (OTUs) that responded to heavy metals both positively and negatively. This study also confirmed that Actinobacteria appear to be less affected by heavy metals than other bacteria. Because several Actinobacteria were also identified in playing a significant role in cellulose and lignocellulose decomposition in soil, they potentially represent important decomposers of organic matter in such environments.
WardleDA, BardgettRD, KlironomosJN, Set?l?H, van der PuttenWH, WallDH ( 2004). Ecological linkages between aboveground and belowground biota Science, 304, 1629-1633. DOI:10.1126/science.1094875URLPMID:15192218 [本文引用: 2] All terrestrial ecosystems consist of aboveground and belowground components that interact to influence community- and ecosystem-level processes and properties. Here we show how these components are closely interlinked at the community level, reinforced by a greater degree of specificity between plants and soil organisms than has been previously supposed. As such, aboveground and belowground communities can be powerful mutual drivers, with both positive and negative feedbacks. A combined aboveground-belowground approach to community and ecosystem ecology is enhancing our understanding of the regulation and functional significance of biodiversity and of the environmental impacts of human-induced global change phenomena.
WymoreAS, SalpasE, CasaburiG, LiuCM, PriceLB, HungateBA, McDowellWH, MarksJC ( 2018). Effects of plant species on stream bacterial communities via leachate from leaf litter Hydrobiologia, 807, 131-144. DOI:10.1007/s10750-017-3386-xURL [本文引用: 3] Leaf litter provides an important resource to forested stream ecosystems. During leaf fall a significant amount of dissolved organic carbon (DOC) enters streams as leaf leachate. We compared the effec
ZhangCH, ZhangLM, LiuXR, XinXP, LiSG ( 2011). Root tissue and shoot litter decomposition of dominant species Stipa baicalensis in Hulun Buir meadow steppe of Inner Mongolia, China. Chinese Journal of Plant Ecology, 35, 1156-1166. DOI:10.3724/SP.J.1258.2011.01156URL [本文引用: 2] 采用凋落物分解袋法,研究了呼伦贝尔草甸草原主要优势种贝加尔针茅(Stipa baicalensis)根系组织和地上部分凋落物分解的季节动态以及凋落物的放置位置(置于地表和15cm土壤表层)对分解的影响。结果表明,置于表层土壤中的根系组织和地上部分凋落物的分解速率比置于地表的快,但是根系组织在两个放置位置分解的差异不显著。无论置于地表还是置于表层土壤中,地上部分凋落物的分解均快于根系组织的凋落物分解。在分解过程中,凋落物碳(C)损失的季节变化模式与重量损失相似;而氮(N)变化模式明显不同,地上部分凋落物表现为释放—累积—释放,根系则表现为释放—累积,并且地上部分或者根系在不同放置位置中N含量变化的差异较小。地上部分和根系组织凋落物的初始化学组成的差异可能是导致其分解过程差异显著的主要原因,其次的原因才是土壤含水量。因此,该地区未来环境温度、湿度因子的变化将会显著影响贝加尔针茅地上部分凋落物的分解过程,而对根系组织凋落物的分解作用较小。 [ 张彩虹, 张雷明, 刘杏认, 辛晓平, 李胜功 ( 2011). 呼伦贝尔草甸草原优势种贝加尔针茅根系组织和地上部分凋落物的分解 植物生态学报, 35, 1156-1166.] DOI:10.3724/SP.J.1258.2011.01156URL [本文引用: 2] 采用凋落物分解袋法,研究了呼伦贝尔草甸草原主要优势种贝加尔针茅(Stipa baicalensis)根系组织和地上部分凋落物分解的季节动态以及凋落物的放置位置(置于地表和15cm土壤表层)对分解的影响。结果表明,置于表层土壤中的根系组织和地上部分凋落物的分解速率比置于地表的快,但是根系组织在两个放置位置分解的差异不显著。无论置于地表还是置于表层土壤中,地上部分凋落物的分解均快于根系组织的凋落物分解。在分解过程中,凋落物碳(C)损失的季节变化模式与重量损失相似;而氮(N)变化模式明显不同,地上部分凋落物表现为释放—累积—释放,根系则表现为释放—累积,并且地上部分或者根系在不同放置位置中N含量变化的差异较小。地上部分和根系组织凋落物的初始化学组成的差异可能是导致其分解过程差异显著的主要原因,其次的原因才是土壤含水量。因此,该地区未来环境温度、湿度因子的变化将会显著影响贝加尔针茅地上部分凋落物的分解过程,而对根系组织凋落物的分解作用较小。
ZhangYG, CongJ, LuH, YangCY, YangYF, ZhouJZ, LiDQ ( 2014). An integrated study to analyze soil microbial community structure and metabolic potential in two forest types PLOS ONE, 9, e93773. DOI: 10.1371/journal.pone.0093773. DOI:10.1371/journal.pone.0093773URLPMID:24743581 [本文引用: 1] Soil microbial metabolic potential and ecosystem function have received little attention owing to difficulties in methodology. In this study, we selected natural mature forest and natural secondary forest and analyzed the soil microbial community and metabolic potential combing the high-throughput sequencing and GeoChip technologies. Phylogenetic analysis based on 16S rRNA sequencing showed that one known archaeal phylum and 15 known bacterial phyla as well as unclassified phylotypes were presented in these forest soils, and Acidobacteria, Protecobacteria, and Actinobacteria were three of most abundant phyla. The detected microbial functional gene groups were related to different biogeochemical processes, including carbon degradation, carbon fixation, methane metabolism, nitrogen cycling, phosphorus utilization, sulfur cycling, etc. The Shannon index for detected functional gene probes was significantly higher (P<0.05) at natural secondary forest site. The regression analysis showed that a strong positive (P<0.05) correlation was existed between the soil microbial functional gene diversity and phylogenetic diversity. Mantel test showed that soil oxidizable organic carbon, soil total nitrogen and cellulose, glucanase, and amylase activities were significantly linked (P<0.05) to the relative abundance of corresponded functional gene groups. Variance partitioning analysis showed that a total of 81.58% of the variation in community structure was explained by soil chemical factors, soil temperature, and plant diversity. Therefore, the positive link of soil microbial structure and composition to functional activity related to ecosystem functioning was existed, and the natural secondary forest soil may occur the high microbial metabolic potential. Although the results can't directly reflect the actual microbial populations and functional activities, this study provides insight into the potential activity of the microbial community and associated feedback responses of the terrestrial ecosystem to environmental changes.
