 ,, 杨庆松, 刘何铭, 王希华,
,, 杨庆松, 刘何铭, 王希华, ,*华东师范大学生态与环境科学学院, 浙江天童森林生态系统国家野外科学观测研究站, 上海 200241
,*华东师范大学生态与环境科学学院, 浙江天童森林生态系统国家野外科学观测研究站, 上海 200241Classification of plant associations based on a 20 hm2 dynamics plot of evergreen broad- leaved forest in Mt. Tiantong, Zhejiang, China
SUN Xiao-Wei,, YANG Qing-Song, LIU He-Ming, WANG Xi-Hua,,*Tiantong National Station for Forest Ecosystem Research, School of Ecological and Environmental Sciences, East China Normal University, Shanghai 200241, China通讯作者: xhwang@des.ecnu.edu.cn
编委: 于明坚
责任编辑: 李敏
收稿日期:2017-10-19修回日期:2018-05-07网络出版日期:2018-05-20
| 基金资助: |
Received:2017-10-19Revised:2018-05-07Online:2018-05-20
| Fund supported: |

摘要
为获取能够代表浙江天童山的森林植被典型群丛类型, 同时也为植被分类中如何发现过渡类型和确定典型类型提供参考, 该研究利用天童20 hm 2森林大样地资料, 运用双向指示种分析(TWINSPAN)与除趋势对应分析(DCA), 剔除过渡群落, 进行群丛划分。结果表明, 去除过渡地段后更利于研究区域典型群丛类型的确定。大样地的植被类型可划分为宜昌荚蒾-厚皮香/港柯+云山青冈群丛(Viburnum erosum-Ternstroemia gymnanthera/Lithocarpus harlandii + Cyclobalanopsis sessilifolia Ass.); 虎皮楠-柯/木荷+米槠群丛(Daphniphyllum oldhami-Lithocarpus glaber/Schima superba + Castanopsis carlesii Ass.); 红毒茴-紫楠/南酸枣+薄叶润楠群丛(Illicium lanceolatum-Phoebe sheareri/Choerospondias axillaries + Machilus leptophylla Ass.)。DCA排序同时能反映各群丛类型分布与环境的相关关系, 结果显示, 海拔和凹凸度对群丛分布有较大影响, 坡度和坡向对群丛分布影响较小。
关键词:
Abstract
Keywords:
PDF (8719KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
孙小伟, 杨庆松, 刘何铭, 王希华. 基于浙江天童20 hm2常绿阔叶林动态监测样地的群丛划分 . 植物生态学报[J], 2018, 42(5): 550-561 DOI:10.17521/cjpe.2017.0263
SUN Xiao-Wei, YANG Qing-Song, LIU He-Ming, WANG Xi-Hua.
植被分类是对植物群落进行类型划分, 并根据各类型间一定相互关系建立相应分类体系的过程。作为基础科学研究的重要工具, 植被分类可以极大地促进人们对生态学格局的理解, 帮助研究者收集和解释生态背景信息(van der Maarel & Franklin, 2013), 推进生态学的发展。一个合适的分类系统可以为基础生态学调查和生态系统多样性研究、保护与评估提供必要的基础(Jennings et al., 2009), 为自然资源的清查与监测、环境保护方案的制定与实施及生态恢复等各项工作提供重要指导。植被分类无论在理论研究还是实践应用中都有重要意义。
植被分类随着植被生态学的发展而深入, 但到目前为止尚未有一种全球公认的标准分类方法和体系。这不仅由于植被本身的复杂性和区域性, 同时也由于植物群落连续性与间断性的争论, 使得植被分类成为植被生态学研究中最复杂的一个问题(宋永昌, 2017)。目前有一种趋势, 认为植被既是连续的, 也是间断的, 植被分类就是要在连续中找间断(van der Maarel & Franklin, 2013; 宋永昌, 2017)。一种比较现实的做法是在连续植被中首先确定其典型类型, 然后根据群落相似性、群落整体组成的相对存在度或多度等指标判定过渡群落的归属(Jennings et al., 2009; van der Maarel & Franklin, 2013)。
群丛是植被分类的基本单位, 相当于植物分类中“种”的分类水平, 进行群丛分类是了解一个地区植被特征最基本、最重要的方法之一(吴征镒, 1980)。植被研究中的不同学派, 也都集中反映在群丛的定义上。建立在特征种(character species)基础上的法瑞学派的群丛定义(Braun-Blanquet, 1964; Westhoff & van der Maarel, 1985), 被认为是正规分类系统单位(Whittker, 1985)。这种以植物区系特征作为植被分类基本单位的划分标准, 强调特征种、鉴别种(diagnostic species)或标志种(indicative species)的作用, 已被我国(吴征镒, 1980; 宋永昌, 2011; 宋永昌等, 2017)及美国(Federal Geographic Data Committee, Vegetation Subcommittee (FGDV-VS), 2008)植被分类系统中的群丛定义所采用。因为基于研究对象本身特征的分类比其他任何分类更加自然, 这样也同时考虑了植物本身的生态特性以及它们对环境的指示意义(吴征镒, 1980)。按此定义划分出的群丛既能表明植物种类组成和结构的一致性, 同时也能够反映出它们占据生境的相似性。
制作群落表(排表法)是法瑞学派划分群丛的主要方法, 但这一方法也常为它的烦琐复杂和一定的主观性被人们所诟病。近几十年来兴起的数量分类与排序为改进这种状况提供了有利的条件, 尤其是双向指示种分析法(two way indicator species analysis, TWINSPAN)和除趋势对应分析(detrended correspondence analysis, DCA)。两者结合, 既符合群落间的界线是间断的观点, 同时又顾及群落间的连续变化, 一方面利于发现过渡类型, 确定典型类型, 另一方面利于更深刻地揭示植物种、植物群落与环境之间的生态关系。
群丛分类的基础是野外样地调查, 它为植被分类提供了必要的原始材料。在通常的植被调查中, 为保证取样的代表性, 选择适当的取样地点是植被调查的关键。如果取样不当, 不仅不利于准确认识植被的总体, 反而可能会歪曲总体, 从而得出错误的结论(宋永昌, 2017)。更重要的是, 这种典型样地取样法不能得到连续的植被数据, 无法客观评价所取样地的代表性及分类结果的典型性。近年来, 国内研究者在各类森林中建立了一系列面积超过20 hm2的大型动态监测样地, 可以将其视为植被连续分布地段, 通过这些样地我们可以获得较大区域内连续森林群落的调查数据, 这为我们研究群丛分类提供了新的平台和有利条件。
常绿阔叶林是亚热带地区的地带性植被, 种类组成复杂, 类型多样, 在人类长期干预下, 次生性强, 更增加了分类的困难。浙江天童20 hm2动态监测样地内的植物群落具有典型的中国东部中亚热带低海拔森林特征, 群落发育成熟, 结构完整, 对其物种成分已有报道(杨庆松等, 2011)。本文以天童20 hm2样地2015年调查数据为基础, 在连续森林群落调查样地的基础上, 运用数量分类(TWINSPAN分析)与排序(DCA排序), 进行群丛划分, 同时分析获得的群丛类型与环境的相关关系, 探讨植被连续分布地段内的群丛分类问题。
1 材料和方法
1.1 研究区域及样地概况
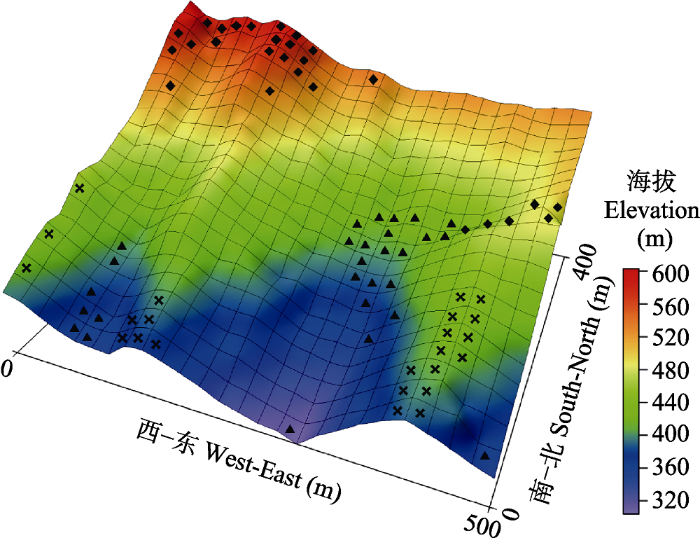
研究区域位于浙江省宁波市鄞州区东南部的天童国家森林公园内, 地处29.80° N, 121.78° E。该地森林植被保存良好, 是浙江省东部丘陵地区地带性植被类型的代表性地段。该区气候为中亚热带季风气候, 全年温和多雨, 四季分明。年平均气温为16.2 ℃, 最热月为7月(月平均气温28.1 ℃), 最冷月为1月(月平均气温4.2 ℃)。区域内的年降水量为1374.7 mm, 主要集中于每年的5-8月(宋永昌和王祥荣, 1995)。天童20 hm2样地位于天童国家森林公园的核心保护区(29.81°-29.82° N, 121.78°-121.79° E), 样地位置及三维地形图如图1所示。样地为长方形, 东西长500 m, 南北宽400 m, 最高海拔602.89 m, 最低海拔304.26 m, 平均海拔447.25 m。样地总体上北高南低, 以东南坡向为主; 样地内包含两条较大的山脊, 南北向纵贯样地, 地形复杂(杨庆松等, 2011)。
图1
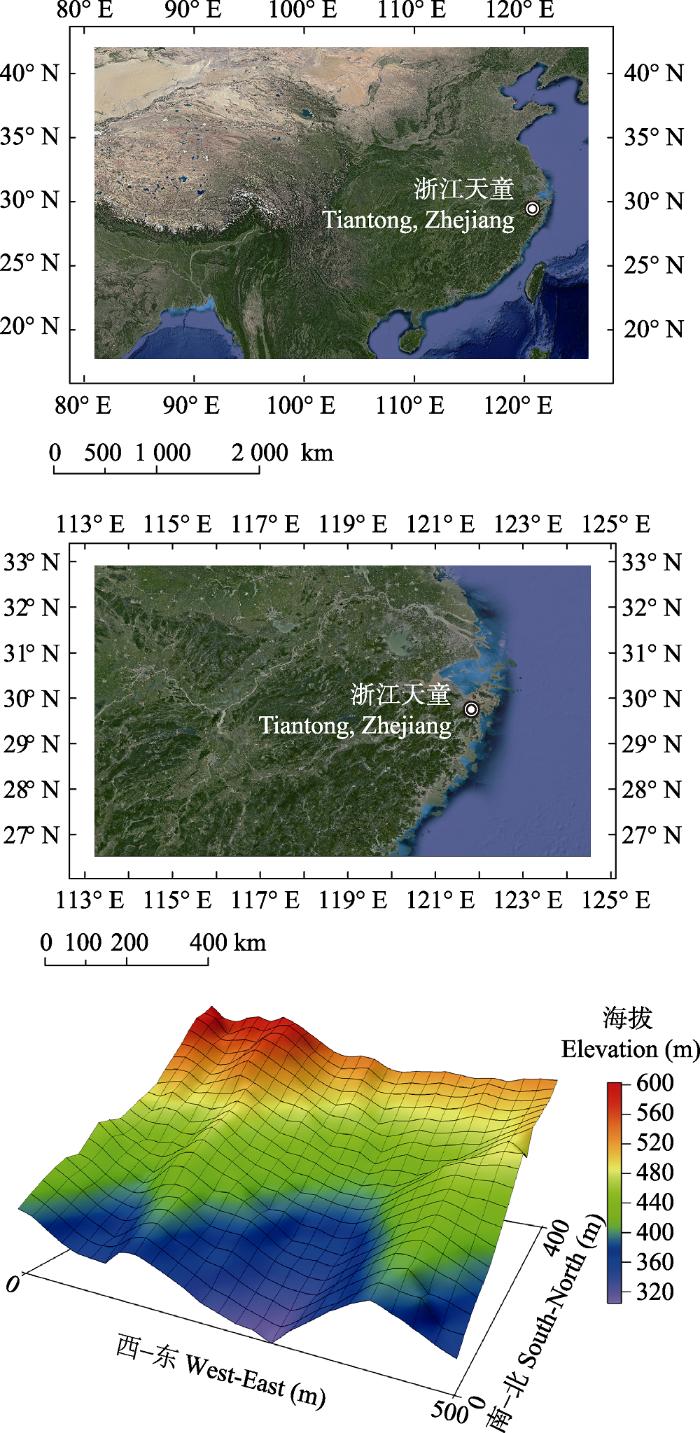 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1浙江省天童20 hm2森林动态样地所在位置及三维地形图。
Fig. 1The location and topography of the 20 hm2 forest dynamics plot in Mt. Tiantong, Zhejiang Province.
