 ,2,*, 胡良2, 孟庆银3
,2,*, 胡良2, 孟庆银3Responses of nitrogen and phosphorus resorption from leaves and branches to long-term nitrogen deposition in a Chinese fir plantation
SHEN Fang-Fang1,2, LI Yan-Yan1,2, LIU Wen-Fei2, DUAN Hong-Lang2, FAN Hou-Bao,2,*, HU Liang2, MENG Qing-Yin3通讯作者: hbfan@nit.edu.cn
编委: 孙建新
责任编辑: 李 敏
收稿日期:2018-07-16修回日期:2018-09-10网络出版日期:2018-09-20
| 基金资助: |
Received:2018-07-16Revised:2018-09-10Online:2018-09-20
| Fund supported: |

摘要
为了解森林养分内循环对全球变化的响应, 基于长期模拟氮沉降试验, 研究了杉木(Cunninghamia lanceolata)人工林不同龄级(一年生、二年生和衰老)叶和枝的氮(N)、磷(P)养分分配及其再吸收特征, 并分析了不同模拟N沉降处理时间(7年和14年)杉木叶N、P养分再吸收差异。在12年生杉木中开展模拟N沉降试验, 以尿素(CO(NH2)2)为N源, 设N0、N1、N2和N3 4个处理水平, 施氮量分别为0、60、120和240 kg·hm -2·a -1, 每个处理重复3次。结果表明: (1)叶和枝在衰老过程中碳(C)、N和P含量逐渐降低, 且叶的C、N和P含量比枝高; N含量大小依次为一年生叶>二年生叶>衰老叶>一年生枝>二年生枝>衰老枝, 且N3 > N2 > N1 > N0, 而C:N则呈现相反的趋势; 衰老器官的C:N、C:P、N:P比新鲜器官高; N沉降增加了不同龄级叶和枝(除二年生叶外)的N、N:P和C:P, 但降低了P和C:N。(2)叶和枝的N、P养分再吸收率(REN、REP)随龄级的增加至衰老有规律地递减, 且REP > REN; 受长期N沉降的影响, REN叶(28.12%) <枝(30.00%), 而REP则为叶(45.82%) >枝(30.42%); 杉木叶和枝N:P与REN:REP之间存在极显著的线性相关关系。(3)随N沉降处理时间的增加, 叶REN呈降低态势, 各处理(N1、N2和N3)分别降低了9.85%、3.17%和11.71%; 而REP则明显上升, 分别增加了71.98%、42.25%和9.60%。研究结果表明: 不同器官、不同龄级的养分再吸收率随氮沉降处理的水平、处理时间而所有不同; REN:REP与N:P之间存在紧密关系。
关键词:
Abstract
Keywords:
PDF (1317KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
沈芳芳, 李燕燕, 刘文飞, 段洪浪, 樊后保, 胡良, 孟庆银. 长期氮沉降对杉木人工林叶、枝氮磷养分再吸收的影响. 植物生态学报[J], 2018, 42(9): 926-937 DOI:10.17521/cjpe.2018.0167
SHEN Fang-Fang, LI Yan-Yan, LIU Wen-Fei, DUAN Hong-Lang, FAN Hou-Bao, HU Liang, MENG Qing-Yin.
养分再吸收(NR)使植物组织中的养分元素(氮(N)、磷(P))在随凋落物进入土壤之前被重新利用, 它缩短了养分元素在植物与环境之间的循环距离(Zhang et al., 2014), 提高了养分利用效率, 减少了养分的损失, 增强了植物对养分贫乏的环境或生态系统的适应性(Reed et al., 2012; 陆姣云等, 2018)。Reed等(2012)采用meta分析得出全球植物N再吸收率(REN)和P再吸收率(REP)平均约为49%和50%, 与Aerts (1996)分析得出REN和REP为50%和52%接近, 而Vergutz等(2012)研究表明, 全球植物REN和REP平均约为62.1%和64.9%, 再吸收的N、P参与到植物生物量、基底面积的生长、树干的生长、果实的生产等过程中。
NR受植物的生活型、器官、年龄等生理因素的影响。大量研究表明, 植物的不同器官, 如叶(Huang et al., 2018)、枝(费世民, 2001)、树干(费世民, 2001; Brant & Chen, 2015)、树皮(Brant & Chen, 2015)、细根(Brant & Chen, 2015; 陈珊等, 2015)都会发生体内养分转移、运输和储存, 其中以叶的贡献最大(Drenovsky & Richards, 2006)。在全球范围内研究叶较多, 研究枝较少。不同生活型植物的NR不同。如, 养分再吸收率呈现木本植物低于非木本植物(江大龙等, 2017); 落叶植物高于常绿植物(Brant & Chen, 2015; 江大龙等, 2017); 常绿植物的REN比落叶植物低, 但REP比落叶植物高, 木本植物REN低于灌木, 针叶树与阔叶树的REN和REP没有差异(Yuan & Chen, 2008)。Yuan等(2005)研究得出中国半干旱地区的不同生活型植物的REN大小依次为草本>灌木>树木>禾本科>固氮植物。不同林龄植物养分再吸收率也不同, 幼龄林有更高的REN, 老龄林具有更高的REP (江大龙等, 2017)。NR在维持植物化学计量比平衡方面起到关键作用, 同时REN:REP的变化为各种空间尺度养分循环和限制的认识提供了一个新的角度(Reed et al., 2012; Yan et al., 2014)。
大气氮沉降在全球范围内普遍发生, 全球总活性N释放量从35 Tg·a-1 (1850年)增加到104 Tg·a-1 (1997?2013年), 在未来一段时期可能继续增加(Galloway et al., 2008), 预计2100年达到114 Tg·a-1 (Wang et al., 2017)。1980-2010年, 中国地区氮沉降量平均每年增加0.41 kg·hm-2, 氮沉降速率与1980年欧洲西北部的沉降高峰值一致(Liu et al., 2013)。中国森林正在经历严重的氮沉降, 过去的20年里, 氮沉降的平均速度从13.2 kg·hm-2增加到21.1 kg·hm-2 (Liu et al., 2013), 并且在很大程度上影响了生态系统的N和P循环(Deng et al., 2017; Wu et al., 2017)。研究表明, 氮沉降降低了受N限制的生态系统的植物叶片N:P, 增加了REN:REP; 但增加了受P限制的生态系统中的叶片N:P, 降低了REN:REP (Zechmeister- Boltenstern et al., 2015)。氮沉降影响下, 不同物种的REN有3种效应: 没有显著影响(Huang et al., 2008; Wang et al., 2014; Chen et al., 2015; 王斌等, 2016)、增加(Yan et al., 2014)、降低(Huang et al., 2008; Li et al., 2010; 赵琼等, 2010); REP也有3种效应: 无反应(Wang et al., 2014; Yan et al., 2014; Chen et al., 2015; 王斌等, 2016)、增加(Wang et al., 2014)、降低(Yan et al., 2014)。NR在植物养分保存中起到关键作用, 探讨其对环境变化的不同反应, 如长期氮沉降等, 是未来研究的重点(Brant & Chen, 2015)。
杉木(Cunninghamia lanceolata)是典型的南方针叶树种, 因其速生和材质优良的特征在中国亚热带地区已经种植1000多年, 目前杉木林占中国人工林面积18%, 占全球人工林面积的5% (Huang et al., 2013)。本研究以杉木为研究对象, 在模拟氮沉降14年的基础上分析杉木一年生、二年生叶和枝的C、N、P生态化学计量, 及其与衰老叶、衰老枝之间的N、P养分再吸收特征, 同时比较不同氮沉降处理时间(7年和14年) N、P养分再吸收的差异, 探讨杉木不同龄级叶和枝的养分再吸收特征对长期模拟氮沉降的响应, 为深入认识植物养分再吸收格局、机制及其对全球变化的响应提供科学依据。
1 研究区概况和研究方法
1.1 试验林概况
试验地设立于福建省沙县官庄林场白溪工区(117.72° E, 26.51° N), 海拔200 m。属中亚热带季风气候, 四季温暖适中, 日照充足, 年平均气温18.8?19.6 ℃, 年降水量1 606?1 650 mm, 无霜期271天。土壤为山地红壤, 土壤立地为I类地。试验地为南坡, 坡度为35.0°左右。样地所处的试验林为1992年由官庄林场统一造林, 树种为杉木, 总造林面积5.73 hm2。于2003年12月在选择好的人工杉木纯林中随机设置了立地条件基本相似的12块20 m × 20 m的试验样地。调查中发现, 样地的林下植被稀疏, 主要以五节芒(Miscanthus floridulus)、芒萁(Dicranopteris dichotoma)、蕨(Pteridium aquilinum var. latiusculum)等为主, 盖度在3%-5%之间。各样处理样地的林分特征及土壤理化性质见表1。Table 1
表1
表1各处理样地的主要林分特征和土壤理化性质本底值
Table 1
| 处理 Treatment | 林分特征 Stand characteristics | 土壤理化性质 Soil physicochemical properties | ||||||
|---|---|---|---|---|---|---|---|---|
| 林龄 Forest age (a) | 密度 Density (No.·hm-2) | 平均胸径 Mean DBH (cm) | 平均树高 Mean tree height (m) | pH | 有机碳 Organic carbon (g·kg-1) | 全氮 Total N (g·kg-1) | 全磷 Total P (g·kg-1) | |
| N0 | 12 | 1 717 | 16.1 | 11.8 | 4.59 | 19.23 | 0.86 | 0.22 |
| N1 | 12 | 1 633 | 16.0 | 12.2 | 4.76 | 17.31 | 0.68 | 0.13 |
| N2 | 12 | 1 683 | 16.3 | 12.2 | 4.65 | 18.88 | 0.80 | 0.17 |
| N3 | 12 | 1 625 | 16.0 | 12.1 | 4.71 | 18.14 | 0.81 | 0.14 |
新窗口打开|下载CSV
1.2 模拟氮沉降处理
本研究选择福建省三明市沙县官庄林场于1992年种植的杉木人工林为研究对象, 于杉木12年生时开始建立样地, 2004年1月开始模拟氮沉降处理, 一直延续至今。氮沉降量的确定参照了国内外同类研究(Fenn et al., 1998; Gundersen et al., 1998; 李德军等, 2004; H?egberg et al., 2006)、当地N的沉降量(樊后保等, 2000; 肖健, 2005)以及杉木对N的年需求量。按N施用量的高低, 分4种处理, 即N0 (0 kg N·hm-2·a-1, 对照), N1 (60 kg N·hm-2·a-1), N2 (120 kg N·hm-2·a-1), N3 (240 kg N·hm-2·a-1), 使用尿素(CO(NH2)2)作为氮源。每个处理重复3次, 4个处理总计12块固定样地, 试验林面积约为6 000 m2。将每个样方每次所需喷施的尿素溶解在20 L水中, 每月月初以背式喷雾器在林地人工来回均匀喷洒。按照处理水平的要求, 对照样方(N0)喷洒相同量的水, 以减少因外加的水而造成对森林生物地球化学循环的影响。1.3 样品采集及处理
1.3.1 成熟叶、枝样品2010年12月和2017年12月, 在12个样地内分别选择有代表性、长势良好的标准株2株, 用修枝剪和高枝剪采集向阳面同高度的杉木枝条若干, 根据枝条的一年生、二年生(包括二年生以上, 以下简称二年生)采集枝条, 用修枝剪剪成小段(长3?5 cm), 每个枝条样品10?15段, 对应分出两个不同龄级(一年生和二年生)的叶片若干。将枝条样品和叶片样品放在透气性好的取样袋中, 带回实验室。新鲜叶片先用10%稀盐酸冲洗, 然后用去离子水反复冲洗以彻底去除表面吸附的尘土和颗粒物。叶片样品和枝条样品均在105 ℃下杀青20 min, 之后在70 ℃下烘干48 h, 及时用微型植物碎样机粉碎, 过100目筛后置于干燥处保存备用。
1.3.2 衰老叶、枝样品
与成熟样品的采集时间和标准株相同, 用修枝剪和高枝剪采集向阳面同高度的衰老枝4?5支, 并采集其枝上叶片混合, 将衰老枝样品和衰老叶样品装入已编号的自封袋中, 采集量约200 g, 带回实验室, 经70 ℃下烘干48 h后, 粉碎, 过100目筛后置于干燥处保存备用。
1.3.3 样品测定方法
叶和枝样品C、N含量采用元素分析仪(2400II, PerkinElmer, Norwalk, USA)测定; P含量采用硝酸-高氯酸消煮-钼锑抗比色法, Lambda25紫外可见分光光度计(Lambda25 UV/VIS Spectrometer, Shelton, USA)测定。
1.4 养分再吸收计算
杉木叶/枝养分再吸收率(RE)计算公式(Huang et al., 2018)如下:RE (%) = (W1 ? W2)/W1 × 100% (1)
式中, W1为成熟叶/枝中的营养元素含量(g·kg-1), W2为衰老叶/枝中的营养元素含量(g·kg-1)。
叶片养分再吸收度(RP)以衰老叶养分转移后的最低水平浓度来表征(Killingbeck, 1996), 其中, 衰老叶的养分浓度越小, 表示养分再吸收度越大。
同时, 还分别计算了杉木不同龄级之间的N (P)养分再吸收率, 计算公式(宁秋蕊等, 2016; 张秋芳等, 2017)如下:
N (P)再吸收率(%) = (1 ? C1/C2) × 100% (2)
式中: C1/C2为衰老叶/枝N (P)含量与一年生叶/枝N (P)含量的比值、衰老叶/枝N (P)含量与二年生叶/枝N (P)含量的比值、二年生叶/枝N (P)含量与一年生叶/枝N (P)含量的比值。
1.5 数据分析
采用Excel 2016软件处理杉木人工林叶和枝的养分含量、化学计量、再吸收率数据, SigmaPlot 13.0软件进行制图, SPSS 19.0统计软件进行相关指标的最小差异显著法检验、双因素方差分析。不同小写字母表示不同氮沉降水平下差异显著(p < 0.05)。2 结果和分析
2.1 杉木叶和枝的C、N、P含量及化学计量
杉木叶的C、N和P含量均高于枝(图1)。随着龄级的增加至衰老(图1), 杉木叶和枝的C、N和P含量均逐渐降低, 从一年生叶至衰老叶, C、N和P平均含量分别降低了3.8%、28.27%和46.98%; 从一年生枝至衰老枝, C、N和P平均含量分别降低了7.10%、37.64%和36.51%。这表明杉木叶和枝在衰老过程中C、N和P含量有减少趋势, 且N和P含量在不同龄级叶和枝之间差异显著(p < 0.01)(表2)。氮沉降条件下, 杉木不同龄级叶和枝的C含量在统计上均未达到显著性差异(表2; 图1, 图2)。一年生叶、衰老叶和一年生枝的N含量均呈现出N3 > N2 > N1 > N0, N3处理显著增加N含量(p < 0.05)(表2; 图1, 图2A、2C、2D)。衰老叶和枝的P含量大小依次为N0 > N3 > N2 > N1, 二年生枝P含量为N3 > N0 > N1 > N2, 其中N1处理降低衰老叶的P含量, N2处理降低二年生枝的P含量, 氮沉降(N1、N2和N3)处理显著降低衰老枝的P含量(p < 0.05)(图1, 图2F)。Table 2
表2
表2基于双因素方差分析得到的组分、氮沉降水平及其交互作用对杉木叶和枝的生态化学计量的影响(F值)
Table 2
| 因子 Factor | F (p)值 F (p) value | |||||
|---|---|---|---|---|---|---|
| C | N | P | C:N | C:P | N:P | |
| 叶组分 Leaf components | 1.173 (0.336) | 20.401 (<0.001) | 24.425 (<0.001) | 13.832 (<0.001) | 24.072 (<0.001) | 5.728 (0.003) |
| 氮沉降 N deposition | 0.573 (0.637) | 3.272 (0.034) | 1.246 (0.309) | 3.158 (0.038) | 0.853 (0.475) | 0.631 (0.601) |
| 叶组分×氮沉降 Leaf components × N deposition | 0.280 (0.976) | 0.893 (0.543) | 0.865 (0.565) | 0.537 (0.837) | 0.963 (0.487) | 0.575 (0.807) |
| 枝组分 Branch components | 1.191 (0.329) | 9.407 (<0.001) | 19.685 (<0.001) | 6.534 (0.001) | 6.010 (0.002) | 8.176 (<0.001) |
| 氮沉降 N deposition | 0.165 (0.919) | 1.782 (0.170) | 2.130 (0.116) | 1.229 (0.315) | 1.063 (0.378) | 5.123 (0.005) |
| 枝组分×氮沉降 Branch components × N deposition | 0.398 (0.927) | 0.608 (0.781) | 0.365 (0.943) | 0.429 (0.910) | 0.371 (0.940) | 0.442 (0.902) |
新窗口打开|下载CSV
图2
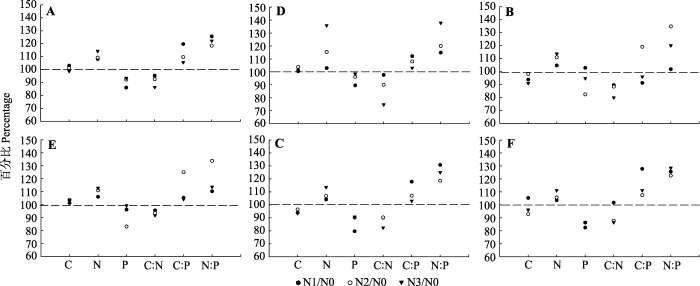 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2杉木叶和枝的测试指标(C、N、P、C:N、C:P、N:P)氮沉降处理平均值与N0处理平均值的百分比(N0/N0 = 100%)。A, 一年生叶。B, 二年生叶。C, 衰老叶。D, 一年生枝。E, 二年生枝。F, 衰老枝。N0、N1、N2、N3, 施氮量分别为0、60、120、240 kg·hm-2·a-1。
Fig. 2Mean percentage changes of measured parameters (C, N, P, C:N, C:P, N:P) of Chinese fir leaf and branch after N deposition treatment for 14 years relative to these values under control (N0) (N0/N0 = 100%). A, One-year old leaf. B, Two-year old leaf. C, Senesced leaf. D, One-year old branch. E, Two-year old branch. F, Senesced branch. N0, N1, N2, N3, nitrogen addition 0, 60, 120, 240 kg·hm-2·a-1, respectively.