ZhaoBY, XingP, WuQL ( 2017). Microbes participated in macrophyte leaf litters decomposition in freshwater habitat FEMS Microbiology Ecology, 93, fix108. DOI: 10.1093/ femsec/fix108. DOI:10.1093/femsec/fix108URLPMID:28961908 [本文引用: 3] Abstract Knowledge of aquatic microbes involved in macrophyte leaf litter decomposition is still scarce in freshwater lakes. In situ experiments (150 days) were conducted to study the decomposition processes of macrophyte leaf litters: Zizania latifolia (Zl), Hydrilla verticillata (Hv), and Nymphoides peltata (Np). The decomposition of Np leaf litter was fastest whereas Zl was slowest. The alpha diversity of both bacterial and fungal communities significantly increased, and their community structures showed significant variations over time. For bacteria, the relative abundance of Gammaproteobacteria decreased, whereas that of Firmicutes, Betaproteobacteria, Deltaproteobacteria, and Alphaproteobacteria increased. The dominant fungal phylum Cryptomycota increased significantly in all of the three macrophytes. Both bacteria and fungi were significantly correlated with the dynamics of total phosphorous in the water and the carbon content of the leaf litters. The dynamics of nitrogen content, phosphorous content, as well as N/P ratio of the leaf litters have more influences on fungal communities than on bacteria. In addition, cellulase and xylanase activities were significantly correlated with bacterial and fungal communities, respectively, thereby reflecting the niches differentiation and cooperation between bacteria and fungi on litter decomposition. This work contributes to the understanding of microbially involved carbon and nutrient cycling in macrophyte-dominated freshwater ecosystems. FEMS 2017. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
ZhaoYY, WuFZ, YangWQ, TanB, HeW ( 2016). Variations in bacterial communities during Foliar litter decomposition in the winter and growing seasons in an alpine forest of the eastern Tibetan Plateau Canadian Journal of Microbiology, 62, 1-14. DOI:10.1139/cjm-2015-0350URLPMID:26553381 [本文引用: 3] Abstract Retroviruses must integrate their cDNA into the host genome to generate proviruses. Viral DNA-protein complexes interact with cellular proteins and produce pre-integration complexes, which carry the viral genome and cross the nuclear pore channel to enter the nucleus and integrate viral DNA into host chromosomal DNA. If the reverse transcripts fail to integrate, linear or circular DNA species such as 1- and 2-long terminal repeats are generated. Such complexes encounter numerous cellular proteins in the cytoplasm, which restrict viral infection and protect the nucleus. To overcome host cell defenses, the pathogens have evolved several evasion strategies. Viral proteins often contain nuclear localization signals, allowing entry into the nucleus. Among more than 1000 proteins identified as required for HIV infection by RNA interference screening, karyopherins, cleavage and polyadenylation specific factor 6, and nucleoporins have been predominantly studied. This review discusses current opinions about the synergistic relationship between the viral and cellular factors involved in nuclear import, with focus on the unveiled mysteries of the host-pathogen interaction, and highlights novel approaches to pinpoint therapeutic targets. Contrasting rRNA gene abundance patterns for aquatic fungi and bacteria in response to leaf-litter chemistry 1 2013
... 凋落物形成后, 通过物理过程、非生物化学过程、生物过程进行分解, 前人的研究发现, 在大的空间尺度上, 气候条件在控制凋落物分解中起更加显著的作用(Meentemeyer, 1978; Tuomi et al., 2009); 在小尺度上, 凋落物质量(如氮(N)含量、C:N、木质素含量等)和微生物群落组成是主要的影响因素(Taylor et al., 1989; Wymore et al., 2018).近年来, 研究表明微生物能代谢和转化叶片C形成微生物生物量(Gessner et al., 1999), 在凋落物分解过程中发挥着不可替代的作用(Sun et al., 2017).关于凋落物分解过程中微生物的研究主要集中在土壤微生物群落结构(Sun et al., 2017), 而对分解过程中凋落物中微生物群落结构的研究涉及较少(张明锦等, 2015).在土壤环境中细菌是数量最大、种类最多的微生物, 其生物学特性各异, 功能多样, 繁殖迅速, 对土壤肥力的形成、植物营养的转化有极其重要的作用(Kennedy, 1999; 丁新景等, 2017).早期的研究结果显示, 细菌对凋落物分解的作用方式与真菌有明显差别, 在分解的初始阶段主要是真菌起作用, 而分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).随后基于荧光定量PCR的研究表明, 细菌在凋落物分解早期同样具有重要作用, 前期的研究低估了细菌在凋落物分解中的作用(Adam et al., 2013; 张明锦等, 2015).且前人的结论主要是基于叶片凋落物基质, 而对于根系分解过程中微生物群落结构的变化尚未见报道. ...
Functional genomics analysis of plant growth-promoting rhizobacterial traits involved in rhizosphere competence 1 2011
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
What type of diversity yields synergy during mixed litter decomposition in a natural forest ecosystem? 3 2007
... 森林凋落物的累积和分解是森林生态系统养分循环的重要环节, 是光合作用固定碳(C)返回到大气的主要途径(Co?teaux et al., 1995), 并且维持生态系统中养分循环过程, 为土壤动物、微生物提供食物来源, 为植物生长提供必要的养分(张彩虹等, 2011), 长期以来受到研究者的广泛关注(张彩虹等, 2011; Gui et al., 2017; 罗永清等, 2017).近10年, 对森林叶片凋落物分解的研究大量涌现(Chapman & Koch, 2007; Wymore et al., 2018), 但是, 根系分解往往被忽略.相比于对地上组织分解的研究, 特殊的位置导致对根系凋落物的研究相对滞后, 但是植物根系周转迅速, 特别是细根, 其周转速度更快, 约占总净初级生产量的33% (罗永清等, 2017; Shen et al., 2017), 且根系凋落物数量庞大, 其更新过程可产生5倍于地上凋落物的凋落量(Grier et al., 1981).可见, 对根系凋落物分解的研究对整个森林生态系统土壤有机质的形成和养分循环至关重要. ...
... 凋落物形成后, 通过物理过程、非生物化学过程、生物过程进行分解, 前人的研究发现, 在大的空间尺度上, 气候条件在控制凋落物分解中起更加显著的作用(Meentemeyer, 1978; Tuomi et al., 2009); 在小尺度上, 凋落物质量(如氮(N)含量、C:N、木质素含量等)和微生物群落组成是主要的影响因素(Taylor et al., 1989; Wymore et al., 2018).近年来, 研究表明微生物能代谢和转化叶片C形成微生物生物量(Gessner et al., 1999), 在凋落物分解过程中发挥着不可替代的作用(Sun et al., 2017).关于凋落物分解过程中微生物的研究主要集中在土壤微生物群落结构(Sun et al., 2017), 而对分解过程中凋落物中微生物群落结构的研究涉及较少(张明锦等, 2015).在土壤环境中细菌是数量最大、种类最多的微生物, 其生物学特性各异, 功能多样, 繁殖迅速, 对土壤肥力的形成、植物营养的转化有极其重要的作用(Kennedy, 1999; 丁新景等, 2017).早期的研究结果显示, 细菌对凋落物分解的作用方式与真菌有明显差别, 在分解的初始阶段主要是真菌起作用, 而分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).随后基于荧光定量PCR的研究表明, 细菌在凋落物分解早期同样具有重要作用, 前期的研究低估了细菌在凋落物分解中的作用(Adam et al., 2013; 张明锦等, 2015).且前人的结论主要是基于叶片凋落物基质, 而对于根系分解过程中微生物群落结构的变化尚未见报道. ...