1.2 样地设置及调查方法
用全站仪将整个样地划分成500个20 m × 20 m的样方, 将样方按1-500编号, 如原点(西南角第一个样方)处的20 m × 20 m样方编号为1, 1号样方右边为21号样方, 东北角最后一个样方编号为500。样地内所有胸径(DBH) ≥ 1 cm的木本植物均鉴定到种, 调查时测量其胸径并挂牌, 以便长期监测, 记录其编号、种名、胸径、坐标、生长状况以及生境概况, 详见巴拿马Barro Colorado岛(BCI)样地建设规范(Condit, 1998)。本样地共记录154种木本植物, 隶属于52科96属(杨庆松等, 2011)。1.3 数据分析方法
1.3.1 物种数据用重要值(IV)表示物种在各群落样方中的相对重要性。计算公式如下:
重要值= (物种相对多度+物种相对胸高断面积)/2
此处考虑到大样地的实际情况, 重要值计算中未采用相对频度参数, 我们将在讨论部分探讨该问题。本研究以20 m × 20 m样方作为分类单元, 这里采用物种的重要值和样方数据, 组成了154 × 500维物种-样方数据矩阵。
1.3.2 环境因子数据
环境数据为每个20 m × 20 m样方的4个地形因子: 海拔、凹凸度、坡度、坡向。地形因子计算方法参照ForestGEO (Forest Global Earth Observatory)样地常用方法(Harms et al., 2001; Valencia et al., 2004)。海拔为样方4个顶点海拔的平均值, 凹凸度为该样方的平均海拔减去与该样方相邻的8个样方平均海拔, 对于边缘的样方, 凹凸度则为该样方中心点的平均海拔减去周围4个顶点的海拔平均值(Yamakura et al., 1995)。样方坡度与坡向的计算方法是: 从样方4个顶点任取3个顶点组成一个平面, 4个顶点可以组合成4个不同的平面, 这4个平面与样方投影水平面夹角的平均值和与正北方向偏差角度的平均值分别为样方的坡度与坡向值(Nee, 2005)。4个地形因子数据与样方数据组成了4 × 500维地形因子-样方数据矩阵。
1.3.3 植被数据的选择
根据已有的植物群落调查规范(方精云等, 2009; 宋永昌, 2017), 样地选择符合以下要求: (1)样地内的植被应尽可能均匀一致, 在样地内没有结构明显的分界线或分层的变化; (2)群落片段内应具有一致的物种种类成分, 样地应是同质的; (3)样地内的生境条件应相对的均匀一致, 样地内不应有大的林窗。根据前人研究, 去除包含林窗面积> 20 m2的样方237个(张志国等, 2012; 刘何铭等, 2015), 受干扰区域的样方42个(谢玉彬等, 2012)以及用ArcGIS 10.0分析去除样地内物种分布不均匀的样方(图2) 28个, 剩余198个样方用于本次分类, 包含133种木本植物。198个样方分布如图3所示, 本文用于分类的样方连续分布于样地内。
图2
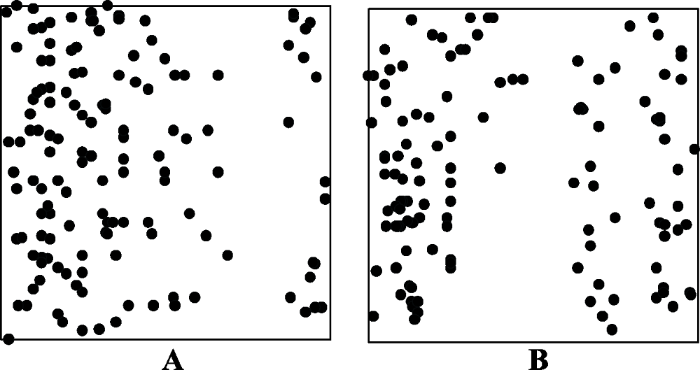 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2物种分布不均匀的样方(A, 样方编号83, B, 样方编号324)。圆点表示样方中的植物个体。
Fig. 2Quadrats with uneven distribution of species (A, stands for quadrat 83; B, stands for quadrat 324). Dots represent the plant individuals in each quadrat.
图3
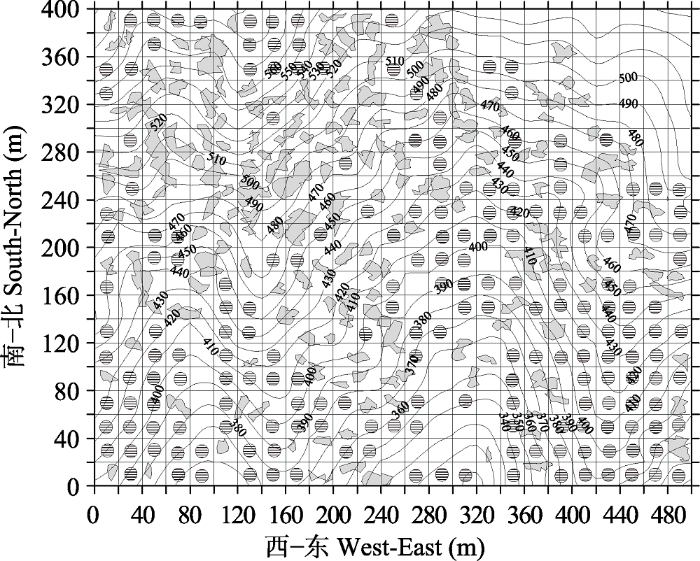 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3天童20 hm2样地中用于分类的20 m × 20 m的样方分布图(198个样方, 等高线间距为10 m)。灰色阴影部分为林窗, 含有阴影圆圈的样方为用于分类的样方。
Fig. 3Map of quadrats for classification at the scale of 20 m × 20 m in Tiantong 20 hm2 plot (198 quadrats in total) (The contour lines with 10 m intervals were shown). Gray polygons represent forest gaps, and shadow circles stand for the quadrats used for classification.
1.3.4 数量分类与排序
TWINSPAN是依据各样方内物种种类组成及各物种重要值进行分类, 最后得到同时包含物种分类和样方分类的一个矩阵(Hill, 1979)。用PC-ORD 5.0软件进行TWINSPAN分析, 程序运行时按: 0; 0.1-1.0; 1.0-2.0; 2.0-5.0; 5.0-10; 10-15; 15-25; 25-50; 50-75将重要值分为9个等级。
DCA是基于植物种类组成数据, 在对应分析(CA)基础上修改而成的特征向量排序, 它克服了弓形效应, 提高了排序精度(张金屯, 2011)。DCA排序结果能够客观反映环境与植物群落间的生态关系, 同时体现植被的过渡性, 为找寻某一研究区域的过渡性群落提供依据。因此本研究采用DCA排序对样方进行分析, 并在排序图上被动加入环境因子, 探讨影响群丛分布格局的环境梯度。DCA排序运用R语言vegan包(Oksanen et al., 2017)计算。
1.3.5 分类结果的评估
群落实体单位确定后, 就需要分析群落样方数据, 以明确所划定群落类型内部是否均一, 以及与其他类型是否差异显著(Lep? & ?milauer, 2003)。本文用轮廓宽度值(silhouette width)来评估分类的有效性。轮廓宽度值能够度量一个样方与所分配类型和其他最相似类型之间相似度的差异程度(Rousseeuw, 1987)。轮廓宽度值范围从-1到1, 该值为正值表示该样方与所分配类型符合度较好, 负值则表示该样方与其他类型更相似。通过计算某一分类类型中全部样方的轮廓宽度均值和轮廓宽度负值数量, 就可以评估该分类结果的好坏。计算公式如下:
S(i) = (bi - ai)/max{bi, ai}
ai是样方i与同一个类型中其他样方的平均距离(相异度); bi是样方i与最相似类型中所有样方的平均距离(相异度)。这里首先使用R语言vegan包(Oksanen et al., 2017)中vegdist函数计算相异度(Bray-Curits系数), 然后使用cluster包(Maechler et al., 2017)中silhouette函数计算轮廓宽度值。
1.3.6 指示值(IndVal)计算
IndVal由Dufrêne和Legendre (1997)创建, de Cáceres和Legendre (2009)以及de Cáceres等(2010)分别进行过改进, 它是植被分类中识别标志种最广泛使用的统计量(van der Maarel & Franklin, 2013)。该指标基于群落物种有无数据计算特异性(A), 基于群落物种多度数据计算确限度(B), A、B两部分综合计算得到IndVal (Dufrêne & Legendre, 1997; de Cáceres & Legendre, 2009; de Cáceres et al., 2010), IndVal最大值为1, 表示该物种对此群丛的指示意义最大。该统计量用R语言indicspecies包(de Cáceres & Jansen, 2016)中multipatt函数计算。
2 结果和分析
2.1 群丛类型的确定
采用TWINSPAN分析将198个样方划分为5个类群(图4), 分别命名为类群A、B、C、D、E。在图5中, 不同字母代表不同类群中的样方并在高程图上显示其在样地中的位置。DCA排序得到的前4个排序轴的特征值分别是: 0.3632、0.2043、0.1063、0.1037。第一排序轴的特征值最大, 第二轴次之, 前两个轴包含了较多的生态信息, 故依据前两个排序轴做出DCA分析二维排序图(图6)。将TWINSPAN分析结果与DCA排序结果结合分析, 即在排序图上圈定各类群的界限。从图6看出, 类群C中的样方位置与类群A、类群B、类群E之间都有重叠区域, 类群D与类群A、类群C、类群E之间有重叠区域, 故认为在这5个类群中, 类群C与类群D为过渡类型, 将这两个过渡类型去除后, 再重复进行数量分类与排序分析。图4
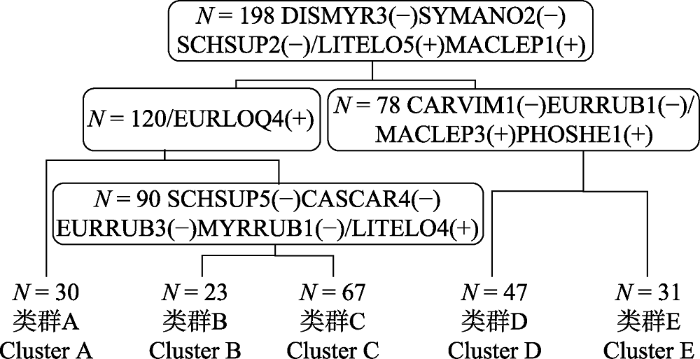 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4天童20 hm2样地内用于分类的198个样方的TWINSPAN分类结果树状示意图。N表示样方总数, 方框里的物种表示各分类水平的正负指示种, 两者用“/”区分。CARVIM, 雷公鹅耳枥; CASCAR, 米槠; DISMYR, 杨梅叶蚊母树; EURLOQ, 细枝柃; EURRUB, 窄基红褐柃; LITELO, 黄丹木姜子; MACLEP, 薄叶润楠; MYRRUB, 杨梅; PHOSHE, 紫楠; SCHSUP, 木荷; SYMANO, 薄叶山矾。
Fig. 4The result of TWINSPAN classification for 198 quadrats in Tiantong 20 hm2 plot. N represents total number of quadrats. Species in the box are indicator species, and a negative group and a positive group are included at each level of classification, they are separated by “/”. CARVIM, Carpinus viminea; CASCAR, Castanopsis carlesii; DISMYR, Distylium myricoides; EURLOQ, Eurya loquaiana; EURRUB, Eurya rubiginosa var. attenuata; LITELO, Litsea elongata; MACLEP, Machilus leptophylla; MYRRUB, Myrica rubra; PHOSHE, Phoebe sheareri; SCHSUP, Schima superba; SYMANO, Symplocos anomala.
