 ,*中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室, 北京 100101
,*中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室, 北京 100101Effects of the supply levels and ratios of nitrogen and phosphorus on seed traits of Chenopodium glaucum
TIAN Da-Shuan,*Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China通讯作者: E-mail:tiands@igsnrr.ac.cn
收稿日期:2018-02-26修回日期:2018-05-29网络出版日期:2018-09-20
| 基金资助: |
Received:2018-02-26Revised:2018-05-29Online:2018-09-20
| Fund supported: |

摘要
全球氮沉降不仅改变土壤氮和磷的有效性, 同时也改变氮磷比例。氮磷供应量、比例及其交互作用可能会影响植物种子性状。该研究在内蒙古草原基于沙培盆栽实验种植灰绿藜(Chenopodium glaucum), 设置3个氮磷供应量水平和3个氮磷比例的正交实验来探究氮磷供应量、比例及其交互作用对灰绿藜种子性状的影响。结果发现氮磷供应量对种子氮浓度、磷浓度和萌发率影响的相对贡献(15%-24%)大于氮磷比例(3%-7%), 而种子大小只受氮磷比例的影响。同时氮磷供应量和比例之间的交互作用显著影响种子氮浓度和磷浓度。同等氮磷比例情况下, 低量养分供应提高种子氮浓度、磷浓度和萌发率。氮磷比例只有在养分匮乏的环境中才会对种子大小和萌发率产生显著影响。总之, 灰绿藜种子不同性状对氮或磷限制的敏感性不同, 同时种子性状也对养分限制表现出适应性和被动响应。
关键词:
Abstract
Keywords:
PDF (900KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
田大栓. 氮磷供应量及比例对灰绿藜种子性状的影响. 植物生态学报[J], 2018, 42(9): 963-970 DOI:10.17521/cjpe.2018.0085
TIAN Da-Shuan.
自工业革命100多年来, 伴随着愈演愈烈的人类工业和农业活动, 氮沉降已经成为一个严重的全球性环境问题(Liu et al., 2013; Pe?uelas et al., 2013)。氮富集正威胁着生物多样性(Stevens et al., 2004; Bobbink et al., 2010)、深刻影响生态系统结构和功能(Bai et al., 2010; Liu & Greaver, 2010; Lu et al., 2011)。氮沉降在增加土壤氮有效性的同时, 也加剧陆地生态系统磷限制(Li et al., 2016)。这暗含着氮沉降背景下土壤氮磷供应水平改变的同时, 它们之间的比例也在改变。但是, 目前考虑解析氮磷供应量、比例及其交互作用对自然生态系统影响的实验还比较少, 主要集中在对植物生长和营养性状的研究(Gusewell, 2005a; Luo et al., 2016; Zhang et al., 2017), 而对野外植物种子性状影响的研究还很少。
植物功能性状紧密关联生态系统功能(Lavorel & Garnier, 2002; Violle et al., 2007; Cornwell et al., 2008)。尽管前人已经检验了氮磷供应水平、比例及其交互作用对植物生长(Gusewell & Bollens, 2003; Fujita et al., 2010)、资源分配(Luo et al., 2016)、植物营养(Gusewell, 2005b)以及养分回收等(Gusewell, 2005a)的影响。但是, 却忽略了氮磷养分对植物生殖性状的作用。植物营养性状变化只关系到当下生态系统功能的变化, 而生殖性状(如种子)变化会影响到未来植物群落乃至生态系统的动态变化(Leishman, 2001; Walck et al., 2011)。例如, 种子大小和氮浓度可以很好地预测幼苗存活和生长, 它们关系到种群和群落的更新, 特别是在不利的环境中(Milberg & Lamont, 1997; Walters & Reich, 2000; 刘志民等, 2004a; Moles & Westoby, 2004; Naegle et al., 2005)。相比于氮浓度, 种子磷浓度可能对幼苗的生长发育更为重要(Gusewell, 2004; Balestri et al., 2009), 这在农业研究中已经得到广泛证实 (Zhang et al., 1990; Grant et al., 2001; Zhu & Smith, 2001; Nadeem et al., 2011)。并且, 萌发快的种子(特别是一年生植物)产生的幼苗在存活、生长和竞争方面都更具有优势(Aarssen & Burton, 1990; Schmid & Dolt, 1994; Verdu & Traveset, 2005)。种子萌发率也可以反映种子质量, 因为营养缺乏的种子通常不能萌发(Howard & Goldberg, 2001; Navarro & Guitian, 2003; 刘志民等, 2004b)。
前人研究表明土壤氮含量增加会显著影响种子大小(增加或降低)、氮浓度(增加)、磷浓度(降低)、萌发率(降低)和萌发速度(降低)(Fortunel et al., 2009; Manning et al., 2009; Hrdlickova et al., 2011; Hejcman et al., 2012; 胡星云等, 2017; Li et al., 2017)。同时, 土壤磷富集也显著改变种子大小(增加或降低)、磷浓度(增加)、氮浓度(增加)和种子萌发率(降低) (Hrdlickova et al., 2011; Hejcman et al., 2012; 刘庆艳等, 2013; Li et al., 2017)。但是, 目前还很少有研究分析氮磷供应比例对野外植物种子性状的影响(孙志高等, 2017)。因此, 全面解析氮磷供应量、比例及其交互作用对植物种子性状的影响显得尤为必要。
灰绿藜(Chenopodium glaucum)广泛分布在全球温带区域, 同时也是内蒙古草原的一年生先锋物种和资源利用型草本植物(Bai et al., 2010)。采用灰绿藜作为本实验研究对象, 并利用野外沙培盆栽实验, 我们设置3个氮磷供应总量和3个供应比例的正交实验, 旨在区分氮磷供应量、比例及其交互作用对灰绿藜种子性状的影响。拟解决的科学问题如下: 1)氮磷供应量和比例对灰绿藜种子性状影响的相对重要性? 2)氮磷供应量和比例对种子性状的影响是否存在交互作用? 3)在不同的氮磷供应比例或供应量情况下, 种子性状对氮磷供应量或比例的响应有何不同?
1 材料和方法
1.1 沙培盆栽实验
本实验在内蒙古草原生态系统定位研究站实施。实验台站位于内蒙古自治区锡林河流域, 具体地理位置为43.63° N, 116.70° E。实验区域年平均气温0.3 ℃, 月平均气温1月份最低(-21.6 ℃), 7月份最高(19 ℃)。年降水量346.1 mm, 生长季降水(5-8月)占到全年降水量的60%-80%。该区域的植被类型为典型草原, 灰绿藜为实验区域的一年生先锋物种和资源利用型草本植物(Bai et al., 2010)。沙培盆栽实验进行时间为2010年6月18日至9月4日。实验包括9个处理, 采用3个氮磷供应量水平和3个氮磷供应比例的完全正交设计(表1; Gusewell & Bollens, 2003; Gusewell, 2005a), 每个处理20个重复。氮磷供应量通过灰绿藜植物个体生命周期所需要的氮量(44.5 mg)和磷量(3.1 mg)的几何平均数计算得出, 选用几何平均数是基于氮和磷对植物生长影响的乘法效应(multiplicative effect)(Gusewell & Bollens, 2003; Gusewell, 2005a)。所以, 我们选取的氮磷供应量为3.93 mg (低量供应), 11.80 mg (中量供应)和35.40 mg (高量供应)。氮磷供应比例基于野外灰绿藜不同器官(叶片、根系、种子和茎)的N:P范围(4.9至46.2), 因而选取N:P为5、15和45。具体的氮磷供应量和比例见表1。
Table 1
表1
表1所有处理的氮磷供应量
Table 1
| N:P | 氮/磷供应量 N/P supply amount (mg·pot-1) | ||
|---|---|---|---|
| 低量 Low | 中量 Middle | 高量 High | |
| 5 | 8.80/1.76 | 26.55/5.28 | 79.15/15.83 |
| 15 | 15.15/1.01 | 45.75/3.05 | 137.11/9.14 |
| 45 | 26.55/0.59 | 79.15/1.76 | 237.62/5.28 |
新窗口打开|下载CSV
氮肥采用KNO3, 磷肥采用KH2PO4。基于Hoagland营养液配方供应足够并且等量的其他植物所需的必需元素。用KCl来平衡溶液中的K+离子。用稀释的KOH溶液来调整pH值到7左右。每周营养液的供应量通过氮磷供应总量除以灰绿藜生活周期的时间来计算。
灰绿藜种子播种在装有洗干净的沙子的盆里(直径20 cm, 高度25 cm), 每盆10粒种子。每个处理20盆, 9个处理共180盆。种子萌发日期为6月18日, 到植物长到3至5片叶时, 每盆只保留一植株, 挑选标准为3片叶的植物个体, 以尽量减少个体间差异所带来的影响。每周用针管将营养液注入, 每盆100 mL。采用塑料布遮掉自然降雨以保证处理效果, 同时每盆供应充足并且等量的水分以排除水分限制效应。
1.2 种子性状测定
当灰绿藜种子成熟时(9月4日), 对所有植物个体都进行取样。在实验室里把种子从植物个体里分离出来, 并在空气中风干。每株随机选取20粒种子称质量, 用来估算种子大小。种子萌发实验在正常室温下进行(18-20 ℃), 同时接受自然光照(Tripathi & Khan, 1990; Wulff et al., 1999; Defalco et al., 2003)。每株随机选取30粒种子用于测定种子萌发率。种子萌发采用直径12 cm的玻璃培养皿, 里面放2层滤纸, 经常喷蒸馏水以保证种子萌发所需的充足水分。每隔6 h对萌发的种子数目进行记录, 标准是胚根伸出。萌发实验持续7天。萌发速度通过Horak和Wax (1991)的方程计算得出。种子氮浓度采用凯氏定氮法测定。种子先用过硫酸盐氧化, 再用钼酸铵分光光度法测定磷浓度(Liu et al., 2016), 并与已知的参考物质进行比对校正。1.3 数据分析
采用两因素方差分析来比较氮磷供应量、比例以及它们之间的交互作用对种子性状的影响。种子磷浓度数据在方差分析之前进行对数转化以保证数据的正态分布。然后, 采用Duncan多重比较分析相同氮磷供应水平下不同氮磷供应比例对种子性状的影响, 同样也分析相同氮磷供应比例情况下不同氮磷供应量对种子性状的影响。所有的统计分析采用SPSS 22.0软件完成, 所有的图形使用SigmaPlot 12.5软件制作。2 结果和分析
总体来说, 氮磷供应量对种子性状影响的相对贡献(15.13%-23.58%)普遍大于氮磷供应比例(4.41%-7.07%)(表2)。氮磷供应量与比例之间的交互作用显著影响种子氮浓度和磷浓度。Table 2
表2
表2氮磷供应量、比例及其交互作用对种子大小、氮浓度、磷浓度、萌发率和萌发速度的影响
Table 2
| 种子性状 Seed trait | 氮磷供应水平 N and P supply level | 氮磷供应比例 N:P supply ratio | 交互作用 Their interaction |
|---|---|---|---|
| 自由度 d.f. | 2 | 2 | 4 |
| 种子大小 Seed size | 0.74%ns | 4.41%* | 5.38%ns |
| 氮浓度 N concentration | 23.58%*** | 2.90%ns | 13.77%** |
| 磷浓度 P concentration | 23.21%*** | 7.07%** | 17.45%*** |
| 萌发率 Germination rate | 15.13%*** | 5.86%** | 2.79%ns |
| 萌发速度 Germination speed | 0.20%ns | 2.00%ns | 4.07%ns |
新窗口打开|下载CSV
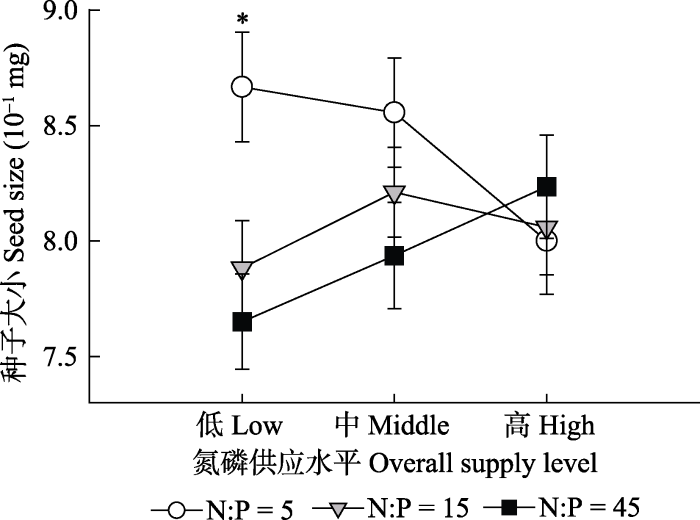
对于形态性状来说, 氮磷供应比例可以解释种子大小变异的4.41%, 而氮磷供应量及它们之间的交互作用影响不显著(表2)。只有在低养分水平下, 随着氮磷比例的提高, 种子显著变小(图1)。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1氮磷供应量或比例对灰绿藜种子大小的影响(平均值±标准误差, n = 20)。*表示相同氮磷供应水平下不同氮磷比例的效应是显著的。
Fig. 1Effects of N and P supply levels or ratios on seed size of Chenopodium glaucum (mean ± SE, n = 20). * represents a significant impact of N:P at the same nutrient supply level.