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
The addition of labile carbon alters litter fungal communities and decreases litter decomposition rates 1 2009
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
Litter decomposition, climate and litter quality 2 1995
... 森林凋落物的累积和分解是森林生态系统养分循环的重要环节, 是光合作用固定碳(C)返回到大气的主要途径(Co?teaux et al., 1995), 并且维持生态系统中养分循环过程, 为土壤动物、微生物提供食物来源, 为植物生长提供必要的养分(张彩虹等, 2011), 长期以来受到研究者的广泛关注(张彩虹等, 2011; Gui et al., 2017; 罗永清等, 2017).近10年, 对森林叶片凋落物分解的研究大量涌现(Chapman & Koch, 2007; Wymore et al., 2018), 但是, 根系分解往往被忽略.相比于对地上组织分解的研究, 特殊的位置导致对根系凋落物的研究相对滞后, 但是植物根系周转迅速, 特别是细根, 其周转速度更快, 约占总净初级生产量的33% (罗永清等, 2017; Shen et al., 2017), 且根系凋落物数量庞大, 其更新过程可产生5倍于地上凋落物的凋落量(Grier et al., 1981).可见, 对根系凋落物分解的研究对整个森林生态系统土壤有机质的形成和养分循环至关重要. ...
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
黄河三角洲刺槐根际与非根际细菌结构及多样性 3 2017
... 凋落物形成后, 通过物理过程、非生物化学过程、生物过程进行分解, 前人的研究发现, 在大的空间尺度上, 气候条件在控制凋落物分解中起更加显著的作用(Meentemeyer, 1978; Tuomi et al., 2009); 在小尺度上, 凋落物质量(如氮(N)含量、C:N、木质素含量等)和微生物群落组成是主要的影响因素(Taylor et al., 1989; Wymore et al., 2018).近年来, 研究表明微生物能代谢和转化叶片C形成微生物生物量(Gessner et al., 1999), 在凋落物分解过程中发挥着不可替代的作用(Sun et al., 2017).关于凋落物分解过程中微生物的研究主要集中在土壤微生物群落结构(Sun et al., 2017), 而对分解过程中凋落物中微生物群落结构的研究涉及较少(张明锦等, 2015).在土壤环境中细菌是数量最大、种类最多的微生物, 其生物学特性各异, 功能多样, 繁殖迅速, 对土壤肥力的形成、植物营养的转化有极其重要的作用(Kennedy, 1999; 丁新景等, 2017).早期的研究结果显示, 细菌对凋落物分解的作用方式与真菌有明显差别, 在分解的初始阶段主要是真菌起作用, 而分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).随后基于荧光定量PCR的研究表明, 细菌在凋落物分解早期同样具有重要作用, 前期的研究低估了细菌在凋落物分解中的作用(Adam et al., 2013; 张明锦等, 2015).且前人的结论主要是基于叶片凋落物基质, 而对于根系分解过程中微生物群落结构的变化尚未见报道. ...
... 长期以来, 由于受技术条件的限制, 对微生物的研究结果不能真实、全面地反映各生态系统中微生物群落多样性和结构及微生物不同类型所起的作用, 特别是在凋落物分解过程中(荣丽等, 2009).高通量测序技术打破了原有技术的局限, 无需分离单一菌株, 直接在基因水平上挖掘环境中的微生物信息, 具有成本低、通量高、信息丰富等优点.近年来, 已经广泛用于不同生态系统微生物群落结构的测定, 如空气(孙翠丽等, 2017)、土壤(丁新景等, 2017; Sun et al., 2017)、水域(Zhao et al., 2017)等.但是高通量测序技术在凋落物分解的研究中应用相对较少, 特别是细根分解中的应用更少.因此, 本研究采用高通量测序技术, 研究了泰山4种优势造林树种细根分解过程中细菌群落组成, 对比分析了4个树种细菌优势种群、细菌多样性的差异, 同时揭示了细根分解速率与初始化学性质和细菌群落结构之间的相关关系, 为研究细根凋落物分解的微生物控制机制提供理论依据. ...
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
黄河三角洲刺槐根际与非根际细菌结构及多样性 3 2017
... 凋落物形成后, 通过物理过程、非生物化学过程、生物过程进行分解, 前人的研究发现, 在大的空间尺度上, 气候条件在控制凋落物分解中起更加显著的作用(Meentemeyer, 1978; Tuomi et al., 2009); 在小尺度上, 凋落物质量(如氮(N)含量、C:N、木质素含量等)和微生物群落组成是主要的影响因素(Taylor et al., 1989; Wymore et al., 2018).近年来, 研究表明微生物能代谢和转化叶片C形成微生物生物量(Gessner et al., 1999), 在凋落物分解过程中发挥着不可替代的作用(Sun et al., 2017).关于凋落物分解过程中微生物的研究主要集中在土壤微生物群落结构(Sun et al., 2017), 而对分解过程中凋落物中微生物群落结构的研究涉及较少(张明锦等, 2015).在土壤环境中细菌是数量最大、种类最多的微生物, 其生物学特性各异, 功能多样, 繁殖迅速, 对土壤肥力的形成、植物营养的转化有极其重要的作用(Kennedy, 1999; 丁新景等, 2017).早期的研究结果显示, 细菌对凋落物分解的作用方式与真菌有明显差别, 在分解的初始阶段主要是真菌起作用, 而分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).随后基于荧光定量PCR的研究表明, 细菌在凋落物分解早期同样具有重要作用, 前期的研究低估了细菌在凋落物分解中的作用(Adam et al., 2013; 张明锦等, 2015).且前人的结论主要是基于叶片凋落物基质, 而对于根系分解过程中微生物群落结构的变化尚未见报道. ...
... 长期以来, 由于受技术条件的限制, 对微生物的研究结果不能真实、全面地反映各生态系统中微生物群落多样性和结构及微生物不同类型所起的作用, 特别是在凋落物分解过程中(荣丽等, 2009).高通量测序技术打破了原有技术的局限, 无需分离单一菌株, 直接在基因水平上挖掘环境中的微生物信息, 具有成本低、通量高、信息丰富等优点.近年来, 已经广泛用于不同生态系统微生物群落结构的测定, 如空气(孙翠丽等, 2017)、土壤(丁新景等, 2017; Sun et al., 2017)、水域(Zhao et al., 2017)等.但是高通量测序技术在凋落物分解的研究中应用相对较少, 特别是细根分解中的应用更少.因此, 本研究采用高通量测序技术, 研究了泰山4种优势造林树种细根分解过程中细菌群落组成, 对比分析了4个树种细菌优势种群、细菌多样性的差异, 同时揭示了细根分解速率与初始化学性质和细菌群落结构之间的相关关系, 为研究细根凋落物分解的微生物控制机制提供理论依据. ...