图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5天童20 hm2样地5个类群的20 m × 20 m的样方分布图。A, B, C, D, E分别表示表示类群A-E包含的样方在样地中的分布。
Fig. 5Distributions of five clusters at the scale of 20 m × 20 m in Tiantong 20 hm2 plot. A, B, C, D, E represent the distributions of the quadrats from clusters A-E, respectively.
图6
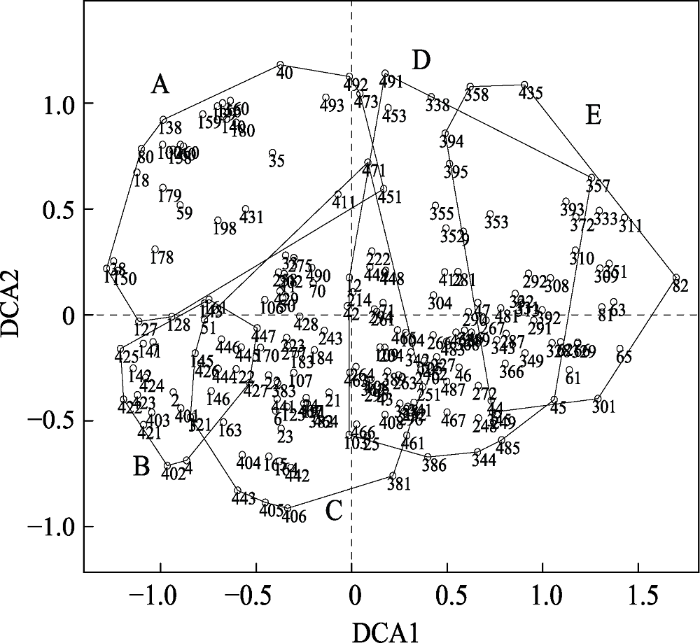 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6天童20 hm2样地内用于分类的198个样方的DCA排序图。阿拉伯数字表示样方编号。A, 类群A; B, 类群B; C, 类群C; D, 类群D; E, 类群E。
Fig. 6DCA ordination diagram for the classification of 198 quadrats in Tiantong 20 hm2 plot with the five clusters. The numbers in the figure are the identification code for each quadrat. A, B, C, D, E represent the distributions of the quadrats in clusters A-E, respectively.
将过渡区域去除后, 剩余78个样方, 共有117种木本植物, 隶属于43科76属。用TWINSPAN分类可分为3种类型(图7), 每种类型在样地中的分布情况如图8所示, 暂将各类型命名为群落1、群落2、群落3。DCA排序的结果显示前4个排序轴的特征值分别为: 0.5395、0.2399、0.1161、0.0864, 前两个排序轴的累积贡献率为76.7%, 大于70%, 可以反映事物的基本信息, 故根据前两个排序轴做二维排序图(图9), 在排序图中3种类型有明显的分布范围和界线, 排序轴1 (DCA1)反映了各样方凹凸度的梯度变化, 即沿第一轴从左向右群落分布生境的凹凸度逐渐减小, 说明样方分布的地形位置从坡面至沟谷的变化; 排序轴2 (DCA2)反映了海拔的梯度变化, 即沿第二轴由下向上, 海拔逐渐降低。
图7
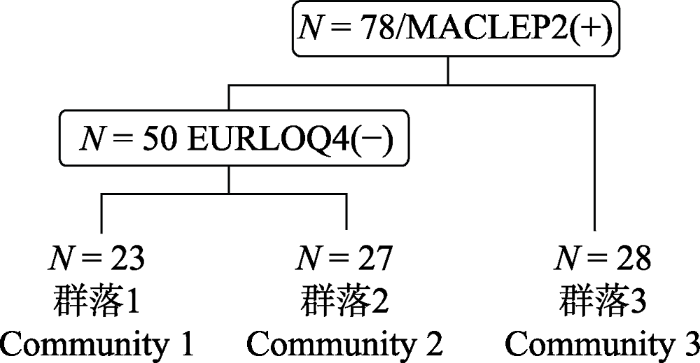 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7去除过渡群落后剩余78个样方的TWINSPAN分类结果树状示意图。N表示样方总数, EURLOQ, 细枝柃; MACLEP, 薄叶润楠。
Fig. 7The results of TWINSPAN classification for 78 quadrats after removing the transitional quadrats. N represents total number of quadrats. EURLOQ, Eurya loquaiana; MACLEP, Machilus leptophylla.
图8
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8天童20 hm2样地3种群落类型的20 m × 20 m的样方分布图。
 ,
,  ,
,  分别表示群落1、群落2、群落3中包含的样方在样地中的分布。
分别表示群落1、群落2、群落3中包含的样方在样地中的分布。Fig. 8Distributions of three community types at the scale of 20 m × 20 m in Tiantong 20 hm2 plot.
 ,
, , and
, and  represent the distributions of the communities 1-3, respectively.
represent the distributions of the communities 1-3, respectively.图9
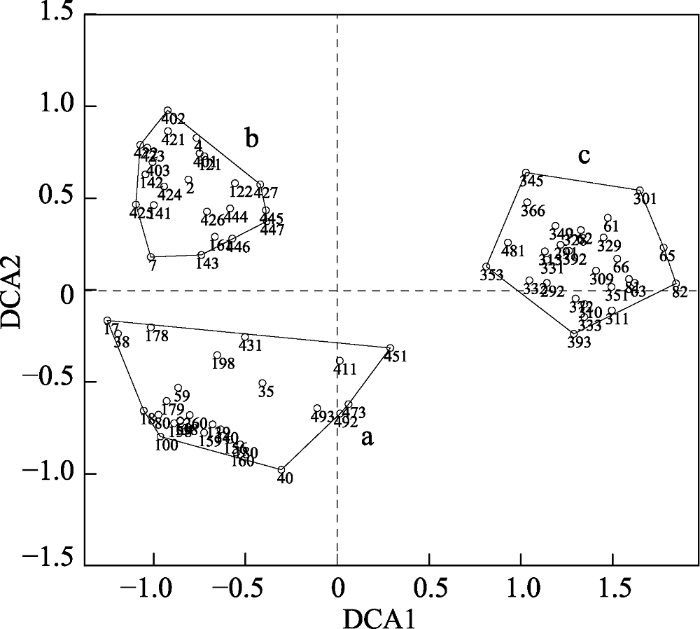 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9去除过渡群落后剩余78个样方的DCA排序图。阿拉伯数字表示样方编号。a, 群落1; b, 群落2; c, 群落3。
Fig. 9DCA ordination diagram of 78 quadrats after removing the transitional quadrats. The numbers in the figure are the identification code for each quadrat. a, community 1; b, community 2; c, community 3.
2.2 分类结果评估
用于评估分类优良度的轮廓宽度值结果见图10。由图可见, 群落1、2、3中各样方轮廓宽度平均值分别为0.33、0.46和0.59, 3种群落类型总的平均轮廓宽度值为0.46。群落2和群落3中各样方的轮廓宽度值均为正值, 群落1中一个样方的轮廓宽度值虽为负值但仅为-0.063, 仍较接近于0, 说明各样方能够较好地与各分类类型符合, 分类结果较好。图10
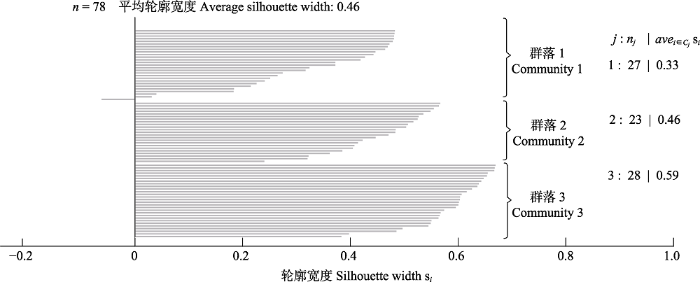 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图103种群落类型的轮廓宽度值图。每一条水平线的长度代表一个样方的轮廓宽度值。n, 样方总数; j, 各类型编号; nj, 各群落内的样方总数; avei∈cj si, 各群落内的平均轮廓宽度值。
Fig. 10The silhouette width of three community types. The length of each horizontal line represents the silhouette width of one quadrat. n, total number of quadrats; j, cluster number; nj, total number of quadrats in the community j; avei∈cj si, the average silhouette width in the community j.
2.3 群丛命名及各群丛特征
群丛命名需包括优势种和标志种两部分, 采用重要值较高的两个乔木树种作为群丛的优势种, 标志种则通过IndVal大小同时结合基于TWINSPAN分类结果的样方区分表(附录I)来确定。参考宋永昌等(2013)提出的命名原则, 标志种在前, 优势种在后, 用“/”相连, 标志种如有两个, 则以“-”相连, 重要者在前; 优势种如有两个, 以“+”相连, 也是重要者在前。结合天童地区的植被特征, 我们分别计算了各群落内物种的重要值(表1: 列出群落内重要值排前5的物种)和指示值(表2: 指示值大于0.5)。依据群丛命名原则, 结合各物种实际生态意义, 根据表1与表2中的结果将群落1、2、3分别命名为: 宜昌荚蒾-厚皮香/港柯+云山青冈群丛(Viburnum erosum-Ternstroemia gymnanthera/Lithocarpus harlandii + Cyclobalanopsis sessilifolia Ass.); 虎皮楠-柯/木荷+米槠群丛(Daphniphyllum oldhami-Lithocarpus glaber/SchimaSuperba + Castanopsis carlesii Ass.); 红毒茴-紫楠/薄叶润楠+南酸枣群丛(Illicium lanceolatum-Phoebe sheareri/Machilus leptophylla + Choerospondias axillaris Ass.)。各群丛在样地内的分布如图8所示。各群丛主要特征如下:Table 1
表1
表1天童20 hm2样地3种群落类型内物种的重要值排序(前5)
Table 1
| 物种 Species | 重要值 Important value (%) | 生长型 Growth form |
|---|---|---|
| 群落1 Community 1 | ||
| 港柯 Lithocarpus harlandii | 12.20 | 乔木 Tree |
| 杨梅叶蚊母树 Distylium myricoides | 10.97 | 小乔木 Small tree |
| 云山青冈 Cyclobalanopsis sessilifolia | 9.27 | 乔木 Tree |
| 木荷 Schima superba | 6.10 | 乔木 Tree |
| 马银花 Rhododendron ovatum | 6.06 | 小乔木 Small tree |
| 群落2 Community 2 | ||
| 细枝柃 Eurya loquaiana | 13.38 | 小乔木 Small tree |
| 木荷 Schima superba | 12.46 | 乔木 Tree |
| 杨梅叶蚊母树 Distylium myricoides | 12.12 | 小乔木 Small tree |
| 米槠 Castanopsis carlesii | 10.04 | 乔木 Tree |
| 栲 Castanopsis fargesii | 6.01 | 乔木Tree |
| 群落3 Community 3 | ||
| 黄丹木姜子 Litsea elongata | 21.46 | 小乔木 Small tree |
| 薄叶润楠 Machilus leptophylla | 12.06 | 乔木 Tree |
| 南酸枣 Choerospondias axillaris | 10.48 | 乔木 Tree |
| 细枝柃 Eurya loquaiana | 9.34 | 小乔木 Small tree |
| 云山青冈 Cyclobalanopsis sessilifolia | 5.90 | 乔木 Tree |
新窗口打开|下载CSV
Table 2
表2
表23种群落类型内物种指示值(大于0.5)排序
Table 2
| 物种 Species | 特异性Specificity | 确限度Fidelity | 指示值Indicator value | p |
|---|---|---|---|---|
| 群落1 Community 1 | ||||
| 杜鹃 Rhododendron simsii | 0.9659 | 0.7037 | 0.824 | 0.001*** |
| 宜昌荚蒾 Viburnum erosum | 0.9298 | 0.5556 | 0.719 | 0.001*** |
| 厚皮香 Ternstroemia gymnanthera | 0.9643 | 0.4444 | 0.655 | 0.001*** |
| 迎春樱桃 Cerasus discoidea | 1.0000 | 0.3704 | 0.609 | 0.001*** |
| 山鸡椒 Litsea cubeba | 0.9070 | 0.4074 | 0.608 | 0.002** |
| 青冈 Cyclobalanopsis glauca | 0.8421 | 0.3704 | 0.558 | 0.003** |
| 大青 Clerodendrum cyrtophyllum | 0.8750 | 0.3333 | 0.540 | 0.007** |
| 群落2 Community 2 | ||||
| 虎皮楠 Daphniphyllum oldhami | 0.9894 | 0.6957 | 0.830 | 0.001*** |
| 柯 Lithocarpus glaber | 0.9792 | 0.6087 | 0.772 | 0.001*** |
| 杨梅 Myrica rubra | 0.8667 | 0.6087 | 0.726 | 0.001*** |
| 总状山矾 Symplocos botryantha | 0.9375 | 0.4783 | 0.670 | 0.001*** |
| 刺毛越桔 Vaccinium trichocladum | 0.8889 | 0.4783 | 0.652 | 0.001*** |
| 铁冬青 Ilex rotunda | 0.5833 | 0.5217 | 0.552 | 0.045* |
| 群落3 Community 3 | ||||
| 薄叶润楠 Machilus leptophylla | 0.9931 | 1.0000 | 0.997 | 0.001*** |
| 红毒茴 Illicium lanceolatum | 0.9695 | 0.8214 | 0.892 | 0.001*** |
| 紫楠 Phoebe sheareri | 1.0000 | 0.5714 | 0.756 | 0.001*** |
| 黄牛奶树 Symplocos cochinchinensis var. laurina | 0.8482 | 0.6429 | 0.738 | 0.001*** |
| 西川朴 Celtis vandervoetiana | 1.0000 | 0.3214 | 0.567 | 0.001*** |
| 锐角枫 Acer acutum | 1.0000 | 0.2500 | 0.500 | 0.002** |
| 胡桃楸 Juglans mandshurica | 1.0000 | 0.2500 | 0.500 | 0.001*** |
