Conduits anatomical structure and leaf traits of diffuse- and ring-porous stems in subtropical evergreen broad-leaved forests
Zhen-Zhen ZHANG,1,*, Ping ZHAO2, Jin-Xiu ZHANG1, Yao SI11 School of Geography and Environmental Sciences, Zhejiang Normal University, Jinhua, Zhejiang 231004, China 2 South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China
Supported by the National Natural Science Foundation of China(41630752) Supported by the National Natural Science Foundation of China(41701226) the Zhejiang Province Public Welfare Technology Application Research Project(GF19C030003)
Abstract Aims The conduits characters are critical for plants to develop their survival strategies. Our current knowledge in this regard remains limited for the subtropical forest. In this study, we set a study objective to quantify the relationship between the conduits characters and the leaf functional traits of the dominant species in the region. Methods Two dominant species, Castanopsis chinensis and Schima superba, in a subtropical forest in Shimentai Nature Reserve were selected to compare their differences in functional traits, including conduits anatomical structure, the leaf morphological characteristics, and leaf physiological characteristics. The study was conducted during the dry season (October to March of the following year) for quantifying the ring-porous and diffuse-porous species. A series of t-tests were performed to quantify the statistical differences of all traits between the two species. Important findings We found that the density of conduits of S. superba (diffuse-porous) was significantly higher than that of C. chinensis (ring-porous), while the diameter of conduits for C. chinensis was much larger than that of S. superba. The leaf water content and the Chlorophyll a/Chlorophyll b ratio were much higher for S. superba than that of C. chinensis; the stomatal density and specific leaf area (SLA) tended to be higher in C. chinensis. In addition, it appeared that the differences in leaf specific net photosynthetic rates and the leaf stomatal conductance were not significant between S. superba and C. chinensis. These results indicated that the ring-porous species C. chinensis maintain a high photosynthetic capacity by maintaining a higher SLA at the expense of low leaf water content in responding to the water stress. The diffuse-porous species S. superba, meanwhile, tended to maintain a high capability of light transform under drought. These functional differences might be responsible for the succession pathways under the gradual changes of global precipitation for the region. Keywords:ring-porous;diffuse-porous;leaf trait;survival strategy;subtropical forest
PDF (4498KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 张振振, 赵平, 张锦秀, 斯瑶. 亚热带常绿阔叶林散孔材和环孔材树种导管及叶片功能性状的比较. 植物生态学报, 2019, 43(2): 131-138. DOI: 10.17521/cjpe.2019.0291 ZHANG Zhen-Zhen, ZHAO Ping, ZHANG Jin-Xiu, SI Yao. Conduits anatomical structure and leaf traits of diffuse- and ring-porous stems in subtropical evergreen broad-leaved forests. Chinese Journal of Plant Ecology, 2019, 43(2): 131-138. DOI: 10.17521/cjpe.2019.0291
Table 1 表1 表1广东省石门台两种亚热带森林优势树种的详细信息 Table 1Community characteristics of the two dominant tree species of the subtropical forest in Shimentai of Guangdong Province
新窗口打开|下载原图ZIP|生成PPT 图3木荷和桂林栲的比叶面积(SLA), 叶片含水量(LWC), 气孔密度(Sd)和气孔大小(Ss)(平均值±标准误差)。*, p < 0.05; **, p < 0.01.
Fig. 3Specific leaf area (SLA), leaf water content (LWC), stomatal quantity (Sd) and size (Ss) of Schima superba and Castanopsis chinensis (mean ± SE). *, p < 0.05; **, p < 0.01.
ArnonDI ( 1949). Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiology, 24, 1-15. DOI:10.1104/pp.24.1.1URL [本文引用: 1]
BlonderB, VasseurF, ViolleC, ShipleyB, EnquistBJ, VileD ( 2015). Testing models for the leaf economics spectrum with leaf and whole-plant traits in Arabidopsis thaliana AoB Plants, 7, plv049. DOI: 10.1093/aobpla/plv049. [本文引用: 1]
BreshearsDD, MyersOB, MeyerCW, BarnesFJ, ZouCB, AllenCD, McDowellNG, PockmanWT ( 2009). Tree die-off in response to global change-type drought: Mortality insights from a decade of plant water potential measurements Frontiers in Ecology and the Environment, 7, 185-189. DOI:10.1890/080016URL [本文引用: 1]
BrodribbTJ, HolbrookNM, EdwardsEJ, GutiérrezMV ( 2010). Relations between stomatal closure, leaf turgor and xylem vulnerability in eight tropical dry forest trees Plant, Cell & Environment, 26, 443-450. [本文引用: 1]
BüssisD, von GrollU, FisahnJ, AltmannT ( 2006). Stomatal aperture can compensate altered stomatal density in Arabidopsis thaliana at growth light conditions. Functional Plant Biology, 33, 1037-1043. [本文引用: 1]
DíazS, CabidoM ( 2001). Vive la différence: Plant functional diversity matters to ecosystem processes Trends in Ecology & Evolution, 16, 646-655. [本文引用: 1]
DíazS, CabidoM, CasanovesF ( 1998). Plant functional traits and environmental filters at a regional scale Journal of Vegetation Science, 9, 113-122. DOI:10.2307/3237229URL [本文引用: 1]
DrakePL, FroendRH, FranksPJ ( 2013). Smaller, faster stomata: Scaling of stomatal size, rate of response, and stomatal conductance Journal of Experimental Botany, 64, 495-505. DOI:10.1093/jxb/ers347URL [本文引用: 1]
EstebanR, García-PlazaolaJI, HernándezA, Fernández-MarínB ( 2018). On the recalcitrant use of Arnon’s method for chlorophyll determination New Phytologist, 217, 474-476. DOI:10.1111/nph.14932URL [本文引用: 1]
FichotR, ChamaillardS, DepardieuC, Le ThiecD, CochardH, BarigahTS, BrignolasF ( 2010). Hydraulic efficiency and coordination with xylem resistance to cavitation, leaf function, and growth performance among eight unrelated Populus deltoides× Populus nigra hybrids. Journal of Experimental Botany, 62, 2093-2106. [本文引用: 1]
FranksPJ, DrakePL, BeerlingDJ ( 2009). Plasticity in maximum stomatal conductance constrained by negative correlation between stomatal size and density: An analysis using Eucalyptus globulus. Plant, Cell & Environment, 32, 1737-1748. [本文引用: 1]
KeyvanS ( 2010). The effects of drought stress on yield, relative water content, proline, soluble carbohydrates and chlorophyll of bread wheat cultivars Journal of Animal and Plant Sciences, 8, 1051-1060. [本文引用: 1]
KleinT ( 2014). The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours Functional Ecology, 28, 1313-1320. DOI:10.1111/1365-2435.12289URL [本文引用: 2]
LiK ( 2011). Community Structure and Leaf Characteristics of a Secondary Broad Leaved Forest in Hilly Area of Central Hunan, China Master degree dissertation, Central South University of Forestry and Technology, Changsha.