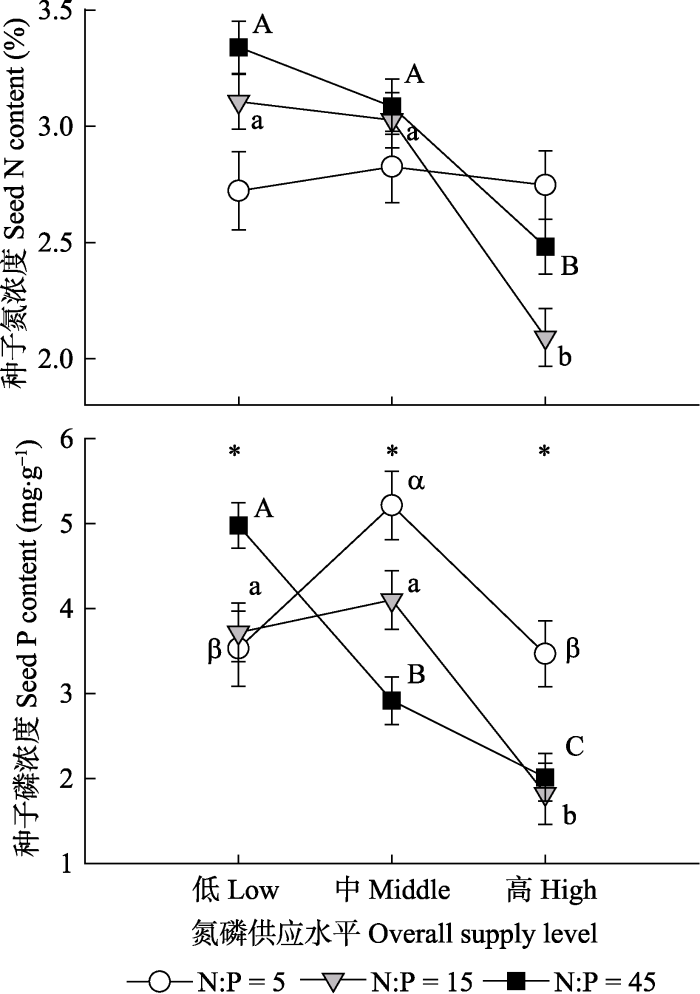
对于养分性状来说, 氮磷供应量分别解释种子氮浓度和磷浓度变异的23.58%和23.21%, 氮磷比例只显著影响种子磷浓度, 它们之间的交互作用分别解释氮浓度和磷浓度变异的13.77%和17.45% (表2)。种子氮浓度在氮磷比例为15或45时, 高量养分供应显著降低种子氮浓度, 而在氮磷比例为5时, 养分供应量没有显著影响(图2)。种子磷浓度在低养分量水平下随着氮磷供应比例的增加而增加, 而在中或高量水平下, 磷浓度对氮磷比例的响应正好相反。氮磷比例为15或45时, 高量养分供应显著降低种子磷浓度, 而在氮磷比例为5时, 磷浓度随着养分供应量没有明显趋势。
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2氮磷供应量或比例对灰绿藜种子氮浓度和磷浓度的影响(平均值±标准误差, n = 20)。*表示相同氮磷供应水平下不同氮磷比例的影响是显著的。不同字母表示相同氮磷供应比例情况下不同氮磷量的效应是显著的。
Fig. 2Effects of nitrogen (N) and phosphorus (P) supply levels or ratios on seed N and P contents of Chenopodium glaucum (mean ± SE, n = 20). * represents a significant effect of N:P at the same nutrient supply level. Different letters indicate a significant effect of nutrient supply level at the same N:P.
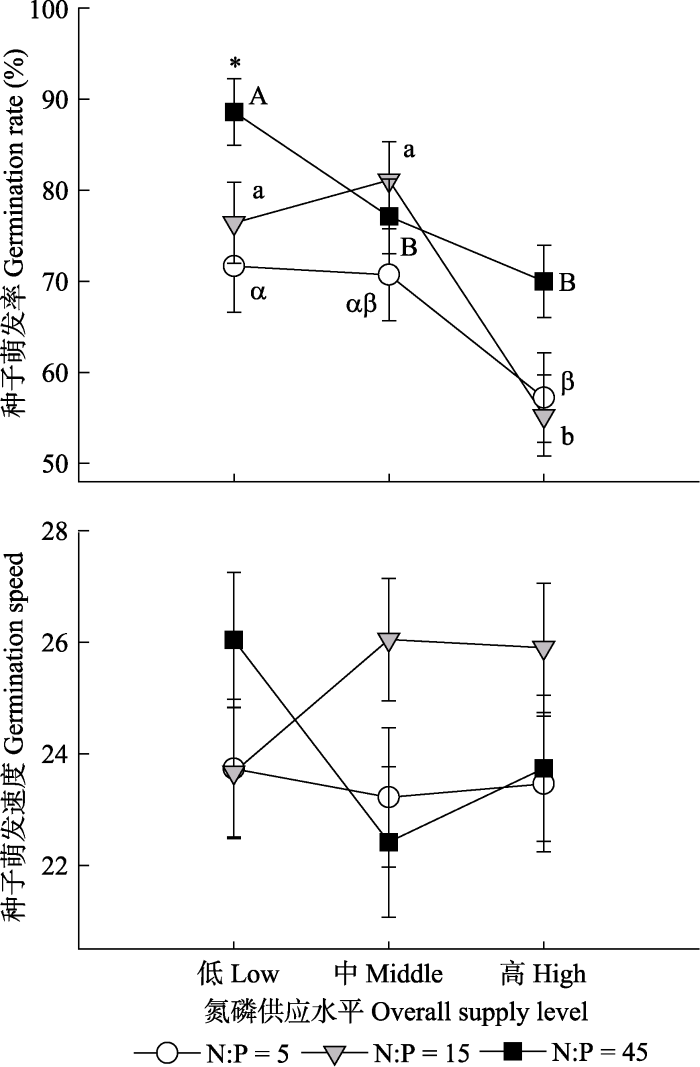
对于萌发性状来说, 氮磷供应量和比例分别解释种子萌发率变异的15.13%和5.86% (表2)。在低量水平, 种子萌发率随着氮磷比例的增加而显著增加(图3), 但是在中量或高量水平下, 氮磷供应比例对萌发率没有显著影响。在氮磷供应比例为5、15或45的情况下, 高量养分供应均显著降低萌发率。种子萌发速度不受任何处理的影响。
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3氮磷供应量或比例对灰绿藜种子萌发率和萌发速度的影响(平均值±标准误差, n = 20)。*表示相同氮磷供应水平下不同氮磷比例的影响是显著的。不同字母表示相同氮磷供应比例情况下不同氮磷供应量的效应是显著的。
Fig. 3Effects of nitrogen (N) and phosphorus (P) supply levels or ratios on seed germination rate and speed of Chenopodium glaucum (mean ± SE, n = 20). * indicates a significant effect of N:P at the same nutrient supply level. Different letters indicate a significant effect of nutrient supply level at the same N:P.
3 讨论
本研究的主要创新性在于系统区分和揭示了氮磷供应量、比例及其交互作用对植物种子性状影响的相对贡献。前人研究主要集中在氮或磷添加水平对种子的影响, 并发现氮或磷供应量显著改变种子性状(Tungate et al., 2006; Breen & Richards, 2008; Fortunel et al., 2009; Hrdlickova et al., 2011; Hejcman et al., 2012)。与以往实验结果类似, 我们也发现氮磷供应量主要影响种子氮浓度、磷浓度和萌发率。但是, 我们发现种子磷浓度和萌发率同时也受到氮磷供应比例的影响, 而种子大小只受氮磷比例的影响, 说明未来土壤氮磷比例变化也会影响种子性状。这在以往相关研究中很少被揭示, 也经常被忽略掉。而且, 我们还发现种子氮浓度和磷浓度受氮磷供应量和比例之间交互作用的影响。这说明即使相同的养分供应水平, 对种子性状的影响效应也会随着氮磷比例的不同而发生变化。或者相同的氮磷供应比例的影响效应也会随不同的养分水平而发生变化。这些研究结果说明全球氮富集背景下自然生态系统中土壤氮磷变化对植物种子性状的影响是复杂的, 不仅仅是土壤氮或磷有效性变化的影响, 同时还掺杂着氮磷比例及其与氮磷供应量之间的交互作用。同时, 种子性状变化紧密关联未来植物种群和群落变化(Galloway, 2001; Leishman, 2001; Naegle et al., 2005; Li et al., 2017), 因此本实验也为我们认识全球养分富集对种群乃至群落的未来动态变化提供新的视角。从种子形态性状来看, 在氮磷比例为5、15或45的情况下, 氮磷供应量对种子大小没有显著影响(图1)。这与前人的一项实验结果类似——Fortunel等(2009)基于地中海生态系统不同演替阶段的18个物种, 利用盆栽实验发现植物种子大小几乎不受养分添加的影响。但是, 我们发现在低量养分水平下, 高氮磷比例显著降低种子大小。这说明灰绿藜植物在贫瘠的环境中, 可能主要是土壤磷的有效性限制其种子大小(Vergeer et al., 2003)。然而, 该显著效应会随着养分供应水平的提高而消失, 指示出氮磷比例只有在养分匮乏的环境中才会对种子大小产生显著作用, 而在养分丰富的环境中对种子的影响不大。
从种子养分性状来看, 在氮磷比例为15或45的情况下, 低量养分供应显著提高种子氮浓度, 尽管在氮磷比例为5的情况下养分供应量的效应不显著。这与以往低养分条件对种子氮浓度影响的实验结果(降低或不影响)(Hrdlickova et al., 2011; Hejcman et al., 2012)不一致。具体原因可能如下: 1)高氮磷供应比例(15和45)比低氮磷比例(5)意味着更少的磷供应, 再随着氮磷供应量的减少就会出现更少的磷输入; 2)在低磷的环境中, 植物可能会主动增加种子氮浓度以增强后代应对不利环境的能力(Violle et al., 2009)。类似地, 氮磷比例为15或45的情况下, 低量养分输入增加种子磷浓度, 而在比例为5的情况下没有明显的增加趋势。这可能也是因为在磷非常匮乏的条件下, 植物主动在种子中累积更多的磷, 来补偿不利环境对植物及其后代的负面影响(Vergeer et al., 2003)。氮磷供应比例对种子氮浓度没有显著影响, 表明灰绿藜种子氮浓度对土壤氮磷有效性之间的相对变化不敏感。有意思的是, 在养分中等或高量水平下, 灰绿藜种子磷浓度随着氮磷供应比例的增加而降低, 说明种子磷浓度可能对土壤磷限制很敏感。但是在养分低量水平下, 种子磷浓度却随着氮磷供应比例的增加而增加, 进一步印证灰绿藜种子主动累积磷养分以应对严重磷缺乏环境这一结论。同样, 前人实验结果也显示种子磷浓度会随着土壤磷有效性的降低而表现出降低(Grant et al., 2001; Ma et al., 2002)或升高(Groom & Lamont, 2010)的响应, 并且相关研究表明同一植物种子的不同性状会对养分添加同时表现出适应性响应(adaptive response)和被动响应(passive response)(Sultan, 1996; Violle et al., 2009)。而我们的实验发现, 即使同一植物的同一种子性状(磷浓度)也会同时表现出对土壤氮磷相对有效性的适应性和被动响应。
从种子萌发性状来看, 我们发现在不同的氮磷比例情况下, 低的氮磷供应量均增加种子萌发率。这与以往实验发现低养分条件促进种子萌发率的结果(Hejcman et al., 2012; Li et al., 2017)一致, 而我们并没有发现氮磷供应量对种子萌发速度的影响(Hrdlickova et al., 2011; Li et al., 2017)。在低养分水平下, 高氮磷比例显著提高种子萌发率, 表明植物在土壤养分匮乏的情况下可能主要是氮有效性限制种子萌发率(Breen & Richards, 2008)。但是, 这些影响效应在养分丰富的环境中不存在, 指示出氮磷比例只有养分匮乏时才会对种子的萌发率产生影响。而灰绿藜种子的萌发速度对氮磷相对有效性不敏感。
总之, 本研究系统阐述氮磷供应量、比例及其交互作用对灰绿藜种子性状影响的相对重要性, 并发现不同种子性状对氮磷绝对有效性和相对有效性的响应不同。不同种子性状受到的氮或磷限制不同, 种子性状同时也对氮或磷限制表现出适应性响应和被动响应。这些实验结果指示出氮或磷富集对植物性状影响的复杂性, 这些复杂效应增加了在内蒙古草原养分富集背景下预测资源利用型草本种群乃至群落未来动态的不确定性。因此, 未来需要更多的氮磷供应量和比例的正交实验来全面解析这些复杂效应。

致谢
感谢乌日娜在野外和室内实验工作中给予的帮助。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 1]

Plants of Senecio vulgaris L. were grown individually within pots to examine effects of maternal soil nutrient level on offspring. Genotypic variability among maternal plants was minimized by using only maternal plants derived from seed of a single inbred line. Significant (P < 0.05) maternal effects of the nutrient gradient were detected at four different levels. Maternal plants grown in lower soil nutrients produced: 1) seeds with lower individual mass; 2) seeds that germinated later; 3) seedling offspring that grew into smaller juvenile plants in nutrient-impoverished soil; and 4) seedling offspring that survived longer in the absence of external nutrients. This last type of maternal effect is reported for the first time in the present study. These data suggest that a compensatory effect of relatively low maternal soil nutrient level may be conferred on the plant's offspring in the form of a "wait and tolerate" strategy through a longer "seedling dormancy" period. This appears to involve a high seedling tolerance threshold level for external nutrient deprivation. To account for these results, we propose two hypotheses based on two different causes of nutrient deprivation, and based on the assumption that if maternal plants experience nutrient deprivation, seedling offspring have a high probability of also experiencing the same cause of nutrient deprivation: 1) Under the "nutrient supply hypothesis," the wait and tolerate strategy in seedling offspring represents an adaptation to selection factors associated with nutrient deprivation resulting from a low nutrient-supplying power of the environment. 2) Under the "nutrient-depletion hypothesis," this strategy represents an adaptation to selection factors associated with nutrient depletion by neighbors.
