 ,*, 倪健
,*, 倪健 ,,*浙江师范大学化学与生命科学学院, 浙江金华 321004
,,*浙江师范大学化学与生命科学学院, 浙江金华 321004What is the optimal number of leaves when measuring leaf area of tree species in a forest community?
GAO Si-Han, GE Yu-Xi, ZHOU Li-Yi, ZHU Bao-Lin, GE Xing-Yu, LI Kai,*, NI Jian,,*College of Chemistry and Life Sciences, Zhejiang Normal University, Jinhua, Zhejiang 321004, China通讯作者: Li K:likai@zjnu.edu.cnNi J:nijian@zjnu.edu.cn倪健ORCID: 0000-0001-5411-7050
编委: 阎恩荣
责任编辑: 李 敏
收稿日期:2018-04-16修回日期:2018-06-09网络出版日期:2018-09-20
| 基金资助: |
Received:2018-04-16Revised:2018-06-09Online:2018-09-20
| Fund supported: |

摘要
植物形态性状叶面积简单易测, 能够反映植物对环境的适应与响应, 指示生态系统的功能与过程。在野外测定叶面积时, 叶片取样数量往往采用约定俗成的10-20片, 但到底采集多少叶片才是最优和最具代表性, 却少有探究。该研究以浙江金华山常绿落叶阔叶混交林的优势树种木荷(Schima superba)与枫香树(Liquidambar formosana)为研究对象, 通过对5个胸径等级植株和每个植株6个方位开展大批量叶片取样(>2 500个), 分析两个树种的叶面积变异特征, 探讨叶片取样数量为多少才能最代表该森林类型的叶片大小性状规律。结果表明, 常绿乔木木荷平均叶面积与变幅均小于落叶乔木枫香树。木荷叶面积与胸径无显著相关性, 而枫香树叶面积与胸径有较显著相关性, 但两个树种均在中胸径等级(15-20 cm)差异不显著; 两个树种的叶面积与采样方位无显著相关性, 但在东、西和底部的差异不显著。因此, 综合考虑代表性与野外可操作性, 叶片采集首选中胸径成树的底部叶片。随机抽样统计可知, 树木叶面积测定的最适叶片采集数量因物种而异, 木荷的最适叶片采集数量为40, 而枫香树最少为170片。因此, 在叶面积测定时, 叶片采集的数量应该不能只局限在10-20片, 在人力、物力和时间等条件允许的情况下, 应该尽可能多地测定较多叶片的叶面积。
关键词:
Abstract
Keywords:
PDF (1456KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
高思涵, 葛珏希, 周李奕, 朱宝琳, 葛星宇, 李凯, 倪健. 测定森林树木叶面积的最适叶片数是多少?. 植物生态学报[J], 2018, 42(9): 917-925 DOI:10.17521/cjpe.2018.0087
GAO Si-Han, GE Yu-Xi, ZHOU Li-Yi, ZHU Bao-Lin, GE Xing-Yu, LI Kai, NI Jian.
叶片是植物光合作用和物质生产的主要器官, 指示叶片大小的性状, 如叶面积(叶片单面投影表面积, mm2), 可反映植物对环境的响应与适应, 指示生态系统的功能与过程, 与环境胁迫、干扰和植物的生长对策及其利用资源的能力密切相关, 是重要的植物功能性状之一(Wright et al., 2004; 孟婷婷等, 2007; 刘晓娟和马克平, 2015; Díaz et al., 2016)。相对于茎干与根系性状, 叶片易于采样, 性状易于测定, 可以通过大批量的采样来比较和分析不同地区、不同生态系统和不同植物种类的功能性状特征。同时, 叶面积性状还可拓展用以计算比叶面积、基于面积的叶片营养元素含量等, 由此与其他叶性状相互关联所形成的叶经济谱(Wright et al., 2004; Donovan et al., 2011; Díaz et al., 2016), 可量化植物利用环境资源的“投资-收益”权衡策略(陈莹婷和许振柱, 2014), 综合反映植物功能性状与环境和生态系统功能的关系, 对模拟和预测气候变化对生态系统的影响, 指导生物多样性保护和生态系统管理政策的制定等具有重要的生态学意义(刘晓娟和马克平, 2015; 王常顺和汪诗平, 2015; Díaz et al., 2016)。
在野外测定叶面积时, 针对一个样地中的目标植物, 除了极其特殊的情形, 如无叶或鳞叶植物、针叶植物等, 通常是每种植物选择10株成熟个体, 随机采摘相对幼嫩但伸展、硬化且无病虫害、无附生植物的健康叶片20个; 至少保证每个物种采集10个叶片, 在叶片稀少时最起码也需要5个植株, 每个植株采摘2个叶片, 或者在成熟个体稀少时最少有4个植株、每个植株采摘5个叶片; 保湿、冷藏储存, 并尽快利用可自动测量的叶面积仪(如LI-COR或Delta-T)或者人工测定的扫描仪进行叶面积测定(Cornelissen et al., 2003; Pérez-Harguindeguy et al., 2013)。然而, 受系统进化地位所决定的科属分类以及诸多环境因素的影响, 植物的叶片大小存在显著的种间变异与较显著的种内变异(Westoby et al., 2002; Albert et al., 2010; Bolnick et al., 2011; Auger & Shipley, 2013; Siefert et al., 2015; Messier et al., 2017)。因此, 在同一地点与不同地点, 同一植物生长于不同生境, 同一植物的不同植株, 甚至同一植株的不同部位(如阴面、阳面), 叶片大小均会存在差异。因此在野外实际采样过程中, 比如针对森林中的优势木本植物, 在理想情况下, 一般会分别选择不同胸径等级或仅大胸径等级的植株3-5株, 在每个植株的向阳部位(冠层)随机采摘5-10个叶片, 或者在每个植株的四个方位与顶部随机分别摘取3-5个叶片, 再进行叶面积的扫描测定。但成熟大树一般较高大, 冠层位置高, 无法有效获取叶片; 而幼小树木的代表性又不强。
那么, 测定叶面积时需要摘取一定数量的叶片作为重复, 但叶片大小具有显著的种内与种间差异, 给野外取样带来一个很大的困惑: 对每一个植株, 叶片取样数量多少才算合适?对一种植物, 采集多少个植株才能代表一个植物种叶片大小的普遍特征?取样数量少, 不一定能完全反映植物叶片性状特征; 而取样数量多, 则耗费太多的人力和时间。叶片取样质与量的权衡, 是目前一个需要解决的问题。
因此, 本研究以浙江省金华市金华山的常绿落叶阔叶混交林中的两种优势树种为对象, 通过设计多植株、大批量叶片采样, 统计分析同一树种内不同植株在不同生境中的叶面积变异特征, 从而回答选择多少植株, 采集多少叶片, 才能代表一种植物的叶片大小特征的真实值这个科学问题。本研究可为今后的叶片性状测定取样数量(最适值)提供证据, 以最优的叶片取样数量、最少的时间与人力成本而达到最适的测定效果。该研究也可进一步对确定其他植物器官性状的取样数量具有借鉴意义。
1 材料和方法
1.1 研究区概况