C:N、C:P和N:P呈现衰老组织>成熟组织, 其中C:N和C:P为枝>叶, 但N:P为叶>枝(图1)。受氮沉降的影响, 不同组织和不同龄级的C:N大小趋势与N含量趋势相反, 表现为衰老枝>二年生枝>一年生枝>衰老叶>二年生叶>一年生叶, 且均表现为N0 > N1 > N2 > N3。N3处理的二年生叶、衰老叶C:N较N0低且达到显著性水平(p < 0.05)(图1; 表2); N2处理的二年生枝C:P比其他处理高且达到显著性水平(p < 0.05); N3处理的一年生枝N:P较N0高且达到显著性水平(p < 0.05)(图1; 表2)。叶组分与氮沉降、枝组分与氮沉降之间的交互作用对C、N和P全量及化学计量均无显著影响(表2)。通过氮沉降处理与N0处理之间的平均值的比值(图2)可知, 氮沉降处理增加了不同龄级叶和枝以及衰老叶和枝的N含量、N:P和C:P (除二年生叶外), 但降低了P含量和C:N。
2.2 杉木叶和枝的N、P再吸收率及其与化学计量的关系
一年生叶/枝和衰老叶/枝之间、二年生叶/枝和衰老叶/枝之间、一年生叶/枝和二年生叶/枝之间的REN、REP见图3。杉木叶和枝的REN、REP表现出随龄级的增加而有规律的递减, 且REP > REN。图3
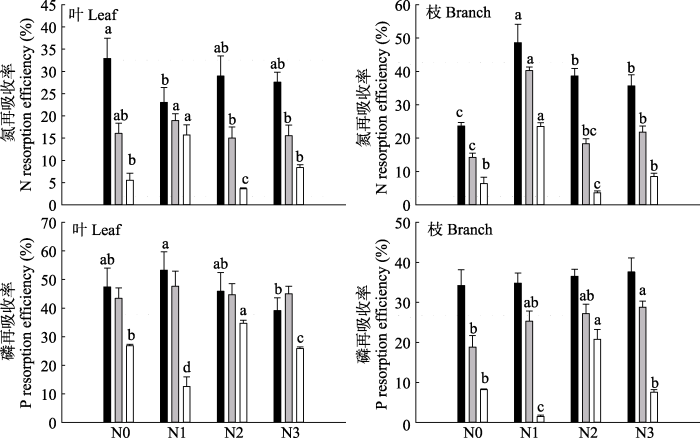 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3长期氮沉降条件下杉木叶和枝的N、P再吸收率(平均值+标准误差)。黑色柱子表示一年生叶/枝与衰老叶/枝之间的养分再吸收率; 浅灰柱子表示二年生叶/枝与衰老叶/枝之间的养分再吸收率; 白色柱子表示一年生叶/枝与二年生叶/枝之间的养分再吸收率。N0、N1、N2、N3, 施氮量分别为0、60、120、240 kg·hm-2·a-1。不同小写字母表示不同氮沉降水平下达到显著差异水平(p < 0.05)。
Fig. 3Effects of long-term nitrogen deposition on N and P resorption efficiency in leaves and branches of Chinese fir (mean + SE). The black column indicates the ratio of the nutrient resorption efficiency between one-year old leaves/branches and senescent leaves/branches; light gray column indicates the ratio of the nutrient resorption efficiency between two-year old leaves/branches and senescent leaves/branches; white column indicates the ratio of the nutrient resorption efficiency between one-year old leaves/branches and two-year-old leaves. N0, N1, N2, N3, nitrogen addition 0, 60, 120, 240 kg·hm-2·a-1, respectively. Different lowercase letters indicate significant differences under different nitrogen addition treatment levels (p < 0.05).
一年生叶和衰老叶之间、二年生叶和衰老叶之间、一年生叶和二年生叶之间的REN平均值分别为28.12%、16.39%和8.25%, REP平均值分别为46.43%、45.21%和24.98%。一年生枝和衰老枝之间、二年生枝和衰老枝之间、一年生枝和二年生枝之间的REN平均值分别为36.36%、23.64%和10.50%, REP平均值分别为35.79%、25.03%和9.50%。
氮沉降显著影响杉木叶和枝REN、REP, 且因植物器官不同而不同。N1显著降低一年生叶和衰老叶之间的REN (p < 0.05); N2和N3显著降低二年生叶和衰老叶之间REN (p < 0.05); N1显著增加一年生叶和二年生叶之间的REN (p < 0.05); 而N2则相反。氮沉降显著增加杉木枝的REN (p < 0.05)。N3显著降低一年生叶和衰老叶之间及一年生叶和二年生叶之间的REP (p < 0.05), 而N2显著增加一年生枝和二年生枝之间的REP, N3显著增加二年生枝和衰老枝之间的REP (p < 0.05)。
通过分析杉木叶和枝的REN、REP之间及其与N:P化学计量之间的关系(图4)可知, 杉木叶和枝的REN与REP之间、成熟叶和枝N:P与REN:REP之间均存在极显著的线性正相关关系(p < 0.001)。
2.3 氮沉降处理7年和14年杉木叶N、P养分再吸收特征
杉木叶N、P含量和REN、REP会随氮沉降处理的时间和杉木生长阶段而有所不同。图4
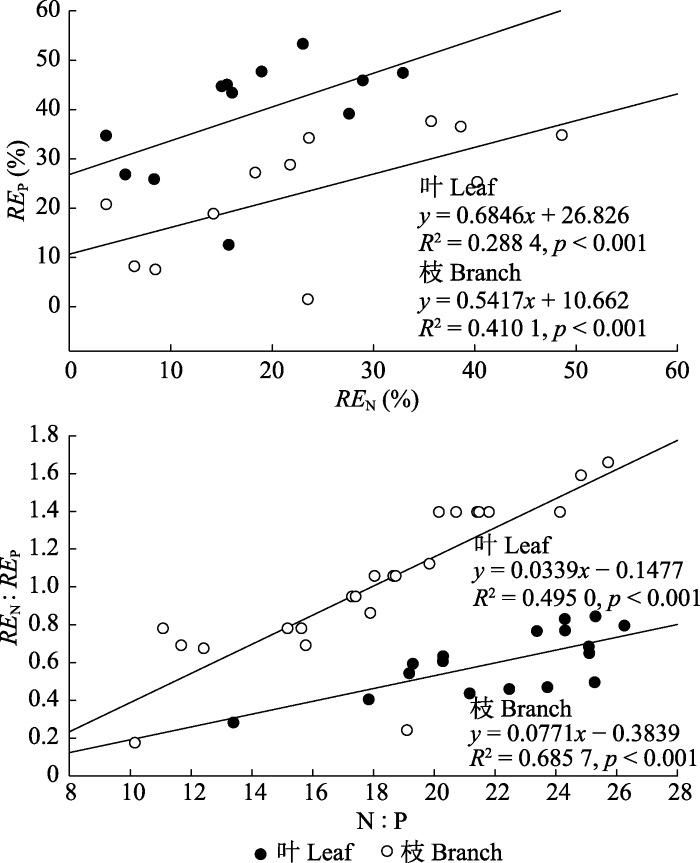 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4长期氮沉降条件下杉木叶和枝REN与REP, N:P与REN:REP的线性关系。
Fig. 4Linear relationship between N resorption efficiency (REN) and P resorption efficiency (REP), N:P and N resorption efficiency to P resorption efficiency ratio (REN:REP) of leaf and branch under long-term nitrogen deposition treatments. Data included treatments for 14 years.
2010年(氮沉降处理7年)成熟绿叶和衰老叶的N、P含量见图5。成熟绿叶的N、P含量均高于衰老叶。成熟绿叶和衰老叶中的N含量均表现为N3 > N2 > N1 > N0, N3处理显著增加了成熟绿叶N含量(p < 0.05)。与N含量相反, N3处理显著降低了成熟绿叶和衰老叶中的P含量(p < 0.05)。
图5
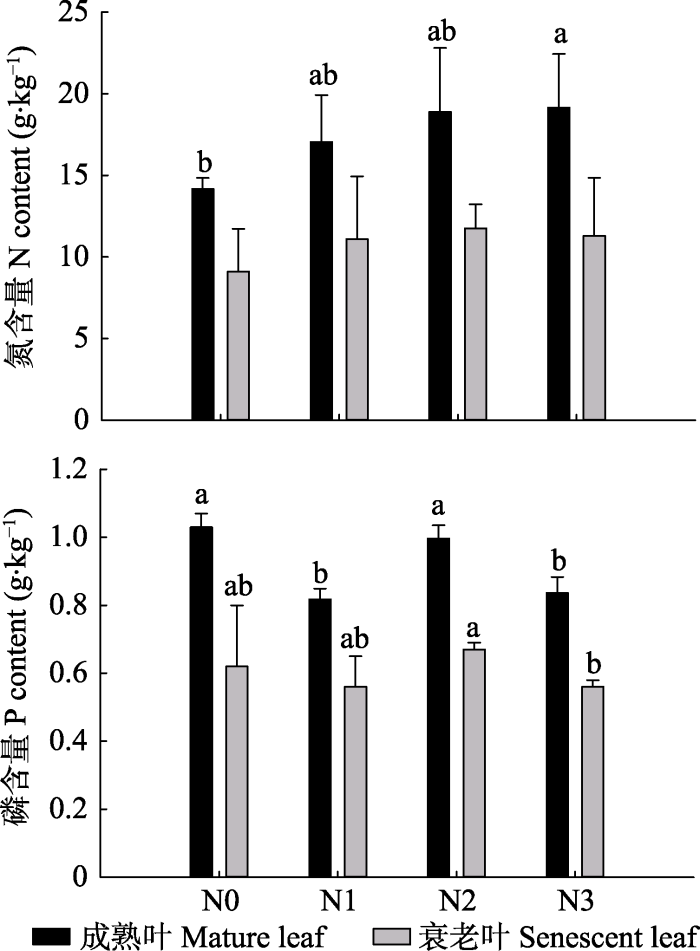 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图52010年成熟绿叶和衰老叶的N、P含量(平均值+标准误差)。N1、N2、N3, 施氮量分别为0、60、120、240 kg·hm-2·a-1。
Fig. 5N and P content of mature living and senescent leaves in 2010 (mean + SE). N0, N1, N2, N3, nitrogen addition 0, 60, 120, 240 kg·hm-2·a-1, respectively.
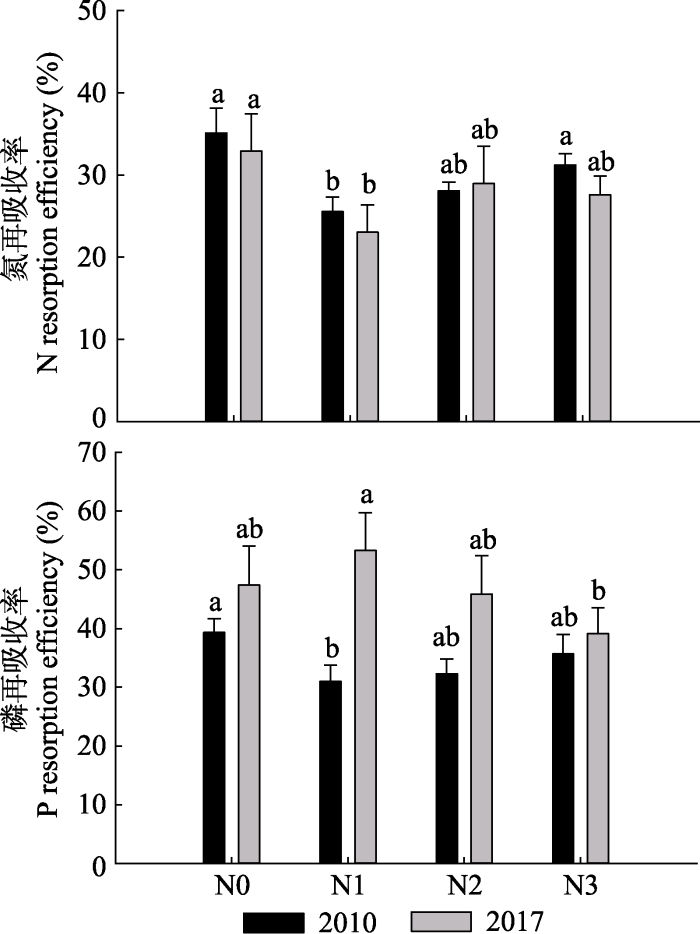
通过对比2010年(氮沉降处理7年)和2017年(氮沉降处理14年)杉木叶的N、P再吸收率(图6)可知, REP > REN。随氮沉降处理时间的增加, REN降低了, 而REP升高了, N1、N2和N3处理的REN降幅分别为9.85%、3.17%和11.71%, 其中N1显著低于N0 (p < 0.05); N1、N2和N3处理的REP增幅分别为71.98%、42.25%和9.60%。
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图62010和2017年的叶N、P再吸收率(平均值+标准误差)。N1、N2、N3, 施氮量分别为0、60、120、240 kg·hm-2·a-1。
Fig. 6N and P resorption efficiency of leaves in 2010 and 2017 (mean + SE). N0, N1, N2, N3, nitrogen addition 0, 60, 120, 240 kg·hm-2·a-1, respectively.
由表3可知, 杉木叶的REN:REP随氮沉降处理时间的增加而降低, 且REN:REP < 1; 杉木枝的REN:REP比叶REN:REP高; 二年生叶与衰老叶的REN:REP比一年生叶与衰老叶低; N1、N2、N3处理增加了一年生枝与衰老枝的REN:REP; N1处理增加了二年生枝与衰老枝的REN:REP。
Table 3
表3
表32010和2017年的叶和枝N再吸收率与P再吸收率的比值
Table 3
| 处理 Treatment | 2010 | 2017 | |||
|---|---|---|---|---|---|
| 成熟-衰叶Mature- senescent leaf | 一-衰叶 One-senesced leaf | 二-衰叶 Two-senesced leaf | 一-衰枝 One-senesced branch | 二-衰枝 Two-senesced branch | |
| N0 | 0.89 | 0.69 | 0.37 | 0.69 | 0.75 |
| N1 | 0.82 | 0.43 | 0.40 | 1.40 | 1.59 |
| N2 | 0.87 | 0.63 | 0.34 | 1.06 | 0.67 |
| N3 | 0.88 | 0.70 | 0.35 | 0.95 | 0.76 |
新窗口打开|下载CSV
3 讨论
3.1 不同龄级杉木叶和枝养分再吸收规律
本研究中, 杉木的N含量、P含量、N:P、REP均表现为叶>枝。N含量大小依次为一年生叶>二年生叶>衰老叶>一年生枝>二年生枝>衰老枝, 且N3 > N2 > N1 > N0, 而C:N则与N相反。说明活跃组织具有更高的N含量。究其原因, 第一, 叶是树木的主要光合器官, 与枝相比, 叶含有较大比例的活生物量(Wright & Westoby, 2003; Chen et al., 2012), 同时, 不同龄级养分含量及再吸收的差异主要受这些组分之间生物量分配的影响(Drenovsky & Richards, 2006); 第二, 叶的再吸收是一个高度生态相关的过程, 特别是在养分受限的生态系统中, 它允许植物减少养分流失并建立养分贮存库(Sohrt et al., 2018); 第三, 在植物的各类器官的养分转移率中, 以叶的贡献最大(费世民, 2001; Brant & Chen, 2015; 林宝平等, 2017)。随着叶和枝的衰老, 养分及化学计量比发生了变化(图1)。养分动力学表明, 养分随植物组织或器官的年龄而变化, 叶片寿命长的物种, 如常绿乔木, 随叶龄的增加, 养分浓度和光合速率可能下降(Reich et al., 2009), 由于光的利用效率通常垂直向下降低直到树冠底部(Chen et al., 2015)。叶和枝逐渐老化, 养分被吸收, 而器官和年龄效应则取决于养分限制的程度(Chen et al., 2015)。有研究表明, 这种机制不随杉木林龄的增加而衰退(林宝平等, 2017)。杉木衰老叶和枝器官的C:N、C:P和N:P比新生器官高(图1), 这是因为衰老叶养分浓度主要是由新生叶的养分状态控制。研究表明, 衰老叶的养分浓度与新生叶养分浓度正相关(相关系数R2为51%-84%)(Kobe et al., 2005), 新生叶养分浓度对衰老叶养分再吸收率影响显著。从理论上来讲, 养分再转移到幼叶会提高植物对养分吸收利用的效率, 因为叶片老化通常会降低植物的养分利用效率(Escudero & Mediavilla, 2003)。Chen等(2015)研究表明2年氮添加会改变养分动态, 但不影响不同林龄杉木叶和枝的养分再吸收率。图1
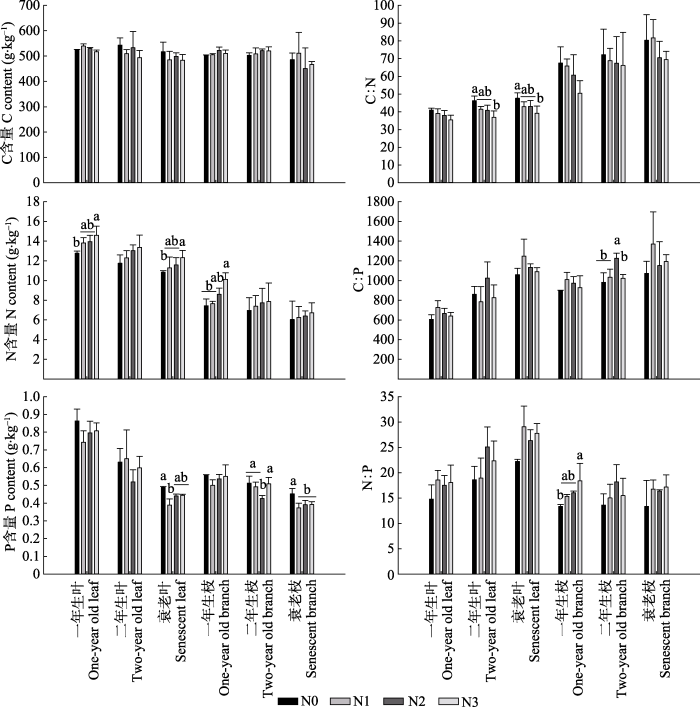 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1长期氮沉降条件下杉木叶和枝的C、N、P含量及化学计量(平均值+标准误差)。不同小写字母表示相同龄级叶或枝不同氮沉降水平下差异显著(p < 0.05)。N0、N1、N2、N3, 施氮量分别为0、60、120、240 kg·hm-2·a-1。
Fig. 1C, N, P content and stoichiometry of leaves and branches of Chinese fir under long-term nitrogen deposition (mean + SE). Different lowercase letters indicate significant differences for the same life span leaf or branch under different nitrogen deposition treatment levels (p < 0.05). N0, N1, N2, N3, nitrogen addition 0, 60, 120, 240 kg·hm-2·a-1, respectively.