新窗口打开|下载CSV
宜昌荚蒾-厚皮香/港柯+云山青冈群丛: 该群丛内样方分布的最低海拔为450 m, 平均海拔550 m, 位于山坡上部, 包含27个样方, 共有84种木本植物, 隶属于31科53属。优势种及标志种有港柯、云山青冈、杨梅叶蚊母树(Distylium myricoides)、杜鹃(Rhododendron simsii)、宜昌荚蒾、厚皮香等。该群丛27个样方(群丛个体)的Jaccard相似性系数介于0.346-0.787之间。
虎皮楠-柯/木荷+米槠群丛: 该群丛位于海拔360-455 m的低海拔山坡上, 包括23个样方, 共有68种木本植物, 隶属于24科43属。在该群丛所含样方内乔木层优势种为木荷、米槠、栲(Castanopsis fargesii)等, 下木层中细枝柃(Eurya loquaiana)、杨梅叶蚊母树也在群落中占有一定的优势, 指示值较高的树种有虎皮楠、柯等。该群丛23个样方的Jaccard相似性系数介于0.346-0.785。
红毒茴-紫楠/薄叶润楠+南酸枣群丛: 该群丛位于海拔440 m以下沟谷中, 最低海拔320 m, 这里地下水位较高, 生境较湿润, 包含28个样方, 共有83种木本植物, 隶属于34科60属。优势种包括南酸枣、黄丹木姜子(Litsea elongata)、薄叶润楠等, 标志种为薄叶润楠、红毒茴、紫楠、黄牛奶树(Symplocos cochinchinensis var. laurina)等。该群丛28个样方的Jaccard相似性系数介于0.192-0.680, 其中仅样方301的相似性系数为0.192, 该样方位于样地边缘, 样方内裸露岩石较多, 物种数仅为10种, 这可能是它与其他样方的相似性较低的原因。在去除样方301后, 该群丛的Jaccard相似性系数介于0.235-0.680。
2.4 各群丛与环境间关系
为了更直观地显示每个环境因子对群丛分布的影响, 将海拔、凹凸度、坡度与坡向4个环境因子被动加入DCA排序图(图9)中, 结果见图11。图中箭头表示环境因子, 箭头所处的象限表示环境因子与排序轴间的正负相关性, 箭头连线的长度代表某个环境因子与群落分布和种类分布相关程度的大小, 连线越长, 说明相关性越大, 反之越小。从图中可以看出海拔对群丛分布的影响最大, 其次是凹凸度, 坡度和坡向对群丛分布影响较小。凹凸度与第一轴的方向相反, 表明沿第一轴从左向右, 凹凸度逐渐降低, 海拔与第二轴的方向相反, 表明沿第二轴从下到上, 海拔逐渐降低, 说明位于排序图左下方的宜昌荚蒾-厚皮香/港柯+云山青冈群丛分布在较高海拔坡面上, 位于排序图左上方的虎皮楠-柯/木荷+米槠群丛分布在低海拔坡面上, 位于排序图左侧的红毒茴-紫楠/南酸枣+薄叶润楠群丛适宜于沟谷生境中。我们所划分的3个群丛不仅有相对一致的植物种类组成, 也分布在相对一致的生境中。图11
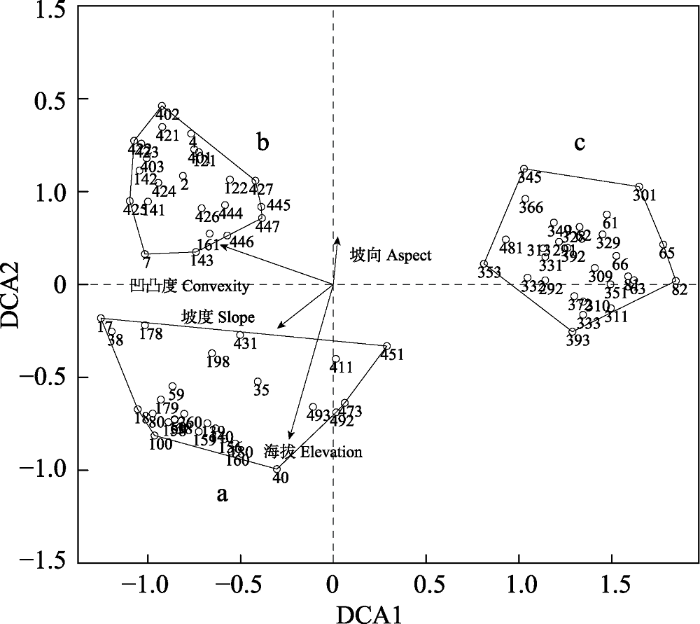 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1178个样方的DCA排序轴与环境因子的相关性图。阿拉伯数字表示样方编号。a, 宜昌荚蒾-厚皮香/港柯+云山青冈群丛; b, 虎皮楠-柯/木荷+米槠群丛; c, 红毒茴-紫楠/南酸枣+薄叶润楠群丛。
Fig. 11DCA ordination diagram of 78 quadrats with environmental variables. The numbers in the figure are the identification code for each quadrat. a, Viburnum erosum-Ternstroemia gymnanthera/Lithocarpus harlandii + Cyclobalanopsis sessilifolia Association; b, Daphniphyllum oldhami-Lithocarpus glaber/Schima superba + Castanopsis carlesii Association; c, Illicium lanceolatum-Phoebe sheareri/Choerospondias axillaris + Machilus leptophylla Association.
3 讨论
植被在时间和空间上是随环境不断变化的, 并不存在清楚明确的界限, 亦即群落是连续的, 但在植被的连续之中也有间断(宋永昌, 2017)。对于森林植被而言, 在连续分布中, 过渡群落是客观存在的, 它会对确定典型群丛类型产生影响。本文对未去除过渡类型的5个类群进行了IndVal分析(结果见表3), 发现类群A (群落1)、类群B (群落2)、类群E (群落3)中各标志种的IndVal明显低于去除过渡区域后的各群落中标志种的IndVal, 且类群C和类群D没有独立的标志种, 它们就是我们通过DCA排序分析得出的过渡群落。当去除过渡类型后, 各群落类型中的标志种与TWINSPAN分类结果(附录I)中得到的区别种(即标志种)能够较好吻合。由此可见, 过渡类型的存在对分类结果影响很大, 不利于典型群丛的确定。但过渡群落又是客观存在的, 对这部分群落如何处理是一个值得研究的问题。据前人研究结果, 基于以下几点可将过渡群落划分到已确定的群丛中: (1)用排序或聚类方法进行相似性度量, (2)基于标志种的出现以及多度或组成, (3)根据生境和群落外貌特征(Jennings et al., 2009; van der Maarel & Franklin, 2013)。此外, Mueller-Dombois和Ellenberg (1974)也曾提议, 根据Jaccard有-无指数(与最典型样地的相似性)在25%到50%之间的群落地段可以成为同一群丛的一部分(Mueller-Dombois & Ellenberg,1974), 具体的量化指标有待进一步研究。此外, 由于干扰(包括自然干扰和人为干扰)造成样地内植被的不同质性, 为识别植被类型造成了困扰, 在植被分类时需尽量避免, 故本文将林窗以及人为干扰影响较大的群落地段去除后再进行群丛划分。Table 3
表3
表35个类群内物种指示值排序
Table 3
| 物种 Species | 特异性Specificity | 确限度Fidelity | 指示值Indicator value | p | ||
|---|---|---|---|---|---|---|
| 类群A Cluster A | ||||||
| 杜鹃 Rhododendron simsii | 0.8788 | 0.6667 | 0.765 | 0.001*** | ||
| 宜昌荚蒾 Viburnum erosum | 0.8281 | 0.5000 | 0.643 | 0.001*** | ||
| 厚皮香 Ternstroemia gymnanthera | 0.8056 | 0.5000 | 0.635 | 0.001*** | ||
| 山鸡椒 Litsea cubeba | 0.5362 | 0.4000 | 0.463 | 0.001*** | ||
| 迎春樱桃 Cerasus discoidea | 0.6071 | 0.3333 | 0.450 | 0.001*** | ||
| 南烛 Vaccinium bracteatum | 0.5143 | 0.3333 | 0.414 | 0.003** | ||
| 豆腐柴 Premna microphylla | 0.6000 | 0.1333 | 0.283 | 0.045* | ||
| 野鸦椿 Euscaphis japonica | 0.7500 | 0.1000 | 0.274 | 0.026* | ||
| 木蜡树 Toxicodendron sylvestre | 1.0000 | 0.0667 | 0.258 | 0.029* | ||
| 类群B Cluster B | ||||||
| 柯 Lithocarpus glaber | 0.7833 | 0.6087 | 0.691 | 0.001*** | ||
| 杨梅 Myrica rubra | 0.6047 | 0.6087 | 0.607 | 0.001*** | ||
| 刺毛越桔 Vaccinium trichocladum | 0.5333 | 0.4783 | 0.505 | 0.001*** | ||
| 总状山矾 Symplocos botryantha | 0.4688 | 0.4783 | 0.473 | 0.001*** | ||
| 类群E Cluster E | ||||||
| 紫楠 Phoebe sheareri | 0.9000 | 0.5806 | 0.723 | 0.001*** | ||
| 胡桃楸 Juglans mandshurica | 0.9167 | 0.2581 | 0.486 | 0.001*** | ||
| 锐角槭 Acer acutum | 0.8333 | 0.2258 | 0.434 | 0.001*** | ||
| 中华卫矛 Euonymus nitidus | 0.7000 | 0.1613 | 0.336 | 0.006** | ||
| 枳椇 Hovenia acerba | 0.8750 | 0.1290 | 0.336 | 0.002** | ||
| 类群B + C Cluster B + C | ||||||
| 虎皮楠 Daphniphyllum oldhami | 0.8685 | 0.5889 | 0.715 | 0.001*** | ||
| 皱柄冬青 Ilex kengii | 0.7812 | 0.1889 | 0.384 | 0.013* | ||
| 类群D + E Cluster D + E | ||||||
| 薄叶润楠 Machilus leptophylla | 0.9809 | 0.7436 | 0.854 | 0.001*** | ||
| 红毒茴 Illicium lanceolatum | 0.9031 | 0.6539 | 0.768 | 0.001*** | ||
| 黄牛奶树 Symplocos cochinchinensis var. laurina | 0.8273 | 0.5513 | 0.675 | 0.001*** | ||
| 西川朴 Celtis vandervoetiana | 0.9730 | 0.3077 | 0.547 | 0.001*** | ||
| 无患子 Sapindus saponaria | 0.8529 | 0.2564 | 0.468 | 0.001*** | ||
| 糙叶树 Aphananthe aspera | 0.8846 | 0.2308 | 0.452 | 0.001*** | ||
| 紫弹树 Celtis biondii | 0.8889 | 0.1923 | 0.413 | 0.003** | ||
| 杭州榆 Ulmus changii | 0.9853 | 0.1410 | 0.373 | 0.007** | ||
| 青钱柳 Cyclocarya paliurus | 0.8667 | 0.1282 | 0.333 | 0.009** | ||
| 大叶旱樱 Cerasus subhirtella | 0.8750 | 0.0897 | 0.280 | 0.04* | ||
| 黐花 Mussaenda esquirolii | 1.0000 | 0.0769 | 0.277 | 0.036* | ||
新窗口打开|下载CSV
群丛是植被分类的基本单位, 由于各个学派的分类途径不同, 植物群丛的定义各有侧重, 如法瑞学派强调特征种, 苏联学派强调各层优势种, 美国国家植被分类系统强调鉴别种等, 但总的来看, 植物群丛应该包括4个主要特点: (1)一致的群落外貌和结构, (2)一致的生境, (3)确定的植物种类组成, (4)在一个景观或区域内重复出现(Jennings et al., 2009)。本文所划分的3个群丛与以上4个基本特点相契合: (1)宜昌荚蒾-厚皮香/港柯+云山青冈群丛及虎皮楠-柯/木荷+米槠群丛属于常绿阔叶林, 红毒茴-紫楠/南酸枣+薄叶润楠群丛属于常绿落叶阔叶混交林, 其群落外貌存在明显差异; (2)从3个群丛在样地中的分布(图8)及DCA排序轴与环境因子的相关性(图11)可以看出, 各群丛分布的生境内部基本一致, 但各群丛之间差异明显。另外, 标志种对环境有指示意义, 也保证了不同群丛具有各自一致的生态特征、生境条件和动态特点; (3)从平均轮廓宽度值(图10)可以看出, 3个群丛各自内部种类组成相似, 群丛之间差异明显, 同时3个群丛各自的Jaccard相似性系数也表明各群丛内部具有大体一致的植物种类组成。各群丛内物种的重要值(表1)及指示值(表2)也证明其各自具有独立的优势种及标志种; (4)从3个群丛在样地中的分布(图8)可以看出, 同一群丛的样方并不完全集中在一起, 而是间断地分布在相似的生境中。同时也满足确定一个群丛至少需要5个以上群落样地资料的要求(宋永昌, 2017)。
TWINSPAN分类是迄今为止最受欢迎的一种数量分类方法(van der Maarel & Franklin, 2013), 适于揭示植物群落的间断性, DCA排序可展示植物群落的连续性。本文首先用TWINSPAN将植被划分成类群, 再用DCA排序展示各类群之间的相互关系, 可以明显发现有些类群与其他类群之间有重叠部分, 过渡群落去除后, 再重新用TWINSPAN划分群落, 发现DCA排序更能明确显示群落间断性, 说明两者结合能更直观地发现连续群落中的过渡群落, 该方法具有较高实用性。将本文所划分的群丛类型与《浙江天童国家森林公园的植被和区系》(宋永昌和王祥荣, 1995)一书中, 对天童国家森林公园内分布在海拔300-600 m的植被所划分出的群丛进行对比, 可以发现其中的港柯° 云山青冈群丛与本文的宜昌荚蒾-厚皮香/港柯+云山青冈群丛, 木荷° 栲树群丛(含杨梅叶蚊母树的亚群丛)与本文的虎皮楠-柯/木荷+米槠群丛, 红毒茴? 南酸枣群丛与本文的红毒茴-紫楠/薄叶润楠+南酸枣群丛, 两两之间具有大体一致的植物种类组成, 优势种与标志种基本相同, 同时也分布在相似的生境中。这表明本文所划分出的群丛与前人研究结果能较好地吻合, 由于本文采用了数量分析方法, 使得分类结果具有更高的客观性。
