 ,*, 刘雨桐新疆大学资源与环境科学学院, 绿洲生态教育部重点实验室, 乌鲁木齐 830046
,*, 刘雨桐新疆大学资源与环境科学学院, 绿洲生态教育部重点实验室, 乌鲁木齐 830046Effects of seasonal snow cover on decomposition and carbon, nitrogen and phosphorus release of Picea schrenkiana leaf litter in Mt. Tianshan, Northwest China
CHEN Wen-Jing, GONG Lu,*, LIU Yu-TongCollege of Resources and Environment Science, Xinjiang University, Key Laboratory of Oasis Ecology, Ministry of Education, ürümqi 830046, China通讯作者: (gonglu721@163.com)
编委: 李胜功
责任编辑: 王葳
| 基金资助: |
Online:2018-04-20
| Fund supported: |

摘要
季节性雪被下显著的冻融格局差异可能对干旱区山地森林凋落叶分解过程产生重要影响, 但一直未见深入研究。2015年10月至2016年10月, 采用凋落物分解袋法, 研究了天山典型树种雪岭云杉(Picea schrenkiana)凋落叶在季节性雪被覆盖下的3个关键时期(冻融期、深冻期、融冻期)以及生长季(生长季早期和生长季末期)的分解动态和碳、氮、磷释放特征。结果表明: (1)经过一年的分解, 不同雪被厚度下雪岭云杉凋落叶分解率为24.6%-29.2%, 且存在显著性差异。分解系数k值厚雪被覆盖最大, 无雪被覆盖最小。(2)冬季雪被覆盖期雪岭云杉凋落叶分解对当年分解总量的贡献达46.0%- 48.5%, 其中对冻融期凋落叶分解影响较为明显。(3)随着凋落叶的分解, 雪岭云杉凋落叶氮含量总体呈增加趋势; 碳含量和碳氮比大致呈下降趋势, 在深冻期和生长季末期不同雪被处理下碳含量呈显著性差异; 而凋落叶磷含量呈不规则变化趋势, 且在冻融期和融冻期不同雪被厚度下呈显著性差异。(4)整个雪被覆盖季节凋落叶氮元素表现为富集, 碳和磷元素表现为释放; 其中, 在融冻期薄雪被和中雪被处理下碳元素富集率最大, 在冻融期薄雪被、中雪被和厚雪被处理下, 融冻期无雪被和厚雪被下以及生长季早期中雪被和厚雪被下氮元素富集率最大, 而雪被对凋落叶磷释放的影响不显著。
关键词:
Abstract
Keywords:
PDF (1212KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
陈文静, 贡璐, 刘雨桐. 季节性雪被对天山雪岭云杉凋落叶分解和碳氮磷释放的影响. 植物生态学报[J], 2018, 42(4): 487-497 DOI:10.17521/cjpe.2017.0298
CHEN Wen-Jing, GONG Lu, LIU Yu-Tong.
凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(贾丙瑞等, 2016; Paudel et al., 2016)。迄今为止, 国内外专家已经对凋落物分解进行了大量研究(Parton et al., 2007; Barrett, 2008; Zhang & Wang, 2015; 刘涛等, 2017), 研究对象包括不同气候带和植被类型, 所涉及的环境因素有光照、大气氮沉降、土壤温湿度、土壤类型以及雪被斑块等(Parton et al., 2007; Berg & McClaugherty, 2008; Baptist et al., 2010)。已有研究表明, 气候主要控制区域尺度的凋落物分解速率, 而小尺度的凋落物分解过程则由不同物种凋落物基质质量所控制(Aerts, 2006)。传统观点认为, 凋落叶分解速率随温度和湿度的升高而加快, 冬季冻结环境下凋落叶分解缓慢甚至停滞(Vitousek et al., 1994)。然而越来越多的冬季凋落物分解实验表明, 高寒山地森林生态系统中雪被下的绝热保温环境以及由气温变化等因素产生的冻融循环作用成为影响凋落物分解过程的重要因素(Bleak, 1970; Taylor & Parkinson, 1988; Wu et al., 2010; 胡霞等, 2012b; Saccone et al., 2013; 武启骞等, 2013), 因此, 冬季雪被斑块下凋落物质量损失在年际分解率中可能占较大比例。当前关于季节性雪被下凋落物分解研究主要集中在青藏高原中高海拔山地森林生态系统(胡霞等, 2012b; 吴启骞等, 2013; 刘涛等, 2017), 而对干旱区山地温带针叶林凋落物分解及其碳氮磷释放影响的研究鲜有报道。