杉木叶和枝均呈现REP > REN。这是因为养分投入与器官寿命长短之间的协调关系以及养分在植物体内的停留时间与植物的生产力之间的协调关系密切相关(Brant & Chen, 2015)。比如, 与枝、干等植物器官相比, 叶的养分停留时间短, 寿命也短, 会增加对养分再吸收的需求。植物器官中REN比REP更灵活(Mayor et al., 2014), REP基本没有变化(Brant & Chen, 2015; Chen et al., 2015)。4年N处理降低了落叶松叶的REN和REP, REN变化幅度大于REP (赵琼等, 2010)。本研究中对杉木针叶的研究也得出类似结果。这是因为P在土壤圈中总量丰富, 但有效态P普遍匮乏, 主要原因是P大部分处于固定态, 难以被植物直接吸收利用。热带和亚热带地区的土壤一般受P限制(Zechmeister-Boltenstern et al., 2015)。本研究对杉木叶(REN 28.12%、REP 45.82%)和枝(REN 30.00%、REP 30.42%)的研究结果也是如此, 但均低于全球REN (62.1%)、REP (64.9%)(Vergutz et al., 2012)。
叶的养分再吸收途径有两条: 从衰老叶转移到干和(或)根; 从衰老叶或者成熟叶转移到新叶(Brant & Chen, 2015)。本研究中N含量、P含量、N:P和REN:REP均表现为幼叶/枝比老叶/枝高。说明杉木优先将N、P从老叶/枝中转移到幼叶/枝中, 而不是从土壤吸收N、P。可能原因是, 从土壤中吸收N、P相比老叶/枝中转移会消耗更多的能量(Chen et al., 2015)。此外, 养分再吸收率取决于叶养分库与其他器官养分库之间的养分转移, 这些库的大小可引起植物养分的转移(Vergutz et al., 2012)。因植物新芽对养分的需求而激发木质部液流速率的增加从而使营养物质从旧叶转移到新叶。养分再吸收率取决于叶片碳水化合物通量来源的强度和同化物形成库(如生长组织和储存组织)的强度, 而源-库关系可以影响韧皮部运输, 植物的养分正是通过韧皮部的运输来实现的(张凌云和张大鹏, 2003; Yan et al., 2014)。植物对养分的吸收受植物生长阶段的影响, 比如, 地中海地区常绿落叶乔木, NR最先发生在6个月时, 从幼叶转移到其他组织, 2年后从成熟叶中转移出, 然后在叶衰老和叶凋落之前从老叶中吸收养分, 最后前三个阶段叶的营养物质重新转移到新芽, 反过来支持新叶的生长(Fife et al., 2008)。研究表明随林龄的增加, REN、REP呈现一定的趋势, 表现为幼龄林REN最高, 老龄林或成熟林以REP最高(Reed et al., 2012; 江大龙等, 2017)。本研究结果与之类似, 氮处理7年和14年时, 杉木分别为18林龄和25林龄, 此时杉木经历了中龄林和近熟林两个阶段, 相比来讲中龄林时杉木叶的REN更高, 而近熟林时杉木叶的REP更高。
3.2 长期氮沉降对杉木叶和枝养分内循环的影响
经过14年的模拟氮沉降试验, 杉木叶和枝的N含量增加、P含量降低, 这与短期(2-3年)氮沉降的影响趋势一致(樊后保等, 2007; 刘文飞等, 2008; Chen et al., 2015)。长期(14年)氮沉降条件下, 针叶N含量(平均为13.83 g·kg-1)低于短期(3年)氮沉降N含量(平均为15.4 g·kg-1), 说明随着氮沉降处理时间的增加, 杉木针叶N含量增加的幅度会逐渐降低。这与杉木不同生长阶段对N的需求有关。本试验发现, 与氮沉降处理初期(1?3年) P含量(平均为1.27 g·kg-1)(樊后保等, 2007)相比, 氮沉降处理7年和14年时杉木叶的P含量分别为0.92 g·kg-1、0.70 g·kg-1, 均有所下降, 这表明杉木叶的P含量会随氮沉降处理时间的延长而降低, 证实了氮沉降会加剧P的限制。究其原因, 氮沉降加速土壤酸化, 土壤中Al3+和NH4+溶出量增加, 抑制了杉木对P的吸收。衰老叶N、P含量和衰老枝P含量对氮沉降的响应显著, 说明杉木的N、P养分再吸收度对氮沉降引起的养分利用变化敏感。N添加增加了凋落物的N和P的再吸收能力, 但未产生显著影响(Chen et al., 2015)。14年氮沉降处理增加了不同龄级叶和枝(除二年生叶外)的N、N:P、C:P, 但降低了P和C:N。这与叶片衰老期间, N添加降低了N:P的研究结果一致(Wang et al., 2014)。与氮沉降7年相比, 氮沉降14年时杉木叶片的REN更低, 而REP更高。这与凋落物N含量在氮沉降处理时间的延长基础上仍然具有较高浓度, 并且通过落叶返回到土壤的N可能增加有关, 而P含量相反(van Heerwaarden et al., 2003)。研究表明, 13年氮沉降增加了土壤有效氮(沈芳芳等, 2018), 这也意味着衰老叶N含量随土壤速效养分的增加而增加, 导致叶片REN随土壤养分的有效性增加而降低。长期(13年)氮添加没有改变热带森林内物种的叶片N含量, 但降低了REN(Mayor et al., 2014)。生态系统长期而过量的氮沉降会降低森林P内循环率, 加剧了土壤受P限制(Matson et al., 2002; Wang et al., 2014)。研究表明, 在细胞水平上, 植物P跨液泡膜运输以实现P在体内转移再利用是植物在低P生境中的实现P素高效利用的机制(吴鹏飞和马祥庆, 2009)。在这种情况下, 植物的REP 高于REN, 全球范围内的养分富集地区也证实了这一点(Zechmeister- Boltenstern et al., 2015)。随氮沉降处理的时间增加, 杉木叶REN:REP逐渐下降(表3)。这主要是因为N、P再吸收受最后的演替阶段所驱动, 在演替过程中, REN趋于稳定, 但REP受森林林龄的增加而持续增加, 这导致成熟林中REN:REP再吸收率最低。氮沉降条件下, 杉木叶和枝的N:P与REN:REP之间存在极显著的线性相关关系(图4), 说明养分再吸收与养分状态之间存在紧密关系, 这与Vergutz等(2012)的研究结果相似。在一定的程度上, REN:REP比N:P更能说明植物的养分限制。无论植物的器官或龄级如何, 在外源养分输入的影响下, 植物叶和枝的养分水平在生理学上相互协调(Wright et al., 2006)。高养分再吸收率在营养贫乏的环境中起着至关重要的作用, 在这种环境中, 从土壤中吸取养分会需要更高的能量(Wang et al., 2014)。在热带和亚热带森林中, 植物更容易从老化、衰老叶或枝中吸收养分, 而不是从土壤中获取(Chen et al., 2015)。
4 结论
在长期氮沉降条件下, 通过比较杉木叶和枝的养分动态, 以及幼叶/枝与衰老叶/枝之间的养分动态和养分再吸收特征, 发现不同组分(叶和枝)对氮沉降的响应不同, 且叶>枝。叶和枝的REN、REP随龄级的增加至衰老而有规律递减, 且REP > REN。氮沉降处理7年, 杉木叶和枝的REN更高, 而氮沉降处理14年, REP更高。随氮沉降处理的时间增加, 杉木叶REN:REP逐渐下降, 且杉木叶和枝N:P与REN:REP之间存在密切的关系。养分再吸收作为一个过程为植物养分利用和循环提供了独特的见解, 而不同养分再吸收率的比值为估算长期环境变化条件下植物的养分循环提供了一个更好的途径。
致谢
感谢福建省三明市沙县官庄国有林场的帮助和江西省教育厅科技项目(GJJ161118)资助。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 1]
DOIURL [本文引用: 9]

Nutrient resorption (NR) plays a key role in the nutrient conservation of plants. However, a fundamental understanding of the mechanisms that control NR remains limited. In this review, we examine how intrinsic controls (e.g., genetic variability and plant development) and extrinsic environmental controls (e.g., climate and soil fertility) influence NR. We also examined conceptual NR advances, mass loss correction, measurement in non-leaf plant tissues for whole-plant nutrient budget accounting, and the use of stoichiometric ratios in place of individual elements. Nutrient resorption from senescing leaves is greater than that from stems/culms or roots. Nutrients resorbed from stems and roots in woody plants are lower than in non-woody plants. Deciduous plants are more efficient in resorbing leaf nutrients prior to senescence than are evergreen plants. Furthermore, reproductive efforts tend to increase NR. Along a latitudinal gradient of terrestrial biomes, nitrogen resorption efficiency decreases and phosphorus resorption efficiency increases with increasing temperature and precipitation; however, latitudinal patterns reflect the influences of several coupling factors such as genetic variation, climate, soil, and disturbance history. Nutrient fertilization experiments have demonstrated that increased soil fertility reduces NR. The inquiries into the impacts of ongoing climate change on NR are still at a nascent stage. Future NR studies are needed to better understand the independent effects of a wide range of genetic variation, plant development, and environment, and possibly the different responses of plants to environmental change; particularly elevated atmospheric CO2concentrations and global warming.
DOIURL [本文引用: 1]
Abstract02green and Pgreen in spring, summer, and winter, but not in autumn; N resorption proficiency generally correlated positively with Ngreen in each season, while P resorption efficiency correlated positively with Pgreen in spring and summer, but not in autumn and winter. Soil nitrogen availability correlated negatively with leaf nutrient traits in some seasons. In conclusion, leaf trait relationships varied among the seasons and among PFGs. Seasonal dynamics of leaf traits as well as soil nutrients’ relations must be considered when exploring plant feedback to soil nutrients.
DOIURLPMID [本文引用: 10]
Abstract It is unclear how or even if phosphorus (P) input alters the influence of nitrogen (N) deposition in a forest. In theory, nutrients in leaves and twigs differing in age may show different responses to elevated nutrient input. To test this possibility, we selected Chinese fir (Cunninghamia lanceolata) for a series of N and P addition experiments using treatments of +N1 - P (50 kg N ha(-1) year(-1)), +N2 - P (100 kg N ha(-1) year(-1)), -N + P (50 kg P ha(-1) year(-1)), +N1 + P, +N2 + P and -N - P (without N and P addition). Soil samples were analyzed for mineral N and available P concentrations. Leaves and twigs in summer and their litters in winter were classified as and sorted into young and old components to measure N and P concentrations. Soil mineral N and available P increased with N and P additions, respectively. Nitrogen addition increased leaf and twig N concentrations in the second year, but not in the first year; P addition increased leaf and twig P concentrations in both years and enhanced young but not old leaf and twig N accumulations. Nitrogen and P resorption proficiencies in litters increased in response to N and P additions, but N and P resorption efficiencies were not significantly altered. Nitrogen resorption efficiency was generally higher in leaves than in twigs and in young vs old leaves and twigs. Phosphorus resorption efficiency showed a minimal variation from 26.6 to 47.0%. Therefore, P input intensified leaf and twig N enrichment with N addition, leaf and twig nutrients were both gradually resorbed with aging, and organ and age effects depended on the extent of nutrient limitation. The Author 2015. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
DOIURL [本文引用: 1]
为了给林地覆盖经营雷竹(Phyllostachys violascens)林可持续经营提供理论参考,探讨了休养式覆盖经营(覆盖3a后休养3a)、长期覆盖经营(覆盖6a)和不覆盖雷竹林(CK)2年生壮龄竹鞭及其1级、2级根N、P、K、Mg、Ca、Fe浓度和养分迁移、内循环率的差异。结果表明:不同覆盖经营年限雷竹林N、P、K、Mg、Ca和Fe浓度总体上1级根显著高于2级根。1级根和2级根中均存在N、P、K、Mg的养分内循环,且1级根养分内循环率大于2级根,Fe、Ca内循环不明显。N、P、K、Mg养分浓度与养分迁移速率随时间的推延,1级根为持续降低,2级根为先升高后降低。与不覆盖雷竹林相比,休养式林地覆盖经营总体上提高了1级、2级根的N、P、K、Ca的浓度和P、K、Mg的迁移速率、N、P、K的迁移量、P、K的养分内循环率以及1级根Mg的浓度和迁移量、2级根N的迁移速率和Mg的内循环率;长期林地覆盖经营虽提高了雷竹1级根N、K的浓度和N的迁移量及2级根N的浓度和内循环率,但总体上降低了1级根P、K、Mg和2级根N、P、Mg的迁移量与1级、2级根P、Mg的迁移速率及P、K、Mg的养分内循环率。研究表明:雷竹林鞭根中存在明显的养分内循环,且1级根对养分内循环的贡献较大。休养式林地覆盖经营利于雷竹林对养分的循环利用,而长期覆盖经营阻碍了根系对养分的平衡吸收,减弱了根系养分的内循环,不利于雷竹林的生长更新。
DOIURL [本文引用: 1]
为了给林地覆盖经营雷竹(Phyllostachys violascens)林可持续经营提供理论参考,探讨了休养式覆盖经营(覆盖3a后休养3a)、长期覆盖经营(覆盖6a)和不覆盖雷竹林(CK)2年生壮龄竹鞭及其1级、2级根N、P、K、Mg、Ca、Fe浓度和养分迁移、内循环率的差异。结果表明:不同覆盖经营年限雷竹林N、P、K、Mg、Ca和Fe浓度总体上1级根显著高于2级根。1级根和2级根中均存在N、P、K、Mg的养分内循环,且1级根养分内循环率大于2级根,Fe、Ca内循环不明显。N、P、K、Mg养分浓度与养分迁移速率随时间的推延,1级根为持续降低,2级根为先升高后降低。与不覆盖雷竹林相比,休养式林地覆盖经营总体上提高了1级、2级根的N、P、K、Ca的浓度和P、K、Mg的迁移速率、N、P、K的迁移量、P、K的养分内循环率以及1级根Mg的浓度和迁移量、2级根N的迁移速率和Mg的内循环率;长期林地覆盖经营虽提高了雷竹1级根N、K的浓度和N的迁移量及2级根N的浓度和内循环率,但总体上降低了1级根P、K、Mg和2级根N、P、Mg的迁移量与1级、2级根P、Mg的迁移速率及P、K、Mg的养分内循环率。研究表明:雷竹林鞭根中存在明显的养分内循环,且1级根对养分内循环的贡献较大。休养式林地覆盖经营利于雷竹林对养分的循环利用,而长期覆盖经营阻碍了根系对养分的平衡吸收,减弱了根系养分的内循环,不利于雷竹林的生长更新。
DOIURL [本文引用: 1]
Abstract Anthropogenic additions of nitrogen (N) are expected to drive terrestrial ecosystems toward greater phosphorus (P) limitation. However, a comprehensive understanding of how an ecosystem's P cycle responds to external N inputs remains elusive, making model predictions of the anthropogenic P limitation and its impacts largely uncertain. Global. 1986-2015. Terrestrial ecosystems. We conducted a meta-analysis including 288 independent study sites from 192 articles to evaluate global patterns and controls of 10 variables associated with ecosystem P cycling under N addition. Overall, N addition increased biomass in plants (+34%) and litter (+15%) as well as plant P content (+17%), while decreasing P concentrations in plants and litter ( 8% and 11%, respectively). N addition did not change soil labile P or microbial P, but enhanced phosphatase activity (+24%). The effects of N addition on the litter P pool and soil total P remained unclear due to significant publication biases. The response of P cycling to N addition in tropical forests was different from that in other ecosystem types. N addition did not change plant biomass or phosphatase activity in tropical forests but significantly reduced plant P and soil labile P concentrations. The shift in plant P concentration under N addition was negatively correlated with the N application rate or total N load. N-induced change in soil labile P was strongly regulated by soil pH value at the control sites, with a significant decrease of 14% only in acidic soils (pH < 5.5). Our results suggest that as anthropogenic N enhancement continues in the future it could induce P limitation in terrestrial ecosystems while accelerating P cycling, particularly in tropical forests. A quantitative framework generated on the basis of this meta-analysis is useful for our understanding of ecosystem P cycling with N addition, and for incorporating the anthropogenic P limitation into ecosystem models used to analyse effects of future climate change.
DOIURL [本文引用: 2]
Both water and nutrients are limiting in arid environments, and desert plants have adapted to these limitations through numerous developmental and physiological mechanisms. In the Mono Basin, California, USA, co-dominant Sarcobatus vermiculatus and Chrysothamnus nauseosus ssp. consimilis are differentially N and P limited. We hypothesized that low leaf N resorption contributes to N-limitation in Sarcobatus and that low leaf P resorption contributes to P-limitation in Chrysothamnus. As predicted, Sarcobatus resorbed proportionally 1.7-fold less N than Chrysothamnus, but reduced leaf P in senescent leaves to lower levels than Chrysothamnus (8.0-10.8-fold lower based on leaf area or mass, respectively), consistent with N, but not P limitations in Sarcobatus. Again, as predicted, Chrysothamnus resorbed proportionally 2.0-fold less P than Sarcobatus yet reduced leaf N in senescent leaves to lower levels than Sarcobatus (1.8-1.3-fold lower based on leaf area or mass, respectively), consistent with P, but not N limitations in Chrysothamnus. Leaf N and P pools were approximately 50% of aboveground pools in both species during the growing season, suggesting leaf resorption can contribute significantly to whole plant nutrient retention. This was consistent with changes in leaf N vs. P concentration as plants grew from seedlings to adults. Our results support the conclusion that N-limitation in Sarcobatus and P-limitation in Chrysothamnus are in part caused by physiological (or other) constraints that prevent more efficient resorption of N or P, respectively. For these species, differential nutrient resorption may be a key physiological component contributing to their coexistence in this saline, low resource habitat.
DOIURL [本文引用: 1]
1 Cost-benefit models predict that leaf life span depends on its initial photosynthetic rate and construction cost and on the rate of decline in photosynthesis with age. Leaf gas exchange rates and N contents were measured in nine woody evergreen Mediterranean species with different leaf life spans to determine the effects of leaf ageing on photosynthetic N use efficiency (PNUE). N costs of leaf construction were assumed to be in part dependent on N resorption from senescing leaves. 2 Leaf ageing had significant negative effects on photosynthetic rates per unit leaf area. As N content per unit leaf area did not decline until the end of leaf life, PNUE also decreased with age. PNUE generally declined faster in species with a shorter leaf life span. There were no significant interspecific differences in maximum CO2assimilation rates per unit leaf area and in N resorption that could be related to differences in leaf life span. 3 As PNUE decreases with leaf age, shedding of the older leaves and retranslocation of N to the current year's leaf biomass would result in an increase in the mean instantaneous efficiency of use of the N retranslocated. However, total CO2assimilation can be improved by such shedding only when the increase in the efficiency of use of the remobilized N compensates for the remaining N lost in the shed leaves. 4 The photosynthesis of the old leaf cohorts exceeded the increase in photosynthesis that would be obtained from the N retranslocated to the younger leaves, given the observed efficiencies of N resorption. The retention of old leaves thus resulted in a higher whole-canopy CO2assimilation, despite their low PNUE.