LiMC, ZhuJJ, SunYR ( 2009). Responses of specific leaf area of dominant tree species in Northeast China secondary forests to light intensity Chinese Journal of Ecology , 28, 1437-1442. [本文引用: 1]
LiWJ, ZuoJQ, SongYL, LiuJP, LiY, ShenYS, LiJX ( 2015). Changes in spatio temporal distribution of drought/flood disaster in southern China under global climate warming Meteorological Monthly , 41, 261-271. [本文引用: 1]
MathenyAM, BohrerG, VogelCS, MorinTH, HeL, FrassonRPDM, MirfenderesgiG, Sch?ferKVR, GoughCM, IvanovVY ( 2014). Species-specific transpiration responses to intermediate disturbance in a northern hardwood forest Journal of Geophysical Research Biogeosciences, 119, 2292-2311. DOI:10.1002/jgrg.v119.12URL [本文引用: 1]
MilcuA, AllanE, RoscherC, JenkinsT, MeyerST, FlynnD, BesslerH, BuscotF, EngelsC, GubschM ( 2013). Functionally and phylogenetically diverse plant communities key to soil biota Ecology, 94, 1878-1885. DOI:10.1890/12-1936.1URL [本文引用: 2]
NiuCY, MeinzerFC, HaoGY ( 2017). Divergence in strategies for coping with winter embolism among co-occurring temperate tree species: The role of positive xylem pressure, wood type and tree stature Functional Ecology, 31, 1550-1560. DOI:10.1111/fec.2017.31.issue-8URL [本文引用: 1]
PérezharguindeguyN, DíazS, GarnierE, LavorelS, PoorterH, JaureguiberryP, BretharteMS, CornwellWK, CraineJM, GurvichDE ( 2013). New handbook for standardised measurement of plant functional traits worldwide Australian Journal of Botany, 61, 167-234. DOI:10.1071/BT12225URL [本文引用: 2]
PetersEB, McfaddenJP, MontgomeryRA ( 2015). Biological and environmental controls on tree transpiration in a suburban landscape Journal of Geophysical Research Biogeosciences, 115, G04006. DOI: 10.1029/2009JG001266. [本文引用: 1]
PoorterL, McdonaldI, AlarcónA, FichtlerE, LiconaJC, Pe?a-ClarosM, SterckF, VillegasZ, Sass-KlaassenU ( 2010). The importance of wood traits and hydraulic conductance for the performance and life history strategies of 42 rainforest tree species New Phytologist, 185, 481-492. DOI:10.1111/j.1469-8137.2009.03092.xURL [本文引用: 1]
SperryJS, NicholsKL, SullivanJEM, EastlackSE ( 1994). Xylem embolism in ring-porous, diffuse-porous, and coniferous trees of northern Utah and interior Alaska Ecology, 75, 1736-1752. DOI:10.2307/1939633URL [本文引用: 1]
SperryJS, PockmanWT ( 1993). Limitation of transpiration by hydraulic conductance and xylem cavitation in Betula occidentalis. Plant, Cell & Environment, 16, 279-287. [本文引用: 1]
SteppeK, LemeurR ( 2007). Effects of ring-porous and diffuse-?porous stem wood anatomy on the hydraulic parameters used in a water flow and storage model Tree Physiology, 27, 43-52. DOI:10.1093/treephys/27.1.43URL [本文引用: 1]
TakahashiS, OkadaN, NobuchiT ( 2013). Relationship between the timing of vessel formation and leaf phenology in ten ring-porous and diffuse-porous deciduous tree species Ecological Research, 28, 615-624. DOI:10.1007/s11284-013-1053-xURL [本文引用: 1]
TateishiM, KumagaiTO, UtsumiY, UmebayashiT, ShiibaY, InoueK, KajiK, ChoK, OtsukiK ( 2008). Spatial variations in xylem sap flux density in evergreen oak trees with radial-porous wood: Comparisons with anatomical observations Trees, 22, 23-30. DOI:10.1007/s00468-007-0165-8URL [本文引用: 1]
von AllmenEI, SperryJS, BushSE ( 2013). Contrasting whole- tree water use, hydraulics, and growth in a co-dominant diffuse-porous vs. ring-porous species pair Trees, 29, 717-728. [本文引用: 1]
WangJ, IvesNE, LechowiczMJ ( 1992). The relation of foliar phenology to xylem embolism in trees Functional Ecology, 6, 469-475. DOI:10.2307/2389285URL [本文引用: 1]
WardleDA, BarkerGM, BonnerKI, NicholsonKS ( 1998). Can comparative approaches based on plant ecophysiological traits predict the nature of biotic interactions and individual plant species effects in ecosystems? Journal of Ecology, [本文引用: 1]
WestobyM, FalsterDS, MolesAT, AndPAV, WrightIJ ( 2002). Plant ecological strategies: Some leading dimensions of variation between species Annual Review of Ecology & Systematics, 33, 125-159. [本文引用: 1]
WrightIJ, AckerlyDD, BongersF, HarmsKE, Ibarra-?ManriquezG, Martinez-RamosM, MazerSJ, Muller-?LandauHC, PazH, PitmanNCA, PoorterL, SilmanMR, VriesendorpCF, WebbCO, WestobyM, WrightSJ ( 2007). Relationships among ecologically important dimensions of plant trait variation in seven neotropical forests Annals of Botany, 99, 1003-1015. DOI:10.1093/aob/mcl066URL [本文引用: 1]
YanDL, RenYY, LianJF, ShengLJ ( 2012). Leaf functional traits and their associated characteristics of three dominant families Journal of Forestry Engineering , 26(3), 34-37. [本文引用: 1]
ZengXM, ZhaoP, OuyangL, ZhuLW, NiGY, ZhaoXH ( 2017). Soil water use and adaptive regulation of Schima superba in the dry and wet seasons. Journal of Tropical and Subtropical Botany, 25(2), 105-114. [本文引用: 1]
ZengXP, ZhaoP, CaiXA, RaoXQ, LiuH, MaL, LiCH ( 2006). A preliminary study on the tolerance of 25 species of South Asian tropical plants Journal of Beijing Forestry University , 28(4), 92-99. [本文引用: 2]
ZhangJL, PoorterL, CaoKF ( 2012). Productive leaf functional traits of Chinese savanna species Plant Ecology, 213, 1449-1460. DOI:10.1007/s11258-012-0103-8URL [本文引用: 1]
ZhouG, WeiX, WuY, LiuS, HuangY, YanJ, ZhangD, ZhangQ, LiuJ, MengZ, WangC, ChuG, LiuS, TangX, WangC ( 2011). Quantifying the hydrological responses to climate change in an intact forested small watershed in Southern China Global Change Biology, 17, 3736-3746. DOI:10.1111/j.1365-2486.2011.02499.xURL [本文引用: 1]
ZhuL, HuY, ZhaoX, ZengX, ZhaoP, ZhangZ, JuY ( 2017). The impact of drought on sap flow of cooccurring Liquidambar formosana Hance and Quercus variabilis Blume in a temperate forest, Central China. Ecohydrology, 10, 1828. DOI: 10.1002/eco.1828. [本文引用: 1]
Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. 1 1949
Testing models for the leaf economics spectrum with leaf and whole-plant traits in Arabidopsis thaliana 1 2015
... 在全球范围内, 生态****从个体到整个生态系统对多个层次的植物功能性状进行了大量研究(Wright et al., 2007; Chave et al., 2009; Poorter et al., 2010; Pérezharguindeguy et al., 2013).其中, 叶片功能性状直接反映了植物为适应环境变化而采取的生存策略(Wardle et al., 1998).植物不同叶片性状间关系的研究一直是植物生态学的重要领域(Zhang et al., 2012; Blonder et al., 2015).然而叶片的功能性状并不能单独决定植物的生存策略, 当其与木质部结构特征无法耦合时, 就会导致植物的死亡(Breshears et al., 2009).例如, 叶片气孔开度、叶片水势与植物木质部结构紧密相关(Brodribb et al., 2010), 当植物气孔特性不能够随着木质部导水率的丧失而产生相应调整时, 就会导致叶片凋落甚至植物死亡(Sperry & Pockman, 1993).因此, 深入了解植物叶片和木质部结构之间的耦合关系是分析气候变化背景下森林群落结构变化的前提. ...
Tree die-off in response to global change-type drought: Mortality insights from a decade of plant water potential measurements 1 2009
... 在全球范围内, 生态****从个体到整个生态系统对多个层次的植物功能性状进行了大量研究(Wright et al., 2007; Chave et al., 2009; Poorter et al., 2010; Pérezharguindeguy et al., 2013).其中, 叶片功能性状直接反映了植物为适应环境变化而采取的生存策略(Wardle et al., 1998).植物不同叶片性状间关系的研究一直是植物生态学的重要领域(Zhang et al., 2012; Blonder et al., 2015).然而叶片的功能性状并不能单独决定植物的生存策略, 当其与木质部结构特征无法耦合时, 就会导致植物的死亡(Breshears et al., 2009).例如, 叶片气孔开度、叶片水势与植物木质部结构紧密相关(Brodribb et al., 2010), 当植物气孔特性不能够随着木质部导水率的丧失而产生相应调整时, 就会导致叶片凋落甚至植物死亡(Sperry & Pockman, 1993).因此, 深入了解植物叶片和木质部结构之间的耦合关系是分析气候变化背景下森林群落结构变化的前提. ...