天山森林在涵养水源、保育土壤、固碳释氧以及保护生物多样性、维持绿洲稳定性等方面发挥着不可替代的作用(宋新章等, 2008; 许文强等, 2016)。在新疆天山地区, 季节性积雪是最活跃且敏感的环境要素, 雪被覆盖期长达5-6个月, 并在林窗-林下形成天然的雪被梯度(Tan et al., 2010)。而该区域雪被期雪岭云杉(Picea schrenkiana)凋落叶分解特征如何? 碳氮磷释放呈现何种规律? 仍未见报道。本文以天山北麓中山带优势物种雪岭云杉凋落叶为研究对象, 采用凋落物分解袋法, 探讨季节性雪被对凋落叶分解和碳氮磷释放特征的影响, 以期为深入了解中高海拔山地森林物质循环等过程对气候变暖的响应提供理论依据。
1 材料和方法
1.1 研究区概况
研究区位于新疆天山北麓中山带, 横跨83°- 94° E, 纵贯42°-45° N, 距乌鲁木齐市61 km的水西沟。其海拔1 300-4 200 m, 属于温带大陆性气候, 年总辐射量达5.85 × 105 J·cm-2, 年平均气温为2-3 ℃, 历年最高气温为30.5 ℃, 最低气温为-30.2 ℃, 年降水量400-600 mm, 雨季集中在6-8月份, 年蒸发量150-980 mm, 年平均相对湿度65%, 干燥度1.4, 无霜期89天, 积温1 170.5 ℃, 最大积雪深度为65 cm。塔里木河和伊犁河是天山南北坡最重要的两条水系。林下土壤为山地灰褐色森林土, 腐殖质层较厚。当地主要森林类型为北方温带针叶林, 其中雪岭云杉是研究区域数量最多、分布最广的物种, 主要见于中高海拔地区的背阴面。1.2 实验设计和方法
于实验区内选取具有代表性的雪岭云杉纯林作为实验样地, 并在样地内分别选取环境(坡向、坡度相似, 树龄、海拔相同等)相对一致的大约400 m2的林窗, 每个林窗间隔大于1 km, 同时沿顺风方向从林窗中央至林下依次设置4个4 m × 4 m的小样方, 每个样方间距为5 m左右, 以保证在冬季可以形成从厚到无4个雪被梯度, 分别记作厚雪被、中雪被、薄雪被、无雪被, 雪被厚度变化见图1。于2015年9月下旬在雪岭云杉林中采集地表新鲜凋落叶带回实验室, 彻底风干后称取5份质量为10 g的凋落物, 于65 ℃烘箱烘干至恒质量, 用以计算凋落物初始质量。称取相当于烘干质量为10 g的凋落物装入大小为400 cm2、孔径为0.1 mm的尼龙网袋中备用, 共计300袋(3个样地×4个样方×1个物种×5次采样×5个重复)。在2015年10下旬, 将凋落叶袋平铺于提前划分标记好的样方上, 袋间距为5 cm左右, 以免其相互影响。同时, 在每种雪被处理下设置纽扣式温度记录器(iButton DS1923-F5, Maxim/Dallas Semiconductor, Sunnyvale, USA), 将其放置于相应的凋落叶分解袋内, 每隔2 h记录一次土壤温度数据(图2)。图1
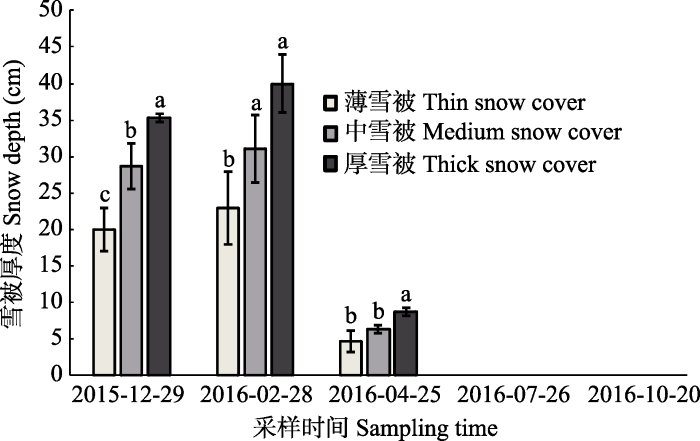 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同采样时间雪被厚度变化动态(平均值±标准偏差, n = 5)。小写字母表示相同采样日期不同雪被厚度间的差异性(p < 0.05)。
Fig. 1Thickness changes of snow cover in different sampling time (mean ± SD, n = 5). The lowercase letters indicate significant difference among different snow cover thickness for the same sampling date (p < 0.05).