[本文引用: 2]
[本文引用: 2]
DOIURL [本文引用: 1]
在福建南平地区 ,对两片杉木人工林 (标记为FFC和XQF)的氮素沉降进行了为期 3a(1994~ 1996 )的定位研究 .结果表明 ,降水通过林冠后 ,穿透雨的氮素含量没有发生明显变化 ,但树干径流的养分富集现象则十分显著 .降雨、穿透雨和树干径流中的氮素含量表现出明显的季节变化 ,均以夏季最低 ,冬季最高 ,春秋季居中 ,这种格局强烈受降雨量的控制 .在FFC和XQF监测场 ,每年从降雨中输入的NH+4 N分别为 8.73kghm-2 和 4.6 2kghm-2 ,NO-3 N分别为 9.36kghm-2 和 6 .83kghm-2 ,总计氮素输入分别为 18.0 9kghm-2 及 11.45kghm-2 ,其中16 .4% (2 .97kghm-2 )和 4.9% (0 .5 6kghm-2 )在降水过程中直接被两林分的林冠所吸收 .吸收行为主要出现在春、夏两季 ,这是杉木生长旺盛的阶段 ,而在秋、冬季则主要表现为淋溶 .林龄和密度是影响林冠对氮素吸收量的两个主要因子 .图 3表 1参 2 2
DOIURL [本文引用: 1]
在福建南平地区 ,对两片杉木人工林 (标记为FFC和XQF)的氮素沉降进行了为期 3a(1994~ 1996 )的定位研究 .结果表明 ,降水通过林冠后 ,穿透雨的氮素含量没有发生明显变化 ,但树干径流的养分富集现象则十分显著 .降雨、穿透雨和树干径流中的氮素含量表现出明显的季节变化 ,均以夏季最低 ,冬季最高 ,春秋季居中 ,这种格局强烈受降雨量的控制 .在FFC和XQF监测场 ,每年从降雨中输入的NH+4 N分别为 8.73kghm-2 和 4.6 2kghm-2 ,NO-3 N分别为 9.36kghm-2 和 6 .83kghm-2 ,总计氮素输入分别为 18.0 9kghm-2 及 11.45kghm-2 ,其中16 .4% (2 .97kghm-2 )和 4.9% (0 .5 6kghm-2 )在降水过程中直接被两林分的林冠所吸收 .吸收行为主要出现在春、夏两季 ,这是杉木生长旺盛的阶段 ,而在秋、冬季则主要表现为淋溶 .林龄和密度是影响林冠对氮素吸收量的两个主要因子 .图 3表 1参 2 2
DOIURLMagsci [本文引用: 3]
<p>对江苏省下蜀林场火炬松人工林(15年生)地上部分养分体内转移和内循环特点进行了初步研究,结果表明:(1)在体内养分转移中,叶的贡献最大,N、P、K和Mg元素的平均转移量均大于枝和干;地上部分养分N、P、K和Mg元素的转移率分别为44.9% ,34.5% ,20.8%和30.7%。(2)地上部分养分归还量N、P、K、Ca和Mg元素分别55.787,6.13,26.70,70.79和20.30kg·hm<sup>-2</sup>·a<sup>-1</sup> 分别占需要量的36.67% ,44.65% 55.3% ,78.02%和42.84% ;地上实际消耗土壤中的养分量N、P、K、Ca和Mg分别为22.48,2.13,4.64,26.98和7.41kg·hm<sup>-2</sup>·a<sup>-1</sup>,分别占吸收量的23.99% ,22.76% ,11.84% ,24.32%和21.09%。(3)与当地的杉木林和次生栎林相比,该火炬松人工林养分利用率高,并处于养分消耗阶段。</p>
DOIURLMagsci [本文引用: 3]
<p>对江苏省下蜀林场火炬松人工林(15年生)地上部分养分体内转移和内循环特点进行了初步研究,结果表明:(1)在体内养分转移中,叶的贡献最大,N、P、K和Mg元素的平均转移量均大于枝和干;地上部分养分N、P、K和Mg元素的转移率分别为44.9% ,34.5% ,20.8%和30.7%。(2)地上部分养分归还量N、P、K、Ca和Mg元素分别55.787,6.13,26.70,70.79和20.30kg·hm<sup>-2</sup>·a<sup>-1</sup> 分别占需要量的36.67% ,44.65% 55.3% ,78.02%和42.84% ;地上实际消耗土壤中的养分量N、P、K、Ca和Mg分别为22.48,2.13,4.64,26.98和7.41kg·hm<sup>-2</sup>·a<sup>-1</sup>,分别占吸收量的23.99% ,22.76% ,11.84% ,24.32%和21.09%。(3)与当地的杉木林和次生栎林相比,该火炬松人工林养分利用率高,并处于养分消耗阶段。</p>
DOIURL [本文引用: 1]
DOIURLPMID [本文引用: 1]
Abstract Internal nutrient recycling through retranslocation (resorption) is important for meeting the nutrient demands of new tissue production in trees. We conducted a comparative study of nutrient retranslocation from leaves of five tree species from three genera grown in plantation forests for commercial or environmental purposes in southern Australia--Acacia mearnsii De Wild., Eucalyptus globulus Labill., E. fraxinoides H. Deane & Maiden, E. grandis W. Hill ex Maiden and Pinus radiata D. Don. Significant amounts of nitrogen, phosphorus and potassium were retranslocated during three phases of leaf life. In the first phase, retranslocation occurred from young leaves beginning 6 months after leaf initiation, even when leaves were physiologically most active. In the second phase, retranslocation occurred from mature green leaves during their second year, and in the third phase, retranslocation occurred during senescence before leaf fall. Nutrient retranslocation occurred mainly in response to new shoot production. The pattern of retranslocation was remarkably similar in the leaves of all study species (and in the phyllodes of Casuarina glauca Sieber ex Spreng.), despite their diverse genetics, leaf forms and growth rates. There was no net retranslocation of calcium in any of the species. The amounts of nutrients at the start of each pre-retranslocation phase had a strong positive relationship with the amounts subsequently retranslocated, and all species fitted a common relationship. The percentage reduction in concentration or content (retranslocation efficiency) at a particular growth phase is subject to many variables, even within a species, and is therefore not a meaningful measure of interspecific variation. It is proposed that the pattern of retranslocation and its governing factors are similar among species in the absence of interspecies competition for growth and crown structure which occurs in mixed species stands.
DOIURLPMID [本文引用: 1]
Humans continue to transform the global nitrogen cycle at a record pace, reflecting an increased combustion of fossil fuels, growing demand for nitrogen in agriculture and industry, and pervasive inefficiencies in its use. Much anthropogenic nitrogen is lost to air, water, and land to cause a cascade of environmental and human health problems. Simultaneously, food production in some parts of the world is nitrogen-deficient, highlighting inequities in the distribution of nitrogen-containing fertilizers. Optimizing the need for a key human resource while minimizing its negative consequences requires an integrated interdisciplinary approach and the development of strategies to decrease nitrogen-containing waste.
DOIURL [本文引用: 1]
Impact of nitrogen (N) deposition was studied by comparing N fluxes, N concentrations and N pool sizes in vegetation and soil in five coniferous forest stands at the NITREX sites: G02rdsj02n (GD), Sweden, Klosterhede (KH), Denmark, Aber (AB), Wales, UK, Speuld (SP), the Netherlands, and Ysselsteyn (YS), the Netherlands. The sites span a N- deposition gradient from 13 to 59 kg N ha 611 yr 611 . Measurements of soil N transformation rates by laboratory and field incubations were part of the site comparison. Further, results from 4–5 yr of NH 4 NO 3 addition (35 kg N ha 611 yr 611 ) at low deposition sites (GD, KH, AB) and 6 yr of N removal (roofs) at high deposition sites (SP, YS) were included in the analysis. Significant correlations were found between a range of variables including N concentrations in foliage and litter, soil N transformation rates and forest floor characteristics. Using the methods from principal component analysis (PCA) these variables were summarized to an index of site N status that assigned the lowest N status to GD and the highest to YS. Site N status increased with N deposition with the exception that AB was naturally rich in N. Nitrate leaching was significantly correlated with N status but not correlated with N deposition. Forest floor mass and root biomass decreased with increased N status. Characteristics of the mineral soil were not correlated with vegetation and forest floor variables. High ja:math ratios in the mineral soil at the high-N deposition sites (SP, YS) suggest that the mineral soil pool changes slowly and need not change for N saturation to occur. Nitrogen transformation rates measured in laboratory incubations did not agree well with rates measured in the field except for a good correlation between ‘gross’ mineralization in the laboratory and ‘net’ mineralization in the field. The changes in N concentrations and fluxes after manipulation of N input followed the direction expected from the site comparison: increases at N addition and decreases at N removal sites. Nitrate leaching responded within the first year of treatment at all sites, whereas responses in vegetation and soil were delayed. Changes in N status by the manipulation treatments were small compared to the differences between sites. Changes in nitrate leaching were small at the low-N status sites and substantial at the high-N status sites. Nitrogen-limited and N-saturated forest ecosystems could be characterized quantitatively.
DOIURL [本文引用: 1]
Relations among nitrogen load, soil acidification and forest growth have been evaluated based on short-term (<15 years) experiments, or on surveys across gradients of N deposition that may also include variations in edaphic conditions and other pollutants, which confound the interpretation of effects of N per se . We report effects on trees and soils in a uniquely long-term (30 years) experiment with annual N loading on an un-polluted boreal forest. Ammonium nitrate was added to replicated ( N =3) 0.09 ha plots at two doses, N1 and N2, 34 and 68 kg N ha 611 yr 611 , respectively. A third treatment, N3, 108 kg N ha 611 yr 611 , was terminated after 20 years, allowing assessment of recovery during 10 years. Tree growth initially responded positively to all N treatments, but the longer term response was highly rate dependent with no gain in N3, a gain of 50 m 3 ha 611 stemwood in N2 and a gain of 100 m 3 ha 611 stemwood in excess of the control (N0) in N1. High N treatments caused losses of up to 70% of exchangeable base cations (Ca 2+ , Mg 2+ , K + ) in the mineral soil, along with decreases in pH and increases in exchangeable Al 3+ . In contrast, the organic mor-layer (forest floor) in the N-treated plots had similar amounts per hectare of exchangeable base cations as in the N0 treatment. Magnesium was even higher in the mor of N-treated plots, providing evidence of up-lift by the trees from the mineral soil. Tree growth did not correlate with the soil Ca/Al ratio (a suggested predictor of effects of soil acidity on tree growth). A boron deficiency occurred on N-treated plots, but was corrected at an early stage. Extractable NH 4 + and NO 3 61 were high in mor and mineral soils of on-going N treatments, while NH 4 + was elevated in the mor only in N3 plots. Ten years after termination of N addition in the N3 treatment, the pH had increased significantly in the mineral soil; there were also tendencies of higher soil base status and concentrations of base cations in the foliage. Our data suggest the recovery of soil chemical properties, notably pH, may be quicker after removal of the N-load than predicted. Our long-term experiment demonstrated the fundamental importance of the rate of N application relative to the total amount of N applied, in particular with regard to tree growth and C sequestration. Hence, experiments adding high doses of N over short periods do not mimic the long-term effects of N deposition at lower rates.
DOIURL [本文引用: 2]
How nitrogen deposition and increasing precipitation would affect leaf nutrient concentration and internal nutrient cycling of desert plants is still unclear. The aim of our study was to test the resp
DOIURL [本文引用: 2]
DOIURL [本文引用: 1]
This study aimed to determine the influence of different harvest residue management strategies on tree growth, soil carbon (C) concentrations, soil nitrogen (N) availability and ecosystem C stocks 15 years after replanting second rotation Chinese fir (Cunninghamia lanceolata), an important plantation species in subtropical China. Such information is needed for designing improved management strategies for reforestation programmes in subtropical environments aimed at mitigating CO2 emissions.Four harvest residue management treatments including slash burning, whole tree, stem-only and double residue retention were applied to sixteen 20 m x 30 m plots in a randomized complete block design with four replicates. Tree growth was measured annually and soil properties were measured at 3 year intervals over a 15 year period after re-planting.Cumulative diameter growth at age 15 years was significantly smaller in the slash burning than the whole tree and double residue harvest treatments. Hot water extractable N concentrations increased with the increased organic residue retention levels and significant differences were observed between double residue and slash burning treatments. Harvest residue management had no significant effect on the soil C concentrations to 40 cm depth. ANOVA showed that harvest residue management had no significant effect on total biomass carbon at age 15, but the plantation ecosystem (soil C at 0-40 cm depth plus forest biomass C) had significantly lower C mass in the slash burning treatment compared with whole tree, stem only harvest and double residue harvest treatments.These observations suggest that organic residue retention during the harvesting could improve the growth and ecosystem C stocks of Chinese fir in second rotation forest plantations in subtropical China and highlight the importance of viewing the ecosystem as a whole when evaluating the impact of harvest residue management on C stocks.
DOIURL [本文引用: 4]
【目的】植物养分重吸收是指植物落叶前将养分转移到其他活组织中保存或直接利用的一种现象。对养分重吸收控制机制进行的系统了解,可为完善植物养分循环模型提供一定的理论依据。【方法】通过对国内外已有研究进行分析,全面比较不同生活型、林龄以及外界的环境(气候和土壤肥力)对植物的养分重吸收的影响。【结果】1木本植物茎、根的养分重吸收要小于非木本植物,非木本植物养分重吸收对不同的土壤肥力表现出更强的可塑性;落叶植物比常绿植物重吸收更多的养分;2幼龄林有更高的氮元素重吸收率;老龄林能重吸收更多的磷元素;3随着年均温和年均降水量的升高,磷元素重吸收率增加,氮元素重吸收率下降;4干旱、洪水、飓风等极端天气都会显著影响养分重吸收;5土壤肥力与养分重吸收呈负相关。相对于叶片,木本植物根、茎养分以及微量元素重吸收的研究还比较缺乏,对于养分吸收和重吸收对能量的消耗也缺乏了解。此外,长期气候变化对养分重吸收的影响也有待研究。【结论】养分重吸收是植物养分循环的一个重要部分,对它的精确估计将有利于完善全球气候变化模型,对进一步准确预测未来全球变化的趋势具有重要意义。
DOIURL [本文引用: 4]
【目的】植物养分重吸收是指植物落叶前将养分转移到其他活组织中保存或直接利用的一种现象。对养分重吸收控制机制进行的系统了解,可为完善植物养分循环模型提供一定的理论依据。【方法】通过对国内外已有研究进行分析,全面比较不同生活型、林龄以及外界的环境(气候和土壤肥力)对植物的养分重吸收的影响。【结果】1木本植物茎、根的养分重吸收要小于非木本植物,非木本植物养分重吸收对不同的土壤肥力表现出更强的可塑性;落叶植物比常绿植物重吸收更多的养分;2幼龄林有更高的氮元素重吸收率;老龄林能重吸收更多的磷元素;3随着年均温和年均降水量的升高,磷元素重吸收率增加,氮元素重吸收率下降;4干旱、洪水、飓风等极端天气都会显著影响养分重吸收;5土壤肥力与养分重吸收呈负相关。相对于叶片,木本植物根、茎养分以及微量元素重吸收的研究还比较缺乏,对于养分吸收和重吸收对能量的消耗也缺乏了解。此外,长期气候变化对养分重吸收的影响也有待研究。【结论】养分重吸收是植物养分循环的一个重要部分,对它的精确估计将有利于完善全球气候变化模型,对进一步准确预测未来全球变化的趋势具有重要意义。
DOIURL [本文引用: 1]
Analyses of nitrogen and phosphorus in the senesced leaves of 89 species of deciduous and evergreen woody perennials were used (1) to discover the limits of ultimate potential resorption (maximal withdrawal of nutrients from senescing leaves), (2) to determine a means by which resorption can be categorized as complete or incomplete, (3) to develop the concept of resorption proficiency (measured as the levels to which nutrients have been reduced in senesced leaves), (4) to compare resorption in evergreen vs. deciduous species, (5) to assess the impact of phylogeny on resorption, (6) to compare resorption in actinorhizal vs. non-nitrogen-fixing species, and (7) to consider the efficacy of using multiple measures of resorption to answer questions regarding the function and evolution of this process, rather than relying solely on analyses of resorption efficiency (percentage reduction of nutrients between green and senesced leaves). Concentrations of 0.3% nitrogen and 0.01% phosphorus in senesced leaves represent ultimate potential resorption of these nutrients in woody perennials. Resorption proficiency and potential resorption were quantitatively defined in two models that describe both resorption that is maximal and biochemically complete, and that which is not. Resorption is highly proficient in plants that have reduced nitrogen and phosphorus in their senescing leaves to concentrations below 0.7% and 0.05%, respectively. An important feature of knowing the levels to which nutrients can be reduced in senescing leaves is that these values offer an objective gauge by which to measure the success of resorption as a nutrient conservation mechanism. Evergreens were significantly more proficient at resorbing phosphorus than were deciduous species (0.045% vs. 0.067% P in senesced leaves, respectively) and plants capable of symbiotic nitrogen fixation were significantly less proficient at resorbing nitrogen than were nonfixers (1.6% vs. 0.9% N in senesced leaves, respectively). Resorption proficiency appeared to parallel some phylogenic trends, yet the influence of phylogeny was not so significant as to overwhelm the effects of recent selection. The ability of plants to reduce nitrogen in senescing leaves was significantly correlated with their ability to reduce phosphorus. Measurement and analysis of resorption proficiency, when coupled with concurrent consideration of potential resorption and resorption efficiency, should facilitate and expedite the ongoing attempt to resolve complex questions regarding the environmental constraints that influence resorption, and the selection pressures that have directed the evolution of this process.
DOIURL [本文引用: 1]
.
DOIURL [本文引用: 1]
探讨了3种南亚热带优势树种荷木(Schima superba)、锥栗(Castanopsis chinensis)和黄果厚壳桂(Cryptocarya concinna)的幼苗对模拟氮沉降增加的响应.实验分为对照组A和处理组B、C、D和E,分别以NH4NO3形式人为喷施0、5、10、15和30g N/(m2·a).研究结果表明,高氮处理组的幼苗生长逐渐受到抑制,而中氮处理则大大促进了幼苗的生长.经过7个月的处理,3种树苗的净光合速率呈现出随氮处理水平增加而先增加后减小的特点,即C组的净光合速率最高,而D组则开始下降.荷木幼苗的水分利用效率和锥栗幼苗的气孔导度的变化趋势与净光合速率一样,但黄果厚壳桂幼苗的气孔导度和水分利用效率及荷木幼苗气孔导度各处理间差异不明显.荷木和黄果厚壳桂幼苗的光合色素含量随处理水平增加而增加,锥栗幼苗的Chl a 和Chl(a+b)含量以B组最高,总体趋势呈现出随处理水平先增加后减小的特点,但类胡萝卜素随处理水平增加而增加.
DOIURL [本文引用: 1]
探讨了3种南亚热带优势树种荷木(Schima superba)、锥栗(Castanopsis chinensis)和黄果厚壳桂(Cryptocarya concinna)的幼苗对模拟氮沉降增加的响应.实验分为对照组A和处理组B、C、D和E,分别以NH4NO3形式人为喷施0、5、10、15和30g N/(m2·a).研究结果表明,高氮处理组的幼苗生长逐渐受到抑制,而中氮处理则大大促进了幼苗的生长.经过7个月的处理,3种树苗的净光合速率呈现出随氮处理水平增加而先增加后减小的特点,即C组的净光合速率最高,而D组则开始下降.荷木幼苗的水分利用效率和锥栗幼苗的气孔导度的变化趋势与净光合速率一样,但黄果厚壳桂幼苗的气孔导度和水分利用效率及荷木幼苗气孔导度各处理间差异不明显.荷木和黄果厚壳桂幼苗的光合色素含量随处理水平增加而增加,锥栗幼苗的Chl a 和Chl(a+b)含量以B组最高,总体趋势呈现出随处理水平先增加后减小的特点,但类胡萝卜素随处理水平增加而增加.