大型动态监测样地在进行群落调查时是按种记录DBH ≥ 1 cm的每一个体的胸径, 而不是像一般森林群落调查中分层(乔木层、灌木层及草本层)记录每一种多度、显著度/盖度, 在不分层情况下, 包含相对频度计算的重要值会放大小个体、分布广的种的重要性, 弱化大个体、有限分布种的重要性。Mueller-Dombois和Ellenberg (1974)在《植被生态学的目的与方法》一书中也曾在谈论重要值计算的定量参数选择时提出频度对下木层中的树苗意义更大, 而只用相对优势度/相对胸高断面积来衡量物种在样方中的作用, 又会强化大个体在样地内的重要性。故本文中, 重要值计算我们采用了相对多度与相对胸高断面积两个参数。在国内其他对大样地进行的植被分类研究中, 例如, 用TWINSPAN方法对台湾福山25 hm2样地进行的植被类型划分研究(Su et al., 2007), 用多元回归树(MRT)对古田山24 hm2样地、弄岗15 hm2样地进行的群丛划分研究(赖江山等, 2010; 黄甫昭等, 2014), 重要值计算也都是采用的多度与胸高断面积参数。此外, 从以往的群丛划分结果来看, 群丛的标志种并非都是木本植物, 有可能是草本植物, 而大样地调查记录大都缺少草本层调查, 这对于研究群丛分类是一缺陷(宋永昌等, 2015), 本研究也不例外, 希望在将来的研究中得到补充和改进。
综上所述, 植物群落既是连续的, 也是间断的, 本研究表明过渡群落的去除对确定典型群丛具有重要意义; 群丛是植被分类的基本单位, 群落种类组成, 包括优势种和标志种是划分群丛的基础, 与环境因子相结合是对所划分群丛结果的有效检验; 此外, 将基于植物群落种类组成的数量分类与DCA排序结合进行群丛类型划分有利于更加直观地发现过渡群落; 利用数量分类作为工具进行植被类型划分是当前植被分类发展的趋势。
附件
附录I 天童20 hm2样地内78个样方区分表-基于双向指示种分类结果Supplement I Differential table of 78 quadrats in Tiantong 20 hm2 plot-based on the result of TWINSPAN

致谢
宋永昌教授对本文进行多次审阅, 并提出宝贵修改意见, 特此致谢!参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
DOIURLPMID [本文引用: 2]

Ecologists often face the task of studying the association between single species and one or several groups of sites representing habitat types, community types, or other categories. Besides characterizing the ecological preference of the species, the strength of the association usually presents a lot of interest for conservation biology, landscape mapping and management, and natural reserve design, among other applications. The indices most frequently employed to assess these relationships are the phi coefficient of association and the indicator value index (IndVal). We compare these two approaches by putting them into a broader framework of related measures, which includes several new indices. We present permutation tests to assess the statistical significance of species-site group associations and bootstrap methods for obtaining confidence intervals. Correlation measures, such as the phi coefficient, are more context-dependent than indicator values but allow focusing on the preference of the species. In contrast, the two components of an indicator value index directly assess the value of the species as a bioindicator because they can be interpreted as its positive predictive value and sensitivity. Ecologists should select the most appropriate index of association strength according to their objective and then compute confidence intervals to determine the precision of the estimate.
DOIURL [本文引用: 2]
[本文引用: 2]
DOIURL [本文引用: 1]
植物群落是不同植物在长期环境变化中相互作用、相互适应而形成的组合.它提供着人类赖以生存的主要物质资源,维系着地球生态系统的健康和功能,也为各种动物和其他生物提供食物来源和栖息地,是人类生存和发展不可或缺的物质基础,具有不可替代的作用.我国植物群落类型多样,在世界上首屈一指,但我国至今尚没有一次全面和系统的植物群落清查,不仅影响了人们对我国植物资源的了解、利用和保护,也不利于我国生态学、环境科学和地理学等相关学科的发展.采用统一的方法体系和技术规范开展我国植物群落的清查工作势在必行,并具有紧迫性.本文基于作者长期的野外工作实践和国内外的群落调查方法,首先简要定义了与植物群落清查有关的重要概念,在此基础上,论述了调查样地的设置原则和体系、群落清查的技术指标和方法、主要优势种生态属性的测定方法和规范,并介绍了大样地调查的主要步骤.通过本文的介绍、归纳和总结,试图为制定我国植物群落清查的技术规范提供基础材料和技术储备.
DOIURL [本文引用: 1]
植物群落是不同植物在长期环境变化中相互作用、相互适应而形成的组合.它提供着人类赖以生存的主要物质资源,维系着地球生态系统的健康和功能,也为各种动物和其他生物提供食物来源和栖息地,是人类生存和发展不可或缺的物质基础,具有不可替代的作用.我国植物群落类型多样,在世界上首屈一指,但我国至今尚没有一次全面和系统的植物群落清查,不仅影响了人们对我国植物资源的了解、利用和保护,也不利于我国生态学、环境科学和地理学等相关学科的发展.采用统一的方法体系和技术规范开展我国植物群落的清查工作势在必行,并具有紧迫性.本文基于作者长期的野外工作实践和国内外的群落调查方法,首先简要定义了与植物群落清查有关的重要概念,在此基础上,论述了调查样地的设置原则和体系、群落清查的技术指标和方法、主要优势种生态属性的测定方法和规范,并介绍了大样地调查的主要步骤.通过本文的介绍、归纳和总结,试图为制定我国植物群落清查的技术规范提供基础材料和技术储备.
URL
DOIURL [本文引用: 1]
Summary 1 Tests of habitat association among species of tropical trees and shrubs often assume that individual stems can be treated as independent sample units, even though limited dispersal conflicts with this assumption by causing new recruits to occur near maternal parents and siblings. 2 We developed methods for assessing patterns of association between mapped plants and mapped habitat types that explicitly incorporate spatial structure, thereby eliminating the need to assume independence among stems. 3 We used these methods to determine habitat-association patterns for 171 species of trees and shrubs within the permanent 50-ha Forest Dynamics Project plot on Barro Colorado Island, Panama. 4 Many fewer significant habitat associations result from the new methods than from traditional, but inappropriate, chi-square tests. The low-lying plateau, the most extensive habitat on the 50-ha plot, had nine species positively associated with it and 19 species negatively associated, leaving 143 species whose distributions were not biased with respect to this habitat. A small swamp in the plot was the most distinct habitat, with 32 species positively and 20 species negatively associated, leaving more than two-thirds of the species neither positively nor negatively associated. 5 To the extent that habitat association reflects habitat specialization, our results suggest that local habitat specialization plays a limited role in the maintenance of species diversity in this forest.
[本文引用: 1]
DOIURL [本文引用: 1]
以弄岗15 ha北热带喀斯特季节性雨林监测样地各样方的相对海拔、坡度、坡向、凹凸度和物种重要值数据为变量,采用多元回归树方法将喀斯特季节性雨林森林群落分为8 个群丛.同时,按乔木层、亚乔木层优势种为主,灌木层指示种为辅的方法分别把8类群丛命名为:(A)南方紫金牛(Ardisia thyrsiflora)-苹婆(Sterculia monosperma)-中国无忧花(Saraca dives)群丛,该群丛类型主要分布于土壤和空气的湿度都较大的谷底边缘;(B)对叶榕(Ficus hispida)-苹婆-董棕(Caryota obtusa)群丛,该群丛类型主要分布于伴有季节性水淹、土壤和空气湿度都很大的谷底;(C)山石榴(Catunaregam spinosa)-广西牡荆-蚬木(Excentrodendron tonkinense)群丛,该群丛类型主要分布于受太阳直射时间较长、水分条件中等偏旱的中下坡阳坡地带;(D)垂茉莉(Clerodendrum wallichii)-苹婆-广西棋子豆(Cychidendron guangxiensis)群丛,该群丛类型主要分布于受太阳直射时间稍短、水分条件适中的中下坡阴坡地带;(E)米仔兰(Aglaia odorata)-闭花木(Cleistanthus sumatranus)-蚬木群丛,该群丛类型主要分布于土壤湿度中等偏旱的中上坡半阴坡地带;(F)割舌树(Walsura robusta)-闭花木-蚬木群丛,该群丛类型主要分布于土壤湿度中等偏旱、比较陡峭的半阳坡地带;(G)枝花流苏树(Chionanthus ramiflorus)-广西牡荆-蚬木群丛,该群丛类型主要分布于坡度缓和、太阳直射的时间较短、水分条件适中的垭口部位;(H)齿叶黄皮 (Clausena dunniana)-黄梨木Boniodendron minius)-蚬木群丛,该群丛类型主要分布于受到太阳直射时间最长、岩石裸露度最大、气温干燥、土壤水分严重不足的山顶及其周围.群丛划分说明该区域 植被差异主要源于相对海拔和坡向变化,各群丛物种组成并不存在明显界限,而是随环境梯度逐渐变化,反映了植被分布具有连续性,间断只是相对的.