图2
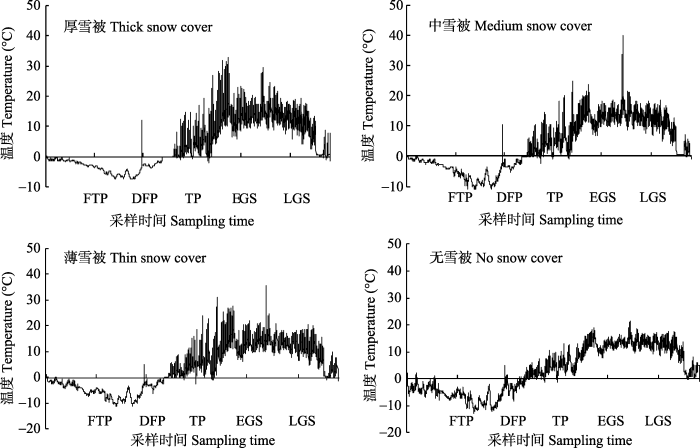 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2研究样地不同雪被厚度下土壤温度动态。DFP, 深冻期; EGS, 生长季早期; FTP, 冻融期; LGS, 生长季末期; TP, 融冻期。
Fig. 2Dynamics of soil temperature under different depths of snow cover in the sampling forest. DFP, deep-freeze period; EGS, early growing season; FTP, freeze-thaw period; LGS, late growing season; TP, thawing period.
为探究雪被形成、覆盖、融化以及生长季凋落叶分解和碳氮磷释放特征, 在前期观测基础上, 将冬季分为3个阶段, 生长季分为2个阶段。即2015- 10-30-2015-12-29 (冻融期, FTP)、2015-12-29- 2016-02-28 (深冻期, DFP)、2016-02-28-2016-04-25(融冻期, TP)、2016-04-25-2016-07-26 (生长季早期, EGS)和2016-07-26-2016-10-20 (生长季末期, LGS)。每次采样随机从每个样方内采集雪岭云杉凋落物分解袋各5袋, 采样时测量林窗至林下不同位置的雪被厚度, 由于样地处于中高海拔森林, 无法进行适时监测, 雪被厚度用钢卷尺多点测量, 并读取各样方下的土壤温度数据。
1.3 样品分析
将已采集到实验室的凋落叶分解袋打开, 小心去除泥土杂物及新生根系, 于65 ℃烘干至恒质量, 称量并计算凋落叶质量损失率。氮含量采用半微量凯氏定氮法测定, 磷含量采用钼锑抗比色法测定, 碳含量采用硫酸-重铬酸钾外加热法测定。1.4 计算公式及数据分析
质量损失贡献率公式为:$G(%)=\frac{{{M}_{i-1}}-{{M}_{i}}}{{{M}_{0}}-{{M}_{I}}}\times 100%$
其中G为质量损失率(%), M0为凋落叶初始质量(g), Mi为第i次回收凋落叶袋时凋落叶烘干质量(g), MI为最后一次回收凋落叶袋时凋落叶烘干质量(g) (Zhu et al., 2012)。
凋落物质量损失率(M)公式为:
$M(%)=\frac{{{M}_{0}}-{{M}_{i}}}{{{M}_{0}}}\times 100%$
养分释放率公式(E)为:
$E(%)=\frac{{{E}_{i-1}}-{{E}_{i}}}{{{E}_{0}}}\times 100%$
式中E0为凋落叶初始元素含量(g·kg-1), Ei为第i次回收凋落叶袋时凋落叶元素含量(g·kg-1), 元素表现为净释放时E为正值, 元素为净富集时E为负值。
凋落叶分解Olson衰减指数模型为:
y=ae-kt...