Relations between stomatal closure, leaf turgor and xylem vulnerability in eight tropical dry forest trees 1 2010
... 在全球范围内, 生态****从个体到整个生态系统对多个层次的植物功能性状进行了大量研究(Wright et al., 2007; Chave et al., 2009; Poorter et al., 2010; Pérezharguindeguy et al., 2013).其中, 叶片功能性状直接反映了植物为适应环境变化而采取的生存策略(Wardle et al., 1998).植物不同叶片性状间关系的研究一直是植物生态学的重要领域(Zhang et al., 2012; Blonder et al., 2015).然而叶片的功能性状并不能单独决定植物的生存策略, 当其与木质部结构特征无法耦合时, 就会导致植物的死亡(Breshears et al., 2009).例如, 叶片气孔开度、叶片水势与植物木质部结构紧密相关(Brodribb et al., 2010), 当植物气孔特性不能够随着木质部导水率的丧失而产生相应调整时, 就会导致叶片凋落甚至植物死亡(Sperry & Pockman, 1993).因此, 深入了解植物叶片和木质部结构之间的耦合关系是分析气候变化背景下森林群落结构变化的前提. ...
Stomatal aperture can compensate altered stomatal density in Arabidopsis thaliana at growth light conditions. 1 2006
... 除此之外, 对水分条件变化的应对能力同样在植物生存策略中占有十分重要的地位(Milcu et al., 2013).两种植物导管密度和直径的差异决定了两者的输水策略, 即木荷具有较大的Kn, 具有较强的整树输水潜力, 从而影响叶片的水分利用策略.从叶片结构上, 我们发现, 作为环孔材的桂林栲, 其Sd显著低于散孔材的木荷(图3; p = 0.022).研究表明, 较低的叶片Sd能够使植物具有较高的Cond (Franks et al., 2009), 但当光照较强时, 其光合作用将会受到极大的影响(Büssis et al., 2006).而Sd较大的植物, 由于有较好的气孔调控能力, 气孔不仅能够在环境恶劣的条件下迅速关闭, 同时也可以在环境较好的条件下迅速打开(Drake et al., 2013).因此, 桂林栲在水分限制条件下, 也更有可能出现水分胁迫.与此同时, 我们发现木荷的LWC也显著高于桂林栲(图2), 因此对干旱的耐受性更强(Keyvan, 2010).这与上述较强的气孔调控能力相一致.植物叶片这种对干旱的应对能力, 与其导管特征是密不可分的(Klein, 2014).研究发现, 散孔材树种Acer grandidentatum的蒸腾量是环孔材树种Quercus gambelii的两倍(von Allmen et al., 2013).而在我们的另一项研究中也发现, 样地中木荷的树干液流速率也显著高于桂林栲(待发表).这就使得木荷在干季能够维持较高的Tr以及较低WUEi (图4).此外, Klein (2014)通过大量的研究发现, 当植物Cond下降50%时, 环孔材植物的叶片水势下降量是散孔材的2倍.这些结果都为木荷较高的叶片耐旱特性提供了支撑.因此, 从这些结果中我们可以发现, 散孔材较大的导管密度能够增强其叶片的耐旱能力. ...
Towards a worldwide wood economics spectrum 1 2009
... 在全球范围内, 生态****从个体到整个生态系统对多个层次的植物功能性状进行了大量研究(Wright et al., 2007; Chave et al., 2009; Poorter et al., 2010; Pérezharguindeguy et al., 2013).其中, 叶片功能性状直接反映了植物为适应环境变化而采取的生存策略(Wardle et al., 1998).植物不同叶片性状间关系的研究一直是植物生态学的重要领域(Zhang et al., 2012; Blonder et al., 2015).然而叶片的功能性状并不能单独决定植物的生存策略, 当其与木质部结构特征无法耦合时, 就会导致植物的死亡(Breshears et al., 2009).例如, 叶片气孔开度、叶片水势与植物木质部结构紧密相关(Brodribb et al., 2010), 当植物气孔特性不能够随着木质部导水率的丧失而产生相应调整时, 就会导致叶片凋落甚至植物死亡(Sperry & Pockman, 1993).因此, 深入了解植物叶片和木质部结构之间的耦合关系是分析气候变化背景下森林群落结构变化的前提. ...
Vive la différence: Plant functional diversity matters to ecosystem processes 1 2001
... 环境胁迫诱导的基于植物功能性状的生存策略变化一直是生态学研究中的关键问题(Díaz et al., 1998; Díaz & Cabido, 2001).通过揭示不同物种采用的生存策略, 可为了解气候变化下的森林的结构和演替规律提供基础(Hulot et al., 2000; Westoby et al., 2002; Milcu et al., 2013). ...
Plant functional traits and environmental filters at a regional scale 1 1998
... 环境胁迫诱导的基于植物功能性状的生存策略变化一直是生态学研究中的关键问题(Díaz et al., 1998; Díaz & Cabido, 2001).通过揭示不同物种采用的生存策略, 可为了解气候变化下的森林的结构和演替规律提供基础(Hulot et al., 2000; Westoby et al., 2002; Milcu et al., 2013). ...
Smaller, faster stomata: Scaling of stomatal size, rate of response, and stomatal conductance 1 2013
... 除此之外, 对水分条件变化的应对能力同样在植物生存策略中占有十分重要的地位(Milcu et al., 2013).两种植物导管密度和直径的差异决定了两者的输水策略, 即木荷具有较大的Kn, 具有较强的整树输水潜力, 从而影响叶片的水分利用策略.从叶片结构上, 我们发现, 作为环孔材的桂林栲, 其Sd显著低于散孔材的木荷(图3; p = 0.022).研究表明, 较低的叶片Sd能够使植物具有较高的Cond (Franks et al., 2009), 但当光照较强时, 其光合作用将会受到极大的影响(Büssis et al., 2006).而Sd较大的植物, 由于有较好的气孔调控能力, 气孔不仅能够在环境恶劣的条件下迅速关闭, 同时也可以在环境较好的条件下迅速打开(Drake et al., 2013).因此, 桂林栲在水分限制条件下, 也更有可能出现水分胁迫.与此同时, 我们发现木荷的LWC也显著高于桂林栲(图2), 因此对干旱的耐受性更强(Keyvan, 2010).这与上述较强的气孔调控能力相一致.植物叶片这种对干旱的应对能力, 与其导管特征是密不可分的(Klein, 2014).研究发现, 散孔材树种Acer grandidentatum的蒸腾量是环孔材树种Quercus gambelii的两倍(von Allmen et al., 2013).而在我们的另一项研究中也发现, 样地中木荷的树干液流速率也显著高于桂林栲(待发表).这就使得木荷在干季能够维持较高的Tr以及较低WUEi (图4).此外, Klein (2014)通过大量的研究发现, 当植物Cond下降50%时, 环孔材植物的叶片水势下降量是散孔材的2倍.这些结果都为木荷较高的叶片耐旱特性提供了支撑.因此, 从这些结果中我们可以发现, 散孔材较大的导管密度能够增强其叶片的耐旱能力. ...