金华山地处中亚热带北缘, 位于浙江省中西部金华市北15 km处, 金衢盆地东部, 系龙门山脉的支脉, 因横亘于金华市北, 故俗称北山。整个山体跨越119.48°-119.82° E, 29.15°-29.27° N, 海拔通常在100-1 200 m, 主峰大盘尖海拔1 314 m。研究区位于金华山南坡双龙洞国家森林公园内, 属亚热带山地季风气候。据金华气象站(119.65° E, 29.12° N, 海拔626 m) 1953-2014年的纪录, 该地区年平均气温为17.7 ℃, 1月平均气温5.2 ℃, 7月平均气温29.5 ℃, 极端低温达-9.6 ℃, 极端高温为41.5 ℃; 年降水量可达1 436.8 mm, 主要集中在3-9月, 占全年降水量的76.7%, 但以6月份梅雨季节的降水量最大, 平均达252.2 mm, 3-6月平温期占降水集中期降水量的70% (平均月降水量191 mm), 而7-9月高温期降水反而不是最高(平均月降水量112 mm); 年日照率为43%, 无霜期256天。该地区土壤以亚热带山地黄壤为主, 植被具有亚热带常绿阔叶林、常绿落叶阔叶林和灌丛等垂直带谱, 但受人为干扰影响大, 是人为干扰下典型的亚热带常绿阔叶林退化植被, 山脚至中坡(200-700 m)以次生性的常绿阔叶林、常绿落叶阔叶混交林和针阔混交林、次生灌丛为主, 中坡至上坡(600-1000 m)主要为马尾松(Pinus massoniana)人工林、杉木(Cunninghamia lanceolata)林、柳杉(Cryptomeria fortunei)林、黄山松(Pinus taiwanensis)林和山地灌丛(郭水良等, 1993; 方芳和郭水良, 2004)。1.2 野外采样与测试
取样点位于119.62° E、29.20° N附近山谷两侧的常绿落叶阔叶混交林, 山体较陡峭, 坡度20°-40°。群落乔木层优势树种以木荷(Schima superba)、青冈(Cyclobalanopsis glauca)、枫香树(Liquidambar formosana)为主, 其次是苦槠栲(Castanopsis sclerophylla)、柯(Lithocarpus glaber)、锥栗(Castanea henryi)等, 少数马尾松大树散生其间。树高10-15 m, 胸径10-25 cm, 郁闭度为0.7。灌木层主要由格药柃(Eurya muricata)、南烛(Vaccinium bracteatum)、荚蒾(Viburnum dilatatum)、算盘子(Glochidion puberum)等组成, 草本层由蕨(Pteridium sp.)、鳞毛蕨(Dryopteris sp.)、三脉紫菀(Aster ageratoides)、芒(Miscanthus sp.)等组成。选取常绿树种木荷与落叶树种枫香树为研究对象, 前者取样海拔范围305-794 m, 后者457- 635 m。在2017年7月中下旬, 按照不同的胸径等级: 5-10 cm (D2)、10-15 cm (D3)、15-20 cm (D4)、20- 30 cm (D5)和30-45 cm (D6), 对前3个等级各选择20株、后2个等级分别选择15和10株进行取样(大树个体较少), 等级D1 (0-5 cm)无植物个体。对每个植株记录其编号和位置、胸径大小、树高、经纬度、海拔、坡度和坡向等。同时, 将树木冠层划分为上中下三部分, 利用高枝剪等采样工具, 在其树冠的底部, 树冠中部的东、南、西、北方位, 以及树冠上部开阔区域的顶部, 共6个方位, 分别采摘成熟、健康、无病害的叶片5片, 每株树合计采集叶片30片, 编号封装在密封袋内, 置于便携式冷藏箱内储存。共采集叶片5 087片, 其中木荷2 543片, 枫香树2 544片。在采集叶片的当天, 利用便携式叶面积仪(LI-3000C, LI-COR, Lincoln, USA)扫描测量每个叶片的叶面积。
1.3 统计分析
利用SPSS 20.0计算木荷与枫香树2个树种、5个胸径等级、10-20株树木、6个采样方位的叶面积平均值、标准误差、四分位统计及其95%置信区间; 基于独立样本t检验计算不同胸径、采样方位叶面积的差异性, 获取不同树种在不同胸径等级与不同方位的单个叶面积与其总平均值间的偏离程度, 并区分未成熟树木(胸径<20 cm)和成熟树木(胸径≥20 cm)进一步分析统计。利用R 3.3软件中的Sample程序进行所有叶面积样本以及有限制条件的叶面积样本、有放回的随机抽样, 以比较不同取样数量下叶面积均值及其置信区间的偏移, 利用t检验获取目标样本的均值和均值的95%置信区间, 并将目标样本与总体样本之间进行独立样本t检验。如果随机选取的叶面积与总体叶面积没有显著性差异, 则为最优取样数量。所有图形均利用Golden Software Grapher 9.1.5软件绘制。2 结果
2.1 叶面积特征
所有叶片的统计分析表明, 木荷和枫香树叶面积总体呈正态分布, 木荷叶面积平均为(41.60 ± 10.88) cm2, 介于16.74-100.80 cm2之间(图1A), 而枫香树平均值为(57.65 ± 19.35) cm2, 变化范围为11.31-129.51 cm2 (图1B)。二者叶面积差异较大, 无论是叶面积还是变幅, 常绿乔木木荷均小于落叶乔木枫香树。图1
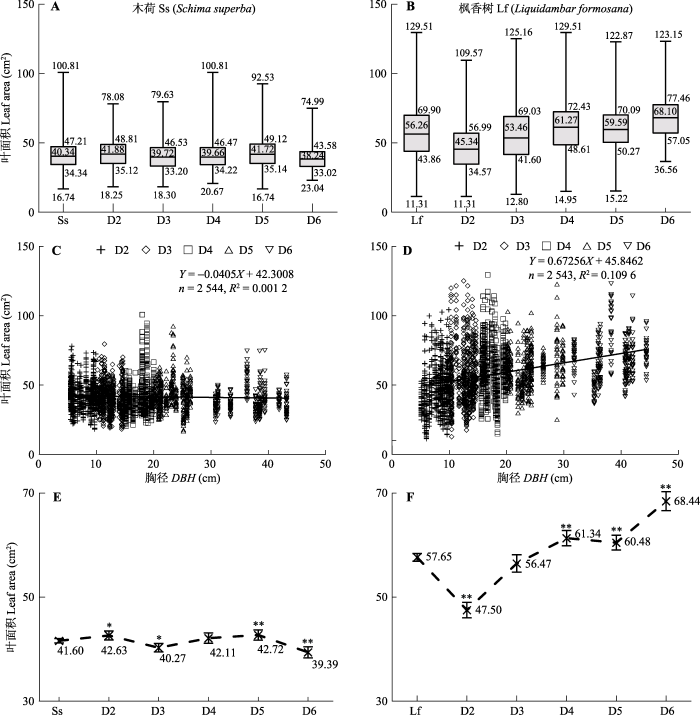 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1两个树种叶面积与胸径的关系。A, B, 四分位图; C, D, 散点图; E, F, 置信区间图。Ss, Lf, 木荷与枫香树所有胸径等级样品的平均值。D2-D6, 胸径等级D2-D6。*, p < 0.05; **, p < 0.01。
Fig. 1Relationship between leaf area and diameter at breast height of two tree species. A, B, Quartile map; C, D, Scatter plot; E, F, Confidence interval. Ss, Lf, mean values of all samples in all classes of diameter at breast height for Schima superba and Liquidambar formosana, respectively. D2-D6, D2-D6 classes of diameter at breast height. *, p < 0.05; **, p < 0.01.
木荷的叶片大小与胸径等级之间呈现极不显著的负相关性(图1C), 而枫香树的叶面积却与胸径之间呈现较显著的正相关性(图1D)。在不同胸径等级之间, 木荷的叶面积差异不显著, 仅在D4与D5两个胸径等级表现出较大的变化区间(图1E); 但枫香树在不同胸径等级却有较显著的差异(图1D), 在所有胸径等级均表现出较大的变化区间, 尤其是D2和D5两个胸径等级。木荷叶面积的平均值和置信区间在第3和第6胸径等级有轻微异常偏移(图1E), 而枫香树叶面积则在第2和第6胸径等级处有较大的异常位移, 胸径越大, 置信区间也越向上偏移(图1F), 说明其叶面积与胸径间的正相关关系越密切。
木荷与枫香树的叶面积与采样方位无相关性(图2A、2B), 在不同采样方位之间差异也不显著(图2C、2D), 但木荷在南部方向的叶面积大于其他方位的, 顶部和西部的叶面积最小(图2C); 枫香树的叶面积在南北两个方向均较大, 在东西两个方向较小(图2D), 且上述所有方位的置信区间均较显著上移或下移(图2E、2F)。总体来看, 叶面积随采样方位的偏移(图2E、2F)明显小于随胸径的偏移(图1E、1F)。
图2
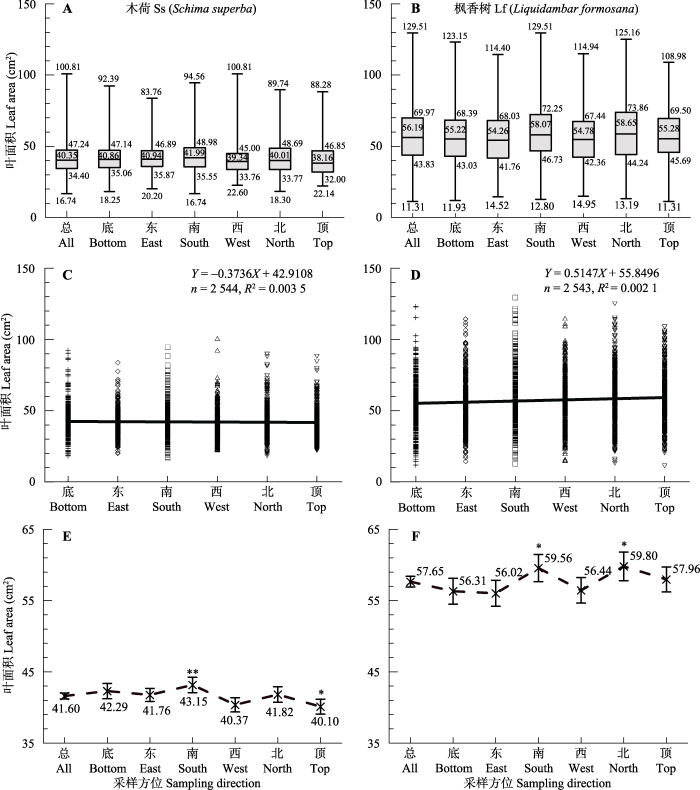 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2两个树种叶面积与采样方位的关系。A, B, 四分位图; C, D, 散点图; E, F, 置信区间图。*, p < 0.05; **, p < 0.01。
Fig. 2Relationship between leaf area and sampling direction of two tree species. A, B, Quartile map; C, D, Scatter plot; E, F, Confidence interval. *, p < 0.05; **, p < 0.01.