DOIURL [本文引用: 3]
Nitrogen (N) deposition is widely considered an environmental problem that leads to biodiversity loss and reduced ecosystem resilience; but, N fertilization has also been used as a management tool for enhancing primary production and ground cover, thereby promoting the restoration of degraded lands. However, empirical evaluation of these contrasting impacts is lacking. We tested the dual effects of N enrichment on biodiversity and ecosystem functioning at different organizational levels (i.e., plant species, functional groups, and community) by adding N at 0, 1.75, 5.25, 10.5, 17.5, and 28.0 g N m612 yr611 for four years in two contrasting field sites in Inner Mongolia: an undisturbed mature grassland and a nearby degraded grassland of the same type. N addition had both quantitatively and qualitatively different effects on the two communities. In the mature community, N addition led to a large reduction in species richness, accompanied by increased dominance of early successional annuals and loss of perennial grasses and forbs at all N input rates. In the degraded community, however, N addition increased the productivity and dominance of perennial rhizomatous grasses, with only a slight reduction in species richness and no significant change in annual abundance. The mature grassland was much more sensitive to N-induced changes in community structure, likely as a result of higher soil moisture accentuating limitation by N alone. Our findings suggest that the critical threshold for N-induced species loss to mature Eurasian grasslands is below 1.75 g N m612 yr611, and that changes in aboveground biomass, species richness, and plant functional group composition to both mature and degraded ecosystems saturate at N addition rates of approximately 10.5 g N m612 yr611. This work highlights the tradeoffs that exist in assessing the total impact of N deposition on ecosystem function.
DOIURL [本文引用: 1]
Seedlings ; seagrass ; elemental analysis ; nutritional contrainsts ; sexual reproduction
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
DOIURLPMID [本文引用: 1]
Worldwide decomposition rates depend both on climate and the legacy of plant functional traits as litter quality. To quantify the degree to which functional differentiation among species affects their litter decomposition rates, we brought together leaf trait and litter mass loss data for 818 species from 66 decomposition experiments on six continents. We show that: (i) the magnitude of species-driven differences is much larger than previously thought and greater than climate-driven variation; (ii) the decomposability of a species' litter is consistently correlated with that species' ecological strategy within different ecosystems globally, representing a new connection between whole plant carbon strategy and biogeochemical cycling. This connection between plant strategies and decomposability is crucial for both understanding vegetation oil feedbacks, and for improving forecasts of the global carbon cycle.
[本文引用: 1]
DOIURL [本文引用: 3]
Species performance depends on ecological strategies, revealed by suites of traits, conferring different relative ecological advantages in different environments. Although current knowledge on plant strategies along successional gradients is derived from studies conducted Here we tested whether allocation strategies and seed traits differ among successional stages and nitrogen levels. To this aim, we assessed biomass and nitrogen allocations and seed traits variations for 18 species, differing in life history and belonging to three stages of a Mediterranean old-field succession. These species were grown as monocultures in an experimental garden under limiting and non-limiting nitrogen supply.Early successional species allocated allometrically more nitrogen and proportionally more biomass to reproduction, and set more seeds than later successional species. Seed mass increased with successional status and was negatively related to seed number. Early successional species thus produced more but less-provisioned seeds, suggesting better colonization abilities. These patterns were not the sole consequence of the replacement of annuals by perennials along the successional gradient, since comparable trends were also observed within each life history. Allocation patterns were generally not altered by nitrogen supply and the higher nitrogen content in vegetative organs of plants grown under high nitrogen supply was not retranslocated from leaves to seeds during seed development.We therefore conclude that differences in plant ecological strategies in species characteristics from contrasting successional stages appear to be intrinsic properties of the studied species, and independent from environmental conditions.
DOIURL [本文引用: 1]
N and P have different eutrophication effects on grassland communities, yet the underlying mechanisms are poorly understood. To examine plant growth in response to the varying (relative) supply of N and P, we conducted a two-year greenhouse experiment. Five grasses and three herbs were grown with three N:P supply ratios at two overall nutrient supply levels. During the first year the plant growth was relatively low at both high and low N:P supply ratios, whereas during the second year the growth was especially low at a high N:P supply ratio. This second-year low growth was attributed to the high root death rate, which was influenced by a high N:P supply ratio rather than by the nutrient supply level. Species responded differently, especially in P uptake and loss at a high N:P supply ratio. Each species seemed to have a different strategy for P limitation, e.g. an efficient P uptake or a high P resorption rate. Species typical of P-limited grasslands had neither better P uptake nor better P retention at a high N:P supply ratio. This study quantitatively demonstrates an increased plant root death triggered by strong P limitation. This finding indicates a possible extra effect of N eutrophication on ecosystem functioning via changed N:P stoichiometry.
[本文引用: 1]
DOIURL [本文引用: 2]
DOIURL [本文引用: 1]
The family Proteaceae dominates the nutrient-poor, Mediterranean-climate floristic regions of southwestern Australia (SWA) and the Cape of South Africa. It is well-recognised that mediterranean Proteaceae have comparatively large seeds that are enriched with phosphorus (P), stored mainly as salts of phytic acid in protein globoids. Seed P can contribute up to 48% of the total aboveground P, with the fraction allocated depending on the species fire response. For SWA species, 70–80% of P allocated to fruiting structures is invested in seeds, compared with 30–75% for Cape species, with SWA species storing on average 4.7 times more P per seed at twice the concentration. When soil P is less limiting for growth, seed P reserves may be less important for seedling establishment, and hence plants there tend to produce smaller seeds with less P. For Australian Hakea and Grevillea species the translocation of P from the fruit wall to the seed occurs in the days/weeks before final fruit dry mass is reached, and accounts for 4–36% of seed P. Seed P content increases with the level of serotiny, though it decreases marginally as a fraction of the total reproductive structure. The greater occurrence of serotiny and higher seed P content within the Proteaceae in SWA supports the notion that SWA soils are more P-impoverished than those of the Cape.
DOIURL [本文引用: 1]
Nitrogen (N) and phosphorus (P) availability limit plant growth in most terrestrial ecosystems. This review examines how variation in the relative availability of N and P, as reflected by N: P ratios of plant biomass, influences vegetation composition and functioning. Plastic responses of plants to N and P supply cause up to 50-fold variation in biomass N: P ratios, associated with differences in root allocation, nutrient uptake, biomass turnover and reproductive output. Optimal N: P ratios - those of plants whose growth is equally limited by N and P - depend on species, growth rate, plant age and plant parts. At vegetation level, N: P ratios <10 and >20 often (not always) correspond to N- and P-limited biomass production, as shown by short-term fertilization experiments; however long-term effects of fertilization or effects on individual species can be different. N: P ratios are on average higher in graminoids than in forbs, and in stress-tolerant species compared with ruderals; they correlate negatively with the maximal relative growth rates of species and with their N-indicator values. At vegetation level, N: P ratios often correlate negatively with biomass production; high N: P ratios promote graminoids and stress tolerators relative to other species, whereas relationships with species richness are not consistent N: P ratios are influenced by global change, increased atmospheric N deposition, and conservation managment.
DOIURL [本文引用: 4]
DOIURL [本文引用: 1]
The biomass production of wetland vegetation can be limited by nitrogen or phosphorus. Some species are most abundant in N-limited vegetation, and others in P-limited vegetation, possibly because growth-related traits of these species respond differently to N versus P supply. Two growth experiments were carried out to examine how various morphological and physiological traits respond to the relative supply of N and P, and whether species from sites with contrasting nutrient availability respond differently. In experiment 1, four Carex species were grown in nutrient solutions at five N:P supply ratios (1.7, 5, 15, 45, 135) combined with two levels of supply (geometric means of N and P supply). In experiment 2, two Carex and two grass species were grown in sand at the same five N:P supply ratios combined with three levels of supply and two light intensities (45% or 5% day-light). After 12-13 weeks of growth, plant biomass, allocation, leaf area, tissue nutrient concentrations and rates and nutrient uptake depended significantly on the N:P supply ratio, but the type and strength of the responses differed among these traits. The P concentration and the N:P ratio of shoots and roots as well as the rates of N and P uptake were mainly determined by the N:P supply ratio; they showed little or no dependence on the supply level and relatively small interspecific variation. By contrast, the N concentration, root mass ratio, leaf dry matter content and specific leaf area were only weakly related to the N:P supply ratio; they mainly depended on plant species and light, and partly on overall nutrient supply. Plant biomass was determined by all factors together. Within a level of light and nutrient supply, biomass was generally maximal (i.e. co-limited by N and P) at a N:P supply ratio of 15 or 45. All species responded in a similar way to the N:P supply ratio. In particular, the grass species Phalaris arundinacea and Molinia caerulea showed no differences in response that could clearly explain why P. arundinacea tends to invade P-rich (N-limited) sites, and M. caerulea P-limited sites. This may be due to the short duration of the experiments, which investigated growth and nutrient acquisition but not nutrient conservation.
DOIURL [本文引用: 3]
Changes in the relative availability of essential nutrients such as nitrogen, phosphorus, and potassium can be associated with changes in the species composition of wetland vegetation, but responsible mechanisms are not always clear. This study examines how the relative supply of N and P influences the composition of mixtures of four herbaceous wetland species ( Carex flava, Centaurea angustifolia, Lycopus europaeus and Selinum carvifolia). Mixtures were grown in sand fertilised with nutrient solutions containing N and P in varying proportions (N:P mass ratios of 1.7–135) and total amounts (‘overall supply’, geometric mean of N and P supply). The total above-ground biomass of the mixtures after 11 weeks of growth depended mainly on overall nutrient supply, but also on the N:P supply ratio; it was maximal at a N:P supply ratio of 15 with high overall nutrient supply, 45 with intermediate, and 135 with low nutrient supply. The relative contributions of the four species to total above-ground biomass as well as the root weight ratio differed significantly among nutrient treatments. Treatment effects mostly remained significant when biomass was used as covariate to account for differences in productivity. With increasing N:P supply ratio, the contribution of C. flava increased, while the contribution of L. europaeus and the root weight ratio tended to decrease. These differences were unrelated to interspecific differences in biomass N and P concentrations since L. europaeus had lower concentrations of both N and P than C. flava. Differences in the rates of N and P uptake were apparently most important in this short-term experiment and probably due to differences in biomass allocation to roots. The results also show that the effects of N:P supply ratios should always be investigated at several levels of overall nutrient supply since the two factors interact with each other. Ver01nderungen der relativen Verfügbarkeit wichtiger N01hrstoffe wie Stickstoff, Phosphor und Kalium in Feuchtgebieten k02nnen mit Ver01nderungen der Artenzusammensetzung der Vegetation verbunden sein, doch die verantwortlichen Mechanismen sind nicht immer offensichtlich. Hier wurde untersucht, wie das Verh01ltnis von N- zu P-Versorgung die Zusammensetzung (Biomasseanteile) von Mischungen aus vier Feuchtgebietsarten ( Carex flava, Centaurea angustifolia, Lycopus europaeus und Selinum carvifolia) beeinflusst. Die Mischungen wurden in Quarzsand kultiviert und mit N und P in unterschiedlichen Verh01ltnissen (Massenverh01ltnisse von 1.7–135) und Gesamtmengen (geometrisches Mittel von N- und P-Menge) versorgt. Die gesamte oberirdische Biomasse nach elf-w02chigem Wachstum hing haupts01chlich von der Gesamtversorgung ab, aber auch vom N:P-Verh01ltnis; sie war am h02chsten wenn das N:P-Verh01ltnis bei hoher Gesamtversorgung 15 war, bei mittlerer Gesamtversorgung 45 und bei niedriger Gesamtversorgung 135. Die Biomasseanteile der vier Arten sowie der Wurzelanteil hingen von den Behandlungen ab. Die Behandlungseffekte blieben zum gr0208ten Teil signifikant wenn die Biomasse als Kovariable verwendet wurde, um den Einfluss von Produktivit01tsunterschieden auszuschlie08en. Mit zunehmendem N:P-Verh01ltnis nahm der Biomasseanteil von C. flava zu, w01hrend der Anteil von L. europaeus und der Wurzelanteil abnahmen. Diese Verschiebungen hingen nicht mit unterschiedlichen N01hrstoffkonzentrationen in der Biomasse zusammen, da sowohl die N- als auch die P-Konzentration bei L. europaeus niedriger war als bei C. flava. Demnach waren Unterschiede bei der N- und P-Aufnahme entscheidend, und diese lagen vermutlich an unterschiedlichen Wurzelanteilen. Die Ergebnisse zeigen auch, dass der Einfluss der relativen Verfügbarkeit von N und P stets bei mehreren Gesamtgaben untersucht werden sollte, da die Wirkung der zwei Faktoren voneinander abh01ngt.
DOIURL [本文引用: 5]
Hejcman M, K00i0802álová V, 09ervená K, Hrdli00ková J & Pavl01 V (2012). Effect of nitrogen, phosphorus and potassium availability on mother plant size, seed production and germination ability of Rumex crispus. Weed Research52, 260–268.SummaryRumex crispus is believed to be a highly nutrient-demanding weedy species that spreads mainly by seeds. However, the effects of nutrient availability on its performance and seed production have never been fully investigated. In this study, we investigated how plant size, seed production and germination were affected by the supply of N, P and K. In May 2008, a pot N, P and K fertiliser experiment was established in Prague (Czech Republic). During 2009, plant growth data were collected, and fully ripe seeds were tested for germination and N, P and K concentrations. Rumex crispus showed high phenotypic plasticity in plant height, number of leaves, leaf length and number of stems per plant in relation to N, P and K supply. Seed production per plant ranged from <2000 in the control and low P treatment up to almost 25 000 in the high NPK treatment. More than 16 000 seeds were produced per plant in all treatments where N and P were applied together. To produce a high quantity of rapidly germinating seeds, R. crispus requires a balanced N, P and K supply, as a deficiency of P and K together with a high N supply can result in the production of P- and K-deficient seeds (P and K <3 g kg611) with lower germination ability. In central Europe, at least some populations of R. crispus do not flower in the seeding year and are strictly monocarpic. Given the short lifespan and monocarpic character of the species, control is probably most effective if applied at the rosette stage, prior to the production of viable seeds.
DOIURL [本文引用: 1]
Studies were undertaken to determine effects of scarification, temperature, stratification, pH, and osmotic potential on seed germination; to determine the effect of planting depth on emergence; to describe the emergence sequence; and to quantify seedling development of bigroot morningglory. Mechanical and chemical scarification caused increased germination but cool, moist stratification did not break dormancy. Optimum germination occurred at 20 and 25 C and with alternating temperatures of 20/10 and 30/20 C. The optimum pH range for germination was between 6 and 8.5. Increasing the solution osmotic potential decreased germination linearly from 90% at -0.08 MPa to 0% at -0.8 MPa. Plant establishment was best at 2 to 4 cm and decreased at greater seeding depths. Seedling emergence was observed and recorded and found not to fit the classical models of hypogeal or epigeal emergence. Only 3% of 5-day-old seedlings sprouted after being clipped to the root crown. When seedlings were 11 days old at clipping, over 85% of the plants sprouted. These results indicate that bigroot morningglory has the potential to successfully reproduce by seed and that the rapid development of the plant should be considered when designing control programs.