DOIURL [本文引用: 1]
以弄岗15 ha北热带喀斯特季节性雨林监测样地各样方的相对海拔、坡度、坡向、凹凸度和物种重要值数据为变量,采用多元回归树方法将喀斯特季节性雨林森林群落分为8 个群丛.同时,按乔木层、亚乔木层优势种为主,灌木层指示种为辅的方法分别把8类群丛命名为:(A)南方紫金牛(Ardisia thyrsiflora)-苹婆(Sterculia monosperma)-中国无忧花(Saraca dives)群丛,该群丛类型主要分布于土壤和空气的湿度都较大的谷底边缘;(B)对叶榕(Ficus hispida)-苹婆-董棕(Caryota obtusa)群丛,该群丛类型主要分布于伴有季节性水淹、土壤和空气湿度都很大的谷底;(C)山石榴(Catunaregam spinosa)-广西牡荆-蚬木(Excentrodendron tonkinense)群丛,该群丛类型主要分布于受太阳直射时间较长、水分条件中等偏旱的中下坡阳坡地带;(D)垂茉莉(Clerodendrum wallichii)-苹婆-广西棋子豆(Cychidendron guangxiensis)群丛,该群丛类型主要分布于受太阳直射时间稍短、水分条件适中的中下坡阴坡地带;(E)米仔兰(Aglaia odorata)-闭花木(Cleistanthus sumatranus)-蚬木群丛,该群丛类型主要分布于土壤湿度中等偏旱的中上坡半阴坡地带;(F)割舌树(Walsura robusta)-闭花木-蚬木群丛,该群丛类型主要分布于土壤湿度中等偏旱、比较陡峭的半阳坡地带;(G)枝花流苏树(Chionanthus ramiflorus)-广西牡荆-蚬木群丛,该群丛类型主要分布于坡度缓和、太阳直射的时间较短、水分条件适中的垭口部位;(H)齿叶黄皮 (Clausena dunniana)-黄梨木Boniodendron minius)-蚬木群丛,该群丛类型主要分布于受到太阳直射时间最长、岩石裸露度最大、气温干燥、土壤水分严重不足的山顶及其周围.群丛划分说明该区域 植被差异主要源于相对海拔和坡向变化,各群丛物种组成并不存在明显界限,而是随环境梯度逐渐变化,反映了植被分布具有连续性,间断只是相对的.
DOIURL [本文引用: 4]
This article provides guidelines for the description, documentation, and review of proposals for new or revised plant associations and alliances to be recognized as units of vegetation within the U.S. National Vegetation Classification (NVC). By setting forth standards for field records, analysis, description, peer review, and archiving, the Ecological Society of America's Vegetation Classification Panel, in collaboration with the U.S. Federal Geographic Data Committee, NatureServe, and others, seeks to advance our common understanding of vegetation and improve our capability to sustain and restore natural systems. We provide definitions for the two floristic levels of the NVC hierarchy: associations and alliances. This is followed by a description of standards for field plot records and the identification and classification of vegetation types. Procedures for review and evaluation of proposed additions and revisions of types are provided, as is a structure for data archiving and dissemination. These procedures provide a dynamic and practical way to publish new or revised descriptions of vegetation types while maintaining a current, authoritative list of types for multiple users to access and apply.
DOIURL [本文引用: 1]
常绿阔叶林植被分类一直是植被生态学研究中的难题,尤其是基本单位——群丛的分类。该文以地形因子和物种组成数据为变量的多元回归树方法,将浙江古田山24hm2森林监测样地的森林群落分为3个群丛。所得群丛既反映了群落在时间和空间上的相对间断分布,也符合植被分类基本单位的特点,为常绿阔叶林的群丛分类提供了新思路。并首次引入了指示值算法,解决了以往指示种无法量化的难题。最终以优势层优势种为主、下木层指示种为辅的命名原则,将3个群丛命名为:1)石斑木(Raphiolepis indica)+柳叶蜡梅(Chimonanthus salicifolius)-格药柃(Eurya muricata)+赤楠(Syzygium buxifolium)-木荷(Schima superba)+甜槠(Castanopsis eyrei)群丛;2)映山红(Rhododendron simsii)+满山红(Rhododendron mariesii)-短柄枹(Quercus serrata var.brevipetiolata)+灰白蜡瓣花(Corylopsis glandulifera var.hypoglauca)-马尾松(Pinus massoniana)+甜槠群丛;3)短尾越桔(Vaccinium carlesii)+毛花连蕊茶(Camellia fraterna)-杨梅叶蚊母树(Distylium myricoides)+浙江新木姜子(Neolitsea aurata)-木荷+甜槠群丛。
DOIURL [本文引用: 1]
常绿阔叶林植被分类一直是植被生态学研究中的难题,尤其是基本单位——群丛的分类。该文以地形因子和物种组成数据为变量的多元回归树方法,将浙江古田山24hm2森林监测样地的森林群落分为3个群丛。所得群丛既反映了群落在时间和空间上的相对间断分布,也符合植被分类基本单位的特点,为常绿阔叶林的群丛分类提供了新思路。并首次引入了指示值算法,解决了以往指示种无法量化的难题。最终以优势层优势种为主、下木层指示种为辅的命名原则,将3个群丛命名为:1)石斑木(Raphiolepis indica)+柳叶蜡梅(Chimonanthus salicifolius)-格药柃(Eurya muricata)+赤楠(Syzygium buxifolium)-木荷(Schima superba)+甜槠(Castanopsis eyrei)群丛;2)映山红(Rhododendron simsii)+满山红(Rhododendron mariesii)-短柄枹(Quercus serrata var.brevipetiolata)+灰白蜡瓣花(Corylopsis glandulifera var.hypoglauca)-马尾松(Pinus massoniana)+甜槠群丛;3)短尾越桔(Vaccinium carlesii)+毛花连蕊茶(Camellia fraterna)-杨梅叶蚊母树(Distylium myricoides)+浙江新木姜子(Neolitsea aurata)-木荷+甜槠群丛。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
[本文引用: 3]
DOIURL [本文引用: 1]
URL [本文引用: 2]
DOIURL [本文引用: 1]
A new graphical display is proposed for partitioning techniques. Each cluster is represented by a so-called silhouette, which is based on the comparison of its tightness and separation. This silhouette shows which objects lie well within their cluster, and which ones are merely somewhere in between clusters. The entire clustering is displayed by combining the silhouettes into a single plot, allowing an appreciation of the relative quality of the clusters and an overview of the data configuration. The average silhouette width provides an evaluation of clustering validity, and might be used to select an ‘appropriate’ number of clusters.
DOIURL [本文引用: 1]
《中国植被志》的编写工作已经启动,藉此征求修订《中国植被分类系统》意见之际,提出几点看法与建 议:1)贯彻原先制定的中国植被“高级分类单位偏重于生态外貌,而中、低级分类单位则着重种类组成和群落结构”的分类原则.这一思想与当前国际上植被分类 的发展是一致的.2)中国植被分类既要符合中国植被的特点,又要适应全球植被分类的发展,名词概念应尽量与国际上的广泛理解相一致.3)“群丛”是《中国 植被志》描述的基本对象,其概念需要统一,以避免歧义和可能导致的南北“群丛”的不等质.资料不足的类型应组织野外补点调查.4)植被分类等级系统是严格 的,但各等级又是开放的,可适当地增设高级分类单位,以适应类型扩展之需要.该文附有从植被型纲到植被型的建议草案.
DOIURL [本文引用: 1]
《中国植被志》的编写工作已经启动,藉此征求修订《中国植被分类系统》意见之际,提出几点看法与建 议:1)贯彻原先制定的中国植被“高级分类单位偏重于生态外貌,而中、低级分类单位则着重种类组成和群落结构”的分类原则.这一思想与当前国际上植被分类 的发展是一致的.2)中国植被分类既要符合中国植被的特点,又要适应全球植被分类的发展,名词概念应尽量与国际上的广泛理解相一致.3)“群丛”是《中国 植被志》描述的基本对象,其概念需要统一,以避免歧义和可能导致的南北“群丛”的不等质.资料不足的类型应组织野外补点调查.4)植被分类等级系统是严格 的,但各等级又是开放的,可适当地增设高级分类单位,以适应类型扩展之需要.该文附有从植被型纲到植被型的建议草案.
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 2]
URL [本文引用: 2]
DOIURL [本文引用: 1]
本文通过汇总中国常绿阔叶林8大动态监测样地的植物名录,在规整植物名称的基础上,统计了8大样地的植物种类组成,比较了它们的相似性及其与环境立地的关系。利用生长型和生态指示值,分析了8大样地组成植物的生态特征及其与生境的联系。从区系组成和生态特征看,8大样地可分为两组,一组包括天童、古田山、百山祖、八大公山,属于典型常绿阔叶林;另一组包括黑石顶、鼎湖山、福山、莲花池,属于季节常绿阔叶林和适雨常绿阔叶林(亚热带雨林)。另外还讨论了动态观测样地的群落类型划分问题。最后提出的建议是:增加草本和藤本植物调查,重视植物分类和生态指示值及种间相互关系的研究,更为重要的是制定长期的协作研究计划。
DOIURL [本文引用: 1]
本文通过汇总中国常绿阔叶林8大动态监测样地的植物名录,在规整植物名称的基础上,统计了8大样地的植物种类组成,比较了它们的相似性及其与环境立地的关系。利用生长型和生态指示值,分析了8大样地组成植物的生态特征及其与生境的联系。从区系组成和生态特征看,8大样地可分为两组,一组包括天童、古田山、百山祖、八大公山,属于典型常绿阔叶林;另一组包括黑石顶、鼎湖山、福山、莲花池,属于季节常绿阔叶林和适雨常绿阔叶林(亚热带雨林)。另外还讨论了动态观测样地的群落类型划分问题。最后提出的建议是:增加草本和藤本植物调查,重视植物分类和生态指示值及种间相互关系的研究,更为重要的是制定长期的协作研究计划。
DOIURL [本文引用: 7]
在《对中国植被分类系统的认知和建议》(简称《认知和建议》)的基础上,参考近期国内外植被分类研究成果,再次讨论了中国植被分类系统和单位,统一了各级单位划分依据,增补了高、中、低各等级分类的具体建议,并对《认知和建议》一文中相关部分,特别是植被型一级做了修订。将该修订方案与联合国教科文组织《世界植被分类与制图》、美国联邦地理数据委员会《美国植被分类规范》,以及Braun-Blanquet的分类方案进行对比,以便于同行间相互了解和交流。
DOIURL [本文引用: 7]
在《对中国植被分类系统的认知和建议》(简称《认知和建议》)的基础上,参考近期国内外植被分类研究成果,再次讨论了中国植被分类系统和单位,统一了各级单位划分依据,增补了高、中、低各等级分类的具体建议,并对《认知和建议》一文中相关部分,特别是植被型一级做了修订。将该修订方案与联合国教科文组织《世界植被分类与制图》、美国联邦地理数据委员会《美国植被分类规范》,以及Braun-Blanquet的分类方案进行对比,以便于同行间相互了解和交流。
[本文引用: 1]
DOIURL [本文引用: 1]
1 We mapped and identified all trees greater than or equal to 10 mm in diameter in 25 ha of lowland wet forest in Amazonian Ecuador, and found 1104 morphospecies among 152 353 individuals. The largest number of species was mid-sized canopy trees with maximum height 10-20 m and understorey treelets with maximum height of 5-10 m.