2.2 叶片取样对象与方位
木荷的大胸径等级(D5-D6)个体的叶面积偏离均值极显著(p < 0.01), 小胸径等级(D2-D3)显著(p < 0.05), 而中等胸径(D4)则不显著(图3A); 其在南向和顶部分别偏离极显著与显著, 其他方位则不显著(图3B)。对枫香树而言, 除胸径等级D3之外, 其他所有胸径的个体叶面积均极显著偏离均值(图3C); 而其只有南和北两个方位显著偏离均值, 其他方位都不显著(图3D)。图3
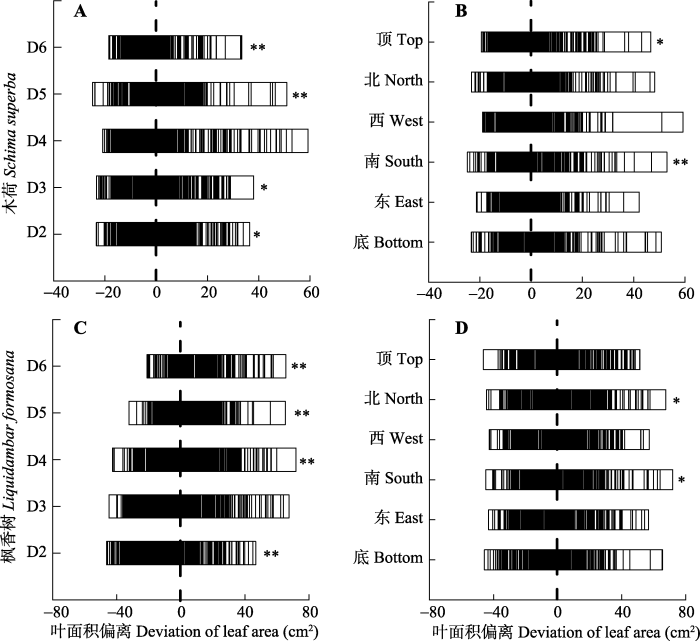 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3两个树种叶面积在胸径与采样方位的偏离。A, 木荷胸径。B, 木荷采样方位。C, 枫香树胸径。D, 枫香树采样方位。*, p < 0.05; **, p < 0.01。竖线表示归零后的均值。
Fig. 3Deviation of leaf area to diameter and sampling direction of two tree species. A, Diameter of Ss; B, Sampling direction of Ss; C, Diameter of Lf; D, Sampling direction of Lf. *, p < 0.05; **, p < 0.01. The vertical line indicates the mean value returning to zero.
以成树和非成树单独进行统计检验(图4)发现, 两种树木的成树与非成树在不同采样方位的叶面积差异不显著(未展示)。木荷由于不同胸径和叶面积差异较小, 所以成树与非成树的均值与所有树木总均值之间的差异不大。而枫香树叶面积在成树和非成树间表现出明显的差异性, 若以成树叶面积均值代替总叶面积均值则导致总叶面积呈显著的负偏移, 若以非成树叶面积均值代替总叶面积均值则导致叶面积出现整体正偏移。
图4
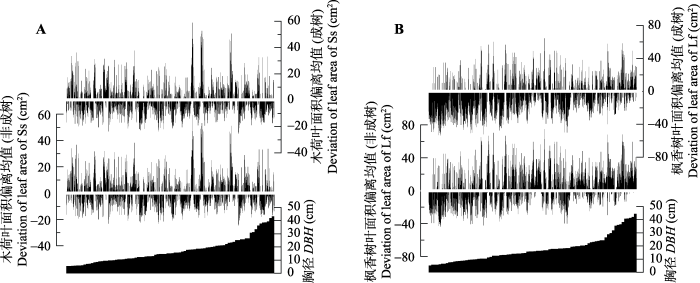 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4两个树种成树与非成树叶面积在胸径上的偏离。A, 木荷。B, 枫香树。横线表示归零后的均值。
Fig. 4Deviation of mature and non-mature leaf areas to diameter at breast height (DBH) of two tree species. A, Schima superba (Ss). B, Liquidambar formosana (Lf). The horizontal line indicates the mean value returning to zero.
综合比较可以看出, 中等胸径的植株叶片是最适的取样材料, 而东西向与底部的叶片是可选的, 但考虑到东方与西方叶片位置较高, 那么选择底部的叶片也不失一个良策。
2.3 叶片取样数量
以所有胸径等级、采样方位的叶片样本为基础进行的随机抽样与检验表明, 木荷随机抽样大于734片(图5A), 枫香树大于1 157片(图5C), 所获得的目标样本与总样本间才无显著性差异。因此, 在不限定胸径等级和取样方位前提下, 734和1 157分别是木荷与枫香树叶面积性状研究所需要的最低取样数量。如果只考虑最佳的取样对象和方位, 即以木荷D4胸径和枫香树D3胸径等级的底部叶片进行随机取样, 那么木荷与枫香树叶片的取样数量以40 (图5B)和170片(图5D)为最适。图5
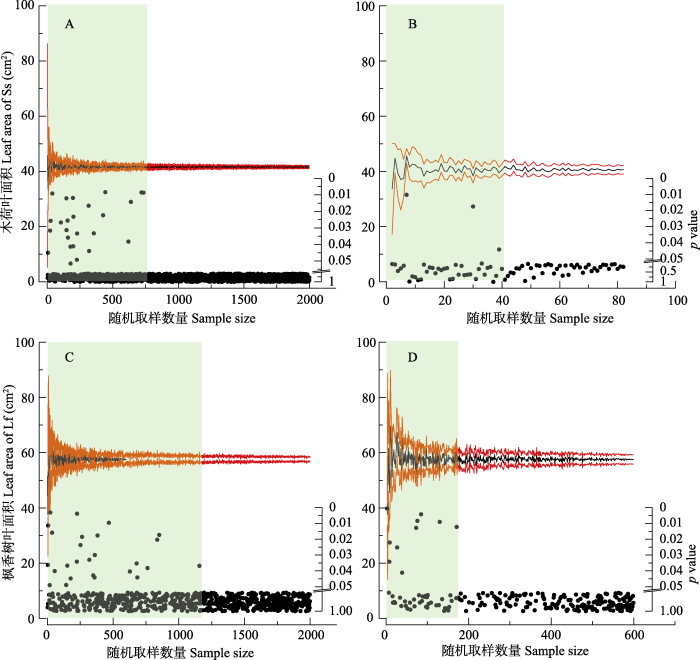 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5两个树种叶面积的随机抽样均值和95%置信区间及其差异性检验。A, C, 全部叶片样品。B, D, 中等胸径等级、底部叶片样品。叶面积图上的黑色线代表随机抽样均值, 红色线代
Fig. 5Mean of random sampling, 95% confidence interval and significant test of leaf areas of two tree species. A, C, All leaf samples. B, D, Leaf samples of medium diameter class and bottom sampling direction. The black line of the leaf area plot indicates the mean value of random sampling, and the red line indicates the 95% confidence interval; the light blue block is the area of significant difference, indicating that the sample size of the right position of the block is the optimal sampling number of leaves. Ss, Schima superba; Lf, Liquidambar formosana.
3 讨论
植物的分类地位与环境条件决定其性状特征, 包括叶面积。植物叶面积存在较大的种间变异(Westoby et al., 2002; Albert et al., 2010; Bolnick et al., 2011; Violle et al., 2012; Siefert et al., 2015;Messier et al., 2017), 因此不同物种叶性状测定时的叶片取样数量应该不同。而叶片大小的种内变异也同样不容忽视(Auger & Shipley, 2013; Bu et al., 2017), 因此在采集同一个物种的叶片时也应该考虑不同植株的差异。另外, 气候、土壤、光照、海拔、地形等要素也会影响植物叶片的大小(McDonald et al., 2003; Royer et al., 2008; Peppe et al., 2011), 同一物种的不同植株间和同一个植株内, 叶片大小也可能存在差异。因此, 在测定植物叶片大小时, 按照传统和约定俗成地采摘成熟植株的健康叶片10-20片(Cornelissen et al., 2003; Pérez-Harguindeguy et al., 2013), 在有限的人力、物力和时间条件下, 是可以接受的; 但对于植物性状的精细研究来说, 10-20片叶片是远远不足的, 应该针对不同植物采集不同数量的叶片开展研究。本研究中木荷的最适采样数量为40片叶片, 枫香树为170片叶片, 最适采样数量与树种有关。木荷为常绿树种, 叶片为较小单叶, 叶面积变异小, 因此最适的叶片数量较小。而枫香树为落叶树种, 叶片为较大单叶, 叶面积变异大, 因此最适的叶片数量大于木荷。同时, 常绿树种叶寿命长, 叶面积建设成本高、消耗大, 叶片变异大则不利于其适应环境; 而落叶树种叶寿命短, 建设成本低, 在不同环境、不同树冠部位, 乃至不同发育阶段采取不同方式适应环境, 因而其叶片变异比常绿树种大(Aerts, 1995; Givnish, 2002), 这也是木荷叶片最适采样数量小于枫香树的一个重要原因。
另外, 传统的方法对采样植株的年龄与采样方位没有具体规定, 通常在野外操作时都是针对大树向阳的叶片。本研究通过对亚热带常绿落叶阔叶混交林的一个常绿与一个落叶优势树种的研究发现, 中龄植株树冠底部是较合适的叶片采样对象与方位。树龄影响叶片大小, 在种间表现出一定的差异, 木荷叶片大小与树龄关系不密切, 而枫香树则较密切, 二者中龄树的叶片大小与总叶片大小的关系一致。而取样方位, 亦即光照强度同样影响叶片大小(李明财等, 2009), 两个树种的叶片大小与取样方位的关系也与上述树龄关系相同, 但密切程度较小; 通常只是在南、北与顶部的叶片大小变异较大, 东、西、底部的变异相对较小, 考虑到野外的可操作性, 以底部采样为宜。
上述研究结果可能也适用于其他器官性状测定的采样, 在今后的植物性状研究中, 无论是叶片还是茎干、枝条或者根系, 都应该采集较多的植物样本, 并兼顾不同树龄和不同方位。但对不同的植物器官来说, 到底采集多少数量才是最适的, 仍需要进行更多植物的观测与实验。同时, 也需要更多注意不同气候带的优势植物的性状., 包括乔木、灌木和草本植物, 以期发现不同气候带、不同植被类型中, 植物器官采样数量的差异。

致谢
感谢浙江师范大学重点建设项目(2017XM023和2017PT009)资助。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURLPMID [本文引用: 1]

Recent research shows that the dominance of evergreen species in nutrient-poor environments can be explained by their low nutrient loss rates. From this work It appears that the plant traits that are associated with low nutrient loss rates lead to low maximum-dry-matter production and to low rates of litter decomposition. This suggests a positive feedback between the evergreen habit and low nutrient availability. The growth characteristics of evergreens lead to a low responsiveness to environmental changes. As a result, global warming may lead to changes in the distribution of evergreens.