DOIURL [本文引用: 1]
Among plants, differences in competition intensity and in the ranking of competitive ability for traits such as germination, growth, and survival may suggest that the importance of competition for population success varies with the trait measured. If a species is a good competitor when measured by one component of fitness (e.g., seedling growth), but a poor competitor when measured by a different component of fitness (e.g., seedling survival), understanding the relative importance of each component of fitness for persistence in the community is critical to understanding how communities are structured. Using a field and a garden experiment in southeastern Michigan, we generated competitive response hierarchies among 4-8 old-field perennials for five components of fitness (germination, seedling growth, seedling survival, adult growth, and adult survival). We examined how the overall response to neighbors changes among components of fitness, the concordance of hierarchies within and among components of fitness, and the correlations between competitive hierarchies and natural abundance. We found little to no overall effect of neighbors on germination and seedling survival, indicating a high tolerance (= strong response competitive ability) of species for neighbors as measured by these components of fitness. We found a strong effect of neighbors on seedling growth, and on adult survival and growth, indicating poorer response competitive ability overall for these components. Although rankings of competitive ability were concordant among species across all components of fitness, certain demographic parameters were more consistent in their rankings than others. Specifically, competitive response hierarchies based on size were strongly concordant, while those based on survival were not. This suggests that future studies may effectively estimate response competitive ability for growth with a single estimate, but that estimates of response competitive ability for survival at both the seedling and adult stages may be required. Finally, competitive response rankings based on germination and seedling growth were most strongly correlated with abundance, suggesting that these components of fitness more strongly influence success in the community than do seedling survival and adult growth and survival.
DOIURL [本文引用: 5]
Rumex obtusifolius is believed to be a nitrophilous weedy species that spreads mainly by seed. However, the effects of nutrient availability on seed production never have been investigated. In this study, how the amount of seed production per plant, the chemical composition of the seeds, and their size and germination are affected by the supply of N, P, and K was investigated. A pot fertilizer experiment with 10 treatments (combinations of 0, 150 [N1], and 300 [N2] kg02N02ha611, 0, 40 [P1], and 80 [P2] kg02P02ha611, and 0 and 100 [K] kg02K02ha611 applied twice per vegetation season) was carried out in Prague, Czech Republic, in 2008 and 2009. The seeds from plants that were cultivated since the spring were collected each August. The germination of the seeds was determined in long-daylight conditions at a stable temperature of 20°C. The number of seeds per plant ranged from 0 in the P1 and P2 treatments and <200 in the control and K treatments to almost 9000 in the N2P2K treatment. The number of seeds per plant was positively correlated with the number of stems per plant and the number of seeds per stem. The relationship between the amount of seeds per plant and the individual seed weight was positive. The concentration of N, P, and K in the seeds was affected by the treatments. The weight of 1000 seeds ranged from 1.3 to 1.702g. The seed length ranged from 2.11 to 2.3402mm and the seed width varied from 1.26 to 1.4202mm. With the exception of the control (with 94% germination), the amount of seed germination was >98% in all the other treatments in 2008. In 2009, the amount of seed germination was >95% in all the treatments, except N1 and N2, where 93% and 89% germination, respectively, was recorded. The germination rate was positively related to the P concentration in the seeds. The amount of time that was necessary for 50% seed germination ranged from 1.53 to 2.8902days and was not related to the N concentration in the seeds, but it was negatively related to the P and K concentrations. A balanced N, P, and K supply to the mother plant strongly affected the number of produced seeds and their chemical composition and germination ability.
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
This paper examines four key mechanisms of the seed size/number trade-off (SSNT) models to assess their relevance to a general understanding of plant community structure. Mechanism 1 is that large seeds have a greater probability of winning in competition against smaller seeds. I provide interspecific experimental evidence that there is a competitive hierarchy among seedlings based on seed size. Mechanism 2 is that a trade-off exists between the number and size of seeds produced for a given reproductive allocation. Negative correlations between seed size and number were found consistently across a range of species from a range of habitats, from published literature. Mechanism 3, that seedling-seedling competition is an important influence on species composition, was found to exist potentially in a range of environments, including annual-dominated, post-fire and gap-dynamic communities. However, there is little quantitative evidence available and this is likely to be a restrictive mechanism. Mechanism 4, that small seeds are superior colonists due to their greater number, was tested in a field experiment in a calcareous grassland community. No supporting evidence was found, suggesting that the SSNT is not an important determinant of structure in this community. Thus two of the four mechanisms can be considered to hold true generally, while the third mechanism may be valid in particular environments. The fourth mechanism did not apply in the community tested, but could be tested in a wider range of communities.
DOIURLPMID [本文引用: 5]
Plants adapt to environment by plastic growth which will be transferred to offspring through transgenerational effect. Performance and response of maternal and offspring plant will affect population dynamics and community composition. However, it is scarcely understood how maternal nutrient environment affect the performance and response of offspring through transgenerational effect. Here we studied the impacts of nitrogen (N) and phosphorus (P) enrichment on maternal and offspring performances and responses ofStipa kryloviiandArtemisia frigida. Seeds were collected from maternal plant experiencing N or/and P addition for three years in Inner Mongolia grassland. We found that maternal nutrient addition significantly affected seed traits, offspring biomass, and offspring responses ofA. frigida. Maternal N addition significantly affected maternal reproductive biomass, seed traits ofS. kryloii. Maternal P addition ofS. kryloiisignificantly affected seed qualities, seedling biomass and seeding response to N addition. Our results suggested that transgenerational effects of N and P enrichment to the two dominant plant species existed in this ecosystem. Furthermore, the two species exhibited different adaptive strategies to future nutrient addition. These findings indicate that maternal environmental effect should be considered into the model projection of vegetation dynamics in response to ongoing environmental change.
DOIURLPMID [本文引用: 1]
Abstract Nitrogen (N) and phosphorus (P), either individually or in combination, have been demonstrated to limit biomass production in terrestrial ecosystems. Field studies have been extensively synthesized to assess global patterns of N impacts on terrestrial ecosystem processes. However, to our knowledge, no synthesis has been done so far to reveal global patterns of P impacts on terrestrial ecosystems, especially under different nitrogen (N) levels. Here, we conducted a meta-analysis of impacts of P addition, either alone or with N addition, on aboveground (AGB) and belowground biomass production (BGB), plant and soil P concentrations, and N :聽P ratio in terrestrial ecosystems. Overall, our meta-analysis quantitatively confirmed existing notions: (i) colimitation of N and P on biomass production and (ii) more P limitation in tropical forest than other ecosystems. More importantly, our analysis revealed new findings: (i) P limitation on biomass production was aggravated by N enrichment and (ii) plant P concentration was a better indicator of P limitation than soil P availability. Specifically, P addition increased AGB and BGB by 34% and 13%, respectively. The effect size of P addition on biomass production was larger in tropical forest than grassland, wetland, and tundra and varied with P fertilizer forms, P addition rates, or experimental durations. The P-induced increase in biomass production and plant P concentration was larger under elevated than ambient N. Our findings suggest that the global limitation of P on biomass production will become severer under increasing N fertilizer and deposition in the future.
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
通过温室萌发实验,研究磷养分输入对兴凯湖狭叶甜茅(Glyceria spiculosa)沼泽、槽鳞扁莎(Pycreus korshinskyi)沼泽、恢复沼泽、大豆地和水稻田土壤种子库种子萌发的影响。结果表明,磷养分输入并没有显著影响狭叶甜茅沼泽、槽鳞扁莎沼泽土壤种子库种子萌发的物种组成,但是随着磷养分由低—中—高浓度的输入,两者的种子库萌发幼苗数呈现减小—增加—显著减小的变化。其中,8g/m3磷养分输入使二者的种子库种子萌发幼苗数量显著减少(p0.05)。磷养分输入对恢复沼泽和农田的土壤种子库种子萌发影响不明显。2g/m3磷养分输入显著减少了狭叶甜茅沼泽土壤种子库中野稗(Echinochloa crusgalli)的幼苗萌发数量,而磷养分浓度达到8g/m3时,槽鳞扁莎和三裂慈菇(Sagittaria trifolia)的幼苗萌发数量显著减少。磷养分输入可能会抑制狭叶甜茅和槽鳞扁莎沼泽物种的种子萌发数量,进而改变沼泽植物群落结构。应注意控制沼泽中营养物质的输入,以维持沼泽植物群落结构和生态系统的稳定。
DOIURL [本文引用: 2]
通过温室萌发实验,研究磷养分输入对兴凯湖狭叶甜茅(Glyceria spiculosa)沼泽、槽鳞扁莎(Pycreus korshinskyi)沼泽、恢复沼泽、大豆地和水稻田土壤种子库种子萌发的影响。结果表明,磷养分输入并没有显著影响狭叶甜茅沼泽、槽鳞扁莎沼泽土壤种子库种子萌发的物种组成,但是随着磷养分由低—中—高浓度的输入,两者的种子库萌发幼苗数呈现减小—增加—显著减小的变化。其中,8g/m3磷养分输入使二者的种子库种子萌发幼苗数量显著减少(p0.05)。磷养分输入对恢复沼泽和农田的土壤种子库种子萌发影响不明显。2g/m3磷养分输入显著减少了狭叶甜茅沼泽土壤种子库中野稗(Echinochloa crusgalli)的幼苗萌发数量,而磷养分浓度达到8g/m3时,槽鳞扁莎和三裂慈菇(Sagittaria trifolia)的幼苗萌发数量显著减少。磷养分输入可能会抑制狭叶甜茅和槽鳞扁莎沼泽物种的种子萌发数量,进而改变沼泽植物群落结构。应注意控制沼泽中营养物质的输入,以维持沼泽植物群落结构和生态系统的稳定。
DOIURLPMID
China is experiencing intense air pollution caused in large part by anthropogenic emissions of reactive nitrogen(1,2). These emissions result in the deposition of atmospheric nitrogen (N) in terrestrial and aquatic ecosystems, with implications for human and ecosystem health, greenhouse gas balances and biological diversity(1,3-5). However, information on the magnitude and environmental impact of N deposition in China is limited. Here we use nationwide data sets on bulk N deposition, plant foliar N and crop N uptake (from long-term unfertilized soils) to evaluate N deposition dynamics and their effect on ecosystems across China between 1980 and 2010. We find that the average annual bulk deposition of N increased by approximately 8 kilograms of nitrogen per hectare (P < 0.001) between the 1980s (13.2 kilograms of nitrogen per hectare) and the 2000s (21.1 kilograms of nitrogen Per hectare). Nitrogen deposition rates in the industrialized and agriculturally intensified regions of China are as high as the peak levels of deposition in northwestern Europe in the 1980s(6), before the introduction of mitigation measures(7,8). Nitrogen from ammonium (NH4+) is the dominant form of N in bulk deposition, but the rate of increase is largest for deposition of N from nitrate (NO3-), in agreement with decreased ratios of NH3 to NOx emissions since 1980. We also find that the impact of N deposition on Chinese ecosystems includes significantly increased plant foliar N concentrations in natural and semi-natural (that is, non-agricultural) ecosystems and increased crop N uptake from long-term-unfertilized crop-lands. China and other economies are facing a continuing challenge to reduce emissions of reactive nitrogen, N deposition and their negative effects on human health and the environment.
DOIURLPMID [本文引用: 1]
Soil total nitrogen (STN) and total phosphorus (STP) are important indicators of soil nutrients and the important indexes of soil fertility and soil quality evaluation. Using geographic information system (GIS) and geostatistics, the spatial heterogeneity distribution ofSTNandSTPin the Yaoxiang watershed in a hilly area of northern China was studied. The results showed that: (1) TheSTNandSTPcontents showed a declining trend with the increase in soil depth; the variation coefficients (Cv) ofSTNandSTPin the 0‐ to 10‐cm soil layer (42.25% and 14.77%, respectively) were higher than in the 10‐ to 30‐cm soil layer (28.77% and 11.60%, respectively). Moreover, theCvofSTNwas higher than that ofSTP. (2) The maximumC0/(C0+C1) ofSTNandSTPin the soil layers was less than 25%, this indicated that a strong spatial distribution autocorrelation existed forSTNandSTP; and theSTPshowed higher intensity and more stable variation than theSTN. (3) From the correlation analysis, we concluded that the topographic indexes such as elevation and slope direction all influenced the spatial distribution ofSTNandSTP(correlation coefficients were 0.688 and 0.518, respectively). (4) The overall distribution ofSTNandSTPin the Yaoxiang watershed decreased from the northwest to the southeast. This variation trend was similar to the watershedDEMtrend and was significantly influenced by vegetation and topographic factors. These results revealed the spatial heterogeneity distribution ofSTNandSTP, and addressed the influences of forest vegetation coverage, elevation, and other topographic factors on the spatial distribution ofSTNandSTPat the watershed scale.