On the recalcitrant use of Arnon’s method for chlorophyll determination 1 2018
Hydraulic efficiency and coordination with xylem resistance to cavitation, leaf function, and growth performance among eight unrelated Populus deltoides× Populus nigra hybrids. 1 2010
... 基于木质部导管解剖特征, 被子植物大致被分为两类, 即环孔材物种和散孔材物种.这两种植物的导管结构特征差异导致水分生理调节的显著差异(Matheny et al., 2014; Peters et al., 2015).研究发现环孔材植物具有高的水分传输能力(Niu et al., 2017), 而在水分胁迫条件下更容易遭受空穴化和栓塞胁迫, 从而降低植物对树冠的供水能力(Wang et al., 1992; Sperry et al., 1994; Steppe & Lemeur, 2007).导管直径较小的散孔材植物具有较低的水传输速率, 其水分传输速率的最大值、最小值和季节变化都很小(Sack & Holbrook, 2006).干湿季节环孔材和散孔材植物的水分蒸腾差异表明他们对水的调控机制不同(Zhu et al., 2017).近年来, 很多****对具有不同木质部特征的树木的水分运输效率和水分利用策略进行了广泛的研究, 希望从植物水力结构的角度探讨植物的水分运输, 评价树木的耐旱性(Fichot et al., 2010).有研究曾阐明早春时节环孔材和散孔材物种导管形成的时间与叶物候之间的关系(Takahashi et al., 2013), 但尚不清楚植物这种物候特征的耦合能否反映在功能性状特征上.我国亚热带地区受季风气候的影响, 干湿季分明(曾小敏等, 2017), 尤其是全球气候变化导致的降水格局变化(李维京等, 2015), 更加剧了这种干湿季节降水的差异性.如何适应这种短时间的极端环境变化是亚热带地区植被生存演替所面临的严峻挑战. ...
Plasticity in maximum stomatal conductance constrained by negative correlation between stomatal size and density: An analysis using Eucalyptus globulus. 1 2009
... 除此之外, 对水分条件变化的应对能力同样在植物生存策略中占有十分重要的地位(Milcu et al., 2013).两种植物导管密度和直径的差异决定了两者的输水策略, 即木荷具有较大的Kn, 具有较强的整树输水潜力, 从而影响叶片的水分利用策略.从叶片结构上, 我们发现, 作为环孔材的桂林栲, 其Sd显著低于散孔材的木荷(图3; p = 0.022).研究表明, 较低的叶片Sd能够使植物具有较高的Cond (Franks et al., 2009), 但当光照较强时, 其光合作用将会受到极大的影响(Büssis et al., 2006).而Sd较大的植物, 由于有较好的气孔调控能力, 气孔不仅能够在环境恶劣的条件下迅速关闭, 同时也可以在环境较好的条件下迅速打开(Drake et al., 2013).因此, 桂林栲在水分限制条件下, 也更有可能出现水分胁迫.与此同时, 我们发现木荷的LWC也显著高于桂林栲(图2), 因此对干旱的耐受性更强(Keyvan, 2010).这与上述较强的气孔调控能力相一致.植物叶片这种对干旱的应对能力, 与其导管特征是密不可分的(Klein, 2014).研究发现, 散孔材树种Acer grandidentatum的蒸腾量是环孔材树种Quercus gambelii的两倍(von Allmen et al., 2013).而在我们的另一项研究中也发现, 样地中木荷的树干液流速率也显著高于桂林栲(待发表).这就使得木荷在干季能够维持较高的Tr以及较低WUEi (图4).此外, Klein (2014)通过大量的研究发现, 当植物Cond下降50%时, 环孔材植物的叶片水势下降量是散孔材的2倍.这些结果都为木荷较高的叶片耐旱特性提供了支撑.因此, 从这些结果中我们可以发现, 散孔材较大的导管密度能够增强其叶片的耐旱能力. ...
Functional diversity governs ecosystem response to nutrient enrichment 1 2000
... 环境胁迫诱导的基于植物功能性状的生存策略变化一直是生态学研究中的关键问题(Díaz et al., 1998; Díaz & Cabido, 2001).通过揭示不同物种采用的生存策略, 可为了解气候变化下的森林的结构和演替规律提供基础(Hulot et al., 2000; Westoby et al., 2002; Milcu et al., 2013). ...
The effects of drought stress on yield, relative water content, proline, soluble carbohydrates and chlorophyll of bread wheat cultivars 1 2010
... 除此之外, 对水分条件变化的应对能力同样在植物生存策略中占有十分重要的地位(Milcu et al., 2013).两种植物导管密度和直径的差异决定了两者的输水策略, 即木荷具有较大的Kn, 具有较强的整树输水潜力, 从而影响叶片的水分利用策略.从叶片结构上, 我们发现, 作为环孔材的桂林栲, 其Sd显著低于散孔材的木荷(图3; p = 0.022).研究表明, 较低的叶片Sd能够使植物具有较高的Cond (Franks et al., 2009), 但当光照较强时, 其光合作用将会受到极大的影响(Büssis et al., 2006).而Sd较大的植物, 由于有较好的气孔调控能力, 气孔不仅能够在环境恶劣的条件下迅速关闭, 同时也可以在环境较好的条件下迅速打开(Drake et al., 2013).因此, 桂林栲在水分限制条件下, 也更有可能出现水分胁迫.与此同时, 我们发现木荷的LWC也显著高于桂林栲(图2), 因此对干旱的耐受性更强(Keyvan, 2010).这与上述较强的气孔调控能力相一致.植物叶片这种对干旱的应对能力, 与其导管特征是密不可分的(Klein, 2014).研究发现, 散孔材树种Acer grandidentatum的蒸腾量是环孔材树种Quercus gambelii的两倍(von Allmen et al., 2013).而在我们的另一项研究中也发现, 样地中木荷的树干液流速率也显著高于桂林栲(待发表).这就使得木荷在干季能够维持较高的Tr以及较低WUEi (图4).此外, Klein (2014)通过大量的研究发现, 当植物Cond下降50%时, 环孔材植物的叶片水势下降量是散孔材的2倍.这些结果都为木荷较高的叶片耐旱特性提供了支撑.因此, 从这些结果中我们可以发现, 散孔材较大的导管密度能够增强其叶片的耐旱能力. ...
The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours 2 2014
... 除此之外, 对水分条件变化的应对能力同样在植物生存策略中占有十分重要的地位(Milcu et al., 2013).两种植物导管密度和直径的差异决定了两者的输水策略, 即木荷具有较大的Kn, 具有较强的整树输水潜力, 从而影响叶片的水分利用策略.从叶片结构上, 我们发现, 作为环孔材的桂林栲, 其Sd显著低于散孔材的木荷(图3; p = 0.022).研究表明, 较低的叶片Sd能够使植物具有较高的Cond (Franks et al., 2009), 但当光照较强时, 其光合作用将会受到极大的影响(Büssis et al., 2006).而Sd较大的植物, 由于有较好的气孔调控能力, 气孔不仅能够在环境恶劣的条件下迅速关闭, 同时也可以在环境较好的条件下迅速打开(Drake et al., 2013).因此, 桂林栲在水分限制条件下, 也更有可能出现水分胁迫.与此同时, 我们发现木荷的LWC也显著高于桂林栲(图2), 因此对干旱的耐受性更强(Keyvan, 2010).这与上述较强的气孔调控能力相一致.植物叶片这种对干旱的应对能力, 与其导管特征是密不可分的(Klein, 2014).研究发现, 散孔材树种Acer grandidentatum的蒸腾量是环孔材树种Quercus gambelii的两倍(von Allmen et al., 2013).而在我们的另一项研究中也发现, 样地中木荷的树干液流速率也显著高于桂林栲(待发表).这就使得木荷在干季能够维持较高的Tr以及较低WUEi (图4).此外, Klein (2014)通过大量的研究发现, 当植物Cond下降50%时, 环孔材植物的叶片水势下降量是散孔材的2倍.这些结果都为木荷较高的叶片耐旱特性提供了支撑.因此, 从这些结果中我们可以发现, 散孔材较大的导管密度能够增强其叶片的耐旱能力. ...
... ).此外, Klein (2014)通过大量的研究发现, 当植物Cond下降50%时, 环孔材植物的叶片水势下降量是散孔材的2倍.这些结果都为木荷较高的叶片耐旱特性提供了支撑.因此, 从这些结果中我们可以发现, 散孔材较大的导管密度能够增强其叶片的耐旱能力. ...