其中y为t时间回收凋落叶袋的残留率(%), k为分解系数, t为回收凋落叶袋时间(a) (Olson, 1963)。
分解半衰期(50%分解): t0.05 = ln0.5/(-k)
完全分解时间(95%分解): t0.95 = ln0.5/(-k) (宋新章等, 2009)
数据统计与分析使用SPSS 19.0和Excel完成, 采用单因素方差分析和最小显著差异法(LSD)比较不同雪被下凋落叶分解率以及雪被斑块对凋落叶元素释放的影响。
2 结果分析
2.1 不同雪被下雪岭云杉凋落叶的分解率
经过一年的分解, 不同雪被覆盖下雪岭云杉凋落叶分解率达24.6%-29.2%, 且存在显著性差异 (p < 0.05), 各时期雪岭云杉凋落叶分解率随雪被厚度的减小而降低, 即厚雪被中质量损失最大, 无雪被中质量损失最小(图3)。同一时期, 厚雪被下凋落叶分解率均显著高于无雪被和中雪被(p < 0.05); 除生长季后期外, 薄雪被和中雪被下凋落叶分解率在其余4个时期中无明显差异(p > 0.05); 厚雪被与中雪被下凋落叶分解率在冻融期、生长季前期和生长季后期中呈显著性差异(p < 0.05); 在生长季后期, 厚雪被下凋落叶分解率远大于无雪被下凋落叶分解率; 在深冻期和融冻期, 无雪被、薄雪被和中雪被下凋落叶分解率均无显著差异, 而在厚雪被下存在显著性差异(p < 0.05)。图3
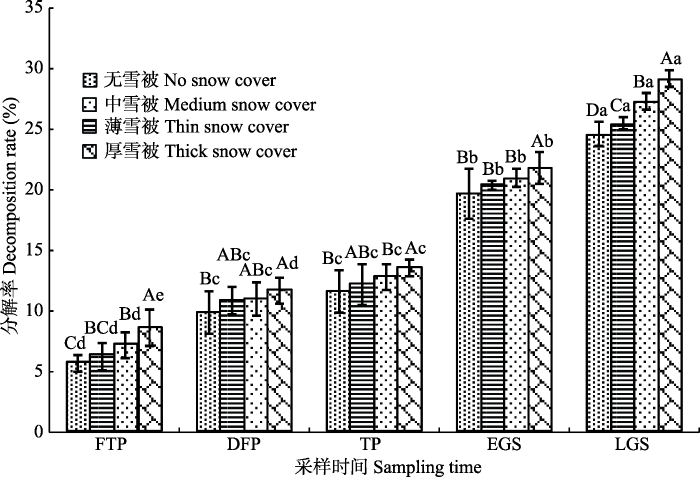 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同雪被厚度下雪岭云杉凋落叶质量损失率(平均值±标准偏差, n = 5)。DFP, 深冻期; EGS, 生长季早期; FTP, 冻融期; LGS, 生长季末期; TP, 融冻期。大写字母表示相同时期不同雪被厚度差异性; 小写字母表示相同雪被厚度不同时期差异性(p < 0.05)。
Fig. 3Mass loss rates of Picea schrenkiana leaf litter under different depths of snow cover (mean ± SD, n = 5). DFP, deep-freeze period; EGS, early growing season; FTP, freeze-thaw period; LGS, late growing season; TP, thawing period. The capital letters indicate the difference of same period in different snow thickness. The lowercase letters indicate the difference of same snow thickness in different periods (p < 0.05).
利用Olson指数衰减模型对雪岭云杉凋落叶分解过程的残留率(1-质量损失率%)进行拟合(Olson, 1963), 并估算完成50%和95%分解所需的时间。天山雪岭云杉的k值以厚雪被覆盖为最大(0.310), 完成50%和95%分解所需时间较短, 而无雪被下的k值最小(0.273), 完成50%和95%分解所需时间较长(表1)。
Table 1
表1
表1雪岭云杉凋落叶非线性回归拟合Olson指数模型
Table 1
| 雪被 Snow cover | 回归方程 Regression model | 分解系数 k Decomposition constant k | 相关系数 r Correlation coefficient r | 半分解时间 Time of half decomposition (a) | 95%分解时间 Time of 95% decomposition (a) |
|---|---|---|---|---|---|
| 厚雪被 Thick snow cover | y = 98.009e-0.310x | 0.310 | 0.97 | 2.336 | 9.664 |
| 中雪被 Medium snow cover | y = 98.592e-0.296x | 0.296 | 0.98 | 2.342 | 10.121 |