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
Influence of plant polymers on the distribution and cultivation of bacteria in the phylum Acidobacteria 1 2011
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
Growth rate-stoichiometry couplings in diverse biota 1 2003
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
Toward an ecological classification of soil bacteria 1 2007
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
A perspective on leaf litter breakdown in streams 1 1999
... 凋落物形成后, 通过物理过程、非生物化学过程、生物过程进行分解, 前人的研究发现, 在大的空间尺度上, 气候条件在控制凋落物分解中起更加显著的作用(Meentemeyer, 1978; Tuomi et al., 2009); 在小尺度上, 凋落物质量(如氮(N)含量、C:N、木质素含量等)和微生物群落组成是主要的影响因素(Taylor et al., 1989; Wymore et al., 2018).近年来, 研究表明微生物能代谢和转化叶片C形成微生物生物量(Gessner et al., 1999), 在凋落物分解过程中发挥着不可替代的作用(Sun et al., 2017).关于凋落物分解过程中微生物的研究主要集中在土壤微生物群落结构(Sun et al., 2017), 而对分解过程中凋落物中微生物群落结构的研究涉及较少(张明锦等, 2015).在土壤环境中细菌是数量最大、种类最多的微生物, 其生物学特性各异, 功能多样, 繁殖迅速, 对土壤肥力的形成、植物营养的转化有极其重要的作用(Kennedy, 1999; 丁新景等, 2017).早期的研究结果显示, 细菌对凋落物分解的作用方式与真菌有明显差别, 在分解的初始阶段主要是真菌起作用, 而分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).随后基于荧光定量PCR的研究表明, 细菌在凋落物分解早期同样具有重要作用, 前期的研究低估了细菌在凋落物分解中的作用(Adam et al., 2013; 张明锦等, 2015).且前人的结论主要是基于叶片凋落物基质, 而对于根系分解过程中微生物群落结构的变化尚未见报道. ...
Biomass distribution and above- and below-?ground production in young and mature Abies amabilis zone ecosystems of the Washington Cascades. 1 1981
... 森林凋落物的累积和分解是森林生态系统养分循环的重要环节, 是光合作用固定碳(C)返回到大气的主要途径(Co?teaux et al., 1995), 并且维持生态系统中养分循环过程, 为土壤动物、微生物提供食物来源, 为植物生长提供必要的养分(张彩虹等, 2011), 长期以来受到研究者的广泛关注(张彩虹等, 2011; Gui et al., 2017; 罗永清等, 2017).近10年, 对森林叶片凋落物分解的研究大量涌现(Chapman & Koch, 2007; Wymore et al., 2018), 但是, 根系分解往往被忽略.相比于对地上组织分解的研究, 特殊的位置导致对根系凋落物的研究相对滞后, 但是植物根系周转迅速, 特别是细根, 其周转速度更快, 约占总净初级生产量的33% (罗永清等, 2017; Shen et al., 2017), 且根系凋落物数量庞大, 其更新过程可产生5倍于地上凋落物的凋落量(Grier et al., 1981).可见, 对根系凋落物分解的研究对整个森林生态系统土壤有机质的形成和养分循环至关重要. ...
The arbuscular mycorrhizal fungus Funneliformis mosseae alters bacterial communities in subtropical forest soils during litter decomposition 3 2017
... 森林凋落物的累积和分解是森林生态系统养分循环的重要环节, 是光合作用固定碳(C)返回到大气的主要途径(Co?teaux et al., 1995), 并且维持生态系统中养分循环过程, 为土壤动物、微生物提供食物来源, 为植物生长提供必要的养分(张彩虹等, 2011), 长期以来受到研究者的广泛关注(张彩虹等, 2011; Gui et al., 2017; 罗永清等, 2017).近10年, 对森林叶片凋落物分解的研究大量涌现(Chapman & Koch, 2007; Wymore et al., 2018), 但是, 根系分解往往被忽略.相比于对地上组织分解的研究, 特殊的位置导致对根系凋落物的研究相对滞后, 但是植物根系周转迅速, 特别是细根, 其周转速度更快, 约占总净初级生产量的33% (罗永清等, 2017; Shen et al., 2017), 且根系凋落物数量庞大, 其更新过程可产生5倍于地上凋落物的凋落量(Grier et al., 1981).可见, 对根系凋落物分解的研究对整个森林生态系统土壤有机质的形成和养分循环至关重要. ...
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
... ), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
Multi-omics of permafrost, active layer and thermokarst bog soil microbiomes 1 2015
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
Macroclimate and lignin control of litter decomposition rates 1 1978
... 凋落物形成后, 通过物理过程、非生物化学过程、生物过程进行分解, 前人的研究发现, 在大的空间尺度上, 气候条件在控制凋落物分解中起更加显著的作用(Meentemeyer, 1978; Tuomi et al., 2009); 在小尺度上, 凋落物质量(如氮(N)含量、C:N、木质素含量等)和微生物群落组成是主要的影响因素(Taylor et al., 1989; Wymore et al., 2018).近年来, 研究表明微生物能代谢和转化叶片C形成微生物生物量(Gessner et al., 1999), 在凋落物分解过程中发挥着不可替代的作用(Sun et al., 2017).关于凋落物分解过程中微生物的研究主要集中在土壤微生物群落结构(Sun et al., 2017), 而对分解过程中凋落物中微生物群落结构的研究涉及较少(张明锦等, 2015).在土壤环境中细菌是数量最大、种类最多的微生物, 其生物学特性各异, 功能多样, 繁殖迅速, 对土壤肥力的形成、植物营养的转化有极其重要的作用(Kennedy, 1999; 丁新景等, 2017).早期的研究结果显示, 细菌对凋落物分解的作用方式与真菌有明显差别, 在分解的初始阶段主要是真菌起作用, 而分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).随后基于荧光定量PCR的研究表明, 细菌在凋落物分解早期同样具有重要作用, 前期的研究低估了细菌在凋落物分解中的作用(Adam et al., 2013; 张明锦等, 2015).且前人的结论主要是基于叶片凋落物基质, 而对于根系分解过程中微生物群落结构的变化尚未见报道. ...
光皮桦细根与扁穗牛鞭草草根分解的土壤微生物数量及优势类群 1 2009
... 长期以来, 由于受技术条件的限制, 对微生物的研究结果不能真实、全面地反映各生态系统中微生物群落多样性和结构及微生物不同类型所起的作用, 特别是在凋落物分解过程中(荣丽等, 2009).高通量测序技术打破了原有技术的局限, 无需分离单一菌株, 直接在基因水平上挖掘环境中的微生物信息, 具有成本低、通量高、信息丰富等优点.近年来, 已经广泛用于不同生态系统微生物群落结构的测定, 如空气(孙翠丽等, 2017)、土壤(丁新景等, 2017; Sun et al., 2017)、水域(Zhao et al., 2017)等.但是高通量测序技术在凋落物分解的研究中应用相对较少, 特别是细根分解中的应用更少.因此, 本研究采用高通量测序技术, 研究了泰山4种优势造林树种细根分解过程中细菌群落组成, 对比分析了4个树种细菌优势种群、细菌多样性的差异, 同时揭示了细根分解速率与初始化学性质和细菌群落结构之间的相关关系, 为研究细根凋落物分解的微生物控制机制提供理论依据. ...