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
为探究不同林龄杉木成熟叶与衰老叶之间大量元素的转移规律,选择中国亚热带地区8、14、21、46年生杉木,测定其成熟叶和衰老叶中N、P、K、Ca、Mg含量。结果表明:不同林龄杉木成熟叶中养分含量差异显著(Mg除外),但未表现出有规律的变化,衰老叶中N、K、Mg含量不受林龄的影响。不同林龄杉木针叶大量元素平均转移率表现为REKREPRENREMgRECa,其中,Ca表现为负转移,不同林龄杉木针叶N、K、Mg转移率无显著差异。在杉木成熟叶中,P、K、Ca、Mg含量与其转移率存在极显著的线性正相关,杉木衰老叶片中营养元素均与其转移率存在负相关。研究表明:杉木通过将大量元素从衰老叶片转移到成熟叶片,从而减少养分损失,这种机制不随林龄增加而衰退。
DOIURL [本文引用: 2]
为探究不同林龄杉木成熟叶与衰老叶之间大量元素的转移规律,选择中国亚热带地区8、14、21、46年生杉木,测定其成熟叶和衰老叶中N、P、K、Ca、Mg含量。结果表明:不同林龄杉木成熟叶中养分含量差异显著(Mg除外),但未表现出有规律的变化,衰老叶中N、K、Mg含量不受林龄的影响。不同林龄杉木针叶大量元素平均转移率表现为REKREPRENREMgRECa,其中,Ca表现为负转移,不同林龄杉木针叶N、K、Mg转移率无显著差异。在杉木成熟叶中,P、K、Ca、Mg含量与其转移率存在极显著的线性正相关,杉木衰老叶片中营养元素均与其转移率存在负相关。研究表明:杉木通过将大量元素从衰老叶片转移到成熟叶片,从而减少养分损失,这种机制不随林龄增加而衰退。
DOIURL [本文引用: 1]
由于人类活动加剧,世界许多地区的大气氮沉降出现明显的增加,对森林植物和土壤产生明显影响.在福建沙县,通过野外模拟试验研究了杉木人工林针叶养分含量对氮沉降增加的响应.试验设计为4种处理,分别为N0(N,0kghm-2a-1,对照)、N1(N,60kghm-2a-1)、N2(N,120kghm-2a-1)和N3(N,240kghm-2a-1),每处理重复3次.通过3a监测发现,相对于N0处理,N1、N2和N3处理分别使杉木针叶平均N含量增加18.25%、11.68%和13.14%,而C/N比值分别下降15.07%、9.96%和12.01%.尽管养分含量随时间变化表现出一定的波动,但氮沉降增加在一定程度上降低了针叶中的P、K、Ca、Mg含量,致使N/P、N/K、N/Ca、N/Mg比值上升,影响了针叶的养分平衡.图2表1参33
DOIURL [本文引用: 1]
由于人类活动加剧,世界许多地区的大气氮沉降出现明显的增加,对森林植物和土壤产生明显影响.在福建沙县,通过野外模拟试验研究了杉木人工林针叶养分含量对氮沉降增加的响应.试验设计为4种处理,分别为N0(N,0kghm-2a-1,对照)、N1(N,60kghm-2a-1)、N2(N,120kghm-2a-1)和N3(N,240kghm-2a-1),每处理重复3次.通过3a监测发现,相对于N0处理,N1、N2和N3处理分别使杉木针叶平均N含量增加18.25%、11.68%和13.14%,而C/N比值分别下降15.07%、9.96%和12.01%.尽管养分含量随时间变化表现出一定的波动,但氮沉降增加在一定程度上降低了针叶中的P、K、Ca、Mg含量,致使N/P、N/K、N/Ca、N/Mg比值上升,影响了针叶的养分平衡.图2表1参33
DOIURLPMID [本文引用: 2]
China is experiencing intense air pollution caused in large part by anthropogenic emissions of reactive nitrogen(1,2). These emissions result in the deposition of atmospheric nitrogen (N) in terrestrial and aquatic ecosystems, with implications for human and ecosystem health, greenhouse gas balances and biological diversity(1,3-5). However, information on the magnitude and environmental impact of N deposition in China is limited. Here we use nationwide data sets on bulk N deposition, plant foliar N and crop N uptake (from long-term unfertilized soils) to evaluate N deposition dynamics and their effect on ecosystems across China between 1980 and 2010. We find that the average annual bulk deposition of N increased by approximately 8 kilograms of nitrogen per hectare (P < 0.001) between the 1980s (13.2 kilograms of nitrogen per hectare) and the 2000s (21.1 kilograms of nitrogen Per hectare). Nitrogen deposition rates in the industrialized and agriculturally intensified regions of China are as high as the peak levels of deposition in northwestern Europe in the 1980s(6), before the introduction of mitigation measures(7,8). Nitrogen from ammonium (NH4+) is the dominant form of N in bulk deposition, but the rate of increase is largest for deposition of N from nitrate (NO3-), in agreement with decreased ratios of NH3 to NOx emissions since 1980. We also find that the impact of N deposition on Chinese ecosystems includes significantly increased plant foliar N concentrations in natural and semi-natural (that is, non-agricultural) ecosystems and increased crop N uptake from long-term-unfertilized crop-lands. China and other economies are facing a continuing challenge to reduce emissions of reactive nitrogen, N deposition and their negative effects on human health and the environment.
DOIURL [本文引用: 1]
植物养分重吸收是植物组织或器官在衰老脱落前将部分养分(主要如氮、磷)转移到其他成活组织的过程,能延长养分在植物体内的存留时间,提高养分利用效率,减少植物对土壤养分的依赖,是植物适应环境的重要策略之一。本研究综述了土壤水分和肥力、光照、温度等环境因子和寿命、个体发育等遗传特性对植物叶片氮磷养分重吸收的影响。不同生活型或功能型的叶片养分重吸收不同;施肥会改变土壤速效养分的含量和比例,从而调控叶片养分重吸收;土壤水分通过调节土壤养分有效性而影响叶片养分重吸收;植物在生长过程中,不同时期对养分的需求有所差异,对养分的敏感程度和吸收能力也各不相同,从而表现出不同叶片养分重吸收特征;除了叶片之外,植物的其他组织器官(如,细茎、树木的芯材和能够储存养分的根)也可以进行养分重吸收。因此,植物叶片养分重吸收易受多种因素的影响,不同物种的重吸收能力存在差异,同一物种在不同时、空及相关因素变化下的重吸收特性也不同。深入研究植物养分重吸收规律和调控机制有助于进一步揭示植物的环境适应性,能为提高水肥管理水平,权衡生产力和适应性,实现系统生产和生态可持续发展提供理论依据。
DOIURL [本文引用: 1]
植物养分重吸收是植物组织或器官在衰老脱落前将部分养分(主要如氮、磷)转移到其他成活组织的过程,能延长养分在植物体内的存留时间,提高养分利用效率,减少植物对土壤养分的依赖,是植物适应环境的重要策略之一。本研究综述了土壤水分和肥力、光照、温度等环境因子和寿命、个体发育等遗传特性对植物叶片氮磷养分重吸收的影响。不同生活型或功能型的叶片养分重吸收不同;施肥会改变土壤速效养分的含量和比例,从而调控叶片养分重吸收;土壤水分通过调节土壤养分有效性而影响叶片养分重吸收;植物在生长过程中,不同时期对养分的需求有所差异,对养分的敏感程度和吸收能力也各不相同,从而表现出不同叶片养分重吸收特征;除了叶片之外,植物的其他组织器官(如,细茎、树木的芯材和能够储存养分的根)也可以进行养分重吸收。因此,植物叶片养分重吸收易受多种因素的影响,不同物种的重吸收能力存在差异,同一物种在不同时、空及相关因素变化下的重吸收特性也不同。深入研究植物养分重吸收规律和调控机制有助于进一步揭示植物的环境适应性,能为提高水肥管理水平,权衡生产力和适应性,实现系统生产和生态可持续发展提供理论依据。
DOIURLPMID [本文引用: 1]
The sources and distribution of anthropogenic nitrogen (N), including N fertilization and N fixed during fossil-fuel combustion, are rapidly becoming globally distributed. Responses of terrestrial ecosystems to anthropogenic N inputs are likely to vary geographically. In the temperate zone, long-term N inputs can lead to increases in plant growth and also can result in over-enrichment with N, eventually leading to increased losses of N via solution leaching and trace-gas emissions, and in some cases, to changes in species composition and to ecosystem decline. However, not all ecosystems respond to N deposition similarly; their response depends on factors such as successional state, ecosystem type, N demand or retention capacity, land-use history, soils, topography, climate, and the rate, timing, and type of N deposition. We point to some of the conditions under which anthropogenic impacts can be significant, some of the factors that control variations in response, and some areas where uncertainty is large due to limited information.
DOIURL [本文引用: 2]
The concentration, stoichiometry and resorption of nitrogen (N) and phosphorus (P) in plant leaves are often used as proxies of the availability of these growth-limiting nutrients, but the responses of these metrics to changes in nutrient availability remain largely untested for tropical forest trees.We evaluated changes in N and P concentrations, N/P ratios and resorption for four common tree species after 13 years of factorial N and P additions in a lowland tropical forest in Panama.Chronic P addition increased foliar P concentrations, decreased P resorption proficiency and decreased N/P ratios in three locally common eudicot tree species (Alseis blackiana, Heisteria concinna, Tetragastris panamensis). The increase in foliar P involved similar proportional increases in organic and inorganic P in two species and a disproportionately large increase in inorganic P in A. blackiana.Nitrogen addition did not alter foliar N concentrations in any species, but did decrease N resorption proficiency in H. concinna.A fourth species, the palm Oenocarpus mapora, demonstrated remarkably static foliar nutrient concentrations, responding only with a marginal decrease in P resorption proficiency under N plus P co-addition.Synthesis. Collectively, these results suggest that adjustment of N/P ratios can be expected in eudicots exposed to elevated P, but foliar N appears to already be at optimal levels in these lowland rain forest tree species. The complexity of species-specific responses to altered nutrient availability highlights the difficulty in predicting future responses of tropical forest trees to a changing world.
DOIURL [本文引用: 1]
叶片衰老过程中的养分再吸收是植物适应养分贫瘠生境的一种重要策略,一直是生态学领域的研究热点。以亚热带红壤侵蚀区生态恢复先锋树种马尾松为研究对象,分析4种不同恢复水平下马尾松叶片养分含量随叶龄的变化情况及养分再吸收特征。结果表明:(1)叶片中N、P、K含量变化范围分别在(6.10±0.52)—(12.02±0.85)mg/g、(0.17±0.03)—(1.02±0.01)mg/g、(1.58±0.49)—(9.46±0.90)mg/g,随叶龄增长整体呈先增加后降低的趋势,具有一个快速积累期和一个相对漫长衰减期的动态特征,这表明叶龄也是影响叶片养分含量的重要因素;(2)叶片N、P、K含量随生境恢复水平的提高而增加,且N、P含量在除凋落叶外的叶龄阶段均表现出显著正相关,表明叶片中这两种营养元素在动态变化上存在协同性;(3)在叶片N、P、K再吸收效率中,P、K再吸收效率较高,而N相对较低。养分再吸收受生境中营养元素的含量水平、循环方式等因素的综合影响表现出一定选择性,对生境中较为贫瘠或使其生长受到限制的元素具有较高的再吸收效率。这不仅可以减小植物对外源养分的依赖性,同时也维持了体内重要营养元素的平衡。随叶龄增长叶片中N/P逐渐增大,反映出在叶片衰老过程中对限制元素P具有较强的再吸收能力,这种反馈调节提高了马尾松对养分贫瘠环境的适应性。本文的研究结果可为亚热带红壤侵蚀退化区先锋物种在贫瘠生境条件下的养分利用机制与适应对策方面的研究提供理论依据。
DOIURL [本文引用: 1]
叶片衰老过程中的养分再吸收是植物适应养分贫瘠生境的一种重要策略,一直是生态学领域的研究热点。以亚热带红壤侵蚀区生态恢复先锋树种马尾松为研究对象,分析4种不同恢复水平下马尾松叶片养分含量随叶龄的变化情况及养分再吸收特征。结果表明:(1)叶片中N、P、K含量变化范围分别在(6.10±0.52)—(12.02±0.85)mg/g、(0.17±0.03)—(1.02±0.01)mg/g、(1.58±0.49)—(9.46±0.90)mg/g,随叶龄增长整体呈先增加后降低的趋势,具有一个快速积累期和一个相对漫长衰减期的动态特征,这表明叶龄也是影响叶片养分含量的重要因素;(2)叶片N、P、K含量随生境恢复水平的提高而增加,且N、P含量在除凋落叶外的叶龄阶段均表现出显著正相关,表明叶片中这两种营养元素在动态变化上存在协同性;(3)在叶片N、P、K再吸收效率中,P、K再吸收效率较高,而N相对较低。养分再吸收受生境中营养元素的含量水平、循环方式等因素的综合影响表现出一定选择性,对生境中较为贫瘠或使其生长受到限制的元素具有较高的再吸收效率。这不仅可以减小植物对外源养分的依赖性,同时也维持了体内重要营养元素的平衡。随叶龄增长叶片中N/P逐渐增大,反映出在叶片衰老过程中对限制元素P具有较强的再吸收能力,这种反馈调节提高了马尾松对养分贫瘠环境的适应性。本文的研究结果可为亚热带红壤侵蚀退化区先锋物种在贫瘠生境条件下的养分利用机制与适应对策方面的研究提供理论依据。
DOIURLPMID [本文引用: 4]
090004Nutrient resorption is a fundamental process through which plants withdraw nutrients from leaves before abscission. Nutrient resorption patterns have the potential to reflect gradients in plant nutrient limitation and to affect a suite of terrestrial ecosystem functions.090004Here, we used a stoichiometric approach to assess patterns in foliar resorption at a variety of scales, specifically exploring how N : P resorption ratios relate to presumed variation in N and/or P limitation and possible relationships between N : P resorption ratios and soil nutrient availability.090004N : P resorption ratios varied significantly at the global scale, increasing with latitude and decreasing with mean annual temperature and precipitation. In general, tropical sites (absolute latitudes < 2300°26090005) had N : P resorption ratios of < 1, and plants growing on highly weathered tropical soils maintained the lowest N : P resorption ratios. Resorption ratios also varied with forest age along an Amazonian forest regeneration chronosequence and among species in a diverse Costa Rican rain forest.090004These results suggest that variations in N : P resorption stoichiometry offer insight into nutrient cycling and limitation at a variety of spatial scales, complementing other metrics of plant nutrient biogeochemistry. The extent to which the stoichiometric flexibility of resorption will help regulate terrestrial responses to global change merits further investigation.
DOIURLPMID [本文引用: 1]
The ecophysiological linkage of leaf phosphorus (P) to photosynthetic capacity$\left( {A_{\max } } \right)$and to the$A_{\max } $nitrogen relation remains poorly understood. To address this issue we compiled published and unpublished field data for mass-based$A_{\max } $, nitrogen (N) and P (n = 517 observations) from 314 species at 42 sites in 14 countries. Data were from four biomes: arctic, cold temperate, subtropical (including Mediterranean), and tropical. We asked whether plants with low P levels have low$A_{\max } $, a shallower slope of the$A_{\max } $-N relationship, and whether these patterns have a geographic signature. On average, leaf P was substantially lower in the two warmer than in the two colder biomes, with the reverse true for N: P ratios. The evidence indicates that the response of$A_{\max } $to leaf N is constrained by low leaf P. Using a full factorial model for all data,$A_{\max } $was related to leaf N, but not to leaf P on its own, with a significant leaf N leaf P interaction indicating that the response of$A_{\max } $to N increased with increasing leaf P. This was also found in analyses using one value per species per site, or by comparing only angiosperms or only woody plants. Additionally, the slope of the$A_{\max } $-N relationship was higher in the colder arctic and temperate than warmer tropical and subtropical biomes. Sorting data into low, medium, and high leaf P groupings also showed that the$A_{\max } $-N slope increases with leaf P. These analyses support claims that in P-limited ecosystems the$A_{\max } $-N relationship may be constrained by low P, and are consistent with laboratory studies that show P-deficient plants have limited ribulose-1,5-bisphosphate regeneration, a likely mechanism for the P influence upon the$A_{\max } $-N relation.
URL [本文引用: 1]
凋落物分解的快慢和养分释放的速度决定了生态系统中土壤有效养分的供应。探讨全球变化条件下森林生态系统凋落物与土壤养分的变化规律,有利于深入认识凋落物-土壤相互作用的养分调控因素,从而揭示生态系统C、N、P循环。通过模拟氮沉降增加试验,分4个水平处理,分别为0、60、120、240 kg N hm~(-2)a~(-1)。模拟氮沉降13年后,分析了杉木人工林凋落物中不同组分(落叶、落枝、落果)生态化学计量与土壤有效养分(有效氮、碱解氮、速效磷、速效钾)的关系。结果表明:氮沉降(N1、N2和N3)显著提高了落叶和落枝的N含量,平均增幅分别为35.27%和32.21%;高水平氮沉降(N3)处理显著降低了落叶和落枝的C/N,平均降幅分别为25.95%和22.32%,但N3增加了落枝和落果N/P,平均增幅分别为38.4%和31.7%;氮沉降对凋落物各组分的C、P和C/P均影响不显著。氮沉降处理显著增加了土壤NO_3~--N和NH_4~+-N含量,均表现为N3N2N1N0,其中NO_3~--N含量更容易受氮沉降处理的影响,表现为更大的增幅。N2显著增加0—20 cm土层的碱解氮含量,N1显著降低0—20 cm土层的速效钾,但氮沉降对速效磷含量没有影响。凋落物生态化学计量与土壤有效养分之间的Pearson相关和冗余分析(RDA)表明,凋落物生态化学计量与土壤有效养分之间关系紧密,凋落物P含量(蒙特卡罗检验,P=0.018)和C/P比值(P=0.037)对土壤有效养分影响显著。凋落物中C/N比值、C/P比值与土壤有效养分呈显著负相关,其比值越高越不利于土壤有效养分的累积。
URL [本文引用: 1]
凋落物分解的快慢和养分释放的速度决定了生态系统中土壤有效养分的供应。探讨全球变化条件下森林生态系统凋落物与土壤养分的变化规律,有利于深入认识凋落物-土壤相互作用的养分调控因素,从而揭示生态系统C、N、P循环。通过模拟氮沉降增加试验,分4个水平处理,分别为0、60、120、240 kg N hm~(-2)a~(-1)。模拟氮沉降13年后,分析了杉木人工林凋落物中不同组分(落叶、落枝、落果)生态化学计量与土壤有效养分(有效氮、碱解氮、速效磷、速效钾)的关系。结果表明:氮沉降(N1、N2和N3)显著提高了落叶和落枝的N含量,平均增幅分别为35.27%和32.21%;高水平氮沉降(N3)处理显著降低了落叶和落枝的C/N,平均降幅分别为25.95%和22.32%,但N3增加了落枝和落果N/P,平均增幅分别为38.4%和31.7%;氮沉降对凋落物各组分的C、P和C/P均影响不显著。氮沉降处理显著增加了土壤NO_3~--N和NH_4~+-N含量,均表现为N3N2N1N0,其中NO_3~--N含量更容易受氮沉降处理的影响,表现为更大的增幅。N2显著增加0—20 cm土层的碱解氮含量,N1显著降低0—20 cm土层的速效钾,但氮沉降对速效磷含量没有影响。凋落物生态化学计量与土壤有效养分之间的Pearson相关和冗余分析(RDA)表明,凋落物生态化学计量与土壤有效养分之间关系紧密,凋落物P含量(蒙特卡罗检验,P=0.018)和C/P比值(P=0.037)对土壤有效养分影响显著。凋落物中C/N比值、C/P比值与土壤有效养分呈显著负相关,其比值越高越不利于土壤有效养分的累积。
DOIURL [本文引用: 1]
Key message While there is evidence that foliar P concentration controls foliar P resorption efficiency, this is not found for N. Likewise, the foliar N:P ratio affects P, but not N resorption.