| 薄雪被 Thin snow cover | y = 98.556e-0.277x | 0.277 | 0.98 | 2.502 | 10.815 |
| 无雪被 No snow cover | y = 99.251e-0.273x | 0.273 | 0.98 | 2.539 | 10.973 |
新窗口打开|下载CSV
2.2 不同雪被下雪岭云杉凋落叶在各时期的质量损失贡献率
雪被覆盖期内凋落叶质量损失贡献率略小于生长季, 实验表明, 雪岭云杉凋落叶质量损失在生长季前期消耗最多(图4)。生长季前期凋落叶质量损失贡献率随雪被厚度增加而降低, 其中无雪被下质量损失贡献率最高, 达到了32.1%; 而在生长季后期, 凋落叶质量损失贡献率随雪被厚度增加而增加; 雪被覆盖期凋落叶质量损失主要发生在冻融期和深冻期, 其中融冻期内凋落叶质量损失明显低于其余两个时期; 冻融期内雪岭云杉凋落叶分解贡献率基本随雪被厚度增加而增加, 且厚雪被下凋落叶贡献率要比无雪被下高4%。图4
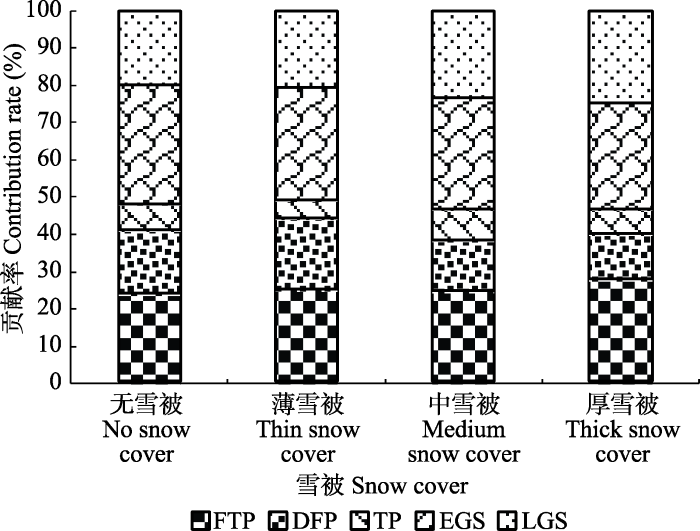 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同雪被厚度下不同时期对雪岭云杉凋落叶质量损失的贡献率。DFP, 深冻期; EGS, 生长季早期; FTP, 冻融期; LGS, 生长季末期; TP, 融冻期。
Fig. 4Proportional contribution (%) of the decomposition in different periods to the total decomposition of Picea schrenkiana leaf litter under different depths of snow cover. DFP, deep- freeze period; EGS, early growing season; FTP, freeze-thaw period; LGS, late growing season; TP, thawing period.
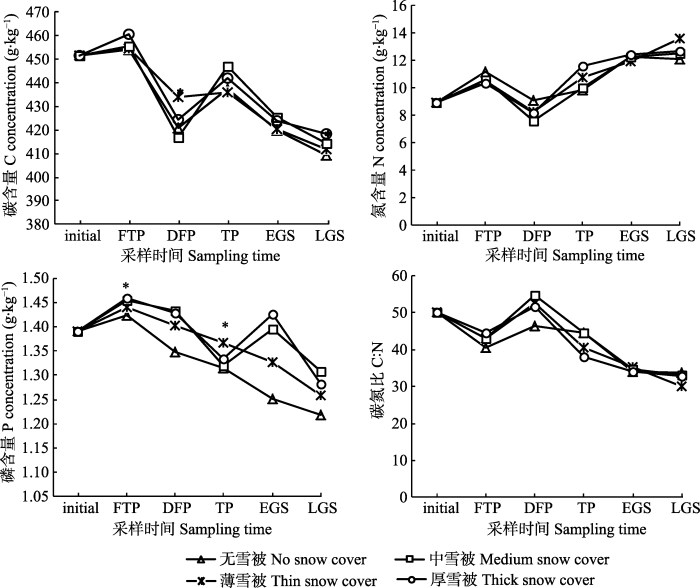
2.3 不同雪被下雪岭云杉凋落叶养分含量的变化
随着凋落叶分解的进行, 雪岭云杉凋落叶氮含量大体呈增加趋势(图5), 分解实验结束后, 不同雪被厚度下氮含量为薄雪被>厚雪被>中雪被>无雪被, 且差异不显著(p > 0.05)。碳含量和C:N在不同雪被厚度下大致呈下降趋势(图5); 凋落叶碳含量在生长季后期大致表现为无雪被下最低, 厚雪被下最高, 在深冻期和生长季后期不同雪被处理下存在显著性差异(p < 0.05); 而在生长季后期, 碳氮比在薄雪被下明显降低, 其他雪被厚度下基本一致。不同雪被梯度下凋落叶磷含量呈不规则变化(图5), 其中冻融期内4种雪被厚度下磷含量均高于初始值, 之后无雪被和薄雪被下磷含量随着时间的变化表现为逐渐降低趋势, 且无雪被下磷含量始终低于有雪被下磷含量, 在冻融期和融冻期不同雪被厚度下具有显著差异(p < 0.05)。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5不同雪被厚度下雪岭云杉凋落叶氮、磷、碳含量及碳氮比的动态变化。DFP, 深冻期; EGS, 生长季早期; FTP, 冻融期; LGS, 生长季末期; TP, 融冻期。initial, 初始期。*, 同一时期不同厚度雪被斑块间差异显著(p < 0.05)。
Fig. 5Dynamics of C, N, P contents and C:N in Picea schrenkiana leaf litter under different depths of snow cover. DFP, deep-freeze period; EGS, early growing season; FTP, freeze-thaw period; LGS, late growing season; TP, thawing period. initial, initial period. *, significant difference among different snow cover thickness in the same period (p < 0.05).