DOIURL [本文引用: 1]
研究了科尔沁沙地 6 9种植物的繁殖体 (30种为种子 ,39种为果实 )重量。结果表明 :1 )传播体为果实的植物可分为 4个组别 (即果实单粒重 0 .1mg、0 .1~ 0 .9999mg、1~ 9.9999mg、1 0~ 99.9999mg) ;黄蒿 (Artemisiasco paria)果实 (0 .0 5 1 7mg)和小香蒲 (Typhaminima)果实 (0 .0 6 82mg)最轻 ,苍耳 (Xanthiumsibiricum)单个果实 (77.894 3mg)最重 ;2 )传播体为种子的植物可分为 3个组别 (即种子单粒重 0 .1~ 0 .9999mg、1~ 9.9999mg、1 0~ 99.9999mg) ;马齿苋 (Portulacaoleracea)种子 (0 .1 5 1 4mg)最轻 ,苦参 (Sophoraflavescens)种子 (4 6 .781 6mg)最重 ;3)黄蒿、马齿苋、轮叶沙参 (Adenophoratetraphylla)、碱地肤 (Kochiasieversiana)、狼尾花 (Lysimachiabarystachys)、灰绿藜 (Chenopodiumglaucum)、刺沙蓬 (Salsolaruthenica)、菟丝子 (Cuscutachinensis)、大籽蒿 (Artemisiasieversiana)、狗尾草 (Setariaviridis)、野古草 (Arundinellahirta)等植物所以广泛分布可能是因为它们繁殖体轻 (1mg)且具有持久土壤种子库 ;4 )流沙上的先锋植物或沙生演替系列前期植物沙蓬 (Agriphyllumsquarrosum)、差巴嘎蒿 (Artemisiahalodendron)、乌丹蒿 (Artemisiawudanica)、狗尾草、雾冰藜 (Bassia
DOIURL [本文引用: 1]
研究了科尔沁沙地 6 9种植物的繁殖体 (30种为种子 ,39种为果实 )重量。结果表明 :1 )传播体为果实的植物可分为 4个组别 (即果实单粒重 0 .1mg、0 .1~ 0 .9999mg、1~ 9.9999mg、1 0~ 99.9999mg) ;黄蒿 (Artemisiasco paria)果实 (0 .0 5 1 7mg)和小香蒲 (Typhaminima)果实 (0 .0 6 82mg)最轻 ,苍耳 (Xanthiumsibiricum)单个果实 (77.894 3mg)最重 ;2 )传播体为种子的植物可分为 3个组别 (即种子单粒重 0 .1~ 0 .9999mg、1~ 9.9999mg、1 0~ 99.9999mg) ;马齿苋 (Portulacaoleracea)种子 (0 .1 5 1 4mg)最轻 ,苦参 (Sophoraflavescens)种子 (4 6 .781 6mg)最重 ;3)黄蒿、马齿苋、轮叶沙参 (Adenophoratetraphylla)、碱地肤 (Kochiasieversiana)、狼尾花 (Lysimachiabarystachys)、灰绿藜 (Chenopodiumglaucum)、刺沙蓬 (Salsolaruthenica)、菟丝子 (Cuscutachinensis)、大籽蒿 (Artemisiasieversiana)、狗尾草 (Setariaviridis)、野古草 (Arundinellahirta)等植物所以广泛分布可能是因为它们繁殖体轻 (1mg)且具有持久土壤种子库 ;4 )流沙上的先锋植物或沙生演替系列前期植物沙蓬 (Agriphyllumsquarrosum)、差巴嘎蒿 (Artemisiahalodendron)、乌丹蒿 (Artemisiawudanica)、狗尾草、雾冰藜 (Bassia
DOIURL [本文引用: 1]
在实验室条件下观测了科尔沁沙地乌兰敖都地区1年生植物当年新采种子的萌发特点.在31种植物中,7种植物包括大籽蒿、虎尾草、冠芒草、刺沙蓬、曼陀罗、龙葵、地锦的发芽率超过80%,9种植物包括苍耳、水稗草、狗尾草、虱子草、沙米、灰绿藜、野西瓜苗、太阳花、鹤虱的发芽率不足10%.1~3d开始发芽的18种植物包括大籽蒿、黄蒿、虎尾草、毛马唐、画眉草、冠芒草、野大豆、黄花草木樨、网果葫芦巴、沙米、雾冰藜、碱地肤、刺沙蓬、苘麻、野西瓜苗、马齿苋、苋菜、地锦.超过10d基本不发芽的5种植物包括苍耳、狗尾草、虱子草、灰绿藜、太阳花.发芽持续期小于10d的12种植物包括毛马唐、水稗草、画眉草、沙米、雾冰藜、苘麻、野西瓜苗、益母草、马齿苋、龙葵、地锦、鹤虱;发芽持续期超过21d的6种植物包括大籽蒿、黄蒿、冠芒草、黄花草木樨、刺沙蓬、碱蓬.高发芽率植物的发芽进程线显示了3种萌发格局:速萌型,包括虎尾草、地锦、曼陀罗、龙葵;缓萌型,包括刺沙蓬;中间型,包括大籽蒿、冠芒草.
DOIURL [本文引用: 1]
在实验室条件下观测了科尔沁沙地乌兰敖都地区1年生植物当年新采种子的萌发特点.在31种植物中,7种植物包括大籽蒿、虎尾草、冠芒草、刺沙蓬、曼陀罗、龙葵、地锦的发芽率超过80%,9种植物包括苍耳、水稗草、狗尾草、虱子草、沙米、灰绿藜、野西瓜苗、太阳花、鹤虱的发芽率不足10%.1~3d开始发芽的18种植物包括大籽蒿、黄蒿、虎尾草、毛马唐、画眉草、冠芒草、野大豆、黄花草木樨、网果葫芦巴、沙米、雾冰藜、碱地肤、刺沙蓬、苘麻、野西瓜苗、马齿苋、苋菜、地锦.超过10d基本不发芽的5种植物包括苍耳、狗尾草、虱子草、灰绿藜、太阳花.发芽持续期小于10d的12种植物包括毛马唐、水稗草、画眉草、沙米、雾冰藜、苘麻、野西瓜苗、益母草、马齿苋、龙葵、地锦、鹤虱;发芽持续期超过21d的6种植物包括大籽蒿、黄蒿、冠芒草、黄花草木樨、刺沙蓬、碱蓬.高发芽率植物的发芽进程线显示了3种萌发格局:速萌型,包括虎尾草、地锦、曼陀罗、龙葵;缓萌型,包括刺沙蓬;中间型,包括大籽蒿、冠芒草.
DOIURLPMID [本文引用: 1]
090004Anthropogenic nitrogen (N) addition may substantially alter the terrestrial N cycle. However, a comprehensive understanding of how the ecosystem N cycle responds to external N input remains elusive.090004Here, we evaluated the central tendencies of the responses of 15 variables associated with the ecosystem N cycle to N addition, using data extracted from 206 peer-reviewed papers.090004Our results showed that the largest changes in the ecosystem N cycle caused by N addition were increases in soil inorganic N leaching (461%), soil NO3090808 concentration (429%), nitrification (154%), nitrous oxide emission (134%), and denitrification (84%). N addition also substantially increased soil NH4+ concentration (47%), and the N content in belowground (53%) and aboveground (44%) plant pools, leaves (24%), litter (24%) and dissolved organic N (21%). Total N content in the organic horizon (6.1%) and mineral soil (6.2%) slightly increased in response to N addition. However, N addition induced a decrease in microbial biomass N by 5.8%.090004The increases in N effluxes caused by N addition were much greater than those in plant and soil pools except soil NO3090808, suggesting a leaky terrestrial N system.
DOIURLPMID [本文引用: 2]
In terrestrial ecosystems, atmospheric nitrogen (N) deposition has greatly increased N availability relative to other elements, particularly phosphorus (P). Alterations in the availability of N relative to P can affect plant growth rate and functional traits, as well as resource allocation to above‐ versus belowground biomass (MAandMB). Biomass allocation among individual plants is broadly size‐dependent, and this can often be described as an allometric relationship betweenMAandMB, as represented by the equationMA=αMBβ, or logMA=02logα02+02βlogMB. Here, we investigated whether the scaling exponent or regression slope may be affected by the N:P supply ratio. We hypothesized that the regression slope betweenMAandMBshould be steeper under a high N:P supply ratio due to P limitation, and shallower under a low N:P supply ratio due to N limitation. To test these hypotheses, we experimentally altered the levels of N, P, and the N:P supply ratio (from 1.7:1 to 135:1) provided to five alpine species representing two functional groups (grasses and composite forbs) under greenhouse conditions; we then measured the effects of these treatments on plant morphology and tissue content (SLA, leaf area, and leaf and root N/P concentrations) and on the scaling relationship betweenMAandMB. Unbalanced N:P supply ratios generally negatively affected plant biomass, leaf area, and tissue nutrient concentration in both grasses and composite forbs. High N:P ratios increased tissue N:P ratios in both functional groups, but more in the two composite forbs than in the grasses. The positive regression slopes between logMAand logMBexhibited by plants raised under a N:P supply ratio of 135:1 were significantly steeper than those observed under the N:P ratio of 1.7:1 and 15:1. Synthesis: Plant biomass allocation is highly plastic in response to variation in the N:P supply ratio. Studies of resource allocation of individual plants should focus on the effects of nutrient ratios as well as the availability of individual elements. The two forb species were more sensitive than grasses to unbalanced N:P supplies. To evaluate the adaptive significance of this plasticity, the effects of unbalanced N:P supply ratio on individual lifetime fitness must be measured.
DOIURL [本文引用: 1]
Narrow-leafed lupin (Lupinus angustifolius L.) is usually grown in sandy, acidic and phosphorus (P) deficient soil with low yield and variable harvest index. This study aimed to examine the effects of varying P supply on lupin growth, seed yield and harvest index. Non-abscission plants (cv. Danja) were grown in Lancelin sand at seven rates of P supply (5, 10, 15, 20, 25, 30 or 40 mg kg-1) in a naturally-lit glasshouse. The rate of leaf emergence, flowering time and flower number were decreased or delayed by low P supply (5, 10 or 15 mg kg-1), with no differences at P rates higher than 20 mg kg-1. High P supply (25, 30 or 40 mg kg-1) increased plant seed yield and harvest index largely by increasing the number of pods and consequently yield on the lateral branches, but had less effect on the number of seeds per pod and seed size. Seed yield and seed P concentration continued to increase up to 40 mg P kg-1but harvest index plateaued at 25 mg P kg-1, indicating that low P supply decreased reproductive growth more than vegetative growth in narrow-leafed lupin.
DOIURL [本文引用: 1]
We tested the hypotheses that increased soil resource availability selects for larger seeded plants by: (1) increasing community seed density and seedling competition, (2) increasing light competition, which favours larger seeded species if their additional seed resources are allocated to shoots. We also tested the hypothesis that plants respond to increased nutrient availability by producing larger seeds. In a controlled environment experiment, we grew monocultures of five species of co-occurring annual plant species at two N addition rates and measured seed size and the number of seeds produced. In a second experiment, we sowed seed of the five species together, manipulated nitrogen (N) addition rate and community seed density and measured species performance. In the first experiment, elevated N addition increased seed size, but only in larger seeded species. In the second experiment, high N addition increased community seed production. However, contrary to our hypothesis small seeded species were selected under high seed density, possibly because they germinated and grew more rapidly thus enabling them to pre-empt limiting resources. Larger seeded species were favoured by high N addition, as hypothesised. However, our data suggest that this was due to their seed reserves boosting survival below the denser canopies of fertile conditions rather than via competitive advantages. Our results point to a largely overlooked role of recruitment in determining community response to fertilisation. Although our results may not generalise to all plant communities they suggest that seed traits play a role in community response to both the direct effect of N addition and the stimulation of seed density caused by increased productivity. These results are also consistent with the view that the advantage of large seed size is not a competitive one, but greater survival in the face of environmental hazards.ZusammenfassungWir untersuchten die Hypothese, dass eine zunehmende Verfügbarkeit von Bodenressourcen zu einer Selektion von Pflanzen mit gr0208eren Samen führt, indem: (1) die Samendichte und damit die Konkurrenz der Keimlinge in der Gemeinschaft erh02ht wird, (2) die Konkurrenz um Licht gr0208er wird, was Arten mit gr0208eren Samen f02rdert, wenn ihre zus01tzlichen Samenressourcen in die Sch02sslinge gehen. Wir untersuchten au08erdem die Hypothese, dass Pflanzen auf eine erh02hte N01hrstoffverfügbarkeit reagieren indem sie gr0208ere Samen produzieren. In einem Experiment unter kontrollierten Umweltbedingungen zogen wir Monokulturen von fünf gemeinsam vorkommenden, annuellen Pflanzenarten bei zwei N Zusatzraten und ma08en die Samengr0208e und die Anzahl der produzierten Samen. In einem zweiten s01ten wir die Samen der fünf Arten gemeinsam und manipulierten die Stickstoff (N) Zusatzrate sowie die Samendichte der Gemeinschaft und ma08en die Artenperformanz. In dem ersten Experiment erh02hte ein h02herer N Zusatz die Samengr0208e, dies jedoch nur bei den Arten mit gr0208eren Samen. Im zweiten Experiment erh02hter ein hoher N Zusatz die Samenproduktion der Gemeinschaft. Im Gegensatz zu unserer Hypothese wurden bei hoher Samendichte Arten mit kleinen Samen selektiert, m02glicherweise weil sie schneller keimten und wuchsen und so in der Lage waren limitierende Ressourcen aufzubrauchen. Arten mit gro08en Samen wurden bei einem hohen N Zusatz gef02rdert, so wie wir vermuteten. Unsere Daten lassen jedoch vermuten, dass dies jedoch eher darauf zurückzuführen war, dass ihre Samenreserven ihr 05berleben unter der dichteren Pflanzendecke bei fruchtbaren Bedingungen erm02glichte, als auf Konkurrenzvorteile. Unsere Ergebnisse weisen auf die weitgehend übersehene Rolle der Rekrutierung bei der Bestimmung der Reaktion einer Gemeinschaft auf Düngung hin. Auch wenn unsere Ergebnisse nicht für alle Pflanzengemeinschaften generalisiert werden k02nnen lassen sie jedoch vermuten, dass die Sameneigenschaften eine Rolle bei der Reaktion einer Gemeinschaft sowohl auf den direkten Effekt eines N Zusatzes als auch auf die Erh02hung der Samendichte, die durch eine erh02hte Produktion verursacht wird, spielen. Diese Ergebnisse stimmen ebenfalls mit der Ansicht überein, dass der Vorteil von gro08en Samen kein Konkurrenzvorteil ist, sondern ein besseres 05berleben im Angesicht von ungünstigen Umweltbedingungen erm02glicht.