DOIURL [本文引用: 1]
1 Plant growth at high-latitude sites is usually strongly nutrient-limited. The increased nutrient availability predicted in response to global warming may affect internal plant nutrient cycling, including nutrient resorption from senescing leaves. 2 The effect of increased N supply (10 g N m-2year-1) on nitrogen and phosphorus resorption efficiency and proficiency in six sub-arctic bog species, belonging to four different growth-forms, was studied in northern Sweden. 3 We hypothesized that while increased N supply would not affect N or P resorption efficiency, it would lead to lower N resorption proficiency (higher N concentrations in leaf litter) and higher P resorption proficiency (lower P concentrations in leaf litter). We also investigated whether the basis on which resorption was expressed (leaf mass, leaf area or unit leaf) influenced the patterns observed. 4 Contrasting with our hypothesis, a general trend of decreased N resorption efficiency occurred in response to increased N supply, but the expected decrease in N resorption proficiency was seen in all species except Betula nana. 5 P resorption efficiency did not change in four species (B. nana, Empetrum hermaphroditum, Eriophorum vaginatum and Rubus chamaemorus) but it decreased in Andromeda polifolia, and increased in Vaccinium uliginosum. P resorption proficiency showed the expected increase in only two species (B. nana and V. uliginosum). 6 Apart from P resorption efficiency, the different calculation methods generally produced similar responses of resorption efficiency and proficiency to N supply. 7 Increased N supply at high-latitude sites clearly leads to more N being returned to the soil through leaf litter production. However, decomposition of such litter will probably become P-limited. 8 Considerable interspecific differences in nutrient resorption proficiency were found, indicating that long-term changes in vegetation composition need to be considered when evaluating plant-mediated effects on ecosystem nutrient cycling in response to increased nutrient supply.
DOIURL [本文引用: 4]
Nutrient resorption in plants influences nutrient availability and cycling and is a key process in biogeochemical models. Improved estimates of resorption parameters are needed for predicting long-term primary productivity and for improving such models. Currently, most models assume a value of 50% resorption for nitrogen (N) and phosphorus (P) and lack resorption data for other nutrients and for specific vegetation types. We provide global estimates of r sorption efficiencies and nutrient concentrations for carbon (C), N, and P and the first global-scale estimates for essential nutrients such as potassium (K), calcium (Ca), and magnesium (Mg). We also examine leaf mass loss during senescence (LML) globally and for different plant types, thus defining a mass loss correction factor (MLCF) needed to quantify unbiased resorption values. We used a global meta-analysis of 86 studies and ~ 1000 data points across climates for green and senesced leaves in six plant types: ferns, forbs, graminoids, conifers, and evergreen and deciduous woody angiosperms. In general, N and P resorption differed significantly from the commonly used global value of 50% (62.1%, 64.9%, respectively; P < 0.05). Ca, C, and Mg showed lower average resorptions of 10.9%, 23.2%, and 28.6%, respectively, while had the highest r sorption, at 70.1%. We also found that resorption of all nutrients except Ca depended on leaf nutrient-status; globally, C, N, P, K, and Mg showed a decrease in resorption with increased nutrient status. On average, global leaf mass loss was 24.2%. Overall, our r sorption data differ substantially from commonly assumed values and should help improve ecological theory and biogeochemical and landsurface models.
URL [本文引用: 2]
人类活动和全球气候变化导致的大气N沉降升高和降水格局的变化,将会影响植物-土壤系统的养分循环。研究了5种荒漠优势短命植物在N添加和增加降水处理下,叶片N、P含量变化及叶片N、P的再吸收特征。结果表明:自然情况下,荒漠植物叶N、P含量相对较高(平均值分别为30.1mg·g-1、3.6mg·g-1),而叶片N∶P很低(6.8~10.5);荒漠短命植物叶N再吸收率(NRE,34.08%)低于P再吸收率(PRE,73.03%);叶片养分含量和再吸收率在物种间的差异极显著;N添加对NRE和PRE没有显著影响;N添加使尖喙牻牛儿苗(Erodium oxyrrhynchum)、条叶庭荠(Alyssum linifolium)和卷果涩荠(Malcolmia scorpioides)N再吸收度(NRP)减小,使丝叶芥(Leptaleum filifolium)和齿稃草(Schismus arabicus)的NRP增加,对P再吸收度(PRP)影响很小。不同含量N添加处理对NRE没有显著影响,但对PRE具有显著影响;增加降水对NRE具有显著影响,使NRE增加(丝叶芥除外),对PRE、NRP和PRP没有显著影响,但PRE和NRP增加,PRP变化很小;N添加和增加降水对NRE没有显著交互作用,对PRE具有显著交互作用,但其变化因物种而异。总之,N输入增加和降水格局的变化,显著改变了荒漠短命植物养分状态及再吸收特征,这将对短命植物群落组成及所处荒漠生态系统养分循环产生重要影响。
URL [本文引用: 2]
人类活动和全球气候变化导致的大气N沉降升高和降水格局的变化,将会影响植物-土壤系统的养分循环。研究了5种荒漠优势短命植物在N添加和增加降水处理下,叶片N、P含量变化及叶片N、P的再吸收特征。结果表明:自然情况下,荒漠植物叶N、P含量相对较高(平均值分别为30.1mg·g-1、3.6mg·g-1),而叶片N∶P很低(6.8~10.5);荒漠短命植物叶N再吸收率(NRE,34.08%)低于P再吸收率(PRE,73.03%);叶片养分含量和再吸收率在物种间的差异极显著;N添加对NRE和PRE没有显著影响;N添加使尖喙牻牛儿苗(Erodium oxyrrhynchum)、条叶庭荠(Alyssum linifolium)和卷果涩荠(Malcolmia scorpioides)N再吸收度(NRP)减小,使丝叶芥(Leptaleum filifolium)和齿稃草(Schismus arabicus)的NRP增加,对P再吸收度(PRP)影响很小。不同含量N添加处理对NRE没有显著影响,但对PRE具有显著影响;增加降水对NRE具有显著影响,使NRE增加(丝叶芥除外),对PRE、NRP和PRP没有显著影响,但PRE和NRP增加,PRP变化很小;N添加和增加降水对NRE没有显著交互作用,对PRE具有显著交互作用,但其变化因物种而异。总之,N输入增加和降水格局的变化,显著改变了荒漠短命植物养分状态及再吸收特征,这将对短命植物群落组成及所处荒漠生态系统养分循环产生重要影响。
DOIURLPMID [本文引用: 6]
Plant resorption of multiple nutrients during leaf senescence has been established but stoichiometric changes among N, P and K during resorption and after fertilization are poorly understood. We anticipated that increased N supply would lead to further P limitation or co-limitation with N or K [i.e. P-(co)limitation], decrease N resorption and increase P and K resorption, while P and K addition would decrease P and K resorption and increase N resorption. Furthermore, Ca would accumulate while Mg would be resorbed during leaf senescence, irrespective of fertilization. We investigated the effect of N, P and K addition on resorption in two evergreen shrubs ( Chamaedaphne calyculata and Rhododendron groenlandicum ) in a long-term fertilization experiment at Mer Bleue bog, Ontario, Canada. In general, N addition caused further P-(co)limitation, increased P and K resorption efficiency but did not affect N resorption. P and K addition did not shift the system to N limitation and affect K resorption, but reduced P resorption proficiency. C. calyculata resorbed both Ca and Mg while R. groenlandicum resorbed neither. C. calyculata showed a higher resorption than R. groenlandicum , suggesting it is better adapted to nutrient deficiency than R. groenlandicum . Resorption during leaf senescence decreased N:P, N:K and K:P ratios. The limited response of N and K and the response of P resorption to fertilization reflect the stoichiometric coupling of nutrient cycling, which varies among the two shrub species; changes in species composition may affect nutrient cycling in bogs.
DOIURLPMID [本文引用: 1]
Abstract Spatial patterns and temporal trend of nitrogen (N) and phosphorus (P) deposition are important for quantifying their impact on forest carbon (C) uptake. In a first step, we modeled historical and future change in the global distributions of the atmospheric deposition of N and P from the dry and wet deposition of aerosols and gases containing N and P. Future projections were compared between two scenarios with contrasting aerosol emissions. Modeled fields of N and P deposition and P concentration were evaluated using globally distributed in situ measurements. N deposition peaked around 1990 in European forests and around 2010 in East Asian forests, and both increased 7-fold relative to 1850. P deposition peaked around 2010 in South Asian forests and increased 3.5-fold relative to 1850. In a second step, we estimated the change in C storage in forests due to the fertilization by deposited N and P (62C ν dep ), based on the retention of deposited nutrients, their allocation within plants, and C:N and C:P stoichiometry. 62C ν dep for 1997-2013 was estimated to be 0.27 ± 0.13 Pg C yr -1 from N and 0.054 ± 0.10 Pg C yr -1 from P, contributing 9% and 2% of the terrestrial C sink, respectively. Sensitivity tests show that uncertainty of 62C ν dep was larger from P than from N, mainly due to uncertainty in the fraction of deposited P that is fixed by soil. 62C P dep was exceeded by 62C N dep over 1960-2007 in a large area of East Asian and West European forests due to a faster growth in N deposition than P. Our results suggest a significant contribution of anthropogenic P deposition to C storage, and additional sources of N are needed to support C storage by P in some Asian tropical forests where the deposition rate increased even faster for P than for N. This article is protected by copyright. All rights reserved. This article is protected by copyright. All rights reserved.
DOIURL [本文引用: 1]
Light availability generally decreases vertically downwards through plant canopies. According to optimisation theory, in order to maximise canopy photosynthesis plants should allocate leaf nitrogen per area (Narea) in parallel with vertical light gradients, and leaf mass per area (LMA) and leaf angles should decrease down through the canopy also. Many species show trends consistent with these predictions, although these are never as steep as predicted. Most studies of canopy gradients in leaf traits have concerned tall herbaceous vegetation or forest trees. But do evergreen species from open habitats also show these patterns? We quantified gradients of light availability, LMA, leaf N and phosphorus (P), and leaf angle along leaf age sequences and vertical canopy profiles, across 28 woody species from open habitats in eastern Australia. The observed trends in LMA, Narea and leaf angle largely conflicted with expectations from canopy optimisation models, whereas trends in leaf P were more consistent with optimal allocation. These discrepancies most likely relate to these species having rather open canopies with quite shallow light gradients, but also suggest that modelling the co-optimisation of resources other than nitrogen is required for understanding plant canopies.
DOIURL [本文引用: 1]
1. Most plants withdraw nutrients from leaves as they age, and redeploy them elsewhere in the plant. The proportion of nutrients resorbed and the residual nutrient concentration in senesced leaves are different but complementary indices of nutrient conservation via this process. A major spectrum of strategic variation runs from plant species with typically long leaf lifespan (LL), high leaf mass per area (LMA), low leaf nutrient concentrations, and low photosynthetic capacity, to species with the opposite characteristics. It is unknown to what extent either facet of resorption covaries with the LL-LMA spectrum. 2. Green-leaf and senesced-leaf N and P concentrations were quantified for 73 evergreen species from four sites in eastern Australia (nutrient-rich and nutrient-poor sites in each of two rainfall zones). Leaf nutrient concentrations in green and senesced leaves were negatively correlated with LL across all species and at most sites, especially if N2-fixing species were excluded from analyses involving leaf N. 3. Proportional resorption did not differ with soil nutrients, as has been found elsewhere, nor was it correlated with LL. Green-leaf and senesced-leaf nutrient concentrations were lower for species on poorer soils. A simple model was described in which the proportion of resorbed vs soil-derived nutrients deployed in new leaves is set by the relative cost of nutrients from the two sources. The model provides a prospective explanation for the observed differences between species from nutrient-rich and nutrient-poor habitats. 4. The results from this study provide support for the argument that selection to minimize nutrient losses has affected the residual nutrient concentration in senesced leaves, rather than proportional resorption per se. Further, variation among species in residual nutrient concentration was correlated with one of the key spectra of strategic variation between plant species, the leaf lifespan-LMA axis of variation.
DOIURLPMID [本文引用: 1]
Recent studies have indicated that tree carbon accumulation in subtropical forests has been negatively affected by global change phenomena such as warming and drought. However, the long-term effect of nitrogen addition on plant carbon storage remains poorly understood in these regions. In this study, we conducted a 10-year field experiment examining the effect of experimental N addition on plant growth and carbon storage in a subtropical Chinese fir forest. The N levels were 0 (control), 60, 120, and 240 g hayr, and the N effects on tree carbon were divided into stand and individual levels. The results indicated that tree carbon storage at the stand scale was not affected by long-term N addition in the subtropical forest. By contrast, significant impacts of different tree size classes on carbon sequestration were found under different N treatments, which indicated that the amount of plant carbon sequestration was significantly enhanced with tree size class. Our findings highlight the importance of community structure and growth characteristics in Chinese fir forests, in which individual size but not additional N regulates tree carbon sequestration in this subtropical forest.
DOIURL [本文引用: 1]
长期进化和环境适应导致不同植物或同种植物不同基因型间养分利用效率(NUE)差异明显,研究筛选植物养分高效利用基因型至关重要,其极大的增产潜力可补充代替传统植物栽培方法所需的能源。目前人们对于植物NUE概念的理解存在一定差异,造成众多研究成果缺乏可比性。通过对植物NUE的概念及其描述方式难以统一原因的分析,提出人工林NUE应采用干材生物量与林分养分总量的比值表示。综合评述了植物体内养分高效利用及植物对生长介质中养分高效吸收的生物生理学适应性机制。对养分逆境植物养分高效利用适应性策略的整个过程进行了描述,进一步阐明了Ca^2+在化学通讯机制中的生物功能,指出Ca^2+可能是启动植物养分高效利用挽救机制的主要调控因子,并就该领域今后研究工作的特点作了展望。
DOIURL [本文引用: 1]
长期进化和环境适应导致不同植物或同种植物不同基因型间养分利用效率(NUE)差异明显,研究筛选植物养分高效利用基因型至关重要,其极大的增产潜力可补充代替传统植物栽培方法所需的能源。目前人们对于植物NUE概念的理解存在一定差异,造成众多研究成果缺乏可比性。通过对植物NUE的概念及其描述方式难以统一原因的分析,提出人工林NUE应采用干材生物量与林分养分总量的比值表示。综合评述了植物体内养分高效利用及植物对生长介质中养分高效吸收的生物生理学适应性机制。对养分逆境植物养分高效利用适应性策略的整个过程进行了描述,进一步阐明了Ca^2+在化学通讯机制中的生物功能,指出Ca^2+可能是启动植物养分高效利用挽救机制的主要调控因子,并就该领域今后研究工作的特点作了展望。
DOIURL [本文引用: 1]
石化燃料排放出氮氧化物是酸雨的主要来源之一,此外,氮氧化物导致氮湿沉降量的增长是近些年来引起生态学家关注的又一个环境问题.以漳州市为例,据以往研究结果,对氮氧化物在闽南地区酸雨中有较大贡献的问题,进行了相关的氮湿沉降量监测,发现漳州地区是国内少有的高氮沉降区,且增长迅速,从而提出应采取的污染控制措施,以避免造成生态危害.
DOIURL [本文引用: 1]
石化燃料排放出氮氧化物是酸雨的主要来源之一,此外,氮氧化物导致氮湿沉降量的增长是近些年来引起生态学家关注的又一个环境问题.以漳州市为例,据以往研究结果,对氮氧化物在闽南地区酸雨中有较大贡献的问题,进行了相关的氮湿沉降量监测,发现漳州地区是国内少有的高氮沉降区,且增长迅速,从而提出应采取的污染控制措施,以避免造成生态危害.
DOIURL [本文引用: 5]
Background Nitrogen (N) and phosphorus (P) availabilities affect plant growth and the balance of elements, yet how these plant traits respond to N and P supply remains unclear.
DOIURL [本文引用: 1]
Aim Senesced-leaf litter plays an important role in the functioning of terrestrial ecosystems. While green-leaf nutrients have been reported to be affected by climatic factors at the global scale, the global patterns of senesced-leaf nutrients are not well understood. Location Global. Methods Here, bringing together a global dataset of senesced-leaf N and P spanning 1253 observations and 638 plant species at 365 sites and of associated mean climatic indices, we describe the world-wide trends in senesced-leaf N and P and their stoichiometric ratios. Results Concentration of senesced-leaf N was highest in tropical forests, intermediate in boreal, temperate, and mediterranean forests and grasslands, and lowest in tundra, whereas P concentration was highest in grasslands, lowest in tropical forests and intermediate in other ecosystems. Tropical forests had the highest N : P and C : P ratios in senesced leaves. When all data were pooled, N concentration significantly increased, but senesced-leaf P concentration decreased with increasing mean annual temperature (MAT) and mean annual precipitation (MAP). The N : P and C : P ratios also increased with MAT and MAP, but C : N ratios decreased. Plant functional type (PFT), i.e. life-form (grass, herb, shrub or tree), phylogeny (angiosperm versus gymnosperm) and leaf habit (deciduous versus evergreen), affected senesced-leaf N, P, N : P, C : N and C : P with a ranking of senesced-leaf N from high to low: forbs ≈ shrubs ≈ trees > grasses, while the ranking of P was forbs ≈ shrubs ≈ trees < grasses. The climatic trends of senesced-leaf N and P and their stoichiometric ratios were similar between PFTs. Main conclusions Globally, senesced-leaf N and P concentrations differed among ecosystem types, from tropical forest to tundra. Differences were significantly related to global climate variables such as MAT and MAP and also related to plant functional types. These results at the global scale suggest that nutrient feedback to soil through leaf senescence depends on both the climatic conditions and the plant composition of an ecosystem.