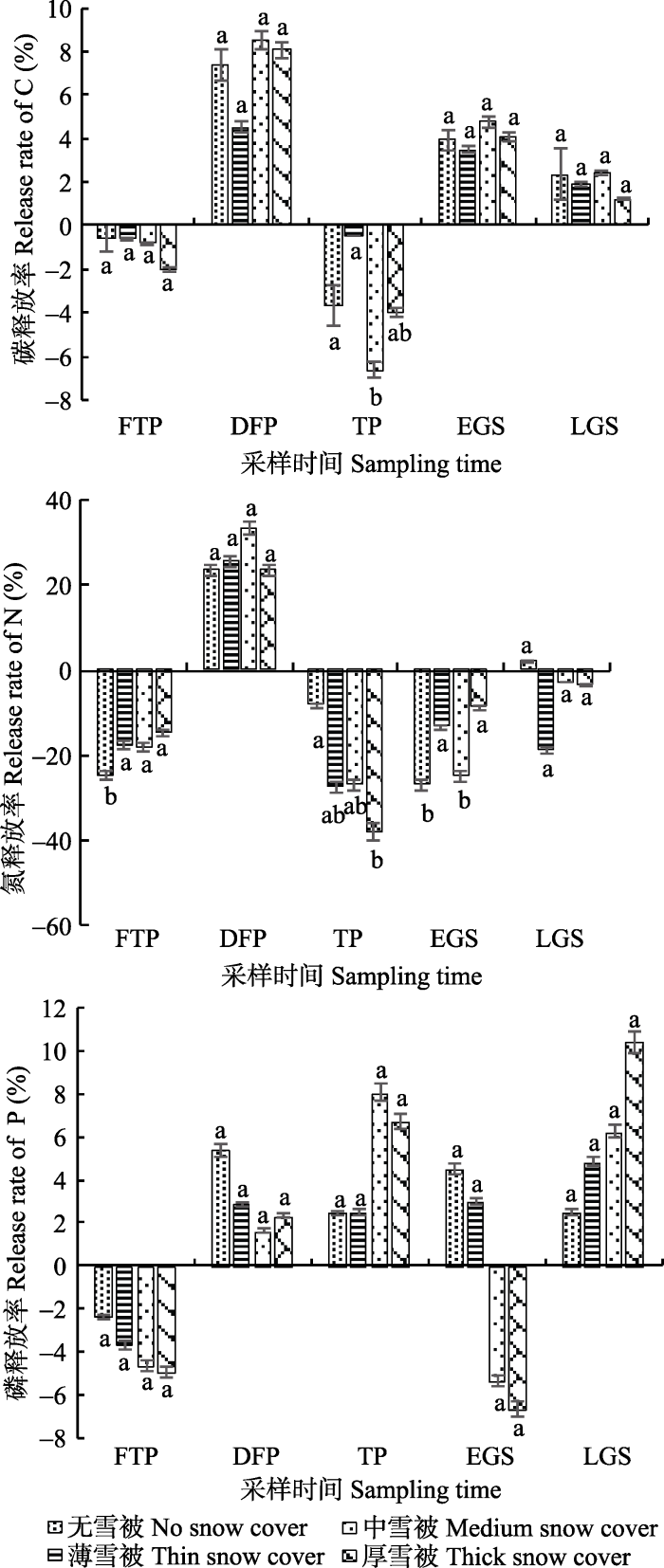
2.4 不同雪被厚度下雪岭云杉凋落叶养分释放动态
在雪岭云杉凋落叶分解过程中, 不同雪被厚度下碳元素经历了富集-释放-再富集-再释放的过程(图6), 且在融冻期, 凋落叶碳富集率以薄雪被和中雪被处理下最大。在冬季雪被覆盖期, 冻融期和融冻期中不同雪被梯度下凋落叶氮表现为富集状态, 深冻期则表现为释放状态, 到生长季时期又表现为富集状态(图6); 其中在冻融期的薄雪被、中雪被和厚雪被处理下氮元素富集率最大, 而在融冻期则是无雪被和厚雪被下氮元素富集率最大, 在生长季前期氮元素富集率以中雪被和厚雪被处理下最大。凋落叶磷在冻融期不同雪被梯度下表现为富集现象, 到深冻期和融冻期时, 不同雪被厚度下磷元素表现为释放状态; 生长季节前期, 无雪被和薄雪被梯度下磷元素表现为释放状态, 而中雪被和厚雪被梯度下磷元素则表现为富集状态, 进入生长季后期, 4种雪被梯度下磷元素均表现为释放状态, 至分解实验结束, 各雪被处理下凋落叶磷释放率无显著差异 (p > 0.05)。
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6不同时期不同雪被厚度下凋落叶分解过程中碳、氮、磷的释放率(平均值±标准偏差, n = 4)。DFP, 深冻期; EGS, 生长季早期; FTP, 冻融期; LGS, 生长季末期; TP, 融冻期。不同小写字母表示同一时期不同雪被厚度间差异显著(p < 0.05)。
Fig. 6Release rates of C, N, P in leaf litter during decomposition under snow cover with different depths in different period (mean ± SD, n = 4). DFP, deep-freeze period; EGS, early growing season; FTP, freeze-thaw period; LGS, late growing season; TP, thawing period. Different small letters meant significant difference among different snow thickness in the same period (p < 0.05).