光皮桦细根与扁穗牛鞭草草根分解的土壤微生物数量及优势类群 1 2009
... 长期以来, 由于受技术条件的限制, 对微生物的研究结果不能真实、全面地反映各生态系统中微生物群落多样性和结构及微生物不同类型所起的作用, 特别是在凋落物分解过程中(荣丽等, 2009).高通量测序技术打破了原有技术的局限, 无需分离单一菌株, 直接在基因水平上挖掘环境中的微生物信息, 具有成本低、通量高、信息丰富等优点.近年来, 已经广泛用于不同生态系统微生物群落结构的测定, 如空气(孙翠丽等, 2017)、土壤(丁新景等, 2017; Sun et al., 2017)、水域(Zhao et al., 2017)等.但是高通量测序技术在凋落物分解的研究中应用相对较少, 特别是细根分解中的应用更少.因此, 本研究采用高通量测序技术, 研究了泰山4种优势造林树种细根分解过程中细菌群落组成, 对比分析了4个树种细菌优势种群、细菌多样性的差异, 同时揭示了细根分解速率与初始化学性质和细菌群落结构之间的相关关系, 为研究细根凋落物分解的微生物控制机制提供理论依据. ...
The dynamics of soil micro-food web structure and functions vary according to litter quality 1 2016
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
Characteristics of fine roots of Pinus massoniana in the Three Gorges Reservoir Area, China 1 2017
... 森林凋落物的累积和分解是森林生态系统养分循环的重要环节, 是光合作用固定碳(C)返回到大气的主要途径(Co?teaux et al., 1995), 并且维持生态系统中养分循环过程, 为土壤动物、微生物提供食物来源, 为植物生长提供必要的养分(张彩虹等, 2011), 长期以来受到研究者的广泛关注(张彩虹等, 2011; Gui et al., 2017; 罗永清等, 2017).近10年, 对森林叶片凋落物分解的研究大量涌现(Chapman & Koch, 2007; Wymore et al., 2018), 但是, 根系分解往往被忽略.相比于对地上组织分解的研究, 特殊的位置导致对根系凋落物的研究相对滞后, 但是植物根系周转迅速, 特别是细根, 其周转速度更快, 约占总净初级生产量的33% (罗永清等, 2017; Shen et al., 2017), 且根系凋落物数量庞大, 其更新过程可产生5倍于地上凋落物的凋落量(Grier et al., 1981).可见, 对根系凋落物分解的研究对整个森林生态系统土壤有机质的形成和养分循环至关重要. ...
Occurrence and distribution of nitrogen fixing bacterial community associated with oat ( Avena sativa) assessed by molecular and microbiological techniques. 1 2006
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
16S rRNA高通量测序方法检测羊圈空气微生物群落结构及多样性 6 2017
... 凋落物形成后, 通过物理过程、非生物化学过程、生物过程进行分解, 前人的研究发现, 在大的空间尺度上, 气候条件在控制凋落物分解中起更加显著的作用(Meentemeyer, 1978; Tuomi et al., 2009); 在小尺度上, 凋落物质量(如氮(N)含量、C:N、木质素含量等)和微生物群落组成是主要的影响因素(Taylor et al., 1989; Wymore et al., 2018).近年来, 研究表明微生物能代谢和转化叶片C形成微生物生物量(Gessner et al., 1999), 在凋落物分解过程中发挥着不可替代的作用(Sun et al., 2017).关于凋落物分解过程中微生物的研究主要集中在土壤微生物群落结构(Sun et al., 2017), 而对分解过程中凋落物中微生物群落结构的研究涉及较少(张明锦等, 2015).在土壤环境中细菌是数量最大、种类最多的微生物, 其生物学特性各异, 功能多样, 繁殖迅速, 对土壤肥力的形成、植物营养的转化有极其重要的作用(Kennedy, 1999; 丁新景等, 2017).早期的研究结果显示, 细菌对凋落物分解的作用方式与真菌有明显差别, 在分解的初始阶段主要是真菌起作用, 而分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).随后基于荧光定量PCR的研究表明, 细菌在凋落物分解早期同样具有重要作用, 前期的研究低估了细菌在凋落物分解中的作用(Adam et al., 2013; 张明锦等, 2015).且前人的结论主要是基于叶片凋落物基质, 而对于根系分解过程中微生物群落结构的变化尚未见报道. ...
Leaf litter decomposition—Estimates of global variability based on Yasso07 model 1 2009
... 凋落物形成后, 通过物理过程、非生物化学过程、生物过程进行分解, 前人的研究发现, 在大的空间尺度上, 气候条件在控制凋落物分解中起更加显著的作用(Meentemeyer, 1978; Tuomi et al., 2009); 在小尺度上, 凋落物质量(如氮(N)含量、C:N、木质素含量等)和微生物群落组成是主要的影响因素(Taylor et al., 1989; Wymore et al., 2018).近年来, 研究表明微生物能代谢和转化叶片C形成微生物生物量(Gessner et al., 1999), 在凋落物分解过程中发挥着不可替代的作用(Sun et al., 2017).关于凋落物分解过程中微生物的研究主要集中在土壤微生物群落结构(Sun et al., 2017), 而对分解过程中凋落物中微生物群落结构的研究涉及较少(张明锦等, 2015).在土壤环境中细菌是数量最大、种类最多的微生物, 其生物学特性各异, 功能多样, 繁殖迅速, 对土壤肥力的形成、植物营养的转化有极其重要的作用(Kennedy, 1999; 丁新景等, 2017).早期的研究结果显示, 细菌对凋落物分解的作用方式与真菌有明显差别, 在分解的初始阶段主要是真菌起作用, 而分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).随后基于荧光定量PCR的研究表明, 细菌在凋落物分解早期同样具有重要作用, 前期的研究低估了细菌在凋落物分解中的作用(Adam et al., 2013; 张明锦等, 2015).且前人的结论主要是基于叶片凋落物基质, 而对于根系分解过程中微生物群落结构的变化尚未见报道. ...
Composition of fungal and bacterial communities in forest litter and soil is largely determined by dominant trees 1 2015
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
An in-depth analysis of actinobacterial communities shows their high diversity in grassland soils along a gradient of mixed heavy metal contamination 1 2015
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
Ecological linkages between aboveground and belowground biota 2 2004
... 凋落物形成后, 通过物理过程、非生物化学过程、生物过程进行分解, 前人的研究发现, 在大的空间尺度上, 气候条件在控制凋落物分解中起更加显著的作用(Meentemeyer, 1978; Tuomi et al., 2009); 在小尺度上, 凋落物质量(如氮(N)含量、C:N、木质素含量等)和微生物群落组成是主要的影响因素(Taylor et al., 1989; Wymore et al., 2018).近年来, 研究表明微生物能代谢和转化叶片C形成微生物生物量(Gessner et al., 1999), 在凋落物分解过程中发挥着不可替代的作用(Sun et al., 2017).关于凋落物分解过程中微生物的研究主要集中在土壤微生物群落结构(Sun et al., 2017), 而对分解过程中凋落物中微生物群落结构的研究涉及较少(张明锦等, 2015).在土壤环境中细菌是数量最大、种类最多的微生物, 其生物学特性各异, 功能多样, 繁殖迅速, 对土壤肥力的形成、植物营养的转化有极其重要的作用(Kennedy, 1999; 丁新景等, 2017).早期的研究结果显示, 细菌对凋落物分解的作用方式与真菌有明显差别, 在分解的初始阶段主要是真菌起作用, 而分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).随后基于荧光定量PCR的研究表明, 细菌在凋落物分解早期同样具有重要作用, 前期的研究低估了细菌在凋落物分解中的作用(Adam et al., 2013; 张明锦等, 2015).且前人的结论主要是基于叶片凋落物基质, 而对于根系分解过程中微生物群落结构的变化尚未见报道. ...