DOIURL [本文引用: 1]
Nitrogen resorption efficiency (NRE) and proficiency of 28 plant species belonging to five different life-forms were studied in a semi-arid region of northern China. NRE in these species ranged from 29.8% to 76.1% and averaged about 48.0%, depending upon the species and the life-form. The pattern of NRE in different life-forms followed the order of herbs>shrubs>trees>graminoids>N fixers. Nitrogen resorption proficiency (NRP) ranged from 8.0 to 20.6 mg g 611, the average value of which was lowest in graminoids and highest in N fixing species. Leaf-level nitrogen use efficiency (NUE) ranged from 48.5 g g 611 to 125.8 g g 611, with the average NUE being lowest in the N fixing species and highest in the graminoids. Our findings show that most of the 28 species examined in this study can be categorized as low N-proficiency plants. The lower nitrogen concentration in living tissues and the greater nitrogen resorption during senescence could have contributed jointly to the leaf-level NUE of the species. It was noted that NRP was negatively correlated to NRE, while a positive correlation between the leaf-level NUE and NRE was found for all the species. We had also found a significant positive relation between NRE and the N concentration in green leaves for all the species pulled together, suggesting that green leaf N content might have partially controlled the leaf N resorption.
DOIURL [本文引用: 3]
Abstract Elemental stoichiometry constitutes an inherent link between biogeochemistry and the structure and processes within food webs, and thus is at the core of ecosystem functioning. Stoichiometry allows for spanning different levels of biological organization, from cellular metabolism to ecosystem structure and nutrient cycling, and is therefore particularly useful for establishing links between different ecosystem compartments. We review elemental carbon66:66nitrogen66:66phosphorus (C:N:P) ratios in terrestrial ecosystems (from vegetation, leaf litter, woody debris, and dead roots, to soil microbes and organic matter). While the stoichiometry of the plant, litter, and soil compartments of ecosystems is well understood, heterotrophic microbial communities, which dominate the soil food web and drive nutrient cycling, have received increasing interest in recent years.This review highlights the effects of resource stoichiometry on soil microorganisms and decomposition, specifically on the structure and function of heterotrophic microbial communities and suggests several general patterns. First, latitudinal gradients of soil and litter stoichiometry are reflected in microbial community structure and function. Second, resource stoichiometry may cause changes in microbial interactions and community dynamics that lead to feedbacks in nutrient availability. Third, global change alters the C:N, C:P, and N:P ratios of primary producers, with repercussions for microbial decomposer communities and critical ecosystem services such as soil fertility. We argue that ecological stoichiometry provides a framework to analyze and predict such global change effects at various scales.
DOIURL [本文引用: 1]
Aims To explore resorption efficiency of nitrogen (NRE) and phosphorus (PRE) of woody plants in relation to soil nutrient availability, climate and evolutionary history, in North China. Methods We measured concentrations of nitrogen ([N]) and phosphorus ([P]) in both full expanded mature green and senescent leaves of the same individuals for 88 woody species from 10 sites of Mt. Dongling, Beijing, China. We built a phylogenetic tree for all these species and compared NRE and PRE among life forms (trees, shrubs and woody lianas) and between functional groups (N-fixers and non-N-fixers). We then explored patterns of NRE and PRE along gradients of mean annual temperature (MAT), soil inorganic N and available P, and phylogeny using a general linear model. Important Findings Mass-based NRE (NREm) and PRE (PREm) averaged 57.4 and 61.4%, respectively, with no significant difference among life forms or functional groups. Neither NREm nor PREm exhibited significant phylogenetic signals, indicating that NREm and PREm were not phylogenetically conserved. NREm was not related to [N] in green leaves; PREm was positively correlated with [P] in green leaves; however, this relationship disappeared for different groups. NREm decreased with [N] in senescent leaves, PREm decreased with [P] in senescent leaves, for all species combined and for trees and shrubs. NREm decreased with soil inorganic N for all species and for shrubs; PREm did not exhibit a significant trend with soil available P for all species or for different plant groups. Neither NREm nor PREm was significantly related to MAT for overall species and for species of different groups.
URL [本文引用: 1]
介绍近年来同化物在库器官中的卸载途径、机制和研究方法的研究进展 ,并简略讨论了该领域研究所面临的问题
URL [本文引用: 1]
介绍近年来同化物在库器官中的卸载途径、机制和研究方法的研究进展 ,并简略讨论了该领域研究所面临的问题
DOIURL [本文引用: 1]
为了解生态恢复对侵蚀红壤恢复的马尾松林叶片碳氮磷化学计量及氮磷养分转移的影响,在福建省长汀县河田镇典型侵蚀红壤区选取恢复13、30、33a的马尾松林为研究对象,并以未治理侵蚀地(CK1)和次生林(CK2)分别作为恢复前和恢复后的对照,通过测定马尾松叶片的碳、氮、磷含量,计算其计量比,内稳性指数和氮磷转移率,分析了侵蚀红壤生长的马尾松养分限制与养分转移的关系。结果表明:在侵蚀红壤恢复过程中,马尾松1年龄叶片C、N、P含量及1年龄叶片C:N、C:P、N:P变化较小,这与马尾松较高的内稳性有关(N和P内稳性指数分别为7.57和3.89)。所有实验地马尾松1年龄叶片N:P处于11.0-13.4之间,表明马尾松的生长受N、P共同限制,其中马尾松叶片N转移率显著低于P转移率,这与生态恢复过程中马尾松养分利用效率、生长需求以及土壤养分供应状况有关。1年龄叶片C:N、C:P分别与马尾松N、P转移率成负相关关系,当马尾松叶片C:N、C:P较低时,表明N、P利用效率较低,叶片衰老时更多的N和P被转移利用;反之,则N、P利用效率较高,转移率低。同时,C:N、C:P分别与树高、胸径成显著负相关关系,即马尾松生长对N、P的需求同样会影响化学计量比的变化,从而影响养分转移。虽然侵蚀地生态恢复过程中土壤N、P含量增加,但仍较贫瘠,不足以满足马尾松的生长,马尾松养分转移率较高,因此,为了提高侵蚀地恢复的马尾松林的生产力,建议下一步恢复措施中适当施加N肥和P肥。该研究将侵蚀红壤不同生态恢复年限的马尾松叶片C、N、P化学计量及养分转移结合,有助于全面、系统地揭示生态恢复过程马尾松林的养分循环,对指导侵蚀红壤恢复和提高马尾松生产力具有重要意义。
DOIURL [本文引用: 1]
为了解生态恢复对侵蚀红壤恢复的马尾松林叶片碳氮磷化学计量及氮磷养分转移的影响,在福建省长汀县河田镇典型侵蚀红壤区选取恢复13、30、33a的马尾松林为研究对象,并以未治理侵蚀地(CK1)和次生林(CK2)分别作为恢复前和恢复后的对照,通过测定马尾松叶片的碳、氮、磷含量,计算其计量比,内稳性指数和氮磷转移率,分析了侵蚀红壤生长的马尾松养分限制与养分转移的关系。结果表明:在侵蚀红壤恢复过程中,马尾松1年龄叶片C、N、P含量及1年龄叶片C:N、C:P、N:P变化较小,这与马尾松较高的内稳性有关(N和P内稳性指数分别为7.57和3.89)。所有实验地马尾松1年龄叶片N:P处于11.0-13.4之间,表明马尾松的生长受N、P共同限制,其中马尾松叶片N转移率显著低于P转移率,这与生态恢复过程中马尾松养分利用效率、生长需求以及土壤养分供应状况有关。1年龄叶片C:N、C:P分别与马尾松N、P转移率成负相关关系,当马尾松叶片C:N、C:P较低时,表明N、P利用效率较低,叶片衰老时更多的N和P被转移利用;反之,则N、P利用效率较高,转移率低。同时,C:N、C:P分别与树高、胸径成显著负相关关系,即马尾松生长对N、P的需求同样会影响化学计量比的变化,从而影响养分转移。虽然侵蚀地生态恢复过程中土壤N、P含量增加,但仍较贫瘠,不足以满足马尾松的生长,马尾松养分转移率较高,因此,为了提高侵蚀地恢复的马尾松林的生产力,建议下一步恢复措施中适当施加N肥和P肥。该研究将侵蚀红壤不同生态恢复年限的马尾松叶片C、N、P化学计量及养分转移结合,有助于全面、系统地揭示生态恢复过程马尾松林的养分循环,对指导侵蚀红壤恢复和提高马尾松生产力具有重要意义。
DOIURLMagsci [本文引用: 2]
<p><font face="Verdana">以东北林业大学帽儿山实验林场18年生兴安落叶松人工林为对象,研究连续5年N添加(NH<sub>4</sub>NO<sub>3</sub>,10 g·m<sup>-2</sup>a<sup>-1</sup>)对落叶松各器官N,P,K,Ca和Mg含量及叶片养分再吸收效率的影响。结果表明:N添加对叶片5种元素含量均无显著影响,但可显著提高树根、树枝和树干的N和P含量; N添加能显著提高凋落叶片的N含量,降低凋落叶片的C/N比和叶片N再吸收效率。研究结果反映出研究区兴安落叶松人工林土壤N并不缺乏。N添加导致兴安落叶松对N和P的奢侈吸收,可提高凋落物的分解速率,加快兴安落叶松人工林生态系统的N循环。</font></p>
DOIURLMagsci [本文引用: 2]
<p><font face="Verdana">以东北林业大学帽儿山实验林场18年生兴安落叶松人工林为对象,研究连续5年N添加(NH<sub>4</sub>NO<sub>3</sub>,10 g·m<sup>-2</sup>a<sup>-1</sup>)对落叶松各器官N,P,K,Ca和Mg含量及叶片养分再吸收效率的影响。结果表明:N添加对叶片5种元素含量均无显著影响,但可显著提高树根、树枝和树干的N和P含量; N添加能显著提高凋落叶片的N含量,降低凋落叶片的C/N比和叶片N再吸收效率。研究结果反映出研究区兴安落叶松人工林土壤N并不缺乏。N添加导致兴安落叶松对N和P的奢侈吸收,可提高凋落物的分解速率,加快兴安落叶松人工林生态系统的N循环。</font></p>
Nutrient resorption from senescing leaves of perennials: Are there general patterns?
1
1996
... 养分再吸收(NR)使植物组织中的养分元素(氮(N)、磷(P))在随凋落物进入土壤之前被重新利用, 它缩短了养分元素在植物与环境之间的循环距离(
Patterns and mechanisms of nutrient resorption in plants
9
2015
... NR受植物的生活型、器官、年龄等生理因素的影响.大量研究表明, 植物的不同器官, 如叶(
... )、树皮(
... )、细根(
... ); 落叶植物高于常绿植物(
... 大气氮沉降在全球范围内普遍发生, 全球总活性N释放量从35 Tg·a-1 (1850年)增加到104 Tg·a-1 (1997?2013年), 在未来一段时期可能继续增加(
... 本研究中, 杉木的N含量、P含量、N:P、REP均表现为叶>枝.N含量大小依次为一年生叶>二年生叶>衰老叶>一年生枝>二年生枝>衰老枝, 且N3 > N2 > N1 > N0, 而C:N则与N相反.说明活跃组织具有更高的N含量.究其原因, 第一, 叶是树木的主要光合器官, 与枝相比, 叶含有较大比例的活生物量(
... 杉木叶和枝均呈现REP > REN.这是因为养分投入与器官寿命长短之间的协调关系以及养分在植物体内的停留时间与植物的生产力之间的协调关系密切相关(
... 基本没有变化(
... 叶的养分再吸收途径有两条: 从衰老叶转移到干和(或)根; 从衰老叶或者成熟叶转移到新叶(
Leaf traits and relationships differ with season as well as among species groupings in a managed Southeastern China forest landscape
1
2012
... 本研究中, 杉木的N含量、P含量、N:P、REP均表现为叶>枝.N含量大小依次为一年生叶>二年生叶>衰老叶>一年生枝>二年生枝>衰老枝, 且N3 > N2 > N1 > N0, 而C:N则与N相反.说明活跃组织具有更高的N含量.究其原因, 第一, 叶是树木的主要光合器官, 与枝相比, 叶含有较大比例的活生物量(
Nitrogen and phosphorus additions alter nutrient dynamics but not resorption efficiencies of Chinese fir leaves and twigs differing in age
10
2015
... 大气氮沉降在全球范围内普遍发生, 全球总活性N释放量从35 Tg·a-1 (1850年)增加到104 Tg·a-1 (1997?2013年), 在未来一段时期可能继续增加(
... ;
... 本研究中, 杉木的N含量、P含量、N:P、REP均表现为叶>枝.N含量大小依次为一年生叶>二年生叶>衰老叶>一年生枝>二年生枝>衰老枝, 且N3 > N2 > N1 > N0, 而C:N则与N相反.说明活跃组织具有更高的N含量.究其原因, 第一, 叶是树木的主要光合器官, 与枝相比, 叶含有较大比例的活生物量(
... ).叶和枝逐渐老化, 养分被吸收, 而器官和年龄效应则取决于养分限制的程度(
... ).
... 杉木叶和枝均呈现REP > REN.这是因为养分投入与器官寿命长短之间的协调关系以及养分在植物体内的停留时间与植物的生产力之间的协调关系密切相关(
... 叶的养分再吸收途径有两条: 从衰老叶转移到干和(或)根; 从衰老叶或者成熟叶转移到新叶(
... 经过14年的模拟氮沉降试验, 杉木叶和枝的N含量增加、P含量降低, 这与短期(2-3年)氮沉降的影响趋势一致(
... 溶出量增加, 抑制了杉木对P的吸收.衰老叶N、P含量和衰老枝P含量对氮沉降的响应显著, 说明杉木的N、P养分再吸收度对氮沉降引起的养分利用变化敏感.N添加增加了凋落物的N和P的再吸收能力, 但未产生显著影响(
... 与氮沉降7年相比, 氮沉降14年时杉木叶片的REN更低, 而REP更高.这与凋落物N含量在氮沉降处理时间的延长基础上仍然具有较高浓度, 并且通过落叶返回到土壤的N可能增加有关, 而P含量相反(
林地覆盖经营对雷竹鞭根主要养分内循环的影响
1
2015
... NR受植物的生活型、器官、年龄等生理因素的影响.大量研究表明, 植物的不同器官, 如叶(
林地覆盖经营对雷竹鞭根主要养分内循环的影响
1
2015
... NR受植物的生活型、器官、年龄等生理因素的影响.大量研究表明, 植物的不同器官, 如叶(
Responses of terrestrial ecosystem phosphorus cycling to nitrogen addition: A meta-analysis
1
2017
... 大气氮沉降在全球范围内普遍发生, 全球总活性N释放量从35 Tg·a-1 (1850年)增加到104 Tg·a-1 (1997?2013年), 在未来一段时期可能继续增加(
Low leaf N and P resorption contributes to nutrient limitation in two desert shrubs
2
2006
... NR受植物的生活型、器官、年龄等生理因素的影响.大量研究表明, 植物的不同器官, 如叶(
... 本研究中, 杉木的N含量、P含量、N:P、REP均表现为叶>枝.N含量大小依次为一年生叶>二年生叶>衰老叶>一年生枝>二年生枝>衰老枝, 且N3 > N2 > N1 > N0, 而C:N则与N相反.说明活跃组织具有更高的N含量.究其原因, 第一, 叶是树木的主要光合器官, 与枝相比, 叶含有较大比例的活生物量(
Decline in photosynthetic nitrogen use efficiency with leaf age and nitrogen resorption as determinants of leaf life span
1
2003
... 本研究中, 杉木的N含量、P含量、N:P、REP均表现为叶>枝.N含量大小依次为一年生叶>二年生叶>衰老叶>一年生枝>二年生枝>衰老枝, 且N3 > N2 > N1 > N0, 而C:N则与N相反.说明活跃组织具有更高的N含量.究其原因, 第一, 叶是树木的主要光合器官, 与枝相比, 叶含有较大比例的活生物量(
亚热带杉木(Cunninghamia lanceolata)人工林生长与土壤养分对氮沉降的响应
2
2007
... 经过14年的模拟氮沉降试验, 杉木叶和枝的N含量增加、P含量降低, 这与短期(2-3年)氮沉降的影响趋势一致(
... )(
亚热带杉木(Cunninghamia lanceolata)人工林生长与土壤养分对氮沉降的响应
2
2007
... 经过14年的模拟氮沉降试验, 杉木叶和枝的N含量增加、P含量降低, 这与短期(2-3年)氮沉降的影响趋势一致(
... )(
杉木人工林生态系统的生物地球化学循环II: 氮素沉降动态
1
2000
... 本研究选择福建省三明市沙县官庄林场于1992年种植的杉木人工林为研究对象, 于杉木12年生时开始建立样地, 2004年1月开始模拟氮沉降处理, 一直延续至今.氮沉降量的确定参照了国内外同类研究(
杉木人工林生态系统的生物地球化学循环II: 氮素沉降动态
1
2000
... 本研究选择福建省三明市沙县官庄林场于1992年种植的杉木人工林为研究对象, 于杉木12年生时开始建立样地, 2004年1月开始模拟氮沉降处理, 一直延续至今.氮沉降量的确定参照了国内外同类研究(
火炬松人工林养分体内转移与内循环研究
3
2001
... NR受植物的生活型、器官、年龄等生理因素的影响.大量研究表明, 植物的不同器官, 如叶(
... )、树干(