DOIURL [本文引用: 2]
1. Functional traits are increasingly used to investigate community structure, ecosystem functioning or to classify species into functional groups. These functional traits are expected to be variable between and within species. Intraspecific functional variability is supposed to influence and modulate species responses to environmental changes and their effects on their environment. However, this hypothesis remains poorly tested and species are mostly described by mean trait values without any consideration of variability in individual trait values.2. In this study, we quantify the extent of intraspecific plant functional trait variability, its spatial structure and its response to environmental factors. Using a sampling design structured along two direct and orthogonal climatic gradients in an alpine valley, we quantified and analysed the intraspecific variability for three functional traits (height, leaf dry matter content and leaf nitrogen content) measured on sixteen plant species with contrasting life histories.3. Results showed a large variability of traits within species with large discrepancies between functional traits and species. This variability did not appear to be structured within populations. Between populations, the overall variability was partly explained by the selected gradients. Despite the strong effects of temperature and radiation on trait intraspecific variability, the response curves of traits along gradients were partly idiosyncratic.4. Synthesis. Giving a comprehensive quantification of intraspecific functional variability through the analysis of an original data set, we report new evidence that using a single trait value to describe a given species can hide large functional variation for this species along environmental gradients. These findings suggest that intraspecific functional variability should be a concern for ecologists and its recognition opens new opportunities to better understand and predict ecological patterns in a changing environment. Further analyses are, however, required to compare inter- and intraspecific variability.
DOIURL [本文引用: 2]
DOIURLPMID [本文引用: 2]
Natural populations consist of phenotypically diverse individuals that exhibit variation in their demographic parameters and intra- and inter-specific interactions. Recent experimental work indicates that such variation can have significant ecological effects. However, ecological models typically disregard this variation and focus instead on trait means and total population density. Under what situations is this simplification appropriate? Why might intraspecific variation alter ecological dynamics? In this review we synthesize recent theory and identify six general mechanisms by which trait variation changes the outcome of ecological interactions. These mechanisms include several direct effects of trait variation per se and indirect effects arising from the role of genetic variation in trait evolution.
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
The concept of leaf economics spectrum (LES) has attracted much attention and debate since its emergence. It for the first time provides quantitative analysis of plant functional traits and their relationships on the global scale, hence quantifying and generalizing the context and variations of the trade-off strategies. This is of great theoretical value, and provides a useful research method and scientific ideas for subsequent study on plant traits and their functions. In this paper, we try to comprehensively review the meaning, contents, relevant verifications and objections about LES, and to explore its underlying mechanisms. In addition, we emphasize the multi-scale and multidimensional extensions, integration and potential applications of LES. Currently there are still several shortcomings about LES research in China, and we outlook the development of LES theory domestically and abroad. It may be of significance for ecological researchers to establish and exploit jointly a global database on plant traits.
DOIURL [本文引用: 1]
The concept of leaf economics spectrum (LES) has attracted much attention and debate since its emergence. It for the first time provides quantitative analysis of plant functional traits and their relationships on the global scale, hence quantifying and generalizing the context and variations of the trade-off strategies. This is of great theoretical value, and provides a useful research method and scientific ideas for subsequent study on plant traits and their functions. In this paper, we try to comprehensively review the meaning, contents, relevant verifications and objections about LES, and to explore its underlying mechanisms. In addition, we emphasize the multi-scale and multidimensional extensions, integration and potential applications of LES. Currently there are still several shortcomings about LES research in China, and we outlook the development of LES theory domestically and abroad. It may be of significance for ecological researchers to establish and exploit jointly a global database on plant traits.
DOIURL [本文引用: 2]
DOIURLPMID [本文引用: 3]
Abstract Earth is home to a remarkable diversity of plant forms and life histories, yet comparatively few essential trait combinations have proved evolutionarily viable in today's terrestrial biosphere. By analysing worldwide variation in six major traits critical to growth, survival and reproduction within the largest sample of vascular plant species ever compiled, we found that occupancy of six-dimensional trait space is strongly concentrated, indicating coordination and trade-offs. Three-quarters of trait variation is captured in a two-dimensional global spectrum of plant form and function. One major dimension within this plane reflects the size of whole plants and their parts; the other represents the leaf economics spectrum, which balances leaf construction costs against growth potential. The global plant trait spectrum provides a backdrop for elucidating constraints on evolution, for functionally qualifying species and ecosystems, and for improving models that predict future vegetation based on continuous variation in plant form and function.
DOIURLPMID [本文引用: 1]
The worldwide leaf economic spectrum (WLES) is a strikingly consistent pattern of correlations among leaf traits. Although the WLES effectively summarizes variation in plant ecological strategies, little is known about its evolution. We reviewed estimates of natural selection and genetic variation for leaf traits to test whether the evolution of the WLES was limited by selection against unfit trait combinations or by genetic constraints. There was significant selection for leaf traits on both ends of the WLES spectrum, as well as significant genetic variation for these traits. In addition, genetic correlations between WLES traits were variable in strength and direction. These data suggest that genetic constraints have had a smaller role than selection in the evolution of the WLES.
DOIURL [本文引用: 1]
调查了浙江金华北山38个样地 的38种主要树种的生态重要值,应用除趋势对应分析方法(detrendedcorrespondenceanalysis,DCA)对该地区的木本植物 群落类型进行了定量分析.结果表明,浙江金华北山存在11个比较典型的群丛,它们分别为:黄山松(Pinustaiwanensis)群丛、马尾松 (Pinusmassoniana)群丛、锥栗(Castaneahennyi)+木荷(Schimasuperba)群丛、木荷+苦槠 (Castanopsisderophylla)群丛、马尾松+木荷群丛、枫香(Liq uidambarformosana)+马尾松群丛、枫香群丛、香樟(Cinnamomumcamphora)群丛、马尾松+短柄抱 (Quercusdlan dulifera)+杜鹃(Phodedendronsinsii)+木荷群丛、木荷+马尾松群丛和马尾松+黄山松群丛.分析了11个群丛的成员组成及其 与环境间的关系,计算了它们的物种多样性指数.
DOIURL [本文引用: 1]
调查了浙江金华北山38个样地 的38种主要树种的生态重要值,应用除趋势对应分析方法(detrendedcorrespondenceanalysis,DCA)对该地区的木本植物 群落类型进行了定量分析.结果表明,浙江金华北山存在11个比较典型的群丛,它们分别为:黄山松(Pinustaiwanensis)群丛、马尾松 (Pinusmassoniana)群丛、锥栗(Castaneahennyi)+木荷(Schimasuperba)群丛、木荷+苦槠 (Castanopsisderophylla)群丛、马尾松+木荷群丛、枫香(Liq uidambarformosana)+马尾松群丛、枫香群丛、香樟(Cinnamomumcamphora)群丛、马尾松+短柄抱 (Quercusdlan dulifera)+杜鹃(Phodedendronsinsii)+木荷群丛、木荷+马尾松群丛和马尾松+黄山松群丛.分析了11个群丛的成员组成及其 与环境间的关系,计算了它们的物种多样性指数.