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
Effects of plant species on stream bacterial communities via leachate from leaf litter 3 2018
... 森林凋落物的累积和分解是森林生态系统养分循环的重要环节, 是光合作用固定碳(C)返回到大气的主要途径(Co?teaux et al., 1995), 并且维持生态系统中养分循环过程, 为土壤动物、微生物提供食物来源, 为植物生长提供必要的养分(张彩虹等, 2011), 长期以来受到研究者的广泛关注(张彩虹等, 2011; Gui et al., 2017; 罗永清等, 2017).近10年, 对森林叶片凋落物分解的研究大量涌现(Chapman & Koch, 2007; Wymore et al., 2018), 但是, 根系分解往往被忽略.相比于对地上组织分解的研究, 特殊的位置导致对根系凋落物的研究相对滞后, 但是植物根系周转迅速, 特别是细根, 其周转速度更快, 约占总净初级生产量的33% (罗永清等, 2017; Shen et al., 2017), 且根系凋落物数量庞大, 其更新过程可产生5倍于地上凋落物的凋落量(Grier et al., 1981).可见, 对根系凋落物分解的研究对整个森林生态系统土壤有机质的形成和养分循环至关重要. ...
... 凋落物形成后, 通过物理过程、非生物化学过程、生物过程进行分解, 前人的研究发现, 在大的空间尺度上, 气候条件在控制凋落物分解中起更加显著的作用(Meentemeyer, 1978; Tuomi et al., 2009); 在小尺度上, 凋落物质量(如氮(N)含量、C:N、木质素含量等)和微生物群落组成是主要的影响因素(Taylor et al., 1989; Wymore et al., 2018).近年来, 研究表明微生物能代谢和转化叶片C形成微生物生物量(Gessner et al., 1999), 在凋落物分解过程中发挥着不可替代的作用(Sun et al., 2017).关于凋落物分解过程中微生物的研究主要集中在土壤微生物群落结构(Sun et al., 2017), 而对分解过程中凋落物中微生物群落结构的研究涉及较少(张明锦等, 2015).在土壤环境中细菌是数量最大、种类最多的微生物, 其生物学特性各异, 功能多样, 繁殖迅速, 对土壤肥力的形成、植物营养的转化有极其重要的作用(Kennedy, 1999; 丁新景等, 2017).早期的研究结果显示, 细菌对凋落物分解的作用方式与真菌有明显差别, 在分解的初始阶段主要是真菌起作用, 而分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).随后基于荧光定量PCR的研究表明, 细菌在凋落物分解早期同样具有重要作用, 前期的研究低估了细菌在凋落物分解中的作用(Adam et al., 2013; 张明锦等, 2015).且前人的结论主要是基于叶片凋落物基质, 而对于根系分解过程中微生物群落结构的变化尚未见报道. ...
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
呼伦贝尔草甸草原优势种贝加尔针茅根系组织和地上部分凋落物的分解 2 2011
... 森林凋落物的累积和分解是森林生态系统养分循环的重要环节, 是光合作用固定碳(C)返回到大气的主要途径(Co?teaux et al., 1995), 并且维持生态系统中养分循环过程, 为土壤动物、微生物提供食物来源, 为植物生长提供必要的养分(张彩虹等, 2011), 长期以来受到研究者的广泛关注(张彩虹等, 2011; Gui et al., 2017; 罗永清等, 2017).近10年, 对森林叶片凋落物分解的研究大量涌现(Chapman & Koch, 2007; Wymore et al., 2018), 但是, 根系分解往往被忽略.相比于对地上组织分解的研究, 特殊的位置导致对根系凋落物的研究相对滞后, 但是植物根系周转迅速, 特别是细根, 其周转速度更快, 约占总净初级生产量的33% (罗永清等, 2017; Shen et al., 2017), 且根系凋落物数量庞大, 其更新过程可产生5倍于地上凋落物的凋落量(Grier et al., 1981).可见, 对根系凋落物分解的研究对整个森林生态系统土壤有机质的形成和养分循环至关重要. ...
... ), 长期以来受到研究者的广泛关注(张彩虹等, 2011; Gui et al., 2017; 罗永清等, 2017).近10年, 对森林叶片凋落物分解的研究大量涌现(Chapman & Koch, 2007; Wymore et al., 2018), 但是, 根系分解往往被忽略.相比于对地上组织分解的研究, 特殊的位置导致对根系凋落物的研究相对滞后, 但是植物根系周转迅速, 特别是细根, 其周转速度更快, 约占总净初级生产量的33% (罗永清等, 2017; Shen et al., 2017), 且根系凋落物数量庞大, 其更新过程可产生5倍于地上凋落物的凋落量(Grier et al., 1981).可见, 对根系凋落物分解的研究对整个森林生态系统土壤有机质的形成和养分循环至关重要. ...
呼伦贝尔草甸草原优势种贝加尔针茅根系组织和地上部分凋落物的分解 2 2011
... 森林凋落物的累积和分解是森林生态系统养分循环的重要环节, 是光合作用固定碳(C)返回到大气的主要途径(Co?teaux et al., 1995), 并且维持生态系统中养分循环过程, 为土壤动物、微生物提供食物来源, 为植物生长提供必要的养分(张彩虹等, 2011), 长期以来受到研究者的广泛关注(张彩虹等, 2011; Gui et al., 2017; 罗永清等, 2017).近10年, 对森林叶片凋落物分解的研究大量涌现(Chapman & Koch, 2007; Wymore et al., 2018), 但是, 根系分解往往被忽略.相比于对地上组织分解的研究, 特殊的位置导致对根系凋落物的研究相对滞后, 但是植物根系周转迅速, 特别是细根, 其周转速度更快, 约占总净初级生产量的33% (罗永清等, 2017; Shen et al., 2017), 且根系凋落物数量庞大, 其更新过程可产生5倍于地上凋落物的凋落量(Grier et al., 1981).可见, 对根系凋落物分解的研究对整个森林生态系统土壤有机质的形成和养分循环至关重要. ...
... ), 长期以来受到研究者的广泛关注(张彩虹等, 2011; Gui et al., 2017; 罗永清等, 2017).近10年, 对森林叶片凋落物分解的研究大量涌现(Chapman & Koch, 2007; Wymore et al., 2018), 但是, 根系分解往往被忽略.相比于对地上组织分解的研究, 特殊的位置导致对根系凋落物的研究相对滞后, 但是植物根系周转迅速, 特别是细根, 其周转速度更快, 约占总净初级生产量的33% (罗永清等, 2017; Shen et al., 2017), 且根系凋落物数量庞大, 其更新过程可产生5倍于地上凋落物的凋落量(Grier et al., 1981).可见, 对根系凋落物分解的研究对整个森林生态系统土壤有机质的形成和养分循环至关重要. ...