... 本研究中, 杉木的N含量、P含量、N:P、REP均表现为叶>枝.N含量大小依次为一年生叶>二年生叶>衰老叶>一年生枝>二年生枝>衰老枝, 且N3 > N2 > N1 > N0, 而C:N则与N相反.说明活跃组织具有更高的N含量.究其原因, 第一, 叶是树木的主要光合器官, 与枝相比, 叶含有较大比例的活生物量(
火炬松人工林养分体内转移与内循环研究
3
2001
... NR受植物的生活型、器官、年龄等生理因素的影响.大量研究表明, 植物的不同器官, 如叶(
... )、树干(
... 本研究中, 杉木的N含量、P含量、N:P、REP均表现为叶>枝.N含量大小依次为一年生叶>二年生叶>衰老叶>一年生枝>二年生枝>衰老枝, 且N3 > N2 > N1 > N0, 而C:N则与N相反.说明活跃组织具有更高的N含量.究其原因, 第一, 叶是树木的主要光合器官, 与枝相比, 叶含有较大比例的活生物量(
Nitrogen excess in North American ecosystems: Predisposing factors, ecosystem response, and management strategies
1
1998
... 本研究选择福建省三明市沙县官庄林场于1992年种植的杉木人工林为研究对象, 于杉木12年生时开始建立样地, 2004年1月开始模拟氮沉降处理, 一直延续至今.氮沉降量的确定参照了国内外同类研究(
Retranslocation of foliar nutrients in evergreen tree species planted in a Mediterranean environment
1
2008
... 叶的养分再吸收途径有两条: 从衰老叶转移到干和(或)根; 从衰老叶或者成熟叶转移到新叶(
Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions
1
2008
... 大气氮沉降在全球范围内普遍发生, 全球总活性N释放量从35 Tg·a-1 (1850年)增加到104 Tg·a-1 (1997?2013年), 在未来一段时期可能继续增加(
Impact of nitrogen deposition on nitrogen cycling in forest: A synthesis of NITREX data
1
1998
... 本研究选择福建省三明市沙县官庄林场于1992年种植的杉木人工林为研究对象, 于杉木12年生时开始建立样地, 2004年1月开始模拟氮沉降处理, 一直延续至今.氮沉降量的确定参照了国内外同类研究(
Tree growth and soil acidification in response to 30 years of experimental nitrogen loading on boreal forest
1
2006
... 本研究选择福建省三明市沙县官庄林场于1992年种植的杉木人工林为研究对象, 于杉木12年生时开始建立样地, 2004年1月开始模拟氮沉降处理, 一直延续至今.氮沉降量的确定参照了国内外同类研究(
Foliar nutrient resorption responses of three life-form plants to water and nitrogen additions in a temperate desert
2
2018
... NR受植物的生活型、器官、年龄等生理因素的影响.大量研究表明, 植物的不同器官, 如叶(
... 杉木叶/枝养分再吸收率(RE)计算公式(
Changes in nitrogen resorption traits of six temperate grassland species along a multi-level N addition gradient
2
2008
... 大气氮沉降在全球范围内普遍发生, 全球总活性N释放量从35 Tg·a-1 (1850年)增加到104 Tg·a-1 (1997?2013年), 在未来一段时期可能继续增加(
... )、降低(
Harvest residue management effects on tree growth and ecosystem carbon in a Chinese fir plantation in subtropical China
1
2013
... 杉木(Cunninghamia lanceolata)是典型的南方针叶树种, 因其速生和材质优良的特征在中国亚热带地区已经种植1000多年, 目前杉木林占中国人工林面积18%, 占全球人工林面积的5% (
植物养分重吸收及其影响研究进展
4
2017
... NR受植物的生活型、器官、年龄等生理因素的影响.大量研究表明, 植物的不同器官, 如叶(
... ;
... (
... 叶的养分再吸收途径有两条: 从衰老叶转移到干和(或)根; 从衰老叶或者成熟叶转移到新叶(
植物养分重吸收及其影响研究进展
4
2017
... NR受植物的生活型、器官、年龄等生理因素的影响.大量研究表明, 植物的不同器官, 如叶(
... ;
... (
... 叶的养分再吸收途径有两条: 从衰老叶转移到干和(或)根; 从衰老叶或者成熟叶转移到新叶(
Nutrients in senesced leaves: Keys to the search for potential resorption and resorption proficiency
1
1996
... 叶片养分再吸收度(RP)以衰老叶养分转移后的最低水平浓度来表征(
Resorption efficiency decreases with increasing green leaf nutrients in a global data set
1
2005
... 本研究中, 杉木的N含量、P含量、N:P、REP均表现为叶>枝.N含量大小依次为一年生叶>二年生叶>衰老叶>一年生枝>二年生枝>衰老枝, 且N3 > N2 > N1 > N0, 而C:N则与N相反.说明活跃组织具有更高的N含量.究其原因, 第一, 叶是树木的主要光合器官, 与枝相比, 叶含有较大比例的活生物量(
模拟氮沉降对三种南亚热带树苗生长和光合作用的影响
1
2004
... 本研究选择福建省三明市沙县官庄林场于1992年种植的杉木人工林为研究对象, 于杉木12年生时开始建立样地, 2004年1月开始模拟氮沉降处理, 一直延续至今.氮沉降量的确定参照了国内外同类研究(
模拟氮沉降对三种南亚热带树苗生长和光合作用的影响
1
2004
... 本研究选择福建省三明市沙县官庄林场于1992年种植的杉木人工林为研究对象, 于杉木12年生时开始建立样地, 2004年1月开始模拟氮沉降处理, 一直延续至今.氮沉降量的确定参照了国内外同类研究(
Effects of nitrogen additions on nitrogen resorption and use efficiencies and foliar litterfall of six tree species in a mixed birch and poplar forest, northeastern China
1
2010
... 大气氮沉降在全球范围内普遍发生, 全球总活性N释放量从35 Tg·a-1 (1850年)增加到104 Tg·a-1 (1997?2013年), 在未来一段时期可能继续增加(
不同林龄杉木针叶大量元素转移特征
2
2017
... 本研究中, 杉木的N含量、P含量、N:P、REP均表现为叶>枝.N含量大小依次为一年生叶>二年生叶>衰老叶>一年生枝>二年生枝>衰老枝, 且N3 > N2 > N1 > N0, 而C:N则与N相反.说明活跃组织具有更高的N含量.究其原因, 第一, 叶是树木的主要光合器官, 与枝相比, 叶含有较大比例的活生物量(
... ).有研究表明, 这种机制不随杉木林龄的增加而衰退(
不同林龄杉木针叶大量元素转移特征
2
2017
... 本研究中, 杉木的N含量、P含量、N:P、REP均表现为叶>枝.N含量大小依次为一年生叶>二年生叶>衰老叶>一年生枝>二年生枝>衰老枝, 且N3 > N2 > N1 > N0, 而C:N则与N相反.说明活跃组织具有更高的N含量.究其原因, 第一, 叶是树木的主要光合器官, 与枝相比, 叶含有较大比例的活生物量(
... ).有研究表明, 这种机制不随杉木林龄的增加而衰退(
杉木人工林针叶养分含量对模拟氮沉降增加的响应
1
2008
... 经过14年的模拟氮沉降试验, 杉木叶和枝的N含量增加、P含量降低, 这与短期(2-3年)氮沉降的影响趋势一致(
杉木人工林针叶养分含量对模拟氮沉降增加的响应
1
2008
... 经过14年的模拟氮沉降试验, 杉木叶和枝的N含量增加、P含量降低, 这与短期(2-3年)氮沉降的影响趋势一致(
Enhanced nitrogen deposition over China
2
2013
... 大气氮沉降在全球范围内普遍发生, 全球总活性N释放量从35 Tg·a-1 (1850年)增加到104 Tg·a-1 (1997?2013年), 在未来一段时期可能继续增加(
... (
植物叶片氮磷养分重吸收规律及其调控机制研究进展
1
2018
... 养分再吸收(NR)使植物组织中的养分元素(氮(N)、磷(P))在随凋落物进入土壤之前被重新利用, 它缩短了养分元素在植物与环境之间的循环距离(
植物叶片氮磷养分重吸收规律及其调控机制研究进展
1
2018
... 养分再吸收(NR)使植物组织中的养分元素(氮(N)、磷(P))在随凋落物进入土壤之前被重新利用, 它缩短了养分元素在植物与环境之间的循环距离(
The globalization of nitrogen deposition: Consequences for terrestrial ecosystems
1
2002
... 与氮沉降7年相比, 氮沉降14年时杉木叶片的REN更低, 而REP更高.这与凋落物N含量在氮沉降处理时间的延长基础上仍然具有较高浓度, 并且通过落叶返回到土壤的N可能增加有关, 而P含量相反(
Species-specific responses of foliar nutrients to long-term nitrogen and phosphorus additions in a lowland tropical forest
2
2014
... 杉木叶和枝均呈现REP > REN.这是因为养分投入与器官寿命长短之间的协调关系以及养分在植物体内的停留时间与植物的生产力之间的协调关系密切相关(
... 与氮沉降7年相比, 氮沉降14年时杉木叶片的REN更低, 而REP更高.这与凋落物N含量在氮沉降处理时间的延长基础上仍然具有较高浓度, 并且通过落叶返回到土壤的N可能增加有关, 而P含量相反(
亚热带红壤侵蚀区马尾松针叶养分含量及再吸收特征
1
2016
... 同时, 还分别计算了杉木不同龄级之间的N (P)养分再吸收率, 计算公式(
亚热带红壤侵蚀区马尾松针叶养分含量及再吸收特征
1
2016
... 同时, 还分别计算了杉木不同龄级之间的N (P)养分再吸收率, 计算公式(
Stoichiometric patterns in foliar nutrient resorption across multiple scales
4
2012
... 养分再吸收(NR)使植物组织中的养分元素(氮(N)、磷(P))在随凋落物进入土壤之前被重新利用, 它缩短了养分元素在植物与环境之间的循环距离(
... ).
... NR受植物的生活型、器官、年龄等生理因素的影响.大量研究表明, 植物的不同器官, 如叶(
... 叶的养分再吸收途径有两条: 从衰老叶转移到干和(或)根; 从衰老叶或者成熟叶转移到新叶(
Leaf phosphorus influences the photosynthesis-nitrogen relation: A cross-biome analysis of 314 species
1
2009
... 本研究中, 杉木的N含量、P含量、N:P、REP均表现为叶>枝.N含量大小依次为一年生叶>二年生叶>衰老叶>一年生枝>二年生枝>衰老枝, 且N3 > N2 > N1 > N0, 而C:N则与N相反.说明活跃组织具有更高的N含量.究其原因, 第一, 叶是树木的主要光合器官, 与枝相比, 叶含有较大比例的活生物量(
杉木人工林凋落物生态化学计量与土壤有效养分对长期模拟氮沉降的响应
1
2018
... 与氮沉降7年相比, 氮沉降14年时杉木叶片的REN更低, 而REP更高.这与凋落物N含量在氮沉降处理时间的延长基础上仍然具有较高浓度, 并且通过落叶返回到土壤的N可能增加有关, 而P含量相反(
杉木人工林凋落物生态化学计量与土壤有效养分对长期模拟氮沉降的响应
1
2018
... 与氮沉降7年相比, 氮沉降14年时杉木叶片的REN更低, 而REP更高.这与凋落物N含量在氮沉降处理时间的延长基础上仍然具有较高浓度, 并且通过落叶返回到土壤的N可能增加有关, 而P含量相反(
Foliar P- but not N resorption efficiency depends on the P-concentration and the N:P ratio in trees of temperate forests
1
2018
... 本研究中, 杉木的N含量、P含量、N:P、REP均表现为叶>枝.N含量大小依次为一年生叶>二年生叶>衰老叶>一年生枝>二年生枝>衰老枝, 且N3 > N2 > N1 > N0, 而C:N则与N相反.说明活跃组织具有更高的N含量.究其原因, 第一, 叶是树木的主要光合器官, 与枝相比, 叶含有较大比例的活生物量(
Nitrogen and phosphorus resorption efficiency and proficiency in six sub-arctic bog species after 4 years of nitrogen fertilization
1
2003
... 与氮沉降7年相比, 氮沉降14年时杉木叶片的REN更低, 而REP更高.这与凋落物N含量在氮沉降处理时间的延长基础上仍然具有较高浓度, 并且通过落叶返回到土壤的N可能增加有关, 而P含量相反(
Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants
4
2012
... 养分再吸收(NR)使植物组织中的养分元素(氮(N)、磷(P))在随凋落物进入土壤之前被重新利用, 它缩短了养分元素在植物与环境之间的循环距离(
... 杉木叶和枝均呈现REP > REN.这是因为养分投入与器官寿命长短之间的协调关系以及养分在植物体内的停留时间与植物的生产力之间的协调关系密切相关(
... 叶的养分再吸收途径有两条: 从衰老叶转移到干和(或)根; 从衰老叶或者成熟叶转移到新叶(
... 与氮沉降7年相比, 氮沉降14年时杉木叶片的REN更低, 而REP更高.这与凋落物N含量在氮沉降处理时间的延长基础上仍然具有较高浓度, 并且通过落叶返回到土壤的N可能增加有关, 而P含量相反(
5种荒漠短命植物养分再吸收对水氮添加的响应
2
2016
... 大气氮沉降在全球范围内普遍发生, 全球总活性N释放量从35 Tg·a-1 (1850年)增加到104 Tg·a-1 (1997?2013年), 在未来一段时期可能继续增加(
... ;
5种荒漠短命植物养分再吸收对水氮添加的响应
2
2016
... 大气氮沉降在全球范围内普遍发生, 全球总活性N释放量从35 Tg·a-1 (1850年)增加到104 Tg·a-1 (1997?2013年), 在未来一段时期可能继续增加(
... ;
Nutrient resorption of two evergreen shrubs in response to long-term fertilization in a bog
6
2014
... 大气氮沉降在全球范围内普遍发生, 全球总活性N释放量从35 Tg·a-1 (1850年)增加到104 Tg·a-1 (1997?2013年), 在未来一段时期可能继续增加(
... 也有3种效应: 无反应(
... )、增加(
... 经过14年的模拟氮沉降试验, 杉木叶和枝的N含量增加、P含量降低, 这与短期(2-3年)氮沉降的影响趋势一致(
... 与氮沉降7年相比, 氮沉降14年时杉木叶片的REN更低, 而REP更高.这与凋落物N含量在氮沉降处理时间的延长基础上仍然具有较高浓度, 并且通过落叶返回到土壤的N可能增加有关, 而P含量相反(
... ).高养分再吸收率在营养贫乏的环境中起着至关重要的作用, 在这种环境中, 从土壤中吸取养分会需要更高的能量(
Global forest carbon uptake due to nitrogen and phosphorus deposition from 1850 to 2100
1
2017
... 大气氮沉降在全球范围内普遍发生, 全球总活性N释放量从35 Tg·a-1 (1850年)增加到104 Tg·a-1 (1997?2013年), 在未来一段时期可能继续增加(
Gradients of light availability and leaf traits with leaf age and canopy position in 28 Australian shrubs and trees
1
2006
... 与氮沉降7年相比, 氮沉降14年时杉木叶片的REN更低, 而REP更高.这与凋落物N含量在氮沉降处理时间的延长基础上仍然具有较高浓度, 并且通过落叶返回到土壤的N可能增加有关, 而P含量相反(
Nutrient concentration, resorption and lifespan: Leaf traits of Australian sclerophyll species
1
2003
... 本研究中, 杉木的N含量、P含量、N:P、REP均表现为叶>枝.N含量大小依次为一年生叶>二年生叶>衰老叶>一年生枝>二年生枝>衰老枝, 且N3 > N2 > N1 > N0, 而C:N则与N相反.说明活跃组织具有更高的N含量.究其原因, 第一, 叶是树木的主要光合器官, 与枝相比, 叶含有较大比例的活生物量(
Individual size but not additional nitrogen regulates tree carbon sequestration in a subtropical forest
1
2017
... 大气氮沉降在全球范围内普遍发生, 全球总活性N释放量从35 Tg·a-1 (1850年)增加到104 Tg·a-1 (1997?2013年), 在未来一段时期可能继续增加(
植物养分高效利用机制研究进展
1
2009
... 与氮沉降7年相比, 氮沉降14年时杉木叶片的REN更低, 而REP更高.这与凋落物N含量在氮沉降处理时间的延长基础上仍然具有较高浓度, 并且通过落叶返回到土壤的N可能增加有关, 而P含量相反(
植物养分高效利用机制研究进展
1
2009
... 与氮沉降7年相比, 氮沉降14年时杉木叶片的REN更低, 而REP更高.这与凋落物N含量在氮沉降处理时间的延长基础上仍然具有较高浓度, 并且通过落叶返回到土壤的N可能增加有关, 而P含量相反(
漳州市氮湿沉降量异常的形成及危害
1
2005
... 本研究选择福建省三明市沙县官庄林场于1992年种植的杉木人工林为研究对象, 于杉木12年生时开始建立样地, 2004年1月开始模拟氮沉降处理, 一直延续至今.氮沉降量的确定参照了国内外同类研究(
漳州市氮湿沉降量异常的形成及危害
1
2005
... 本研究选择福建省三明市沙县官庄林场于1992年种植的杉木人工林为研究对象, 于杉木12年生时开始建立样地, 2004年1月开始模拟氮沉降处理, 一直延续至今.氮沉降量的确定参照了国内外同类研究(
Effects of nitrogen and phosphorus supply on growth rate, leaf stoichiometry, and nutrient resorption of Arabidopsis thaliana
5
2014
... NR受植物的生活型、器官、年龄等生理因素的影响.大量研究表明, 植物的不同器官, 如叶(
... 大气氮沉降在全球范围内普遍发生, 全球总活性N释放量从35 Tg·a-1 (1850年)增加到104 Tg·a-1 (1997?2013年), 在未来一段时期可能继续增加(
... ;
... )、降低(
... 叶的养分再吸收途径有两条: 从衰老叶转移到干和(或)根; 从衰老叶或者成熟叶转移到新叶(
Global trends in senesced-leaf nitrogen and phosphorus
1
2009
... NR受植物的生活型、器官、年龄等生理因素的影响.大量研究表明, 植物的不同器官, 如叶(
Nitrogen resorption from senescing leaves in 28 plant species in a semi-arid region of northern China
1
2005
... NR受植物的生活型、器官、年龄等生理因素的影响.大量研究表明, 植物的不同器官, 如叶(
The application of ecological stoichiometry to plant-microbial- soil organic matter transformations
3
2015
... 大气氮沉降在全球范围内普遍发生, 全球总活性N释放量从35 Tg·a-1 (1850年)增加到104 Tg·a-1 (1997?2013年), 在未来一段时期可能继续增加(
... 杉木叶和枝均呈现REP > REN.这是因为养分投入与器官寿命长短之间的协调关系以及养分在植物体内的停留时间与植物的生产力之间的协调关系密切相关(
... 与氮沉降7年相比, 氮沉降14年时杉木叶片的REN更低, 而REP更高.这与凋落物N含量在氮沉降处理时间的延长基础上仍然具有较高浓度, 并且通过落叶返回到土壤的N可能增加有关, 而P含量相反(
Resorption efficiency of leaf nutrients in woody plants on Mt. Dongling of Beijing, North China
1
2014
... 养分再吸收(NR)使植物组织中的养分元素(氮(N)、磷(P))在随凋落物进入土壤之前被重新利用, 它缩短了养分元素在植物与环境之间的循环距离(
光合同化物韧皮部卸载途径和机制
1
2003
... 叶的养分再吸收途径有两条: 从衰老叶转移到干和(或)根; 从衰老叶或者成熟叶转移到新叶(
光合同化物韧皮部卸载途径和机制
1
2003
... 叶的养分再吸收途径有两条: 从衰老叶转移到干和(或)根; 从衰老叶或者成熟叶转移到新叶(
生态恢复对马尾松叶片化学计量及氮磷转移的影响
1
2017
... 同时, 还分别计算了杉木不同龄级之间的N (P)养分再吸收率, 计算公式(
生态恢复对马尾松叶片化学计量及氮磷转移的影响
1
2017
... 同时, 还分别计算了杉木不同龄级之间的N (P)养分再吸收率, 计算公式(
氮添加对兴安落叶松养分分配和再吸收效率的影响
2
2010
... 大气氮沉降在全球范围内普遍发生, 全球总活性N释放量从35 Tg·a-1 (1850年)增加到104 Tg·a-1 (1997?2013年), 在未来一段时期可能继续增加(
... 杉木叶和枝均呈现REP > REN.这是因为养分投入与器官寿命长短之间的协调关系以及养分在植物体内的停留时间与植物的生产力之间的协调关系密切相关(
氮添加对兴安落叶松养分分配和再吸收效率的影响
2
2010
... 大气氮沉降在全球范围内普遍发生, 全球总活性N释放量从35 Tg·a-1 (1850年)增加到104 Tg·a-1 (1997?2013年), 在未来一段时期可能继续增加(
... 杉木叶和枝均呈现REP > REN.这是因为养分投入与器官寿命长短之间的协调关系以及养分在植物体内的停留时间与植物的生产力之间的协调关系密切相关(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}