DOIURL [本文引用: 1]
We examined the importance of seed size in the early establishment of seedlings of four cotyledonous species restricted to nutrient-impoverished soils in south-western Australia. Experimentally removing cotyledons from seedlings increased mortality and reduced root-penetration, growth rate and final weight of plants after 12 wk, in proportion to seed size. The impact of removal of the cotyledons was least in the smallest-seeded, Eucalyptus loxophleba Benth., grown in the more fertile of two soils, and greatest in the largest-seeded, Hakea psilorrhyncha R. M. Barker, whose growth was otherwise unaffected by soil type. In the smallest-seeded species, cotyledon content of most mineral nutrients increased over time, especially in the more fertile soil, while chlorophyll content showed some decrease by the end of the experiment. In the other species, most nutrients (especially N, P, K and Cu in the less fertile soil) were translocated from the cotyledons to the growing plant, and chlorophyll content declined markedly. The translocated N, P, K, Mg and Cu in these species made up a substantial part of the total plant content by 12 wk after germination, while total Ca and Fe tended to increase in the cotyledons via substantial soil uptake. The results indicate, contrary to reports for some other species, that the cotyledons have an important nutritional function in the early establishment of seedlings in these species. The results also show that, in the larger-seeded species, young seedlings rely to a greater extent on nutrient support from the cotyledons than from the soil. We propose that large seeds might be an adaptation for enhancing establishment in nutrient-poor soils.
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
Adequate phosphorus (P) nutrition during early stages is critical for maize growth. Our objective was to evaluate the relative contribution of seed P reserves and exogenous P to maize nutrition during early growth stages. Seedlings were grown with labeled nutrient solution ( 32 P). Seedlings were harvested periodically over the course of the three-week study. Initially, 87% and 77% of the total C and N in seeds were located in the endosperm, whereas 86% of seed P was located in the scutellum as phytate. Up to the 7th day after sowing, 96% of phytate was hydrolyzed. Hydrolyzed forms of P were temporarily stored in the seed before being translocated to growing organs, suggesting that the hydrolysis of phytate was not a limiting step for P supply to seedlings. Significant P uptake by roots was observed from the 5th day after sowing on. Both sources of P supplied roots and leaves, with a slightly higher proportion of P from seed reserves going to leaves rather than to roots. Of total seed P, 60% and 92% was exported towards newly growing seedlings till 7th and 17th days after sowing and ceased to be a significant source of P for growth thereafter. We conclude that although both P supply processes overlap in time, seed P was the main P source during early growth stages.
DOIURL [本文引用: 2]
In low fertility conditions, germinating seedlings are heavily dependent upon N from seed reserves for growth and development. Experiments were conducted to examine the influence of seed N content and mobilization on leaf development and the ability to take up nitrate and up-regulate leaf growth processes during recovery from the period of N deprivation. Genetically homozygous soybean lines with a range of seed N contents were grown in hydroponics culture without an external N source. Microscopic analyses of the apical meristem and plant exposure to labeled 15 N-nitrate allowed precise descriptions of changes in leaf initiation and nitrate uptake during recovery. In seedlings grown without N, inhibitions of leaf expansion and leaf initiation were detectible in the 2nd week after germination, and both were inversely related to the amount of N released from the seed. The S/R ratio decreased, indicating early N stress, but the S/R adjustments were not proportional to the degree of N stress as occurs with older plants, suggesting limitations in C availability. When external N was supplied to plants after 15 or 23 days of N deprivation, the induction period for nitrate uptake was longer than that normally observed with N-replete seedlings, and rates of growth during recovery from the N stress were correlated with initial seed N contents. The inhibition of individual leaf expansion was released only if leaves were at an early developmental stage. Leaf initiation at the apical meristem immediately responded to the presence of external nitrate, with initiation rates approaching those for +N controls. The results indicate that seedling vigor in infertile conditions and the ability to respond to available N are strongly influenced by seed resources.
DOIURL [本文引用: 1]
We examined germination and seedling survival of Petrocoptis grandiflora and Petrocoptis viscosa (Caryophyllaceae), two narrow endemic species from the northwest Iberian Peninsula. The experiments were carried out with seeds of three of eight populations of P. grandiflora and one of the three sole populations of P. viscosa. Under natural conditions, both P. grandiflora and P. viscosa produce very large numbers of seeds. However, the specific microhabitat of these species (cracks and crevices of limestone rockfaces) has very marked effects on seed germination, and subsequent seedling survival. In the present study, we examined the effects of light, cold treatment and seed weight on germination capacity. In the case of P. grandiflora, we also compared the germination of seeds of the different populations. In addition, we assessed seedling survival over a 1-year period. Seeds maintained in darkness showed higher germination percentages than seeds maintained with a 12:12 h photoperiod. The application of a short period of cold prior to germination had no significant effect in either species. In the case of P. grandiflora, population of origin had a significant effect on germination percentage. Of all the factors considered, seed weight was the most important source of variability, both in seed germination and subsequent seedling survival. In both species in the natural habitat, less than 10% of germinated seeds survived by the end of the year. Seedling survival was affected by microhabitat. Seedlings in non-rockface soil microhabitats were more likely to suffer herbivory or interspecific competition than seedlings in crevices in the rockface.
DOIURLPMID [本文引用: 1]
Abstract The availability of carbon from rising atmospheric carbon dioxide levels and of nitrogen from various human-induced inputs to ecosystems is continuously increasing; however, these increases are not paralleled by a similar increase in phosphorus inputs. The inexorable change in the stoichiometry of carbon and nitrogen relative to phosphorus has no equivalent in Earth's history. Here we report the profound and yet uncertain consequences of the human imprint on the phosphorus cycle and nitrogen:phosphorus stoichiometry for the structure, functioning and diversity of terrestrial and aquatic organisms and ecosystems. A mass balance approach is used to show that limited phosphorus and nitrogen availability are likely to jointly reduce future carbon storage by natural ecosystems during this century. Further, if phosphorus fertilizers cannot be made increasingly accessible, the crop yields projections of the Millennium Ecosystem Assessment imply an increase of the nutrient deficit in developing regions.
DOIURLPMID [本文引用: 1]
To predict the possible evolutionary response of a plant species to a new environment, it is necessary to separate genetic from environmental sources of phenotypic variation. In a case study of the invader Solidago altissima, the influences of several kinds of parental effects and of direct inheritance and environment on offspring phenotype were separated. Fifteen genotypes were crossed in three 5 x 5 diallels excluding selfs. Clonal replicates of the parental genotypes were grown in two environments such that each diallel could be made with maternal/paternal plants from sand/sand, sand/soil, soil/sand, and soil/soil. In a first experiment (1989) offspring were raised in the experimental garden and in a second experiment (1990) in the glasshouse. Parent plants growing in sand invested less biomass in inflorescences but produced larger seeds than parent plants growing in soil. In the garden experiment, phenotypic variation among offspring was greatly influenced by environmental heterogeneity. Direct genetic variation (within diallels) was found only for leaf characters and total leaf mass. Germination probability and early seedling mass were significantly affected by phenotypic differences among maternal plants because of genotype (genetic maternal effects) and soil environment (general environmental maternal effects). Seeds from maternal plants in sand germinated better and produced bigger seedlings than seeds from maternal plants in soil. They also grew taller with time, probably because competition accentuated the initial differences. Height growth and stem mass at harvest (an integrated account of individual growth history) of offspring varied significantly among crosses within parental combinations (specific environmental maternal effects). In the glasshouse experiment, the influence of environmental heterogeneity and competition could be kept low. Except for early characters, the influence of direct genetic variation was large but again leaf characters (= basic module morphology) seemed to be under stricter genetic control than did size characters. Genetic maternal effects, general environmental maternal effects, and specific environmental maternal effects dominated in early characters. The maternal effects were exerted both via seed mass and directly on characters of young offspring. Persistent effects of the general paternal environment (general environmental paternal effects) were found for leaf length and stem and leaf mass at harvest. They were opposite in direction to the general environmental maternal effects, that is the same genotypes produced "better mothers" in sand but "better fathers" in soil. The general environmental paternal effects must have been due to differences in pollen quality, resulting from pollen selection within the male parent or leading to pre- or postzygotic selection within the female parent. The ranking of crosses according to mean offspring phenotypes was different in the two experiments, suggesting strong interaction of the observed effects with the environment. The correlation structure among characters changed less between experiments than did the pattern of variation of single characters, but under the competitive conditions in the garden plant height seemed to be more directly related to fitness than in the glasshouse. Reduced competition could also explain why maternal effects were less persistent in the glasshouse than in the garden experiment. Evolution via selection of maternal effects would be possible in the study population because these effects are in part due to genetic differences among parents.
DOIURLPMID [本文引用: 1]
A transect of 68 acid grasslands across Great Britain, covering the lower range of ambient annual nitrogen deposition in the industrialized world (5 to$35 kg N ha^{-1} year^{-1}$), indicates that long-term, chronic nitrogen deposition has significantly reduced plant species richness. Species richness declines as a linear function of the rate of inorganic nitrogen deposition, with a reduction of one species per 4-m2quadrat for every$2.5 kg N ha^{-1} year^{-1}$of chronic nitrogen deposition. Species adapted to infertile conditions are systematically reduced at high nitrogen deposition. At the mean chronic nitrogen deposition rate of central Europe ($17 kg N ha^{-1} year^{-1}$), there is a 23% species reduction compared with grasslands receiving the lowest levels of nitrogen deposition.
[本文引用: 1]
URL [本文引用: 1]
由于人类活动的影响,近年来黄河口营养盐入海通量一直维持在较高水平。大量营养盐的输入不仅可对河口潮滩的氮、磷营养水平以及氮磷比产生重要影响,而且其对于湿地植物的种子萌发与幼苗发育亦可能产生深刻影响。为此,于2010年10-11月,在人工气候箱控制条件下,研究土壤中氮磷比对采自黄河口中潮滩和低潮滩的不同表现型的碱蓬(Suaeda salsa)种子萌发和幼苗生长的影响。结果表明,随着培养基盐分的增加,采自中潮滩和低潮滩的碱蓬种子的萌发率都在减小;当氮磷比为9.5时,在无或低盐分(0 m M和300 m M)胁迫下,中等浓度的氮磷配比(4.524 m M氮+0.476 m M磷)有利于采自中潮滩的碱蓬种子的萌发和幼苗生长,低浓度的氮磷配比(0.905 m M氮+0.095 m M磷)则有利于采自低潮滩的碱蓬种子的萌发和幼苗生长;在高盐分(600 m M)胁迫下,高浓度的氮磷配比(9.05 m M氮+0.95 m M磷)对于采自两种生境的碱蓬种子萌发和幼苗生长都具有明显促进作用;当氮磷比为17时,无论在高盐分还是在低盐分胁迫下,高浓度的氮磷配比(9.444 m M氮+0.556 m M磷)都有利于采自两种生境的碱蓬种子萌发和幼苗生长,说明高氮低磷的配比(氮磷比为17)可以有效减缓盐分胁迫对碱蓬种子萌发的危害,使得碱蓬种子可以更快地适应环境而萌发。研究发现,采用平均生长量来描述外界条件影响下碱蓬种子萌发和幼苗平均生长状况是可行的。平均生长量的引入可以真实还原碱蓬种子萌发和幼苗生长的整体状况,是对鲜质量和干质量等表征碱蓬幼苗生长状况指标的一个有益补充。当前,伴随着黄河口潮滩环境氮磷配比的改变,采自两种生境的碱蓬种子萌发的抗盐分胁迫能力都呈增强趋势,其中以采自中潮滩的碱蓬种子抗盐分胁迫能力的增强最为明显,说明下一步通过调控氮磷比可以有效提高采17
URL [本文引用: 1]
由于人类活动的影响,近年来黄河口营养盐入海通量一直维持在较高水平。大量营养盐的输入不仅可对河口潮滩的氮、磷营养水平以及氮磷比产生重要影响,而且其对于湿地植物的种子萌发与幼苗发育亦可能产生深刻影响。为此,于2010年10-11月,在人工气候箱控制条件下,研究土壤中氮磷比对采自黄河口中潮滩和低潮滩的不同表现型的碱蓬(Suaeda salsa)种子萌发和幼苗生长的影响。结果表明,随着培养基盐分的增加,采自中潮滩和低潮滩的碱蓬种子的萌发率都在减小;当氮磷比为9.5时,在无或低盐分(0 m M和300 m M)胁迫下,中等浓度的氮磷配比(4.524 m M氮+0.476 m M磷)有利于采自中潮滩的碱蓬种子的萌发和幼苗生长,低浓度的氮磷配比(0.905 m M氮+0.095 m M磷)则有利于采自低潮滩的碱蓬种子的萌发和幼苗生长;在高盐分(600 m M)胁迫下,高浓度的氮磷配比(9.05 m M氮+0.95 m M磷)对于采自两种生境的碱蓬种子萌发和幼苗生长都具有明显促进作用;当氮磷比为17时,无论在高盐分还是在低盐分胁迫下,高浓度的氮磷配比(9.444 m M氮+0.556 m M磷)都有利于采自两种生境的碱蓬种子萌发和幼苗生长,说明高氮低磷的配比(氮磷比为17)可以有效减缓盐分胁迫对碱蓬种子萌发的危害,使得碱蓬种子可以更快地适应环境而萌发。研究发现,采用平均生长量来描述外界条件影响下碱蓬种子萌发和幼苗平均生长状况是可行的。平均生长量的引入可以真实还原碱蓬种子萌发和幼苗生长的整体状况,是对鲜质量和干质量等表征碱蓬幼苗生长状况指标的一个有益补充。当前,伴随着黄河口潮滩环境氮磷配比的改变,采自两种生境的碱蓬种子萌发的抗盐分胁迫能力都呈增强趋势,其中以采自中潮滩的碱蓬种子抗盐分胁迫能力的增强最为明显,说明下一步通过调控氮磷比可以有效提高采17
DOIURL [本文引用: 1]
There is large variation in seed weight within the two species of Quercus (Q. dealbata L. and Q. griffithii Hk. f. & Th.) selected for the present study. Effects on germination and survival of oak seedlings were studied in relation to variation in seed weight and microsite characteristics. Germination showed a strong correlation with seed weight. Heavy seeds germinated earlier and showed better germination than light seeds and seedlings produced from the former survived and grew better. The differences in growth of the seedlings emerged from the seeds having different weights persisted in the second year also. Emergence, survival and growth of the seedlings were highest in the plots devoid of litter and herbaceous cover and lowest in the control plots which were not cleared. Seedling emergence, survival and growth were more adversely affected by herbaceous cover than litter.