... 基于木质部导管解剖特征, 被子植物大致被分为两类, 即环孔材物种和散孔材物种.这两种植物的导管结构特征差异导致水分生理调节的显著差异(Matheny et al., 2014; Peters et al., 2015).研究发现环孔材植物具有高的水分传输能力(Niu et al., 2017), 而在水分胁迫条件下更容易遭受空穴化和栓塞胁迫, 从而降低植物对树冠的供水能力(Wang et al., 1992; Sperry et al., 1994; Steppe & Lemeur, 2007).导管直径较小的散孔材植物具有较低的水传输速率, 其水分传输速率的最大值、最小值和季节变化都很小(Sack & Holbrook, 2006).干湿季节环孔材和散孔材植物的水分蒸腾差异表明他们对水的调控机制不同(Zhu et al., 2017).近年来, 很多****对具有不同木质部特征的树木的水分运输效率和水分利用策略进行了广泛的研究, 希望从植物水力结构的角度探讨植物的水分运输, 评价树木的耐旱性(Fichot et al., 2010).有研究曾阐明早春时节环孔材和散孔材物种导管形成的时间与叶物候之间的关系(Takahashi et al., 2013), 但尚不清楚植物这种物候特征的耦合能否反映在功能性状特征上.我国亚热带地区受季风气候的影响, 干湿季分明(曾小敏等, 2017), 尤其是全球气候变化导致的降水格局变化(李维京等, 2015), 更加剧了这种干湿季节降水的差异性.如何适应这种短时间的极端环境变化是亚热带地区植被生存演替所面临的严峻挑战. ...
气候变暖背景下我国南方旱涝灾害时空格局变化 1 2015
... 基于木质部导管解剖特征, 被子植物大致被分为两类, 即环孔材物种和散孔材物种.这两种植物的导管结构特征差异导致水分生理调节的显著差异(Matheny et al., 2014; Peters et al., 2015).研究发现环孔材植物具有高的水分传输能力(Niu et al., 2017), 而在水分胁迫条件下更容易遭受空穴化和栓塞胁迫, 从而降低植物对树冠的供水能力(Wang et al., 1992; Sperry et al., 1994; Steppe & Lemeur, 2007).导管直径较小的散孔材植物具有较低的水传输速率, 其水分传输速率的最大值、最小值和季节变化都很小(Sack & Holbrook, 2006).干湿季节环孔材和散孔材植物的水分蒸腾差异表明他们对水的调控机制不同(Zhu et al., 2017).近年来, 很多****对具有不同木质部特征的树木的水分运输效率和水分利用策略进行了广泛的研究, 希望从植物水力结构的角度探讨植物的水分运输, 评价树木的耐旱性(Fichot et al., 2010).有研究曾阐明早春时节环孔材和散孔材物种导管形成的时间与叶物候之间的关系(Takahashi et al., 2013), 但尚不清楚植物这种物候特征的耦合能否反映在功能性状特征上.我国亚热带地区受季风气候的影响, 干湿季分明(曾小敏等, 2017), 尤其是全球气候变化导致的降水格局变化(李维京等, 2015), 更加剧了这种干湿季节降水的差异性.如何适应这种短时间的极端环境变化是亚热带地区植被生存演替所面临的严峻挑战. ...
Species-specific transpiration responses to intermediate disturbance in a northern hardwood forest 1 2014
... 基于木质部导管解剖特征, 被子植物大致被分为两类, 即环孔材物种和散孔材物种.这两种植物的导管结构特征差异导致水分生理调节的显著差异(Matheny et al., 2014; Peters et al., 2015).研究发现环孔材植物具有高的水分传输能力(Niu et al., 2017), 而在水分胁迫条件下更容易遭受空穴化和栓塞胁迫, 从而降低植物对树冠的供水能力(Wang et al., 1992; Sperry et al., 1994; Steppe & Lemeur, 2007).导管直径较小的散孔材植物具有较低的水传输速率, 其水分传输速率的最大值、最小值和季节变化都很小(Sack & Holbrook, 2006).干湿季节环孔材和散孔材植物的水分蒸腾差异表明他们对水的调控机制不同(Zhu et al., 2017).近年来, 很多****对具有不同木质部特征的树木的水分运输效率和水分利用策略进行了广泛的研究, 希望从植物水力结构的角度探讨植物的水分运输, 评价树木的耐旱性(Fichot et al., 2010).有研究曾阐明早春时节环孔材和散孔材物种导管形成的时间与叶物候之间的关系(Takahashi et al., 2013), 但尚不清楚植物这种物候特征的耦合能否反映在功能性状特征上.我国亚热带地区受季风气候的影响, 干湿季分明(曾小敏等, 2017), 尤其是全球气候变化导致的降水格局变化(李维京等, 2015), 更加剧了这种干湿季节降水的差异性.如何适应这种短时间的极端环境变化是亚热带地区植被生存演替所面临的严峻挑战. ...
Functionally and phylogenetically diverse plant communities key to soil biota 2 2013
... 环境胁迫诱导的基于植物功能性状的生存策略变化一直是生态学研究中的关键问题(Díaz et al., 1998; Díaz & Cabido, 2001).通过揭示不同物种采用的生存策略, 可为了解气候变化下的森林的结构和演替规律提供基础(Hulot et al., 2000; Westoby et al., 2002; Milcu et al., 2013). ...
... 除此之外, 对水分条件变化的应对能力同样在植物生存策略中占有十分重要的地位(Milcu et al., 2013).两种植物导管密度和直径的差异决定了两者的输水策略, 即木荷具有较大的Kn, 具有较强的整树输水潜力, 从而影响叶片的水分利用策略.从叶片结构上, 我们发现, 作为环孔材的桂林栲, 其Sd显著低于散孔材的木荷(图3; p = 0.022).研究表明, 较低的叶片Sd能够使植物具有较高的Cond (Franks et al., 2009), 但当光照较强时, 其光合作用将会受到极大的影响(Büssis et al., 2006).而Sd较大的植物, 由于有较好的气孔调控能力, 气孔不仅能够在环境恶劣的条件下迅速关闭, 同时也可以在环境较好的条件下迅速打开(Drake et al., 2013).因此, 桂林栲在水分限制条件下, 也更有可能出现水分胁迫.与此同时, 我们发现木荷的LWC也显著高于桂林栲(图2), 因此对干旱的耐受性更强(Keyvan, 2010).这与上述较强的气孔调控能力相一致.植物叶片这种对干旱的应对能力, 与其导管特征是密不可分的(Klein, 2014).研究发现, 散孔材树种Acer grandidentatum的蒸腾量是环孔材树种Quercus gambelii的两倍(von Allmen et al., 2013).而在我们的另一项研究中也发现, 样地中木荷的树干液流速率也显著高于桂林栲(待发表).这就使得木荷在干季能够维持较高的Tr以及较低WUEi (图4).此外, Klein (2014)通过大量的研究发现, 当植物Cond下降50%时, 环孔材植物的叶片水势下降量是散孔材的2倍.这些结果都为木荷较高的叶片耐旱特性提供了支撑.因此, 从这些结果中我们可以发现, 散孔材较大的导管密度能够增强其叶片的耐旱能力. ...
Divergence in strategies for coping with winter embolism among co-occurring temperate tree species: The role of positive xylem pressure, wood type and tree stature 1 2017
... 基于木质部导管解剖特征, 被子植物大致被分为两类, 即环孔材物种和散孔材物种.这两种植物的导管结构特征差异导致水分生理调节的显著差异(Matheny et al., 2014; Peters et al., 2015).研究发现环孔材植物具有高的水分传输能力(Niu et al., 2017), 而在水分胁迫条件下更容易遭受空穴化和栓塞胁迫, 从而降低植物对树冠的供水能力(Wang et al., 1992; Sperry et al., 1994; Steppe & Lemeur, 2007).导管直径较小的散孔材植物具有较低的水传输速率, 其水分传输速率的最大值、最小值和季节变化都很小(Sack & Holbrook, 2006).干湿季节环孔材和散孔材植物的水分蒸腾差异表明他们对水的调控机制不同(Zhu et al., 2017).近年来, 很多****对具有不同木质部特征的树木的水分运输效率和水分利用策略进行了广泛的研究, 希望从植物水力结构的角度探讨植物的水分运输, 评价树木的耐旱性(Fichot et al., 2010).有研究曾阐明早春时节环孔材和散孔材物种导管形成的时间与叶物候之间的关系(Takahashi et al., 2013), 但尚不清楚植物这种物候特征的耦合能否反映在功能性状特征上.我国亚热带地区受季风气候的影响, 干湿季分明(曾小敏等, 2017), 尤其是全球气候变化导致的降水格局变化(李维京等, 2015), 更加剧了这种干湿季节降水的差异性.如何适应这种短时间的极端环境变化是亚热带地区植被生存演替所面临的严峻挑战. ...