林窗干扰对凋落叶分解过程中微生物生物量和细菌群落结构的影响 1 2016
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
林窗干扰对凋落叶分解过程中微生物生物量和细菌群落结构的影响 1 2016
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
林窗对凋落物分解过程中细菌群落结构和多样性的影响 5 2015
... 凋落物形成后, 通过物理过程、非生物化学过程、生物过程进行分解, 前人的研究发现, 在大的空间尺度上, 气候条件在控制凋落物分解中起更加显著的作用(Meentemeyer, 1978; Tuomi et al., 2009); 在小尺度上, 凋落物质量(如氮(N)含量、C:N、木质素含量等)和微生物群落组成是主要的影响因素(Taylor et al., 1989; Wymore et al., 2018).近年来, 研究表明微生物能代谢和转化叶片C形成微生物生物量(Gessner et al., 1999), 在凋落物分解过程中发挥着不可替代的作用(Sun et al., 2017).关于凋落物分解过程中微生物的研究主要集中在土壤微生物群落结构(Sun et al., 2017), 而对分解过程中凋落物中微生物群落结构的研究涉及较少(张明锦等, 2015).在土壤环境中细菌是数量最大、种类最多的微生物, 其生物学特性各异, 功能多样, 繁殖迅速, 对土壤肥力的形成、植物营养的转化有极其重要的作用(Kennedy, 1999; 丁新景等, 2017).早期的研究结果显示, 细菌对凋落物分解的作用方式与真菌有明显差别, 在分解的初始阶段主要是真菌起作用, 而分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).随后基于荧光定量PCR的研究表明, 细菌在凋落物分解早期同样具有重要作用, 前期的研究低估了细菌在凋落物分解中的作用(Adam et al., 2013; 张明锦等, 2015).且前人的结论主要是基于叶片凋落物基质, 而对于根系分解过程中微生物群落结构的变化尚未见报道. ...
Variations in bacterial communities during Foliar litter decomposition in the winter and growing seasons in an alpine forest of the eastern Tibetan Plateau 3 2016
... 研究表明细根分解后期主要是耐受能力更强的细菌起作用(Wardle et al., 2004; Chapman & Koch, 2007).4种细根分解一年后的细菌群落多样性水平较高, 且存在显著差异(表2).张明锦等(2015)对马尾松林叶凋落物分解一年时的细菌群落研究结果显示, 不同树种的Shannon-Wiener指数处于1-4之间, 明显低于我们的结果.一方面, 可能是因为分解组织性质差异,导致两者养分有效性存在差异, 特别是P含量(马尾松针叶为0.045%), 显著低于我们的研究结果(表1), 底物有效性较差, 所能支持的微生物类群及数量相对较少(Taylor et al., 1989).其次, 我们的研究针对细根凋落物, 分解组织放置于土壤15 cm处, 而该研究针对叶凋落物放置于土壤表面, 这就造成分解环境的差异, 土壤环境变化较小, 湿度较大, 这都有利于微生物生长.叶片凋落物分解的研究结果发现针叶凋落物内细菌多样性低于阔叶凋落物(张明锦等, 2015), 我们对细根凋落物的分解结果刚好与其相反, 阔叶树种(刺槐和麻栎)根系分解中细菌群落多样性显著低于针叶树种(赤松和油松)(表2).细菌主要是通过分解凋落物而获取C和N (张明锦等, 2015), 阔叶树种根系分解速度较快(图1), 分解一年后, 养分释放速度快, 可溶性物质含量减少多, 能支持的微生物类群及数量相对较少(Taylor et al., 1989).另外, 可能的原因是刺槐和麻栎的优势菌群变形菌门的丰度显著高于油松和赤松(图4), 其对资源的消耗增加, 细菌群落竞争加剧, 使得其他细菌丰度降低, 细菌多样性较低(丁新景等, 2017).ANOSIM非参数检验结果也显示阔叶树种与针叶树种细根分解的细菌群落结构之间存在极显著差异(r = 0.798 1; p = 0.002), 我们的结果(图4)与之一致.这也可能是导致阔叶树种根分解速率与针叶树种根分解速率存在差异的重要原因(图1).分解者的活动和凋落物分解速度很大程度上依赖于凋落物质量(Wymore et al., 2018).阔叶树种细根分解速率显著高于针叶树种(p < 0.05), 分解快慢表现为刺槐>麻栎>油松>赤松(图1).细根分解速率与初始C含量、木质素含量和C:N显著负相关, 与初始N含量、P含量和N:P显著正相关(表4), 这与前人的研究结果一致(Co?teaux et al., 1995; McLaren & Turkington, 2010; Zhao et al., 2017).阔叶树种初始凋落物质量显著高于针叶树种(表1), 有利于加速打开细菌能量通道, 促进变形菌门的生长(Sauvadet et al., 2016).这有利于加速凋落物分解过程中的C、N循环(Kersters et al., 2006)和分解速率(表4).分解一年后, 变形菌门、放线菌门、拟杆菌门和酸杆菌门是主要的优势细菌门, 尤其是变形菌门平均占整个细菌群落的63.3%.前人的结果显示, 变形菌门、酸杆菌门和放线菌门是最丰富和活跃的分解者(Gui et al., 2017), 在北方和温带森林, 这些细菌门在土壤有机层丰富度最大(Zhang et al., 2014), 且其中大多数细菌能降解凋落物中难分解的C (Barret et al., 2011).