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
通过测定不同光照条件下(15%透光、30%透光、60%透光、全光)东北次生林主要树种(红松、红皮云杉、胡桃楸、水曲柳和黄波罗)的比叶面积,探讨了不同树种对光照变化的响应与适应。结果表明:不同树种比叶面积存在明显的差异,尤其是常绿针叶树种与落叶树种之间差异显著(P〈0.001),指示不同功能型树种存在不同的生存策略;随着光照强度降低,所有树种比叶面积呈现增加的趋势,尤其是弱光照(15%)和低光照(30%)相对于全光和中光照(60%)差异显著(P〈0.05);不同树种比叶面积随光照强度的变化幅度明显不同,且变化最显著的光照区域也有所不同;不同年龄红松比叶面积的变化趋势一致,但变幅不同,4年生幼苗对光照响应则更为明显。本研究结果表明,所有树种均能通过改变叶形态来适应光照条件的变化,在低光条件下通过增加单位质量叶面积,提高叶片光捕获能力;但各树种对光照强度变化的不同响应则表明树种之间不同的光适应机制,这对森林生态系统内部树种的共生具有重要意义;年龄较小的幼苗对光照的强响应指示其在植物更新过程中叶片水平上生理生态学机理研究中的重要性。
URL [本文引用: 1]
通过测定不同光照条件下(15%透光、30%透光、60%透光、全光)东北次生林主要树种(红松、红皮云杉、胡桃楸、水曲柳和黄波罗)的比叶面积,探讨了不同树种对光照变化的响应与适应。结果表明:不同树种比叶面积存在明显的差异,尤其是常绿针叶树种与落叶树种之间差异显著(P〈0.001),指示不同功能型树种存在不同的生存策略;随着光照强度降低,所有树种比叶面积呈现增加的趋势,尤其是弱光照(15%)和低光照(30%)相对于全光和中光照(60%)差异显著(P〈0.05);不同树种比叶面积随光照强度的变化幅度明显不同,且变化最显著的光照区域也有所不同;不同年龄红松比叶面积的变化趋势一致,但变幅不同,4年生幼苗对光照响应则更为明显。本研究结果表明,所有树种均能通过改变叶形态来适应光照条件的变化,在低光条件下通过增加单位质量叶面积,提高叶片光捕获能力;但各树种对光照强度变化的不同响应则表明树种之间不同的光适应机制,这对森林生态系统内部树种的共生具有重要意义;年龄较小的幼苗对光照的强响应指示其在植物更新过程中叶片水平上生理生态学机理研究中的重要性。
DOIURL [本文引用: 2]
Plant functional traits are plant core attributes closely related to plant colonization, survival, growth and mortality. These core attributes could significantly affect ecosystem functions and reflect the response of vegetation to environmental change. There is growing recognition that many ecological questions at various levels can be addressed effectively with key functional traits. In this paper, we review key research progress over the last 10 years addressing important ecological issues based on functional traits, including global distribution patterns of functional traits and their associations, variation of traits distribution along environmental gradients, functional diversity and its application, mechanisms of community assembly and dynamics regarding functional traits, the relationship between traits and phylogeny, the effect and response of functional traits to ecosystem functions and disturbance. In addition, we also provide a future outlook, including research directions, methods and applications based on functional traits in plant science. Specially, we propose that functional traits can also be used as indicators or reference for biodiversity conservation and management of ecosystems under the current global climate change. China has a huge territory, which brings great natural recourses of geography and plants, providing a huge natural database for research on functional traits.
DOIURL [本文引用: 2]
Plant functional traits are plant core attributes closely related to plant colonization, survival, growth and mortality. These core attributes could significantly affect ecosystem functions and reflect the response of vegetation to environmental change. There is growing recognition that many ecological questions at various levels can be addressed effectively with key functional traits. In this paper, we review key research progress over the last 10 years addressing important ecological issues based on functional traits, including global distribution patterns of functional traits and their associations, variation of traits distribution along environmental gradients, functional diversity and its application, mechanisms of community assembly and dynamics regarding functional traits, the relationship between traits and phylogeny, the effect and response of functional traits to ecosystem functions and disturbance. In addition, we also provide a future outlook, including research directions, methods and applications based on functional traits in plant science. Specially, we propose that functional traits can also be used as indicators or reference for biodiversity conservation and management of ecosystems under the current global climate change. China has a huge territory, which brings great natural recourses of geography and plants, providing a huge natural database for research on functional traits.
DOIURL [本文引用: 1]
http://www.jstor.org/stable/3599027
DOIURL [本文引用: 1]
植物性状反映了植物对生长环境的响应和适应,将环境、植物个体和生态系统结构、过程与功能联系起来(所谓的“植物功能性状”)。该文介绍了植物功能性状的分类体系,综述了国内外植物功能性状与气候(包括气温、降水、光照)、地理空间变异(包括地形地貌、生态梯度、海拔)、营养、干扰(包括火灾、放牧、生物入侵、土地利用)等环境因素,以及与生态系统功能之间关系的研究进展,探讨了全球变化(气候变化和CO2浓度升高)对个体和群落植物功能性状的影响。植物功能性状的研究已经取得很多成果,并应用于全球变化、古植被恢复和古气候定量重建、环境监测与评价、生态保护和恢复等研究中,但大尺度、多生境因子下的植物功能性状研究仍有待于加强,同时需要改进性状的测量手段;我国的植物功能性状研究还需要更加明朗化和系统化。
DOIURL [本文引用: 1]
植物性状反映了植物对生长环境的响应和适应,将环境、植物个体和生态系统结构、过程与功能联系起来(所谓的“植物功能性状”)。该文介绍了植物功能性状的分类体系,综述了国内外植物功能性状与气候(包括气温、降水、光照)、地理空间变异(包括地形地貌、生态梯度、海拔)、营养、干扰(包括火灾、放牧、生物入侵、土地利用)等环境因素,以及与生态系统功能之间关系的研究进展,探讨了全球变化(气候变化和CO2浓度升高)对个体和群落植物功能性状的影响。植物功能性状的研究已经取得很多成果,并应用于全球变化、古植被恢复和古气候定量重建、环境监测与评价、生态保护和恢复等研究中,但大尺度、多生境因子下的植物功能性状研究仍有待于加强,同时需要改进性状的测量手段;我国的植物功能性状研究还需要更加明朗化和系统化。
DOIURL [本文引用: 2]
Abstract 1. Plant phenotypic diversity is shaped by the interplay of trade-offs and constraints in evolution. Closely integrated groups of traits (i.e. trait dimensions) are used to classify plant phenotypic diversity into plant strategies, but we do not know the degree of interdependence among trait dimensions. To assess how selection has shaped the phenotypic space, we examine whether trait dimensions are independent. 2. We gathered data on saplings of 24 locally coexisting tree species in a temperate forest, and examined the correlation structure of 20 leaf, branch, stem and root traits. These traits fall into three well-established trait dimensions (the leaf economic spectrum, the wood spectrum, and Corner Rules) that characterize vital plant functions: resource acquisition, sap transport, mechanical support and canopy architecture. Using ordinations, network analyses and Mantel tests, we tested whether the sapling phenotype of these tree species is organized along independent trait dimensions. 3. Across species, the sapling phenotype is not structured into clear trait dimensions. The trait relationships defining the trait dimensions are weak and do not dominate the correlation structure of the sapling phenotype as a whole. Instead traits from the three commonly recognized trait dimensions are organized into an integrated trait network. The effect of phylogeny on trait correlations is minimal. 4. Our results indicate that trait dimensions apparent in broad-based interspecific surveys do not hold up among locally coexisting species. Furthermore, architectural traits appear central to the phenotypic network, suggesting a pivotal role for branching architecture in linking resource acquisition, mechanical support and hydraulic functions. 5. Synthesis. Our study indicates that local and global patterns of phenotypic integration differ and calls into question the use of trait dimensions at local scales. We propose that a network approach to assessing plant function more effectively reflects the multiple trade-offs and constraints shaping the phenotype in locally co-occurring species.
DOIURLPMID [本文引用: 1]
61Paleobotanists have long used models based on leaf size and shape to reconstruct paleoclimate. However, most models incorporate a single variable or use traits that are not physiologically or functionally linked to climate, limiting their predictive power. Further, they often underestimate paleotemperature relative to other proxies.61Here we quantify leaf–climate correlations from 92 globally distributed, climatically diverse sites, and explore potential confounding factors. Multiple linear regression models for mean annual temperature (MAT) and mean annual precipitation (MAP) are developed and applied to nine well-studied fossil floras.61We find that leaves in cold climates typically have larger, more numerous teeth, and are more highly dissected. Leaf habit (deciduous vs evergreen), local water availability, and phylogenetic history all affect these relationships. Leaves in wet climates are larger and have fewer, smaller teeth. Our multivariate MAT and MAP models offer moderate improvements in precision over univariate approaches (± 4.0 vs 4.8°C for MAT) and strong improvements in accuracy. For example, our provisional MAT estimates for most North American fossil floras are considerably warmer and in better agreement with independent paleoclimate evidence.61Our study demonstrates that the inclusion of additional leaf traits that are functionally linked to climate improves paleoclimate reconstructions. This work also illustrates the need for better understanding of the impact of phylogeny and leaf habit on leaf–climate relationships.