New handbook for standardised measurement of plant functional traits worldwide 2 2013
... 在全球范围内, 生态****从个体到整个生态系统对多个层次的植物功能性状进行了大量研究(Wright et al., 2007; Chave et al., 2009; Poorter et al., 2010; Pérezharguindeguy et al., 2013).其中, 叶片功能性状直接反映了植物为适应环境变化而采取的生存策略(Wardle et al., 1998).植物不同叶片性状间关系的研究一直是植物生态学的重要领域(Zhang et al., 2012; Blonder et al., 2015).然而叶片的功能性状并不能单独决定植物的生存策略, 当其与木质部结构特征无法耦合时, 就会导致植物的死亡(Breshears et al., 2009).例如, 叶片气孔开度、叶片水势与植物木质部结构紧密相关(Brodribb et al., 2010), 当植物气孔特性不能够随着木质部导水率的丧失而产生相应调整时, 就会导致叶片凋落甚至植物死亡(Sperry & Pockman, 1993).因此, 深入了解植物叶片和木质部结构之间的耦合关系是分析气候变化背景下森林群落结构变化的前提. ...
Biological and environmental controls on tree transpiration in a suburban landscape 1 2015
... 基于木质部导管解剖特征, 被子植物大致被分为两类, 即环孔材物种和散孔材物种.这两种植物的导管结构特征差异导致水分生理调节的显著差异(Matheny et al., 2014; Peters et al., 2015).研究发现环孔材植物具有高的水分传输能力(Niu et al., 2017), 而在水分胁迫条件下更容易遭受空穴化和栓塞胁迫, 从而降低植物对树冠的供水能力(Wang et al., 1992; Sperry et al., 1994; Steppe & Lemeur, 2007).导管直径较小的散孔材植物具有较低的水传输速率, 其水分传输速率的最大值、最小值和季节变化都很小(Sack & Holbrook, 2006).干湿季节环孔材和散孔材植物的水分蒸腾差异表明他们对水的调控机制不同(Zhu et al., 2017).近年来, 很多****对具有不同木质部特征的树木的水分运输效率和水分利用策略进行了广泛的研究, 希望从植物水力结构的角度探讨植物的水分运输, 评价树木的耐旱性(Fichot et al., 2010).有研究曾阐明早春时节环孔材和散孔材物种导管形成的时间与叶物候之间的关系(Takahashi et al., 2013), 但尚不清楚植物这种物候特征的耦合能否反映在功能性状特征上.我国亚热带地区受季风气候的影响, 干湿季分明(曾小敏等, 2017), 尤其是全球气候变化导致的降水格局变化(李维京等, 2015), 更加剧了这种干湿季节降水的差异性.如何适应这种短时间的极端环境变化是亚热带地区植被生存演替所面临的严峻挑战. ...
The importance of wood traits and hydraulic conductance for the performance and life history strategies of 42 rainforest tree species 1 2010
... 在全球范围内, 生态****从个体到整个生态系统对多个层次的植物功能性状进行了大量研究(Wright et al., 2007; Chave et al., 2009; Poorter et al., 2010; Pérezharguindeguy et al., 2013).其中, 叶片功能性状直接反映了植物为适应环境变化而采取的生存策略(Wardle et al., 1998).植物不同叶片性状间关系的研究一直是植物生态学的重要领域(Zhang et al., 2012; Blonder et al., 2015).然而叶片的功能性状并不能单独决定植物的生存策略, 当其与木质部结构特征无法耦合时, 就会导致植物的死亡(Breshears et al., 2009).例如, 叶片气孔开度、叶片水势与植物木质部结构紧密相关(Brodribb et al., 2010), 当植物气孔特性不能够随着木质部导水率的丧失而产生相应调整时, 就会导致叶片凋落甚至植物死亡(Sperry & Pockman, 1993).因此, 深入了解植物叶片和木质部结构之间的耦合关系是分析气候变化背景下森林群落结构变化的前提. ...
Leaf hydraulics 1 2006
... 基于木质部导管解剖特征, 被子植物大致被分为两类, 即环孔材物种和散孔材物种.这两种植物的导管结构特征差异导致水分生理调节的显著差异(Matheny et al., 2014; Peters et al., 2015).研究发现环孔材植物具有高的水分传输能力(Niu et al., 2017), 而在水分胁迫条件下更容易遭受空穴化和栓塞胁迫, 从而降低植物对树冠的供水能力(Wang et al., 1992; Sperry et al., 1994; Steppe & Lemeur, 2007).导管直径较小的散孔材植物具有较低的水传输速率, 其水分传输速率的最大值、最小值和季节变化都很小(Sack & Holbrook, 2006).干湿季节环孔材和散孔材植物的水分蒸腾差异表明他们对水的调控机制不同(Zhu et al., 2017).近年来, 很多****对具有不同木质部特征的树木的水分运输效率和水分利用策略进行了广泛的研究, 希望从植物水力结构的角度探讨植物的水分运输, 评价树木的耐旱性(Fichot et al., 2010).有研究曾阐明早春时节环孔材和散孔材物种导管形成的时间与叶物候之间的关系(Takahashi et al., 2013), 但尚不清楚植物这种物候特征的耦合能否反映在功能性状特征上.我国亚热带地区受季风气候的影响, 干湿季分明(曾小敏等, 2017), 尤其是全球气候变化导致的降水格局变化(李维京等, 2015), 更加剧了这种干湿季节降水的差异性.如何适应这种短时间的极端环境变化是亚热带地区植被生存演替所面临的严峻挑战. ...
Xylem embolism in ring-porous, diffuse-porous, and coniferous trees of northern Utah and interior Alaska 1 1994
... 基于木质部导管解剖特征, 被子植物大致被分为两类, 即环孔材物种和散孔材物种.这两种植物的导管结构特征差异导致水分生理调节的显著差异(Matheny et al., 2014; Peters et al., 2015).研究发现环孔材植物具有高的水分传输能力(Niu et al., 2017), 而在水分胁迫条件下更容易遭受空穴化和栓塞胁迫, 从而降低植物对树冠的供水能力(Wang et al., 1992; Sperry et al., 1994; Steppe & Lemeur, 2007).导管直径较小的散孔材植物具有较低的水传输速率, 其水分传输速率的最大值、最小值和季节变化都很小(Sack & Holbrook, 2006).干湿季节环孔材和散孔材植物的水分蒸腾差异表明他们对水的调控机制不同(Zhu et al., 2017).近年来, 很多****对具有不同木质部特征的树木的水分运输效率和水分利用策略进行了广泛的研究, 希望从植物水力结构的角度探讨植物的水分运输, 评价树木的耐旱性(Fichot et al., 2010).有研究曾阐明早春时节环孔材和散孔材物种导管形成的时间与叶物候之间的关系(Takahashi et al., 2013), 但尚不清楚植物这种物候特征的耦合能否反映在功能性状特征上.我国亚热带地区受季风气候的影响, 干湿季分明(曾小敏等, 2017), 尤其是全球气候变化导致的降水格局变化(李维京等, 2015), 更加剧了这种干湿季节降水的差异性.如何适应这种短时间的极端环境变化是亚热带地区植被生存演替所面临的严峻挑战. ...