3 讨论和结论
3.1 不同雪被下雪岭云杉凋落叶在各时期的分解速率及其质量损失贡献率
以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(Makoto et al., 2014)。一年的凋落叶分解实验表明, 季节性雪被不仅显著影响冻融期、深冻期和融冻期凋落叶分解, 而且持续影响生长季时期凋落叶分解。冬季分解时期, 雪岭云杉凋落叶分解率随雪被厚度增加而增加, 与前人的研究结论一致(Preston et al., 2009; Aerts et al., 2012; 何伟等, 2013), 这一结果表明凋落物分解对季节性雪被响应较大, 同时气候变暖情景下季节性雪被变化也会进一步影响凋落物分解等山地森林物质循环过程(吴启骞等, 2013)。Olson凋落物分解系数(k)常用来表示凋落物分解速率(Olson, 1963), 本研究中, k值对雪被厚度变化具有明显的响应。可能因为温度波动幅度随着冬季雪被厚度的减小而逐渐增大(图2), 厚雪被具有隔热保温作用, 所提供的稳定环境更益于雪岭云杉凋落叶分解(吴启骞等, 2013)。此外, 各雪被斑块下的土壤动物群落和水热条件有所不同(Colbeck, 1983; Clein & Schimel, 1995; Uchida et al., 2005), 使得起关键作用的微生物群落和数量亦有所差异; 一般分解者群落随雪被厚度的增加而愈加丰富, 生物种群活动及活性也更强(Tomaselli, 1991; Uchida et al., 2005; 胡霞等, 2012a; Zhu et al., 2012)。在生长季, 由于地形地貌和林窗格局等异质性因素导致阳光对样地照射的角度以及对林窗中心的辐射强度不同(He et al., 2016), 因此不同雪被厚度下土壤温度产生了较大波动, 且波动幅度从厚雪被到无雪被逐渐减小, 使得昼夜温差产生较大差异,而土壤动物和微生物活性易受温度的影响(曹宏杰和倪红伟, 2015), 能间接影响凋落物分解速率。武启骞等(2013)对川西高山森林季节性雪被下凋落叶分解动态的研究也得到了相同的结论。雪被下不同时期对凋落叶分解具有不同的作用机制, 因此不同凋落物分解阶段或许对雪被覆盖表现出不同的响应机制。本研究中, 在冬季3个关键时期, 厚雪被处理下凋落叶分解贡献率总是大于无雪被处理下凋落叶分解贡献率, 冬季雪被覆盖显著提高了凋落叶的分解贡献率。可能由于地表雪被为土壤动物以及微生物活性提供了隔热保温且相对稳定的环境(Clein & Schimel, 1995; 夏磊等, 2011), 同时雪被在冻融期和融冻期有更强的淋洗作用。进入生长季前期, 无雪被厚度下凋落叶分解贡献率明显高于有雪被覆盖下凋落叶分解贡献率, 且随雪被厚度增加而降低。可能因为面对冬季严酷环境和频繁的温度变化, 无雪被梯度下的冻融循环更为剧烈, 进而破坏凋落叶物理结构, 提高凋落叶可分解程度(徐李亚等, 2015)并促进下一阶段的分解过程。另外, 由于冬季雪被覆盖期凋落叶分解速度加快, 易分解组分大量消耗, 导致生长季时期雪被覆盖下凋落叶分解缓慢(Tan et al., 2014)。
3.2 不同雪被下雪岭云杉凋落叶碳、氮、磷含量的变化
天山地区频繁的降雪、融化以及冻融循环作用可能对凋落叶质量产生一定的影响, 继而影响整个分解过程。本研究表明, 雪岭云杉凋落叶碳、氮、磷含量对不同雪被厚度均产生了各自的响应。在冻融期, 不同雪被厚度下凋落叶碳含量表现为上升趋势且各雪被厚度间无明显差异, 可能因为冬季初期不同雪被厚度下雪岭云杉凋落叶质量完全相同, 且环境条件基本一致, 雪被厚度对凋落叶碳含量无明显影响。而凋落叶碳含量有升高趋势说明在这一时期发生了大量的质量损失, 导致不同雪被厚度下凋落叶碳含量表现出略微增加的现象。进入深冻期, 各雪被处理下凋落叶碳含量达到最低且存在显著性差异, 这是因为土壤中的分解者随雪被厚度的增加而变得更为活跃(何伟等, 2013), 该时期凋落叶碳含量降低与分解者有关(Aerts et al., 2012)。