DOIURL [本文引用: 2]
Plant functional traits are the features (morphological, physiological, phenological) that represent ecological strategies and determine how plants respond to environmental factors, affect other trophic levels and influence ecosystem properties. Variation in plant functional traits, and trait syndromes, has proven useful for tackling many important ecological questions at a range of scales, giving rise to a demand for standardised ways to measure ecologically meaningful plant traits. This line of research has been among the most fruitful avenues for understanding ecological and evolutionary patterns and processes. It also has the potential both to build a predictive set of local, regional and global relationships between plants and environment and to quantify a wide range of natural and human-driven processes, including changes in biodiversity, the impacts of species invasions, alterations in biogeochemical processes and vegetation-atmosphere interactions. The importance of these topics dictates the urgent need for more and better data, and increases the value of standardised protocols for quantifying trait variation of different species, in particular for traits with power to predict plant-and ecosystem-level processes, and for traits that can be measured relatively easily. Updated and expanded from the widely used previous version, this handbook retains the focus on clearly presented, widely applicable, step-by-step recipes, with a minimum of text on theory, and not only includes updated methods for the traits previously covered, but also introduces many new protocols for further traits. This new handbook has a better balance between whole-plant traits, leaf traits, root and stem traits and regenerative traits, and puts particular emphasis on traits important for predicting species' effects on key ecosystem properties. We hope this new handbook becomes a standard companion in local and global efforts to learn about the responses and impacts of different plant species with respect to environmental changes in the present, past and future.
[本文引用: 1]
DOIURLPMID [本文引用: 2]
Abstract Recent studies have shown that accounting for intraspecific trait variation (ITV) may better address major questions in community ecology. However, a general picture of the relative extent of ITV compared to interspecific trait variation in plant communities is still missing. Here, we conducted a meta-analysis of the relative extent of ITV within and among plant communities worldwide, using a data set encompassing 629 communities (plots) and 36 functional traits. Overall, ITV accounted for 25% of the total trait variation within communities and 32% of the total trait variation among communities on average. The relative extent of ITV tended to be greater for whole-plant (e.g. plant height) vs. organ-level traits and for leaf chemical (e.g. leaf N and P concentration) vs. leaf morphological (e.g. leaf area and thickness) traits. The relative amount of ITV decreased with increasing species richness and spatial extent, but did not vary with plant growth form or climate. These results highlight global patterns in the relative importance of ITV in plant communities, providing practical guidelines for when researchers should include ITV in trait-based community and ecosystem studies.
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
叶片性状反映了植物对环境的高度适应能力及其在复杂生境下的自我调控能力.叶片性状如何响应和适应气候变 化是植物适应性研究的重点内容.该文系统综述了叶片大小、比叶质量、叶片氮含量、碳同位素等指标对气候变化响应的最新研究结果.不同叶片性状对气候变化的 响应结果存在差异,所指示的生态学含义也有所不同.单一叶片性状不能全面地反映植物对气候变化的响应;不同尺度的研究(如环境的修饰或筛选作用的研究)还 存在很多不确定性.高寒地区的研究工作相对缺乏.该文有助于理解植物与气候之间的相互关系、植物对气候变化的响应与适应对策,对了解植物演化、预测植物在 未来气候变化条件下的变化特征具有一定意义.
DOIURL [本文引用: 1]
叶片性状反映了植物对环境的高度适应能力及其在复杂生境下的自我调控能力.叶片性状如何响应和适应气候变 化是植物适应性研究的重点内容.该文系统综述了叶片大小、比叶质量、叶片氮含量、碳同位素等指标对气候变化响应的最新研究结果.不同叶片性状对气候变化的 响应结果存在差异,所指示的生态学含义也有所不同.单一叶片性状不能全面地反映植物对气候变化的响应;不同尺度的研究(如环境的修饰或筛选作用的研究)还 存在很多不确定性.高寒地区的研究工作相对缺乏.该文有助于理解植物与气候之间的相互关系、植物对气候变化的响应与适应对策,对了解植物演化、预测植物在 未来气候变化条件下的变化特征具有一定意义.
DOIURL [本文引用: 2]
An important aim of plant ecology is to identify leading dimensions of ecological variation among species and to understand the basis for them. Dimensions that can readily be measured would be especially useful, because they might offer a path towards improved worldwide synthesis across the thousands of field experiments and ecophysiological studies that use just a few species each. Four dimensions are reviewed here. The leaf mass per area-leaf lifespan (LMA-LL) dimension expresses slow turnover of plant parts (at high LMA and long LL), long nutrient residence times, and slow response to favorable growth conditions. The seed mass-seed output (SM-SO) dimension is an important predictor of dispersal to establishment opportunities (seed output) and of establishment success in the face of hazards (seed mass). The LMA-LL and SM-SO dimensions are each underpinned by a single, comprehensible tradeoff, and their consequences are fairly well understood. The leaf size-twig size (LS-TS) spectrum has obvious consequences for the texture of canopies, but the costs and benefits of large versus small leaf and twig size are poorly understood. The height dimension has universally been seen as ecologically important and included in ecological strategy schemes. Nevertheless, height includes several tradeoffs and adaptive elements, which ideally should be treated separately. Each of these four dimensions varies at the scales of climate zones and of site types within landscapes. This variation can be interpreted as adaptation to the physical environment. Each dimension also varies widely among coexisting species. Most likely this within-site variation arises because the ecological opportunities for each species depend strongly on which other species are present, in other words, because the set of species at a site is a stable mixture of strategies.
DOIURL [本文引用: 2]
The advantages of being evergreen
1
1995
... 因此, 在测定植物叶片大小时, 按照传统和约定俗成地采摘成熟植株的健康叶片10-20片(
Intraspecific functional variability: Extent, structure and sources of variation
2
2010
... 在野外测定叶面积时, 针对一个样地中的目标植物, 除了极其特殊的情形, 如无叶或鳞叶植物、针叶植物等, 通常是每种植物选择10株成熟个体, 随机采摘相对幼嫩但伸展、硬化且无病虫害、无附生植物的健康叶片20个; 至少保证每个物种采集10个叶片, 在叶片稀少时最起码也需要5个植株, 每个植株采摘2个叶片, 或者在成熟个体稀少时最少有4个植株、每个植株采摘5个叶片; 保湿、冷藏储存, 并尽快利用可自动测量的叶面积仪(如LI-COR或Delta-T)或者人工测定的扫描仪进行叶面积测定(