Limitation of transpiration by hydraulic conductance and xylem cavitation in Betula occidentalis. 1 1993
... 在全球范围内, 生态****从个体到整个生态系统对多个层次的植物功能性状进行了大量研究(Wright et al., 2007; Chave et al., 2009; Poorter et al., 2010; Pérezharguindeguy et al., 2013).其中, 叶片功能性状直接反映了植物为适应环境变化而采取的生存策略(Wardle et al., 1998).植物不同叶片性状间关系的研究一直是植物生态学的重要领域(Zhang et al., 2012; Blonder et al., 2015).然而叶片的功能性状并不能单独决定植物的生存策略, 当其与木质部结构特征无法耦合时, 就会导致植物的死亡(Breshears et al., 2009).例如, 叶片气孔开度、叶片水势与植物木质部结构紧密相关(Brodribb et al., 2010), 当植物气孔特性不能够随着木质部导水率的丧失而产生相应调整时, 就会导致叶片凋落甚至植物死亡(Sperry & Pockman, 1993).因此, 深入了解植物叶片和木质部结构之间的耦合关系是分析气候变化背景下森林群落结构变化的前提. ...
Effects of ring-porous and diffuse-?porous stem wood anatomy on the hydraulic parameters used in a water flow and storage model 1 2007
... 基于木质部导管解剖特征, 被子植物大致被分为两类, 即环孔材物种和散孔材物种.这两种植物的导管结构特征差异导致水分生理调节的显著差异(Matheny et al., 2014; Peters et al., 2015).研究发现环孔材植物具有高的水分传输能力(Niu et al., 2017), 而在水分胁迫条件下更容易遭受空穴化和栓塞胁迫, 从而降低植物对树冠的供水能力(Wang et al., 1992; Sperry et al., 1994; Steppe & Lemeur, 2007).导管直径较小的散孔材植物具有较低的水传输速率, 其水分传输速率的最大值、最小值和季节变化都很小(Sack & Holbrook, 2006).干湿季节环孔材和散孔材植物的水分蒸腾差异表明他们对水的调控机制不同(Zhu et al., 2017).近年来, 很多****对具有不同木质部特征的树木的水分运输效率和水分利用策略进行了广泛的研究, 希望从植物水力结构的角度探讨植物的水分运输, 评价树木的耐旱性(Fichot et al., 2010).有研究曾阐明早春时节环孔材和散孔材物种导管形成的时间与叶物候之间的关系(Takahashi et al., 2013), 但尚不清楚植物这种物候特征的耦合能否反映在功能性状特征上.我国亚热带地区受季风气候的影响, 干湿季分明(曾小敏等, 2017), 尤其是全球气候变化导致的降水格局变化(李维京等, 2015), 更加剧了这种干湿季节降水的差异性.如何适应这种短时间的极端环境变化是亚热带地区植被生存演替所面临的严峻挑战. ...
Relationship between the timing of vessel formation and leaf phenology in ten ring-porous and diffuse-porous deciduous tree species 1 2013
... 基于木质部导管解剖特征, 被子植物大致被分为两类, 即环孔材物种和散孔材物种.这两种植物的导管结构特征差异导致水分生理调节的显著差异(Matheny et al., 2014; Peters et al., 2015).研究发现环孔材植物具有高的水分传输能力(Niu et al., 2017), 而在水分胁迫条件下更容易遭受空穴化和栓塞胁迫, 从而降低植物对树冠的供水能力(Wang et al., 1992; Sperry et al., 1994; Steppe & Lemeur, 2007).导管直径较小的散孔材植物具有较低的水传输速率, 其水分传输速率的最大值、最小值和季节变化都很小(Sack & Holbrook, 2006).干湿季节环孔材和散孔材植物的水分蒸腾差异表明他们对水的调控机制不同(Zhu et al., 2017).近年来, 很多****对具有不同木质部特征的树木的水分运输效率和水分利用策略进行了广泛的研究, 希望从植物水力结构的角度探讨植物的水分运输, 评价树木的耐旱性(Fichot et al., 2010).有研究曾阐明早春时节环孔材和散孔材物种导管形成的时间与叶物候之间的关系(Takahashi et al., 2013), 但尚不清楚植物这种物候特征的耦合能否反映在功能性状特征上.我国亚热带地区受季风气候的影响, 干湿季分明(曾小敏等, 2017), 尤其是全球气候变化导致的降水格局变化(李维京等, 2015), 更加剧了这种干湿季节降水的差异性.如何适应这种短时间的极端环境变化是亚热带地区植被生存演替所面临的严峻挑战. ...
Spatial variations in xylem sap flux density in evergreen oak trees with radial-porous wood: Comparisons with anatomical observations 1 2008
Contrasting whole- tree water use, hydraulics, and growth in a co-dominant diffuse-porous vs. ring-porous species pair 1 2013
... 除此之外, 对水分条件变化的应对能力同样在植物生存策略中占有十分重要的地位(Milcu et al., 2013).两种植物导管密度和直径的差异决定了两者的输水策略, 即木荷具有较大的Kn, 具有较强的整树输水潜力, 从而影响叶片的水分利用策略.从叶片结构上, 我们发现, 作为环孔材的桂林栲, 其Sd显著低于散孔材的木荷(图3; p = 0.022).研究表明, 较低的叶片Sd能够使植物具有较高的Cond (Franks et al., 2009), 但当光照较强时, 其光合作用将会受到极大的影响(Büssis et al., 2006).而Sd较大的植物, 由于有较好的气孔调控能力, 气孔不仅能够在环境恶劣的条件下迅速关闭, 同时也可以在环境较好的条件下迅速打开(Drake et al., 2013).因此, 桂林栲在水分限制条件下, 也更有可能出现水分胁迫.与此同时, 我们发现木荷的LWC也显著高于桂林栲(图2), 因此对干旱的耐受性更强(Keyvan, 2010).这与上述较强的气孔调控能力相一致.植物叶片这种对干旱的应对能力, 与其导管特征是密不可分的(Klein, 2014).研究发现, 散孔材树种Acer grandidentatum的蒸腾量是环孔材树种Quercus gambelii的两倍(von Allmen et al., 2013).而在我们的另一项研究中也发现, 样地中木荷的树干液流速率也显著高于桂林栲(待发表).这就使得木荷在干季能够维持较高的Tr以及较低WUEi (图4).此外, Klein (2014)通过大量的研究发现, 当植物Cond下降50%时, 环孔材植物的叶片水势下降量是散孔材的2倍.这些结果都为木荷较高的叶片耐旱特性提供了支撑.因此, 从这些结果中我们可以发现, 散孔材较大的导管密度能够增强其叶片的耐旱能力. ...
The relation of foliar phenology to xylem embolism in trees 1 1992
... 基于木质部导管解剖特征, 被子植物大致被分为两类, 即环孔材物种和散孔材物种.这两种植物的导管结构特征差异导致水分生理调节的显著差异(Matheny et al., 2014; Peters et al., 2015).研究发现环孔材植物具有高的水分传输能力(Niu et al., 2017), 而在水分胁迫条件下更容易遭受空穴化和栓塞胁迫, 从而降低植物对树冠的供水能力(Wang et al., 1992; Sperry et al., 1994; Steppe & Lemeur, 2007).导管直径较小的散孔材植物具有较低的水传输速率, 其水分传输速率的最大值、最小值和季节变化都很小(Sack & Holbrook, 2006).干湿季节环孔材和散孔材植物的水分蒸腾差异表明他们对水的调控机制不同(Zhu et al., 2017).近年来, 很多****对具有不同木质部特征的树木的水分运输效率和水分利用策略进行了广泛的研究, 希望从植物水力结构的角度探讨植物的水分运输, 评价树木的耐旱性(Fichot et al., 2010).有研究曾阐明早春时节环孔材和散孔材物种导管形成的时间与叶物候之间的关系(Takahashi et al., 2013), 但尚不清楚植物这种物候特征的耦合能否反映在功能性状特征上.我国亚热带地区受季风气候的影响, 干湿季分明(曾小敏等, 2017), 尤其是全球气候变化导致的降水格局变化(李维京等, 2015), 更加剧了这种干湿季节降水的差异性.如何适应这种短时间的极端环境变化是亚热带地区植被生存演替所面临的严峻挑战. ...