DOIURL [本文引用: 1]
The low-nitrogen status of highly weathered soils may offer a potential alternative for weed suppression in agricultural systems with N2-fixing crops. In this study, we used sicklepod as a model to evaluate weed response that might occur with managed reductions in nitrogen-soil fertility. A field study was conducted with the parental generation supplied 0, 112, 224, or 448 kg N ha 1. Decreased nitrogen fertility led to reduced shoot biomass, seed number, and total seed mass. Individual seed mass was lower, but seed % nitrogen was not affected. Analysis of seed-mass distribution confirmed that low parental fertility was associated with more small seeds as a proportion of total seeds produced. Additional experiments in hydroponics culture revealed slower growth rates of seedlings produced from small seeds when grown under low-nitrogen conditions. Competitiveness of plants from small (low nitrogen) and large (high nitrogen) seed classes was determined in a replacement-series experiment conducted in sand culture in a controlled environment at two densities and two levels of nitrogen nutrition. Plants produced from smaller seeds were less competitive in low-nitrogen fertility conditions, but plants from small and large seeds competed similarly when grown under high-nitrogen fertility. The results support the hypothesis that comprehensive management strategies to reduce nitrogen availability for weed growth in low-fertility conditions could decrease weed interference by decreasing growth and seed production of parental plants and through maternal effects that lower competitiveness of offspring.
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
1 Succisa pratensis, a characteristic plant species of biodiverse, slightly acidic grasslands (NARDETEA), has declined in the Netherlands over recent decades. This process has mainly been attributed to environmental stress caused by anthropogenic acidification and eutrophication, although genetic deterioration and decreasing population size may also be involved. 2 We analysed the combined effects of environmental stress, genetic variation and population size on the performance of Succisa pratensis. 3 Individual plant performance was negatively affected by eutrophication and, to a smaller extent, by acidification. Plants originating from smaller populations performed less well and showed less genetic variation and higher inbreeding coefficients than plants from large populations. Strong interactions between population size and stress treatments reflected the fact that plants from small populations suffered more from deteriorating soil conditions than those from large populations. 4 Habitat fragmentation may reinforce the negative effects of environmental stress, leading to even the less effective treatment (acidification) having serious consequences.
DOIURL [本文引用: 2]
1. The phenotype of offspring depends on the abiotic and biotic environment in which the parents developed. However, the direct effects of competition experienced by parent plants on single-seed traits are poorly documented despite their impact on plant fitness.
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
At the core of plant regeneration, temperature and water supply are critical drivers for seed dormancy (initiation, break) and germination. Hence, global climate change is altering these environmental cues and will preclude, delay, or enhance regeneration from seeds, as already documented in some cases. Along with compromised seedling emergence and vigour, shifts in germination phenology will influence population dynamics, and thus, species composition and diversity of communities. Altered seed maturation (including consequences for dispersal) and seed mass will have ramifications on life history traits of plants. Predicted changes in temperature and precipitation, and thus in soil moisture, will affect many components of seed persistence in soil, e.g. seed longevity, dormancy release and germination, and soil pathogen activity. More/less equitable climate will alter geographic distribution for species, but restricted migratory capacity in some will greatly limit their response. Seed traits for weedy species could evolve relatively quickly to keep pace with climate change enhancing their negative environmental and economic impact. Thus, increased research in understudied ecosystems, on key issues related to seed ecology, and on evolution of seed traits in nonweedy species is needed to more fully comprehend and plan for plant responses to global warming.
DOIURL [本文引用: 1]
Species differences in seedling survival in deeply shaded understories (i.e., shade tolerance) may depend on both seed size and growth rates, but their relative contributions to survival and how they change with time and with variation in light and belowground resource availability is unknown. With a greenhouse experiment we investigated these relationships by examining responses of growth, growth-related morphology, survival, and their interrelationships to a range of nitrogen (3.4 10-9-3.4 10-3mol/L N fertilizer solutions) and low light (0.6-7.3% of open sky) availabilities for young seedlings of 10 North American tree species that vary in observational shade tolerance rankings and seed size (Populus tremuloides, Betula papyrifera, Betula alleghaniensis, Acer saccharum, Larix laricina, Pinus banksiana, Pinus resinosa, Pinus strobus, Picea mariana, and Abies balsamea). Within all species, relative growth rate (RGR) and survival increased with light. RGR and survival also increased with N supply but only at the two highest light levels, and then only for the shade-intolerant, broad-leaved Populus and Betula spp.. In every species, survival was positively related to RGR. Moreover, each species differed in the relationship of survival to RGR, and these differences were related to seed mass: at any given RGR, large-seeded, shade-tolerant species had higher survival than smaller-seeded, intolerant ones. Across species, in most light and N treatments, seed mass was positively related to young seedling survival, but RGR was not. In very low light the relative benefits of greater seed mass to survival were temporary. As seedlings aged, interspecific mortality rates became more dependent on observational shade tolerance rankings than on seed size, but mortality was still unrelated to RGR. Our results indicate potentially important interactions among light, N, and species that could influence regeneration dynamics. For young seedlings in deeply shaded microsites, N supply does not matter, and only shade-tolerant species survive due, in part, to large seeds and physiological traits other than RGR. In moderate shade, RGR is greater, and survival is high for all species, except that small-seeded, broad-leaved, intolerant species have low survival and RGR at low N supply. This suggests that broad-leaved shade-intolerant species compete more effectively in moderate shade on richer soils than on poorer soils. Although we found that both seed mass and vegetative physiology influenced survival in shade, they did not covary tightly, suggesting that they are under somewhat separate selection pressures.
DOIURL [本文引用: 1]
Morphological shade-avoidance responses have been hypothesized to be a form of adaptive plasticity to improve competition for light; however, little is known about their intraspecific variability and their effect on reproductive fitness. To compare plant responses either at a common age or at a common phenological stage, two experiments were conducted with early- and late-flowering Chenopodium... [Show full abstract]
DOIURL [本文引用: 1]
Effects of nutrient input on Alternanthera philoxeroides were similar between monoculture and mixed culture. Under low nutrient level, effects of N/P ratio on invasive plant growth would be negligible. Nutrient level, rather than N/P ratio, affected the competitive dominance of Alternanthera philoxeroides. Nutrient level cannot alter crucial effects of co-occurring species on the competitive dominance of Alternanthera philoxeroides.
DOIURL [本文引用: 1]
The effect of phosphorus (P) concentration in barley seed on seedling growth has not been much investigated. Consequently, two experiments were conducted in the greenhouse to determine the effect of P concentration in barley seed (Hordeum vulgare L., cv. Empress) on the seedlings grown in sand-filled boxes receiving a culture solution without P. Seeds were selected with three P concentrations: high-P (113.0 mmol P kg6301), medium-P (80.7 mmol P kg6301) and low-P (54.9 mmol P kg6301). At 21 days after sowing, the shoot and root yield or shoot height was the least with seedlings from low-P seed. In the other experiment, high-P and low-P seeds were wetted with distilled water or with a solution of 25.8cmolL6301 of NaH60PO62 for 24 h, and then grown for 31 days. Solution P had been imbibed by seeds whether low or high in native P, but only the imbibed P held by low native P seed benefited seedling dry matter accumulation and shoot elongation. The lack of benefit from seed-imbibed P on seedlings grown from high-P barley seed was associated with low recovery of the imbibed P in those seedlings.
DOIURL [本文引用: 1]
Two experiments were carried out in a growth chamber and a naturally lit glasshouse to investigate the influence of seed phosphorus (P) reserves on growth and P uptake by wheat plants (Triticum aestivum cv Krichauff), and their association with arbuscular mycorrhizal (AM) fungi. Increased seed p reserves improved plant growth at a range of P supply up to over 100 mg P kg鈦宦 soil. Plants grown from seeds with high P reserves tended to accumulate more P from soil, which was mainly attributed to better root system development. Mycorrhizal colonisation did not significantly affect P uptake of plants grown with low irradiance (in growth chamber). However, in the naturally lit glasshouse, mycorrhizal plants had significantly higher P concentrations than non-mycorrhizal plants. Furthermore, mycorrhizal plants grown from seeds low in P accumulated similar amounts of P compared with those grown from seeds with high P, indicating that mycorrhizal colonisation may overcome the disadvantage of having low seed P reserves in the field.
Maternal effects at 4 levels in Senecio vulgaris(Asteraceae) grown on a soil nutrient gradient
1
1990
... 植物功能性状紧密关联生态系统功能(
Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from inner Mongolia Grasslands
3
2010
... 自工业革命100多年来, 伴随着愈演愈烈的人类工业和农业活动, 氮沉降已经成为一个严重的全球性环境问题(
... 灰绿藜(Chenopodium glaucum)广泛分布在全球温带区域, 同时也是内蒙古草原的一年生先锋物种和资源利用型草本植物(
... 本实验在内蒙古草原生态系统定位研究站实施.实验台站位于内蒙古自治区锡林河流域, 具体地理位置为43.63° N, 116.70° E.实验区域年平均气温0.3 ℃, 月平均气温1月份最低(-21.6 ℃), 7月份最高(19 ℃).年降水量346.1 mm, 生长季降水(5-8月)占到全年降水量的60%-80%.该区域的植被类型为典型草原, 灰绿藜为实验区域的一年生先锋物种和资源利用型草本植物(