[本文引用: 6]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
DOIURL [本文引用: 1]
位于亚热带的浙江天童和古田山常绿阔叶林大样地分布有较高比例的落叶树种,那么它们与常绿树种的共存机制是什么?常绿树种和落叶树种生态习性差异较大,二者对生境的选择应有所不同,我们推测生境分化可能是两类植物实现共存的主要机制。为检验该假设,我们以天童20ha动态样地调查数据为依托,选择个体数≥20的55个常绿树种和42个落叶树种作为分析对象,用典范对应分析(CCA)研究了地形因子对二者分布的影响差异,用torus转换检验来分析常绿树种和落叶树种与各类地形生境的关联。结果如下:(1)CCA分析表明地形因子对常绿树种分布的解释量为19.2%,对落叶树种分布的解释量为7.0%。(2)torus转换检验结果表明,与沟谷成正关联的常绿树种和落叶树种的比例分别为16.4%和28.6%,成负关联的比例分别为40%和7%;与山脊成正关联的常绿树种和落叶树种的比例分别为41.8%和4.8%,成负关联的比例分别为10.9%和47.6%;与受干扰生境成正关联的常绿树种和落叶树种的比例分别为16.4%和42.9%。上述结果说明地形对常绿树种分布的影响大于落叶树种;两个植物类群对生境的选择多呈现相反格局,尤其是在沟谷生境和山脊生境,这进一步表明生境分化是常绿树种和落叶树种共存的重要机制之一,生态位理论在一定程度上能较好地解释亚热带常绿阔叶林物种多样性的维持。
DOIURL [本文引用: 1]
位于亚热带的浙江天童和古田山常绿阔叶林大样地分布有较高比例的落叶树种,那么它们与常绿树种的共存机制是什么?常绿树种和落叶树种生态习性差异较大,二者对生境的选择应有所不同,我们推测生境分化可能是两类植物实现共存的主要机制。为检验该假设,我们以天童20ha动态样地调查数据为依托,选择个体数≥20的55个常绿树种和42个落叶树种作为分析对象,用典范对应分析(CCA)研究了地形因子对二者分布的影响差异,用torus转换检验来分析常绿树种和落叶树种与各类地形生境的关联。结果如下:(1)CCA分析表明地形因子对常绿树种分布的解释量为19.2%,对落叶树种分布的解释量为7.0%。(2)torus转换检验结果表明,与沟谷成正关联的常绿树种和落叶树种的比例分别为16.4%和28.6%,成负关联的比例分别为40%和7%;与山脊成正关联的常绿树种和落叶树种的比例分别为41.8%和4.8%,成负关联的比例分别为10.9%和47.6%;与受干扰生境成正关联的常绿树种和落叶树种的比例分别为16.4%和42.9%。上述结果说明地形对常绿树种分布的影响大于落叶树种;两个植物类群对生境的选择多呈现相反格局,尤其是在沟谷生境和山脊生境,这进一步表明生境分化是常绿树种和落叶树种共存的重要机制之一,生态位理论在一定程度上能较好地解释亚热带常绿阔叶林物种多样性的维持。
DOIURL [本文引用: 1]
Topography of a Large-Scale Research Plot Established within a Tropical Rain Forest at Lambir, Sarawak YAMAKURA Takuo , KANZAKI Mamoru , ITOH Akira , OHKUBO Tatsuhiro , OGINO Kazuhiko , CHAI Ernest O. K. , LEE Hua Seng , ASHTON Peter Shaw Tropics 5(1), 41-56, 1995-12-20
[本文引用: 3]
[本文引用: 3]
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
Pflanzensoziologie: Grundzüge der Vegetationskunde
1
1964
... 群丛是植被分类的基本单位, 相当于植物分类中“种”的分类水平, 进行群丛分类是了解一个地区植被特征最基本、最重要的方法之一(
Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and Comparison with Other Plots
1
1998
... 用全站仪将整个样地划分成500个20 m × 20 m的样方, 将样方按1-500编号, 如原点(西南角第一个样方)处的20 m × 20 m样方编号为1, 1号样方右边为21号样方, 东北角最后一个样方编号为500.样地内所有胸径(DBH) ≥ 1 cm的木本植物均鉴定到种, 调查时测量其胸径并挂牌, 以便长期监测, 记录其编号、种名、胸径、坐标、生长状况以及生境概况, 详见巴拿马Barro Colorado岛(BCI)样地建设规范(
Indicspecies: Relationship Between Species and Groups of Sites
1
2016
... IndVal由
Associations between species and groups of sites: Indices and statistical inference
2
2009
... IndVal由
... ;
Improving indicator species analysis by combining groups of sites
2
2010
... IndVal由
... ;
Species assemblages and indicator species: The need for a flexible asymmetrical approach
2
1997
... IndVal由
... 两部分综合计算得到IndVal (
植物群落清查的主要内容、方法和技术规范
1
2009
... 根据已有的植物群落调查规范(
植物群落清查的主要内容、方法和技术规范
1
2009
... 根据已有的植物群落调查规范(
The National Vegetation Classification Standard
2008
Habitat associations of trees and shrubs in a 50-ha neotropical forest plot
1
2001
... 环境数据为每个20 m × 20 m样方的4个地形因子: 海拔、凹凸度、坡度、坡向.地形因子计算方法参照ForestGEO (Forest Global Earth Observatory)样地常用方法(
TWINSPAN-A FORTRAN Program for Arranging Multivariate Data in an Ordered Two-Way Table by Classification of the Individuals and Attributes
1
1979
... TWINSPAN是依据各样方内物种种类组成及各物种重要值进行分类, 最后得到同时包含物种分类和样方分类的一个矩阵(
弄岗北热带喀斯特季节性雨林群系数量分类及与环境的关系
1
2014
... 大型动态监测样地在进行群落调查时是按种记录DBH ≥ 1 cm的每一个体的胸径, 而不是像一般森林群落调查中分层(乔木层、灌木层及草本层)记录每一种多度、显著度/盖度, 在不分层情况下, 包含相对频度计算的重要值会放大小个体、分布广的种的重要性, 弱化大个体、有限分布种的重要性.
弄岗北热带喀斯特季节性雨林群系数量分类及与环境的关系
1
2014
... 大型动态监测样地在进行群落调查时是按种记录DBH ≥ 1 cm的每一个体的胸径, 而不是像一般森林群落调查中分层(乔木层、灌木层及草本层)记录每一种多度、显著度/盖度, 在不分层情况下, 包含相对频度计算的重要值会放大小个体、分布广的种的重要性, 弱化大个体、有限分布种的重要性.
Standards for associations and alliances of the U.S. National Vegetation Classification
4
2009
... 植被分类是对植物群落进行类型划分, 并根据各类型间一定相互关系建立相应分类体系的过程.作为基础科学研究的重要工具, 植被分类可以极大地促进人们对生态学格局的理解, 帮助研究者收集和解释生态背景信息(
... 植被分类随着植被生态学的发展而深入, 但到目前为止尚未有一种全球公认的标准分类方法和体系.这不仅由于植被本身的复杂性和区域性, 同时也由于植物群落连续性与间断性的争论, 使得植被分类成为植被生态学研究中最复杂的一个问题(
... 植被在时间和空间上是随环境不断变化的, 并不存在清楚明确的界限, 亦即群落是连续的, 但在植被的连续之中也有间断(
... 群丛是植被分类的基本单位, 由于各个学派的分类途径不同, 植物群丛的定义各有侧重, 如法瑞学派强调特征种, 苏联学派强调各层优势种, 美国国家植被分类系统强调鉴别种等, 但总的来看, 植物群丛应该包括4个主要特点: (1)一致的群落外貌和结构, (2)一致的生境, (3)确定的植物种类组成, (4)在一个景观或区域内重复出现(
基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例
1
2010
... 大型动态监测样地在进行群落调查时是按种记录DBH ≥ 1 cm的每一个体的胸径, 而不是像一般森林群落调查中分层(乔木层、灌木层及草本层)记录每一种多度、显著度/盖度, 在不分层情况下, 包含相对频度计算的重要值会放大小个体、分布广的种的重要性, 弱化大个体、有限分布种的重要性.
基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例
1
2010
... 大型动态监测样地在进行群落调查时是按种记录DBH ≥ 1 cm的每一个体的胸径, 而不是像一般森林群落调查中分层(乔木层、灌木层及草本层)记录每一种多度、显著度/盖度, 在不分层情况下, 包含相对频度计算的重要值会放大小个体、分布广的种的重要性, 弱化大个体、有限分布种的重要性.
Multivariate Analysis of Ecological Data Using CANOCO
1
2003
... 群落实体单位确定后, 就需要分析群落样方数据, 以明确所划定群落类型内部是否均一, 以及与其他类型是否差异显著(
亚热带常绿阔叶林林窗物种丰富度的影响因素
1
2015
... 根据已有的植物群落调查规范(
亚热带常绿阔叶林林窗物种丰富度的影响因素
1
2015
... 根据已有的植物群落调查规范(
Cluster: Cluster Analysis Basics and Extensions
1
2017
... ai是样方i与同一个类型中其他样方的平均距离(相异度); bi是样方i与最相似类型中所有样方的平均距离(相异度).这里首先使用R语言vegan包(
Aims and Methods of Vegetation Ecology
3
1974
... 植被在时间和空间上是随环境不断变化的, 并不存在清楚明确的界限, 亦即群落是连续的, 但在植被的连续之中也有间断(
... 也曾提议, 根据Jaccard有-无指数(与最典型样地的相似性)在25%到50%之间的群落地段可以成为同一群丛的一部分(
... 大型动态监测样地在进行群落调查时是按种记录DBH ≥ 1 cm的每一个体的胸径, 而不是像一般森林群落调查中分层(乔木层、灌木层及草本层)记录每一种多度、显著度/盖度, 在不分层情况下, 包含相对频度计算的重要值会放大小个体、分布广的种的重要性, 弱化大个体、有限分布种的重要性.
The neutral theory of biodiversity: Do the numbers add up?
1
2005
... 环境数据为每个20 m × 20 m样方的4个地形因子: 海拔、凹凸度、坡度、坡向.地形因子计算方法参照ForestGEO (Forest Global Earth Observatory)样地常用方法(
Vegan: Ordination, Diversity and Dissimilarities
2
2017
... DCA是基于植物种类组成数据, 在对应分析(CA)基础上修改而成的特征向量排序, 它克服了弓形效应, 提高了排序精度(
... ai是样方i与同一个类型中其他样方的平均距离(相异度); bi是样方i与最相似类型中所有样方的平均距离(相异度).这里首先使用R语言vegan包(
Silhouettes: A graphical aid to the interpretation and validation of cluster analysis
1
1987
... 群落实体单位确定后, 就需要分析群落样方数据, 以明确所划定群落类型内部是否均一, 以及与其他类型是否差异显著(
对中国植被分类系统的认知和建议
1
2011
... 群丛是植被分类的基本单位, 相当于植物分类中“种”的分类水平, 进行群丛分类是了解一个地区植被特征最基本、最重要的方法之一(
对中国植被分类系统的认知和建议
1
2011
... 群丛是植被分类的基本单位, 相当于植物分类中“种”的分类水平, 进行群丛分类是了解一个地区植被特征最基本、最重要的方法之一(
2017
2017
中国常绿阔叶林——分类
1
2013
... 群丛命名需包括优势种和标志种两部分, 采用重要值较高的两个乔木树种作为群丛的优势种, 标志种则通过IndVal大小同时结合基于TWINSPAN分类结果的样方区分表(附录I)来确定.参考
中国常绿阔叶林——分类
1
2013
... 群丛命名需包括优势种和标志种两部分, 采用重要值较高的两个乔木树种作为群丛的优势种, 标志种则通过IndVal大小同时结合基于TWINSPAN分类结果的样方区分表(附录I)来确定.参考
浙江天童国家森林公园的植被与区系
2
1995
... 研究区域位于浙江省宁波市鄞州区东南部的天童国家森林公园内, 地处29.80° N, 121.78° E.该地森林植被保存良好, 是浙江省东部丘陵地区地带性植被类型的代表性地段.该区气候为中亚热带季风气候, 全年温和多雨, 四季分明.年平均气温为16.2 ℃, 最热月为7月(月平均气温28.1 ℃), 最冷月为1月(月平均气温4.2 ℃).区域内的年降水量为1374.7 mm, 主要集中于每年的5-8月(
... TWINSPAN分类是迄今为止最受欢迎的一种数量分类方法(
浙江天童国家森林公园的植被与区系
2
1995
... 研究区域位于浙江省宁波市鄞州区东南部的天童国家森林公园内, 地处29.80° N, 121.78° E.该地森林植被保存良好, 是浙江省东部丘陵地区地带性植被类型的代表性地段.该区气候为中亚热带季风气候, 全年温和多雨, 四季分明.年平均气温为16.2 ℃, 最热月为7月(月平均气温28.1 ℃), 最冷月为1月(月平均气温4.2 ℃).区域内的年降水量为1374.7 mm, 主要集中于每年的5-8月(
... TWINSPAN分类是迄今为止最受欢迎的一种数量分类方法(
中国常绿阔叶林8大动态监测样地植被的综合比较
1
2015
... 大型动态监测样地在进行群落调查时是按种记录DBH ≥ 1 cm的每一个体的胸径, 而不是像一般森林群落调查中分层(乔木层、灌木层及草本层)记录每一种多度、显著度/盖度, 在不分层情况下, 包含相对频度计算的重要值会放大小个体、分布广的种的重要性, 弱化大个体、有限分布种的重要性.