Can comparative approaches based on plant ecophysiological traits predict the nature of biotic interactions and individual plant species effects in ecosystems? 1 1998
... 在全球范围内, 生态****从个体到整个生态系统对多个层次的植物功能性状进行了大量研究(Wright et al., 2007; Chave et al., 2009; Poorter et al., 2010; Pérezharguindeguy et al., 2013).其中, 叶片功能性状直接反映了植物为适应环境变化而采取的生存策略(Wardle et al., 1998).植物不同叶片性状间关系的研究一直是植物生态学的重要领域(Zhang et al., 2012; Blonder et al., 2015).然而叶片的功能性状并不能单独决定植物的生存策略, 当其与木质部结构特征无法耦合时, 就会导致植物的死亡(Breshears et al., 2009).例如, 叶片气孔开度、叶片水势与植物木质部结构紧密相关(Brodribb et al., 2010), 当植物气孔特性不能够随着木质部导水率的丧失而产生相应调整时, 就会导致叶片凋落甚至植物死亡(Sperry & Pockman, 1993).因此, 深入了解植物叶片和木质部结构之间的耦合关系是分析气候变化背景下森林群落结构变化的前提. ...
Can comparative approaches based on plant ecophysiological traits predict the nature of biotic interactions and individual plant species effects in ecosystems? 1 1998
... 在全球范围内, 生态****从个体到整个生态系统对多个层次的植物功能性状进行了大量研究(Wright et al., 2007; Chave et al., 2009; Poorter et al., 2010; Pérezharguindeguy et al., 2013).其中, 叶片功能性状直接反映了植物为适应环境变化而采取的生存策略(Wardle et al., 1998).植物不同叶片性状间关系的研究一直是植物生态学的重要领域(Zhang et al., 2012; Blonder et al., 2015).然而叶片的功能性状并不能单独决定植物的生存策略, 当其与木质部结构特征无法耦合时, 就会导致植物的死亡(Breshears et al., 2009).例如, 叶片气孔开度、叶片水势与植物木质部结构紧密相关(Brodribb et al., 2010), 当植物气孔特性不能够随着木质部导水率的丧失而产生相应调整时, 就会导致叶片凋落甚至植物死亡(Sperry & Pockman, 1993).因此, 深入了解植物叶片和木质部结构之间的耦合关系是分析气候变化背景下森林群落结构变化的前提. ...
Plant ecological strategies: Some leading dimensions of variation between species 1 2002
... 环境胁迫诱导的基于植物功能性状的生存策略变化一直是生态学研究中的关键问题(Díaz et al., 1998; Díaz & Cabido, 2001).通过揭示不同物种采用的生存策略, 可为了解气候变化下的森林的结构和演替规律提供基础(Hulot et al., 2000; Westoby et al., 2002; Milcu et al., 2013). ...
Relationships among ecologically important dimensions of plant trait variation in seven neotropical forests 1 2007
... 在全球范围内, 生态****从个体到整个生态系统对多个层次的植物功能性状进行了大量研究(Wright et al., 2007; Chave et al., 2009; Poorter et al., 2010; Pérezharguindeguy et al., 2013).其中, 叶片功能性状直接反映了植物为适应环境变化而采取的生存策略(Wardle et al., 1998).植物不同叶片性状间关系的研究一直是植物生态学的重要领域(Zhang et al., 2012; Blonder et al., 2015).然而叶片的功能性状并不能单独决定植物的生存策略, 当其与木质部结构特征无法耦合时, 就会导致植物的死亡(Breshears et al., 2009).例如, 叶片气孔开度、叶片水势与植物木质部结构紧密相关(Brodribb et al., 2010), 当植物气孔特性不能够随着木质部导水率的丧失而产生相应调整时, 就会导致叶片凋落甚至植物死亡(Sperry & Pockman, 1993).因此, 深入了解植物叶片和木质部结构之间的耦合关系是分析气候变化背景下森林群落结构变化的前提. ...
Productive leaf functional traits of Chinese savanna species 1 2012
... 在全球范围内, 生态****从个体到整个生态系统对多个层次的植物功能性状进行了大量研究(Wright et al., 2007; Chave et al., 2009; Poorter et al., 2010; Pérezharguindeguy et al., 2013).其中, 叶片功能性状直接反映了植物为适应环境变化而采取的生存策略(Wardle et al., 1998).植物不同叶片性状间关系的研究一直是植物生态学的重要领域(Zhang et al., 2012; Blonder et al., 2015).然而叶片的功能性状并不能单独决定植物的生存策略, 当其与木质部结构特征无法耦合时, 就会导致植物的死亡(Breshears et al., 2009).例如, 叶片气孔开度、叶片水势与植物木质部结构紧密相关(Brodribb et al., 2010), 当植物气孔特性不能够随着木质部导水率的丧失而产生相应调整时, 就会导致叶片凋落甚至植物死亡(Sperry & Pockman, 1993).因此, 深入了解植物叶片和木质部结构之间的耦合关系是分析气候变化背景下森林群落结构变化的前提. ...
Quantifying the hydrological responses to climate change in an intact forested small watershed in Southern China 1 2011
 ,1,*, 赵平2, 张锦秀1, 斯瑶1
,1,*, 赵平2, 张锦秀1, 斯瑶1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT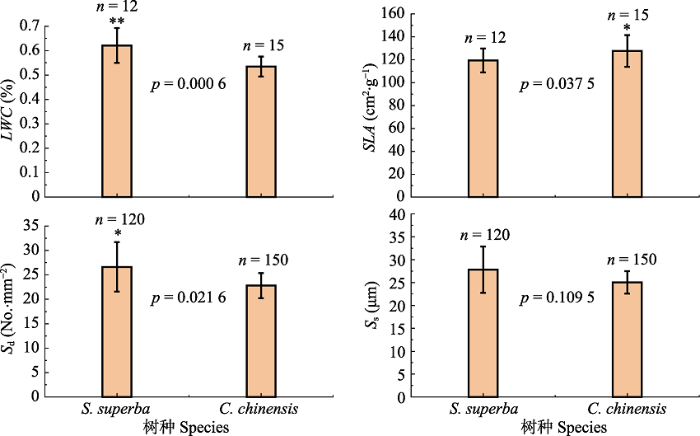 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT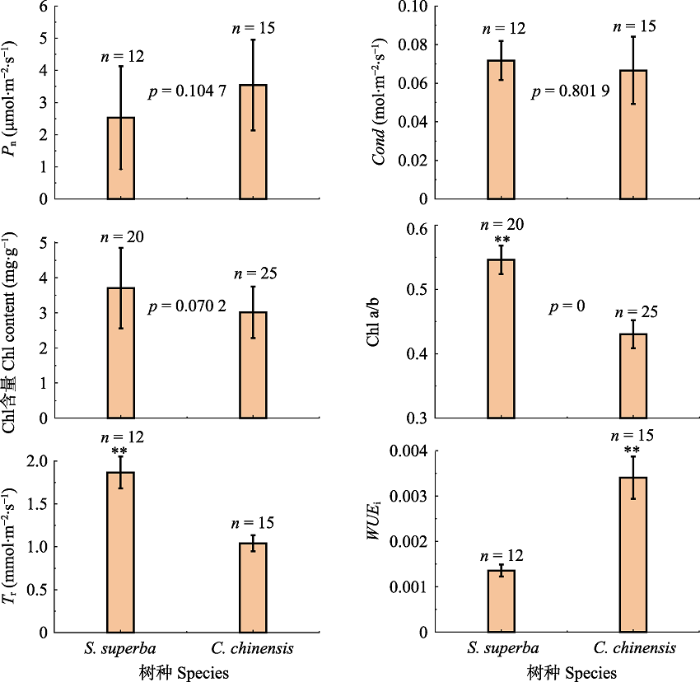 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT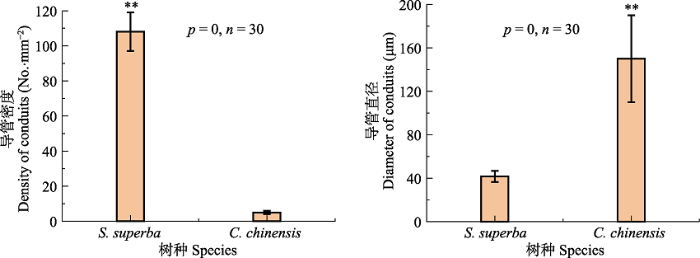 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}