Seed nutrient content and nutritional status of Posidonia oceanica seedlings in the northwestern Mediterranean Sea
1
2009
... 植物功能性状紧密关联生态系统功能(
Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis
1
2010
... 自工业革命100多年来, 伴随着愈演愈烈的人类工业和农业活动, 氮沉降已经成为一个严重的全球性环境问题(
Irrigation and fertilization effects on seed number, size, germination and seedling growth: Implications for desert shrub establishment
2
2008
... 本研究的主要创新性在于系统区分和揭示了氮磷供应量、比例及其交互作用对植物种子性状影响的相对贡献.前人研究主要集中在氮或磷添加水平对种子的影响, 并发现氮或磷供应量显著改变种子性状(
... 从种子萌发性状来看, 我们发现在不同的氮磷比例情况下, 低的氮磷供应量均增加种子萌发率.这与以往实验发现低养分条件促进种子萌发率的结果(
Plant species traits are the predominant control on litter decomposition rates within biomes worldwide
1
2008
... 植物功能性状紧密关联生态系统功能(
Are Mojave Desert annual species equal? Resource acquisition and allocation for the invasive grass Bromus madritensis subsp rubens (Poaceae) and two native species
1
2003
... 当灰绿藜种子成熟时(9月4日), 对所有植物个体都进行取样.在实验室里把种子从植物个体里分离出来, 并在空气中风干.每株随机选取20粒种子称质量, 用来估算种子大小.种子萌发实验在正常室温下进行(18-20 ℃), 同时接受自然光照(
Allocation strategies and seed traits are hardly affected by nitrogen supply in 18 species differing in successional status
3
2009
... 前人研究表明土壤氮含量增加会显著影响种子大小(增加或降低)、氮浓度(增加)、磷浓度(降低)、萌发率(降低)和萌发速度(降低)(
... 本研究的主要创新性在于系统区分和揭示了氮磷供应量、比例及其交互作用对植物种子性状影响的相对贡献.前人研究主要集中在氮或磷添加水平对种子的影响, 并发现氮或磷供应量显著改变种子性状(
... 从种子形态性状来看, 在氮磷比例为5、15或45的情况下, 氮磷供应量对种子大小没有显著影响(
Time-dependent, species-specific effects of N:P stoichiometry on grassland plant growth
1
2010
... 植物功能性状紧密关联生态系统功能(
Parental environmental effects on life history in the herbaceous plant Campanula americana
1
2001
... 本研究的主要创新性在于系统区分和揭示了氮磷供应量、比例及其交互作用对植物种子性状影响的相对贡献.前人研究主要集中在氮或磷添加水平对种子的影响, 并发现氮或磷供应量显著改变种子性状(
The importance of early season phosphorus nutrition
2
2001
... 植物功能性状紧密关联生态系统功能(
... 从种子养分性状来看, 在氮磷比例为15或45的情况下, 低量养分供应显著提高种子氮浓度, 尽管在氮磷比例为5的情况下养分供应量的效应不显著.这与以往低养分条件对种子氮浓度影响的实验结果(降低或不影响)(
Phosphorus accumulation in Proteaceae seeds: A synthesis
1
2010
... 从种子养分性状来看, 在氮磷比例为15或45的情况下, 低量养分供应显著提高种子氮浓度, 尽管在氮磷比例为5的情况下养分供应量的效应不显著.这与以往低养分条件对种子氮浓度影响的实验结果(降低或不影响)(
N:P ratios in terrestrial plants: Variation and functional significance
1
2004
... 植物功能性状紧密关联生态系统功能(
High nitrogen: phosphorus ratios reduce nutrient retention and second-year growth of wetland sedges
4
2005a
... 自工业革命100多年来, 伴随着愈演愈烈的人类工业和农业活动, 氮沉降已经成为一个严重的全球性环境问题(
... 植物功能性状紧密关联生态系统功能(
... 沙培盆栽实验进行时间为2010年6月18日至9月4日.实验包括9个处理, 采用3个氮磷供应量水平和3个氮磷供应比例的完全正交设计(
... ;
Responses of wetland graminoids to the relative supply of nitrogen and phosphorus
1
2005b
... 植物功能性状紧密关联生态系统功能(
Composition of plant species mixtures grown at various N:P ratios and levels of nutrient supply
3
2003
... 植物功能性状紧密关联生态系统功能(
... 沙培盆栽实验进行时间为2010年6月18日至9月4日.实验包括9个处理, 采用3个氮磷供应量水平和3个氮磷供应比例的完全正交设计(
... ), 每个处理20个重复.氮磷供应量通过灰绿藜植物个体生命周期所需要的氮量(44.5 mg)和磷量(3.1 mg)的几何平均数计算得出, 选用几何平均数是基于氮和磷对植物生长影响的乘法效应(multiplicative effect)(
Effect of nitrogen, phosphorus and potassium availability on mother plant size, seed production and germination ability of Rumex crispus
5
2012
... 前人研究表明土壤氮含量增加会显著影响种子大小(增加或降低)、氮浓度(增加)、磷浓度(降低)、萌发率(降低)和萌发速度(降低)(
... ;
... 本研究的主要创新性在于系统区分和揭示了氮磷供应量、比例及其交互作用对植物种子性状影响的相对贡献.前人研究主要集中在氮或磷添加水平对种子的影响, 并发现氮或磷供应量显著改变种子性状(
... 从种子养分性状来看, 在氮磷比例为15或45的情况下, 低量养分供应显著提高种子氮浓度, 尽管在氮磷比例为5的情况下养分供应量的效应不显著.这与以往低养分条件对种子氮浓度影响的实验结果(降低或不影响)(
... 从种子萌发性状来看, 我们发现在不同的氮磷比例情况下, 低的氮磷供应量均增加种子萌发率.这与以往实验发现低养分条件促进种子萌发率的结果(
Germination and seedling development of bigroot morningglory ( Ipomoea pandurata)
1
1991
... 当灰绿藜种子成熟时(9月4日), 对所有植物个体都进行取样.在实验室里把种子从植物个体里分离出来, 并在空气中风干.每株随机选取20粒种子称质量, 用来估算种子大小.种子萌发实验在正常室温下进行(18-20 ℃), 同时接受自然光照(
Competitive response hierarchies for germination, growth, and survival and their influence on abundance
1
2001
... 植物功能性状紧密关联生态系统功能(
Production, size, and germination of broad-leaved dock seeds collected from mother plants grown under different nitrogen, phosphorus, and potassium supplies
5
2011
... 前人研究表明土壤氮含量增加会显著影响种子大小(增加或降低)、氮浓度(增加)、磷浓度(降低)、萌发率(降低)和萌发速度(降低)(
... ).同时, 土壤磷富集也显著改变种子大小(增加或降低)、磷浓度(增加)、氮浓度(增加)和种子萌发率(降低) (
... 本研究的主要创新性在于系统区分和揭示了氮磷供应量、比例及其交互作用对植物种子性状影响的相对贡献.前人研究主要集中在氮或磷添加水平对种子的影响, 并发现氮或磷供应量显著改变种子性状(
... 从种子养分性状来看, 在氮磷比例为15或45的情况下, 低量养分供应显著提高种子氮浓度, 尽管在氮磷比例为5的情况下养分供应量的效应不显著.这与以往低养分条件对种子氮浓度影响的实验结果(降低或不影响)(
... 从种子萌发性状来看, 我们发现在不同的氮磷比例情况下, 低的氮磷供应量均增加种子萌发率.这与以往实验发现低养分条件促进种子萌发率的结果(
黄河口不同氮基质碱蓬种子萌发及幼苗生长对盐分及氮输入的响应
1
2017
... 前人研究表明土壤氮含量增加会显著影响种子大小(增加或降低)、氮浓度(增加)、磷浓度(降低)、萌发率(降低)和萌发速度(降低)(
黄河口不同氮基质碱蓬种子萌发及幼苗生长对盐分及氮输入的响应
1
2017
... 前人研究表明土壤氮含量增加会显著影响种子大小(增加或降低)、氮浓度(增加)、磷浓度(降低)、萌发率(降低)和萌发速度(降低)(
Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the Holy Grail
1
2002
... 植物功能性状紧密关联生态系统功能(
Does the seed size/number trade-off model determine plant community structure? An assessment of the model mechanisms and their generality
2
2001
... 植物功能性状紧密关联生态系统功能(
... 本研究的主要创新性在于系统区分和揭示了氮磷供应量、比例及其交互作用对植物种子性状影响的相对贡献.前人研究主要集中在氮或磷添加水平对种子的影响, 并发现氮或磷供应量显著改变种子性状(
Effects of increased nitrogen and phosphorus deposition on offspring performance of two dominant species in a temperate steppe ecosystem
5
2017
... 前人研究表明土壤氮含量增加会显著影响种子大小(增加或降低)、氮浓度(增加)、磷浓度(降低)、萌发率(降低)和萌发速度(降低)(
... ;
... 本研究的主要创新性在于系统区分和揭示了氮磷供应量、比例及其交互作用对植物种子性状影响的相对贡献.前人研究主要集中在氮或磷添加水平对种子的影响, 并发现氮或磷供应量显著改变种子性状(
... 从种子萌发性状来看, 我们发现在不同的氮磷比例情况下, 低的氮磷供应量均增加种子萌发率.这与以往实验发现低养分条件促进种子萌发率的结果(
... ;
Aggravated phosphorus limitation on biomass production under increasing nitrogen loading: A meta-analysis
1
2016
... 自工业革命100多年来, 伴随着愈演愈烈的人类工业和农业活动, 氮沉降已经成为一个严重的全球性环境问题(
A global perspective on belowground carbon dynamics under nitrogen enrichment
1
2010
... 自工业革命100多年来, 伴随着愈演愈烈的人类工业和农业活动, 氮沉降已经成为一个严重的全球性环境问题(
外源磷输入对兴凯湖沼泽土壤种子库种子萌发的影响
2
2013
... 自工业革命100多年来, 伴随着愈演愈烈的人类工业和农业活动, 氮沉降已经成为一个严重的全球性环境问题(
... 前人研究表明土壤氮含量增加会显著影响种子大小(增加或降低)、氮浓度(增加)、磷浓度(降低)、萌发率(降低)和萌发速度(降低)(
外源磷输入对兴凯湖沼泽土壤种子库种子萌发的影响
2
2013
... 自工业革命100多年来, 伴随着愈演愈烈的人类工业和农业活动, 氮沉降已经成为一个严重的全球性环境问题(
... 前人研究表明土壤氮含量增加会显著影响种子大小(增加或降低)、氮浓度(增加)、磷浓度(降低)、萌发率(降低)和萌发速度(降低)(
Enhanced nitrogen deposition over China
2013
Spatial heterogeneity distribution of soil total nitrogen and total phosphorus in the Yaoxiang watershed in a hilly area of northern China based on geographic information system and geostatistics
1
2016
... 当灰绿藜种子成熟时(9月4日), 对所有植物个体都进行取样.在实验室里把种子从植物个体里分离出来, 并在空气中风干.每株随机选取20粒种子称质量, 用来估算种子大小.种子萌发实验在正常室温下进行(18-20 ℃), 同时接受自然光照(
科尔沁沙地69种植物种子重量比较研究
1
2004a
... 植物功能性状紧密关联生态系统功能(
科尔沁沙地69种植物种子重量比较研究
1
2004a
... 植物功能性状紧密关联生态系统功能(
科尔沁沙地31种1年生植物萌发特性比较研究
1
2004b
... 植物功能性状紧密关联生态系统功能(
科尔沁沙地31种1年生植物萌发特性比较研究
1
2004b
... 植物功能性状紧密关联生态系统功能(
Responses of ecosystem nitrogen cycle to nitrogen addition: A meta-analysis
1
2011
... 自工业革命100多年来, 伴随着愈演愈烈的人类工业和农业活动, 氮沉降已经成为一个严重的全球性环境问题(
Nitrogen: phosphorous supply ratio and allometry in five alpine plant species
2
2016
... 自工业革命100多年来, 伴随着愈演愈烈的人类工业和农业活动, 氮沉降已经成为一个严重的全球性环境问题(
... 植物功能性状紧密关联生态系统功能(
Varying phosphorus supply and development, growth and seed yield in narrow-leafed lupin
1
2002
... 从种子养分性状来看, 在氮磷比例为15或45的情况下, 低量养分供应显著提高种子氮浓度, 尽管在氮磷比例为5的情况下养分供应量的效应不显著.这与以往低养分条件对种子氮浓度影响的实验结果(降低或不影响)(
Shifts in seed size across experimental nitrogen enrichment and plant density gradients
1
2009
... 前人研究表明土壤氮含量增加会显著影响种子大小(增加或降低)、氮浓度(增加)、磷浓度(降低)、萌发率(降低)和萌发速度(降低)(
Seed/cotyledon size and nutrient content play a major role in early performance of species on nutrient-poor soils
1
1997
... 植物功能性状紧密关联生态系统功能(
Seedling survival and seed size: A synthesis of the literature
1
2004
... 植物功能性状紧密关联生态系统功能(
Relative contribution of seed phosphorus reserves and exogenous phosphorus uptake to maize ( Zea mays L.) nutrition during early growth stages
1
2011
... 植物功能性状紧密关联生态系统功能(
Influence of seed nitrogen content on seedling growth and recovery from nitrogen stress
2
2005
... 植物功能性状紧密关联生态系统功能(
... 本研究的主要创新性在于系统区分和揭示了氮磷供应量、比例及其交互作用对植物种子性状影响的相对贡献.前人研究主要集中在氮或磷添加水平对种子的影响, 并发现氮或磷供应量显著改变种子性状(
Seed germination and seedling survival of two threatened endemic species of the northwest Iberian peninsula
1
2003
... 植物功能性状紧密关联生态系统功能(
Human-induced nitrogen-phosphorus imbalances alter natural and managed ecosystems across the globe
1
2013
... 自工业革命100多年来, 伴随着愈演愈烈的人类工业和农业活动, 氮沉降已经成为一个严重的全球性环境问题(
Effects of maternal and paternal environment and genotype on offspring phenotype in Solidago altissima L
1
1994
... 植物功能性状紧密关联生态系统功能(
Impact of nitrogen deposition on the species richness of grasslands
1
2004
... 自工业革命100多年来, 伴随着愈演愈烈的人类工业和农业活动, 氮沉降已经成为一个严重的全球性环境问题(
Phenotypic plasticity for offspring traits in Polygonum persicaria
1
1996
... 从种子养分性状来看, 在氮磷比例为15或45的情况下, 低量养分供应显著提高种子氮浓度, 尽管在氮磷比例为5的情况下养分供应量的效应不显著.这与以往低养分条件对种子氮浓度影响的实验结果(降低或不影响)(
黄河口潮滩碱蓬种子萌发与幼苗生长对土壤中氮磷比的响应
1
2017
... 前人研究表明土壤氮含量增加会显著影响种子大小(增加或降低)、氮浓度(增加)、磷浓度(降低)、萌发率(降低)和萌发速度(降低)(
黄河口潮滩碱蓬种子萌发与幼苗生长对土壤中氮磷比的响应
1
2017
... 前人研究表明土壤氮含量增加会显著影响种子大小(增加或降低)、氮浓度(增加)、磷浓度(降低)、萌发率(降低)和萌发速度(降低)(
Effects of seed weight and microsite characteristics on germination and seedling fitness in 2 species of Quercus in a subtropical wet hill forest
1
1990
... 当灰绿藜种子成熟时(9月4日), 对所有植物个体都进行取样.在实验室里把种子从植物个体里分离出来, 并在空气中风干.每株随机选取20粒种子称质量, 用来估算种子大小.种子萌发实验在正常室温下进行(18-20 ℃), 同时接受自然光照(
Altered weed reproduction and maternal effects under low-nitrogen fertility
1
2006
... 本研究的主要创新性在于系统区分和揭示了氮磷供应量、比例及其交互作用对植物种子性状影响的相对贡献.前人研究主要集中在氮或磷添加水平对种子的影响, 并发现氮或磷供应量显著改变种子性状(
Early emergence enhances plant fitness: A phylogenetically controlled meta-analysis
1
2005
... 植物功能性状紧密关联生态系统功能(
Effects of population size and genetic variation on the response of Succisa pratensis to eutrophication and acidification
2
2003
... 从种子形态性状来看, 在氮磷比例为5、15或45的情况下, 氮磷供应量对种子大小没有显著影响(
... 从种子养分性状来看, 在氮磷比例为15或45的情况下, 低量养分供应显著提高种子氮浓度, 尽管在氮磷比例为5的情况下养分供应量的效应不显著.这与以往低养分条件对种子氮浓度影响的实验结果(降低或不影响)(
Intraspecific seed trait variations and competition: Passive or adaptive response?
2
2009
... 从种子养分性状来看, 在氮磷比例为15或45的情况下, 低量养分供应显著提高种子氮浓度, 尽管在氮磷比例为5的情况下养分供应量的效应不显著.这与以往低养分条件对种子氮浓度影响的实验结果(降低或不影响)(
... ;
Let the concept of trait be functional!
1
2007
... 植物功能性状紧密关联生态系统功能(
Climate change and plant regeneration from seed
1
2011
... 植物功能性状紧密关联生态系统功能(
Seed size, nitrogen supply, and growth rate affect tree seedling survival in deep shade
1
2000
... 植物功能性状紧密关联生态系统功能(
Intraspecific variability and maternal effects in the response to nutrient addition in Chenopodium album
1
1999
... 当灰绿藜种子成熟时(9月4日), 对所有植物个体都进行取样.在实验室里把种子从植物个体里分离出来, 并在空气中风干.每株随机选取20粒种子称质量, 用来估算种子大小.种子萌发实验在正常室温下进行(18-20 ℃), 同时接受自然光照(
Shifts in growth and competitive dominance of the invasive plant Alternanthera philoxeroides under different nitrogen and phosphorus supply
1
2017
... 自工业革命100多年来, 伴随着愈演愈烈的人类工业和农业活动, 氮沉降已经成为一个严重的全球性环境问题(
Phosphorus concentration in barley ( Hordeum vulgare L.) seed: Influence on seedling growth and dry-matter production
1
1990
... 植物功能性状紧密关联生态系统功能(
Seed phosphorus (P) content affects growth, and P uptake of wheat plants and their association with arbuscular mycorrhizal (AM) fungi
1
2001
... 植物功能性状紧密关联生态系统功能(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}