中国常绿阔叶林8大动态监测样地植被的综合比较
1
2015
... 大型动态监测样地在进行群落调查时是按种记录DBH ≥ 1 cm的每一个体的胸径, 而不是像一般森林群落调查中分层(乔木层、灌木层及草本层)记录每一种多度、显著度/盖度, 在不分层情况下, 包含相对频度计算的重要值会放大小个体、分布广的种的重要性, 弱化大个体、有限分布种的重要性.
再议中国的植被分类系统
7
2017
... 植被分类随着植被生态学的发展而深入, 但到目前为止尚未有一种全球公认的标准分类方法和体系.这不仅由于植被本身的复杂性和区域性, 同时也由于植物群落连续性与间断性的争论, 使得植被分类成为植被生态学研究中最复杂的一个问题(
... ;
... 群丛是植被分类的基本单位, 相当于植物分类中“种”的分类水平, 进行群丛分类是了解一个地区植被特征最基本、最重要的方法之一(
... 群丛分类的基础是野外样地调查, 它为植被分类提供了必要的原始材料.在通常的植被调查中, 为保证取样的代表性, 选择适当的取样地点是植被调查的关键.如果取样不当, 不仅不利于准确认识植被的总体, 反而可能会歪曲总体, 从而得出错误的结论(
... 根据已有的植物群落调查规范(
... 植被在时间和空间上是随环境不断变化的, 并不存在清楚明确的界限, 亦即群落是连续的, 但在植被的连续之中也有间断(
... 群丛是植被分类的基本单位, 由于各个学派的分类途径不同, 植物群丛的定义各有侧重, 如法瑞学派强调特征种, 苏联学派强调各层优势种, 美国国家植被分类系统强调鉴别种等, 但总的来看, 植物群丛应该包括4个主要特点: (1)一致的群落外貌和结构, (2)一致的生境, (3)确定的植物种类组成, (4)在一个景观或区域内重复出现(
再议中国的植被分类系统
7
2017
... 植被分类随着植被生态学的发展而深入, 但到目前为止尚未有一种全球公认的标准分类方法和体系.这不仅由于植被本身的复杂性和区域性, 同时也由于植物群落连续性与间断性的争论, 使得植被分类成为植被生态学研究中最复杂的一个问题(
... ;
... 群丛是植被分类的基本单位, 相当于植物分类中“种”的分类水平, 进行群丛分类是了解一个地区植被特征最基本、最重要的方法之一(
... 群丛分类的基础是野外样地调查, 它为植被分类提供了必要的原始材料.在通常的植被调查中, 为保证取样的代表性, 选择适当的取样地点是植被调查的关键.如果取样不当, 不仅不利于准确认识植被的总体, 反而可能会歪曲总体, 从而得出错误的结论(
... 根据已有的植物群落调查规范(
... 植被在时间和空间上是随环境不断变化的, 并不存在清楚明确的界限, 亦即群落是连续的, 但在植被的连续之中也有间断(
... 群丛是植被分类的基本单位, 由于各个学派的分类途径不同, 植物群丛的定义各有侧重, 如法瑞学派强调特征种, 苏联学派强调各层优势种, 美国国家植被分类系统强调鉴别种等, 但总的来看, 植物群丛应该包括4个主要特点: (1)一致的群落外貌和结构, (2)一致的生境, (3)确定的植物种类组成, (4)在一个景观或区域内重复出现(
Fushan Subtropical Forest Dynamics Plot: Tree Species Characteristics and Distribution Patterns
1
2007
... 大型动态监测样地在进行群落调查时是按种记录DBH ≥ 1 cm的每一个体的胸径, 而不是像一般森林群落调查中分层(乔木层、灌木层及草本层)记录每一种多度、显著度/盖度, 在不分层情况下, 包含相对频度计算的重要值会放大小个体、分布广的种的重要性, 弱化大个体、有限分布种的重要性.
Tree species distributions and local habitat variation in the Amazon: Large forest plot in eastern Ecuador
1
2004
... 环境数据为每个20 m × 20 m样方的4个地形因子: 海拔、凹凸度、坡度、坡向.地形因子计算方法参照ForestGEO (Forest Global Earth Observatory)样地常用方法(
Vegetation Ecology. 2nd edn. John Wiley & Sons,
6
2013
... 植被分类是对植物群落进行类型划分, 并根据各类型间一定相互关系建立相应分类体系的过程.作为基础科学研究的重要工具, 植被分类可以极大地促进人们对生态学格局的理解, 帮助研究者收集和解释生态背景信息(
... 植被分类随着植被生态学的发展而深入, 但到目前为止尚未有一种全球公认的标准分类方法和体系.这不仅由于植被本身的复杂性和区域性, 同时也由于植物群落连续性与间断性的争论, 使得植被分类成为植被生态学研究中最复杂的一个问题(
... ;
... IndVal由
... 植被在时间和空间上是随环境不断变化的, 并不存在清楚明确的界限, 亦即群落是连续的, 但在植被的连续之中也有间断(
... TWINSPAN分类是迄今为止最受欢迎的一种数量分类方法(
The Braun-Blanquet approach. In: Whittaker RH ed. Classification of Plant Communities
1
1985
... 群丛是植被分类的基本单位, 相当于植物分类中“种”的分类水平, 进行群丛分类是了解一个地区植被特征最基本、最重要的方法之一(
The Braun-Blanquet approach. In: Whittaker RH ed. Classification of Plant Communities
1
1985
... 群丛是植被分类的基本单位, 相当于植物分类中“种”的分类水平, 进行群丛分类是了解一个地区植被特征最基本、最重要的方法之一(
Classification of Plant Communities
1
1985
... 群丛是植被分类的基本单位, 相当于植物分类中“种”的分类水平, 进行群丛分类是了解一个地区植被特征最基本、最重要的方法之一(
Classification of Plant Communities
1
1985
... 群丛是植被分类的基本单位, 相当于植物分类中“种”的分类水平, 进行群丛分类是了解一个地区植被特征最基本、最重要的方法之一(
Vegetation of China
3
1980
... 群丛是植被分类的基本单位, 相当于植物分类中“种”的分类水平, 进行群丛分类是了解一个地区植被特征最基本、最重要的方法之一(
... ).这种以植物区系特征作为植被分类基本单位的划分标准, 强调特征种、鉴别种(diagnostic species)或标志种(indicative species)的作用, 已被我国(
... )及美国(Federal Geographic Data Committee, Vegetation Subcommittee (FGDV-VS), 2008)植被分类系统中的群丛定义所采用.因为基于研究对象本身特征的分类比其他任何分类更加自然, 这样也同时考虑了植物本身的生态特性以及它们对环境的指示意义(
Vegetation of China
3
1980
... 群丛是植被分类的基本单位, 相当于植物分类中“种”的分类水平, 进行群丛分类是了解一个地区植被特征最基本、最重要的方法之一(
... ).这种以植物区系特征作为植被分类基本单位的划分标准, 强调特征种、鉴别种(diagnostic species)或标志种(indicative species)的作用, 已被我国(
... )及美国(Federal Geographic Data Committee, Vegetation Subcommittee (FGDV-VS), 2008)植被分类系统中的群丛定义所采用.因为基于研究对象本身特征的分类比其他任何分类更加自然, 这样也同时考虑了植物本身的生态特性以及它们对环境的指示意义(
基于地形因子的天童地区常绿树种和落叶树种共存机制研究
1
2012
... 根据已有的植物群落调查规范(
基于地形因子的天童地区常绿树种和落叶树种共存机制研究
1
2012
... 根据已有的植物群落调查规范(
Topography of a large-scale research plot established within a tropical rain forest at Lambir, Sarawak
1
1995
... 环境数据为每个20 m × 20 m样方的4个地形因子: 海拔、凹凸度、坡度、坡向.地形因子计算方法参照ForestGEO (Forest Global Earth Observatory)样地常用方法(
浙江天童20 ha常绿阔叶林动态监测样地的群落特征
3
2011
... 常绿阔叶林是亚热带地区的地带性植被, 种类组成复杂, 类型多样, 在人类长期干预下, 次生性强, 更增加了分类的困难.浙江天童20 hm2动态监测样地内的植物群落具有典型的中国东部中亚热带低海拔森林特征, 群落发育成熟, 结构完整, 对其物种成分已有报道(
... 天童20 hm2样地位于天童国家森林公园的核心保护区(29.81°-29.82° N, 121.78°-121.79° E), 样地位置及三维地形图如
... 用全站仪将整个样地划分成500个20 m × 20 m的样方, 将样方按1-500编号, 如原点(西南角第一个样方)处的20 m × 20 m样方编号为1, 1号样方右边为21号样方, 东北角最后一个样方编号为500.样地内所有胸径(DBH) ≥ 1 cm的木本植物均鉴定到种, 调查时测量其胸径并挂牌, 以便长期监测, 记录其编号、种名、胸径、坐标、生长状况以及生境概况, 详见巴拿马Barro Colorado岛(BCI)样地建设规范(
浙江天童20 ha常绿阔叶林动态监测样地的群落特征
3
2011
... 常绿阔叶林是亚热带地区的地带性植被, 种类组成复杂, 类型多样, 在人类长期干预下, 次生性强, 更增加了分类的困难.浙江天童20 hm2动态监测样地内的植物群落具有典型的中国东部中亚热带低海拔森林特征, 群落发育成熟, 结构完整, 对其物种成分已有报道(
... 天童20 hm2样地位于天童国家森林公园的核心保护区(29.81°-29.82° N, 121.78°-121.79° E), 样地位置及三维地形图如
... 用全站仪将整个样地划分成500个20 m × 20 m的样方, 将样方按1-500编号, 如原点(西南角第一个样方)处的20 m × 20 m样方编号为1, 1号样方右边为21号样方, 东北角最后一个样方编号为500.样地内所有胸径(DBH) ≥ 1 cm的木本植物均鉴定到种, 调查时测量其胸径并挂牌, 以便长期监测, 记录其编号、种名、胸径、坐标、生长状况以及生境概况, 详见巴拿马Barro Colorado岛(BCI)样地建设规范(
数量生态学. 第二版
1
2011
... DCA是基于植物种类组成数据, 在对应分析(CA)基础上修改而成的特征向量排序, 它克服了弓形效应, 提高了排序精度(
数量生态学. 第二版
1
2011
... DCA是基于植物种类组成数据, 在对应分析(CA)基础上修改而成的特征向量排序, 它克服了弓形效应, 提高了排序精度(
天童常绿阔叶林林窗的地形分布格局
1
2013
... 根据已有的植物群落调查规范(
天童常绿阔叶林林窗的地形分布格局
1
2013
... 根据已有的植物群落调查规范(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}