变形菌门被认为是一类富营养菌, 在分解中占优势(Sun et al., 2017), 是土壤中最常见和最丰富的细菌(Gui et al., 2017).阔叶树凋落物产生的富营养生境有利于其变形菌门的生长, 导致阔叶树种根分解的变形菌门显著高于针叶树种(图4B).P是DNA复制和转录的重要元素, P限制将直接影响细菌群落的生长(Elser et al., 2003).N元素的缺乏会对微生物的生长及活性产生负面影响(Taylor et al., 1989), 当细根分解过程中木质素含量高且土壤中可利用N、P含量不足时, 分解微生物的多样性将会下降(Chigineva et al., 2009).分解一年后, 变形菌门相对丰度与细根初始P含量呈显著正相关, 与N含量正相关关系(表4; 图5), 这也与变形菌门需要富营养来满足其需求相一致.放线菌门的细菌具有细丝状的菌丝, 这些菌丝拥有较强的次生代谢功能, 在有机质分解中起重要作用(Větrovsky & Baldrian, 2015).放线菌门相对丰度与初始P含量呈显著负相关关系(表4), 放线菌纲相对丰度与初始C:N呈显著正相关关系(表5), 这与Urbanová (2015)的研究一致.放线菌门大部分属于腐生菌, 部分细菌种群具有分解木质素和纤维素的能力(Zhao et al., 2016), 但是我们的结果并未发现放线菌门(表4)、放线菌纲(表5)与木质素具有相关性, 可能是降解木质素的种群相对丰度较低导致的.但是, 放线菌门相对丰度与分解速率之间并没有明显的相关性(表4), 可能是由于其相对丰度远低于变形菌门(图4).拟杆菌门的细菌能破坏复杂的生物大分子, 因此其与木质纤维素凋落物的分解相关(Lydell et al., 2004).前人的研究表明, 拟杆菌门的相对丰度在阔叶树种凋落物中比在针叶树种凋落物中高(Sun et al., 2017), 我们的结果(图4B)与之不一致.可能的原因是, 该研究通过模拟实验关注凋落物添加后土壤内细菌的变化, 而我们的研究着眼于凋落物内细菌的变化.酸杆菌门被认为是寡营养和嗜酸的细菌, 能生长在复杂的聚合物中, 包括植物半纤维素或纤维素和真菌壳(Eichorst et al., 2011).酸杆菌门在寡营养环境下丰度较高(Jones et al., 2009), 有研究发现酸杆菌门的丰度与土壤有机碳含量和pH值呈显著负相关关系(Jones et al., 2009), 但是, Fierer等(2007)的研究表明土壤酸杆菌相对丰度与土壤有机碳含量和土壤C:N呈显著正相关关系, 而我们并未发现细根分解物内酸杆菌相对丰度与初始化学性质之间具有明显的相关性(表4).这可能是因为研究对象的差异, 该研究主要针对土壤酸杆菌, 而我们的研究涉及细根凋落物内酸杆菌, 两个研究对象养分有效性不同, 从而导致所能支持的微生物类群和数量的差异(Taylor et al., 1989).其次, 我们的研究主要是在固定站点, 样品量少, 这也可能是一个主要原因.在纲水平上, α-变形菌纲、β-变形菌纲、γ-变形菌纲、不明放线菌纲、鞘脂杆菌纲丰度较高, 且α-变形菌纲和不明放线菌纲在不同树种之间差异显著(图4A, p < 0.05).张明锦(2016)的结果显示α-变形菌纲在不同的根系中差异显著, 我们的结果与之一致.刺槐内α-变形菌纲的丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与初始N含量呈显著正相关关系(表5).研究显示, α-变形菌包括大量参与C、N循环的细菌, 以及与植物共生的细菌(如根瘤菌属)(Soares et al., 2006).这也是豆科植物刺槐根系凋落物内α-变形菌纲相对丰度显著高于其他3个树种(图4A), α-变形菌纲相对丰度与凋落物分解速率呈显著正相关关系(表5)的原因.γ-变形菌中绝大多数为假单胞菌(Pseudomonas), 其具有很强的分解能力, 增殖速度快.β-变形菌包括很多好氧或兼性细菌, 通常其降解能力可变.鞘脂杆菌纲有利于凋落物中纤维素的降解, 放线菌纲能促使凋落物迅速腐烂, 是几丁质和木质素分解的重要参与者(Zhao et al., 2016), γ-变形菌纲中的某些种群在磷元素释放中具有重要作用(Zhao et al., 2016), 这些菌群对生态系统C、N循环有重要的作用.Zhao等(2016)的研究结果显示, 川西亚高山森林叶凋落物分解过程中细菌群落以γ-变形菌纲、鞘脂杆菌纲和黄杆菌纲为主要类群.我们的结果与之存在差异, 主要原因可能是: 1)凋落物基质不同: 我们的研究是细根分解, 而该研究是针对叶片凋落物; 2)分解微环境存在差异: 该研究所处环境是亚高山地区, 且分解置于土壤表面, 而我们的实验在暖温带地区, 且放置于土壤中; 3)测定微生物的手段不同.该研究采用PCR-DGGE, 而我们采用高通量测序技术. ...
 ,*黄河下游森林培育国家林业局重点实验室, 山东泰安 271018
,*黄河下游森林培育国家林业局重点实验室, 山东泰安 271018
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT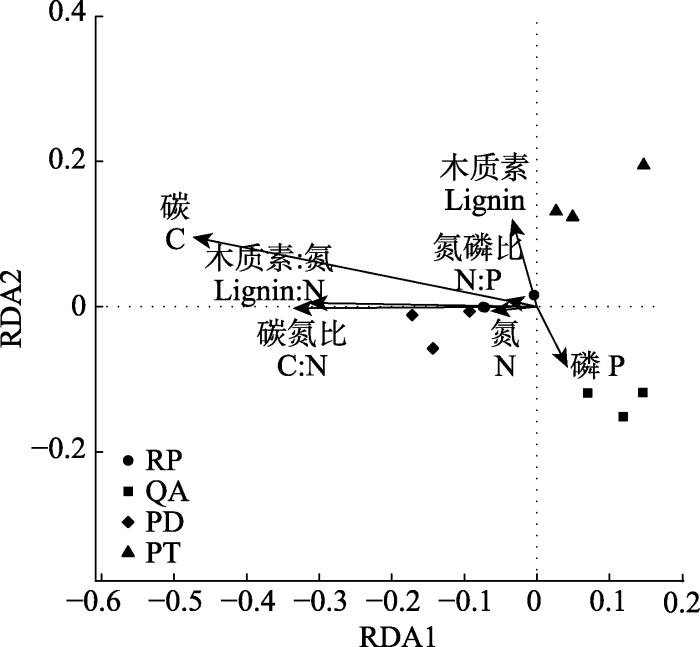 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT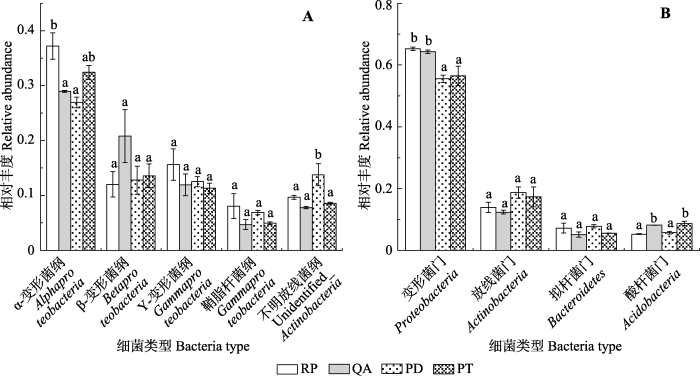 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT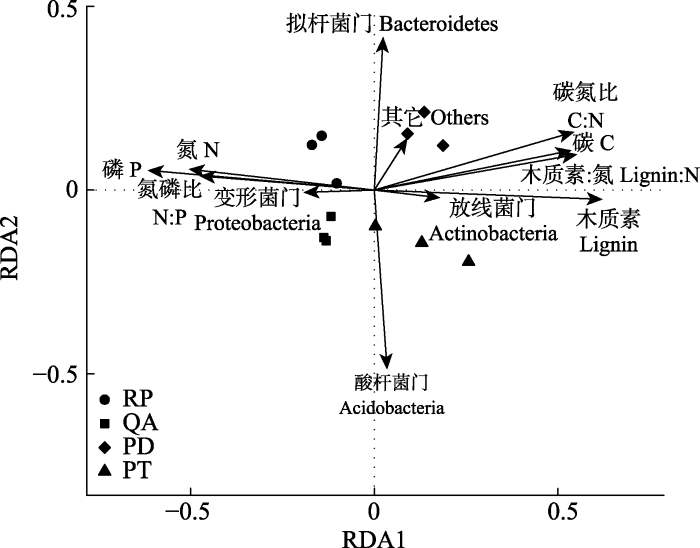 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}