... 植物的分类地位与环境条件决定其性状特征, 包括叶面积.植物叶面积存在较大的种间变异(
Inter-specific and intra-specific trait variation along short environmental gradients in an old-growth temperate forest
2
2013
... 在野外测定叶面积时, 针对一个样地中的目标植物, 除了极其特殊的情形, 如无叶或鳞叶植物、针叶植物等, 通常是每种植物选择10株成熟个体, 随机采摘相对幼嫩但伸展、硬化且无病虫害、无附生植物的健康叶片20个; 至少保证每个物种采集10个叶片, 在叶片稀少时最起码也需要5个植株, 每个植株采摘2个叶片, 或者在成熟个体稀少时最少有4个植株、每个植株采摘5个叶片; 保湿、冷藏储存, 并尽快利用可自动测量的叶面积仪(如LI-COR或Delta-T)或者人工测定的扫描仪进行叶面积测定(
... 植物的分类地位与环境条件决定其性状特征, 包括叶面积.植物叶面积存在较大的种间变异(
Why intraspecific trait variation matters in community ecology
2
2011
... 在野外测定叶面积时, 针对一个样地中的目标植物, 除了极其特殊的情形, 如无叶或鳞叶植物、针叶植物等, 通常是每种植物选择10株成熟个体, 随机采摘相对幼嫩但伸展、硬化且无病虫害、无附生植物的健康叶片20个; 至少保证每个物种采集10个叶片, 在叶片稀少时最起码也需要5个植株, 每个植株采摘2个叶片, 或者在成熟个体稀少时最少有4个植株、每个植株采摘5个叶片; 保湿、冷藏储存, 并尽快利用可自动测量的叶面积仪(如LI-COR或Delta-T)或者人工测定的扫描仪进行叶面积测定(
... 植物的分类地位与环境条件决定其性状特征, 包括叶面积.植物叶面积存在较大的种间变异(
Interspecific and intraspecific variation in specific root length drives aboveground biodiversity effects in young experimental forest stands
1
2017
... 植物的分类地位与环境条件决定其性状特征, 包括叶面积.植物叶面积存在较大的种间变异(
植物叶经济谱的研究进展
1
2014
... 叶片是植物光合作用和物质生产的主要器官, 指示叶片大小的性状, 如叶面积(叶片单面投影表面积, mm2), 可反映植物对环境的响应与适应, 指示生态系统的功能与过程, 与环境胁迫、干扰和植物的生长对策及其利用资源的能力密切相关, 是重要的植物功能性状之一(
植物叶经济谱的研究进展
1
2014
... 叶片是植物光合作用和物质生产的主要器官, 指示叶片大小的性状, 如叶面积(叶片单面投影表面积, mm2), 可反映植物对环境的响应与适应, 指示生态系统的功能与过程, 与环境胁迫、干扰和植物的生长对策及其利用资源的能力密切相关, 是重要的植物功能性状之一(
A handbook of protocols for standardized and easy measurement of plant functional traits worldwide
2
2003
... 在野外测定叶面积时, 针对一个样地中的目标植物, 除了极其特殊的情形, 如无叶或鳞叶植物、针叶植物等, 通常是每种植物选择10株成熟个体, 随机采摘相对幼嫩但伸展、硬化且无病虫害、无附生植物的健康叶片20个; 至少保证每个物种采集10个叶片, 在叶片稀少时最起码也需要5个植株, 每个植株采摘2个叶片, 或者在成熟个体稀少时最少有4个植株、每个植株采摘5个叶片; 保湿、冷藏储存, 并尽快利用可自动测量的叶面积仪(如LI-COR或Delta-T)或者人工测定的扫描仪进行叶面积测定(
... 因此, 在测定植物叶片大小时, 按照传统和约定俗成地采摘成熟植株的健康叶片10-20片(
The global spectrum of plant form and function
3
2016
... 叶片是植物光合作用和物质生产的主要器官, 指示叶片大小的性状, 如叶面积(叶片单面投影表面积, mm2), 可反映植物对环境的响应与适应, 指示生态系统的功能与过程, 与环境胁迫、干扰和植物的生长对策及其利用资源的能力密切相关, 是重要的植物功能性状之一(
... ;
... ;
The evolution of the worldwide leaf economics spectrum
1
2011
... 叶片是植物光合作用和物质生产的主要器官, 指示叶片大小的性状, 如叶面积(叶片单面投影表面积, mm2), 可反映植物对环境的响应与适应, 指示生态系统的功能与过程, 与环境胁迫、干扰和植物的生长对策及其利用资源的能力密切相关, 是重要的植物功能性状之一(
浙江金华北山木本植物群落的DCA排序
1
2004
... 金华山地处中亚热带北缘, 位于浙江省中西部金华市北15 km处, 金衢盆地东部, 系龙门山脉的支脉, 因横亘于金华市北, 故俗称北山.整个山体跨越119.48°-119.82° E, 29.15°-29.27° N, 海拔通常在100-1 200 m, 主峰大盘尖海拔1 314 m.研究区位于金华山南坡双龙洞国家森林公园内, 属亚热带山地季风气候.据金华气象站(119.65° E, 29.12° N, 海拔626 m) 1953-2014年的纪录, 该地区年平均气温为17.7 ℃, 1月平均气温5.2 ℃, 7月平均气温29.5 ℃, 极端低温达-9.6 ℃, 极端高温为41.5 ℃; 年降水量可达1 436.8 mm, 主要集中在3-9月, 占全年降水量的76.7%, 但以6月份梅雨季节的降水量最大, 平均达252.2 mm, 3-6月平温期占降水集中期降水量的70% (平均月降水量191 mm), 而7-9月高温期降水反而不是最高(平均月降水量112 mm); 年日照率为43%, 无霜期256天.该地区土壤以亚热带山地黄壤为主, 植被具有亚热带常绿阔叶林、常绿落叶阔叶林和灌丛等垂直带谱, 但受人为干扰影响大, 是人为干扰下典型的亚热带常绿阔叶林退化植被, 山脚至中坡(200-700 m)以次生性的常绿阔叶林、常绿落叶阔叶混交林和针阔混交林、次生灌丛为主, 中坡至上坡(600-1000 m)主要为马尾松(Pinus massoniana)人工林、杉木(Cunninghamia lanceolata)林、柳杉(Cryptomeria fortunei)林、黄山松(Pinus taiwanensis)林和山地灌丛(
浙江金华北山木本植物群落的DCA排序
1
2004
... 金华山地处中亚热带北缘, 位于浙江省中西部金华市北15 km处, 金衢盆地东部, 系龙门山脉的支脉, 因横亘于金华市北, 故俗称北山.整个山体跨越119.48°-119.82° E, 29.15°-29.27° N, 海拔通常在100-1 200 m, 主峰大盘尖海拔1 314 m.研究区位于金华山南坡双龙洞国家森林公园内, 属亚热带山地季风气候.据金华气象站(119.65° E, 29.12° N, 海拔626 m) 1953-2014年的纪录, 该地区年平均气温为17.7 ℃, 1月平均气温5.2 ℃, 7月平均气温29.5 ℃, 极端低温达-9.6 ℃, 极端高温为41.5 ℃; 年降水量可达1 436.8 mm, 主要集中在3-9月, 占全年降水量的76.7%, 但以6月份梅雨季节的降水量最大, 平均达252.2 mm, 3-6月平温期占降水集中期降水量的70% (平均月降水量191 mm), 而7-9月高温期降水反而不是最高(平均月降水量112 mm); 年日照率为43%, 无霜期256天.该地区土壤以亚热带山地黄壤为主, 植被具有亚热带常绿阔叶林、常绿落叶阔叶林和灌丛等垂直带谱, 但受人为干扰影响大, 是人为干扰下典型的亚热带常绿阔叶林退化植被, 山脚至中坡(200-700 m)以次生性的常绿阔叶林、常绿落叶阔叶混交林和针阔混交林、次生灌丛为主, 中坡至上坡(600-1000 m)主要为马尾松(Pinus massoniana)人工林、杉木(Cunninghamia lanceolata)林、柳杉(Cryptomeria fortunei)林、黄山松(Pinus taiwanensis)林和山地灌丛(
Adaptive significance of evergreen vs. deciduous leaves: Solving the triple paradox
1
2002
... 因此, 在测定植物叶片大小时, 按照传统和约定俗成地采摘成熟植株的健康叶片10-20片(
浙江金华北山植物区系及植被
1
1993
... 金华山地处中亚热带北缘, 位于浙江省中西部金华市北15 km处, 金衢盆地东部, 系龙门山脉的支脉, 因横亘于金华市北, 故俗称北山.整个山体跨越119.48°-119.82° E, 29.15°-29.27° N, 海拔通常在100-1 200 m, 主峰大盘尖海拔1 314 m.研究区位于金华山南坡双龙洞国家森林公园内, 属亚热带山地季风气候.据金华气象站(119.65° E, 29.12° N, 海拔626 m) 1953-2014年的纪录, 该地区年平均气温为17.7 ℃, 1月平均气温5.2 ℃, 7月平均气温29.5 ℃, 极端低温达-9.6 ℃, 极端高温为41.5 ℃; 年降水量可达1 436.8 mm, 主要集中在3-9月, 占全年降水量的76.7%, 但以6月份梅雨季节的降水量最大, 平均达252.2 mm, 3-6月平温期占降水集中期降水量的70% (平均月降水量191 mm), 而7-9月高温期降水反而不是最高(平均月降水量112 mm); 年日照率为43%, 无霜期256天.该地区土壤以亚热带山地黄壤为主, 植被具有亚热带常绿阔叶林、常绿落叶阔叶林和灌丛等垂直带谱, 但受人为干扰影响大, 是人为干扰下典型的亚热带常绿阔叶林退化植被, 山脚至中坡(200-700 m)以次生性的常绿阔叶林、常绿落叶阔叶混交林和针阔混交林、次生灌丛为主, 中坡至上坡(600-1000 m)主要为马尾松(Pinus massoniana)人工林、杉木(Cunninghamia lanceolata)林、柳杉(Cryptomeria fortunei)林、黄山松(Pinus taiwanensis)林和山地灌丛(
浙江金华北山植物区系及植被
1
1993
... 金华山地处中亚热带北缘, 位于浙江省中西部金华市北15 km处, 金衢盆地东部, 系龙门山脉的支脉, 因横亘于金华市北, 故俗称北山.整个山体跨越119.48°-119.82° E, 29.15°-29.27° N, 海拔通常在100-1 200 m, 主峰大盘尖海拔1 314 m.研究区位于金华山南坡双龙洞国家森林公园内, 属亚热带山地季风气候.据金华气象站(119.65° E, 29.12° N, 海拔626 m) 1953-2014年的纪录, 该地区年平均气温为17.7 ℃, 1月平均气温5.2 ℃, 7月平均气温29.5 ℃, 极端低温达-9.6 ℃, 极端高温为41.5 ℃; 年降水量可达1 436.8 mm, 主要集中在3-9月, 占全年降水量的76.7%, 但以6月份梅雨季节的降水量最大, 平均达252.2 mm, 3-6月平温期占降水集中期降水量的70% (平均月降水量191 mm), 而7-9月高温期降水反而不是最高(平均月降水量112 mm); 年日照率为43%, 无霜期256天.该地区土壤以亚热带山地黄壤为主, 植被具有亚热带常绿阔叶林、常绿落叶阔叶林和灌丛等垂直带谱, 但受人为干扰影响大, 是人为干扰下典型的亚热带常绿阔叶林退化植被, 山脚至中坡(200-700 m)以次生性的常绿阔叶林、常绿落叶阔叶混交林和针阔混交林、次生灌丛为主, 中坡至上坡(600-1000 m)主要为马尾松(Pinus massoniana)人工林、杉木(Cunninghamia lanceolata)林、柳杉(Cryptomeria fortunei)林、黄山松(Pinus taiwanensis)林和山地灌丛(
东北次生林主要树种比叶面积对光照强度的响应
1
2009
... 另外, 传统的方法对采样植株的年龄与采样方位没有具体规定, 通常在野外操作时都是针对大树向阳的叶片.本研究通过对亚热带常绿落叶阔叶混交林的一个常绿与一个落叶优势树种的研究发现, 中龄植株树冠底部是较合适的叶片采样对象与方位.树龄影响叶片大小, 在种间表现出一定的差异, 木荷叶片大小与树龄关系不密切, 而枫香树则较密切, 二者中龄树的叶片大小与总叶片大小的关系一致.而取样方位, 亦即光照强度同样影响叶片大小(
东北次生林主要树种比叶面积对光照强度的响应
1
2009
... 另外, 传统的方法对采样植株的年龄与采样方位没有具体规定, 通常在野外操作时都是针对大树向阳的叶片.本研究通过对亚热带常绿落叶阔叶混交林的一个常绿与一个落叶优势树种的研究发现, 中龄植株树冠底部是较合适的叶片采样对象与方位.树龄影响叶片大小, 在种间表现出一定的差异, 木荷叶片大小与树龄关系不密切, 而枫香树则较密切, 二者中龄树的叶片大小与总叶片大小的关系一致.而取样方位, 亦即光照强度同样影响叶片大小(
植物功能性状研究进展
2
2015
... 叶片是植物光合作用和物质生产的主要器官, 指示叶片大小的性状, 如叶面积(叶片单面投影表面积, mm2), 可反映植物对环境的响应与适应, 指示生态系统的功能与过程, 与环境胁迫、干扰和植物的生长对策及其利用资源的能力密切相关, 是重要的植物功能性状之一(
... ), 综合反映植物功能性状与环境和生态系统功能的关系, 对模拟和预测气候变化对生态系统的影响, 指导生物多样性保护和生态系统管理政策的制定等具有重要的生态学意义(
植物功能性状研究进展
2
2015
... 叶片是植物光合作用和物质生产的主要器官, 指示叶片大小的性状, 如叶面积(叶片单面投影表面积, mm2), 可反映植物对环境的响应与适应, 指示生态系统的功能与过程, 与环境胁迫、干扰和植物的生长对策及其利用资源的能力密切相关, 是重要的植物功能性状之一(
... ), 综合反映植物功能性状与环境和生态系统功能的关系, 对模拟和预测气候变化对生态系统的影响, 指导生物多样性保护和生态系统管理政策的制定等具有重要的生态学意义(
Leaf-size divergence along rainfall and soil-nutrient gradients: Is the method of size reduction common among clades?
1
2003
... 植物的分类地位与环境条件决定其性状特征, 包括叶面积.植物叶面积存在较大的种间变异(
植物功能性状与环境和生态系统功能
1
2007
... 叶片是植物光合作用和物质生产的主要器官, 指示叶片大小的性状, 如叶面积(叶片单面投影表面积, mm2), 可反映植物对环境的响应与适应, 指示生态系统的功能与过程, 与环境胁迫、干扰和植物的生长对策及其利用资源的能力密切相关, 是重要的植物功能性状之一(
植物功能性状与环境和生态系统功能
1
2007
... 叶片是植物光合作用和物质生产的主要器官, 指示叶片大小的性状, 如叶面积(叶片单面投影表面积, mm2), 可反映植物对环境的响应与适应, 指示生态系统的功能与过程, 与环境胁迫、干扰和植物的生长对策及其利用资源的能力密切相关, 是重要的植物功能性状之一(
Interspecific integration of trait dimensions at local scales: The plant phenotype as an integrated network
2
2017
... 在野外测定叶面积时, 针对一个样地中的目标植物, 除了极其特殊的情形, 如无叶或鳞叶植物、针叶植物等, 通常是每种植物选择10株成熟个体, 随机采摘相对幼嫩但伸展、硬化且无病虫害、无附生植物的健康叶片20个; 至少保证每个物种采集10个叶片, 在叶片稀少时最起码也需要5个植株, 每个植株采摘2个叶片, 或者在成熟个体稀少时最少有4个植株、每个植株采摘5个叶片; 保湿、冷藏储存, 并尽快利用可自动测量的叶面积仪(如LI-COR或Delta-T)或者人工测定的扫描仪进行叶面积测定(
... 植物的分类地位与环境条件决定其性状特征, 包括叶面积.植物叶面积存在较大的种间变异(
Sensitivity of leaf size and shape to climate: Global patterns and paleoclimatic applications
1
2011
... 植物的分类地位与环境条件决定其性状特征, 包括叶面积.植物叶面积存在较大的种间变异(
New handbook for standardised measurement of plant functional traits worldwide
2
2013
... 在野外测定叶面积时, 针对一个样地中的目标植物, 除了极其特殊的情形, 如无叶或鳞叶植物、针叶植物等, 通常是每种植物选择10株成熟个体, 随机采摘相对幼嫩但伸展、硬化且无病虫害、无附生植物的健康叶片20个; 至少保证每个物种采集10个叶片, 在叶片稀少时最起码也需要5个植株, 每个植株采摘2个叶片, 或者在成熟个体稀少时最少有4个植株、每个植株采摘5个叶片; 保湿、冷藏储存, 并尽快利用可自动测量的叶面积仪(如LI-COR或Delta-T)或者人工测定的扫描仪进行叶面积测定(
... 因此, 在测定植物叶片大小时, 按照传统和约定俗成地采摘成熟植株的健康叶片10-20片(
Sensitivity of leaf size and shape to climate within Acer rubrum and Quercus kelloggii
1
2008
... 植物的分类地位与环境条件决定其性状特征, 包括叶面积.植物叶面积存在较大的种间变异(
A global meta-analysis of the relative extent of intraspecific trait variation in plant communities
2
2015
... 在野外测定叶面积时, 针对一个样地中的目标植物, 除了极其特殊的情形, 如无叶或鳞叶植物、针叶植物等, 通常是每种植物选择10株成熟个体, 随机采摘相对幼嫩但伸展、硬化且无病虫害、无附生植物的健康叶片20个; 至少保证每个物种采集10个叶片, 在叶片稀少时最起码也需要5个植株, 每个植株采摘2个叶片, 或者在成熟个体稀少时最少有4个植株、每个植株采摘5个叶片; 保湿、冷藏储存, 并尽快利用可自动测量的叶面积仪(如LI-COR或Delta-T)或者人工测定的扫描仪进行叶面积测定(
... 植物的分类地位与环境条件决定其性状特征, 包括叶面积.植物叶面积存在较大的种间变异(
The return of the variance: Intraspecific variability in community ecology
1
2012
... 植物的分类地位与环境条件决定其性状特征, 包括叶面积.植物叶面积存在较大的种间变异(
植物叶片性状对气候变化的响应研究进展
1
2015
... 叶片是植物光合作用和物质生产的主要器官, 指示叶片大小的性状, 如叶面积(叶片单面投影表面积, mm2), 可反映植物对环境的响应与适应, 指示生态系统的功能与过程, 与环境胁迫、干扰和植物的生长对策及其利用资源的能力密切相关, 是重要的植物功能性状之一(
植物叶片性状对气候变化的响应研究进展
1
2015
... 叶片是植物光合作用和物质生产的主要器官, 指示叶片大小的性状, 如叶面积(叶片单面投影表面积, mm2), 可反映植物对环境的响应与适应, 指示生态系统的功能与过程, 与环境胁迫、干扰和植物的生长对策及其利用资源的能力密切相关, 是重要的植物功能性状之一(
Plant ecological strategies: Some leading dimensions of variation between species
2
2002
... 在野外测定叶面积时, 针对一个样地中的目标植物, 除了极其特殊的情形, 如无叶或鳞叶植物、针叶植物等, 通常是每种植物选择10株成熟个体, 随机采摘相对幼嫩但伸展、硬化且无病虫害、无附生植物的健康叶片20个; 至少保证每个物种采集10个叶片, 在叶片稀少时最起码也需要5个植株, 每个植株采摘2个叶片, 或者在成熟个体稀少时最少有4个植株、每个植株采摘5个叶片; 保湿、冷藏储存, 并尽快利用可自动测量的叶面积仪(如LI-COR或Delta-T)或者人工测定的扫描仪进行叶面积测定(
... 植物的分类地位与环境条件决定其性状特征, 包括叶面积.植物叶面积存在较大的种间变异(
The worldwide leaf economics spectrum
2
2004
... 叶片是植物光合作用和物质生产的主要器官, 指示叶片大小的性状, 如叶面积(叶片单面投影表面积, mm2), 可反映植物对环境的响应与适应, 指示生态系统的功能与过程, 与环境胁迫、干扰和植物的生长对策及其利用资源的能力密切相关, 是重要的植物功能性状之一(
... ).相对于茎干与根系性状, 叶片易于采样, 性状易于测定, 可以通过大批量的采样来比较和分析不同地区、不同生态系统和不同植物种类的功能性状特征.同时, 叶面积性状还可拓展用以计算比叶面积、基于面积的叶片营养元素含量等, 由此与其他叶性状相互关联所形成的叶经济谱(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}