进入融冻期后, 土壤温度变化剧烈, 强烈的冻融循环和淋洗作用对凋落物叶碳含量产生影响, 到生长季时期, 凋落叶碳含量有所减少, 且不同雪被处理下凋落叶碳含量在生长季后期存在明显差异, 可能与气温逐渐升高所导致的淋溶作用以及更为活跃的分解者有关。凋落叶分解过程中氮、磷元素的释放通常被认为是为分解者提供良好底物的过程, 并从营养方面对凋落物分解过程进行调控。虽然其释放特征能够根据土壤营养情况分别表现为富集和释放, 但释放出来的氮、磷元素常被看作是凋落物对于分解者的可食性指标(Preston et al., 2009; Hobbie et al., 2012)。本研究结果表明, 随着凋落叶分解的进行, 氮含量总体表现为上升趋势; 且雪被覆盖下氮含量基本表现为增加, 这与Sing等(1999)和胡霞等(2012a)的研究结论一致, 这一结果与两个原因有关: 一是冬季无雪被梯度下昼夜热量变化在土壤表层及以下产生反复的冻结-解冻过程, 严重影响了微生物活性以及微生物种群数量和群落结构; 二是当积雪融化时, 雪被中的可溶性无机氮、有机污染物等氮源会释放大量的氮元素(胡霞等, 2012b), 造成凋落叶氮含量增加。凋落叶磷含量在冻融初期表现为上升趋势, 可能因为凋落叶中的磷含量过低导致微生物难以利用, 只能先从土壤中富集, 才能进一步分解。整个时期无雪被和薄雪被下磷含量呈逐渐降低趋势, 而中雪被和厚雪被下磷含量在生长季初期呈上升趋势, 随后在生长季后期表现为下降趋势, 这与前人所得研究结果(武启骞等, 2015)一致。在凋落叶分解过程中, 凋落叶磷含量的动态变化与凋落叶质量损失率有很强的相关性(何洁等, 2014), 凋落叶干质量损失大于磷元素释放率时, 磷含量表现出上升趋势。另外, 本研究中雪被斑块显著影响冻融期和融冻期凋落叶磷含量, 可能由于磷是易于淋溶损失的元素, 不同雪被厚度下这两个时期的淋溶差异导致凋落叶磷的快速损失。
3.3 不同雪被厚度下雪岭云杉凋落叶碳、氮、磷释放动态
凋落叶分解过程中碳释放是区域内碳循环中重要的组成过程。不同时期影响凋落叶碳释放的因素不同。以往很多****认为冬季冻结情况下凋落叶分解处于“停滞”状态, 然而近年来, 已有不少研究指出冬季冻结环境中凋落叶分解仍在进行(Mendon?a & Stott, 2003; 胡霞等, 2012b; 何伟等, 2013; 刘涛等, 2017)。深冻期的温度相对较低, 淋溶作用比较弱, 因此土壤动物、微生物等分解者的活动是这一时期凋落叶碳释放的主要贡献者(Mendon?a & Stott, 2003; 殷秀琴等, 2007; 李俊等, 2016)。本研究中融冻期凋落叶碳富集率以薄雪被和中雪被处理下最大, 可能因为淋溶作用导致凋落叶结构松散、团粒化, 更易被微生物侵入, 且微生物主导的纯分解过程比较复杂, 有时为维持自身浓度平衡, 会向凋落物分解部分输送一些元素, 导致元素发生富集现象, 表现为残留物有所增加或减少缓慢。冬季不同雪被厚度下凋落叶经历了冻融循环以及淋溶作用, 即使不易分解物质不断累积, 物理和化学过程对凋落叶表面结构的破坏也加速了凋落叶分解, 因此很大程度上冬季雪被影响着凋落叶碳释放。凋落叶分解过程中氮的变化状态分为3个过程: 淋溶(释放)、固定(N吸收)、矿化(N释放)(胡霞等, 2012b); 在Berg和McClaugherty (2008)的研究中, 早期凋落叶氮先被固定, 随后表现为净释放; 而在Bosatta和Staaf (1982)的研究中, Pinus sylvestris在分解过程中凋落叶氮表现为先释放再富集, 本研究结论与Berg和McClaugherty (2008)的研究结果不一致, 可能与凋落叶物种和土壤分解者群落的不同有关; 本研究中冻融期有雪被下氮元素富集率最大, 融冻期无雪被和厚雪被下氮元素富集率最大, 到生长季前期氮元素富集率以中雪被和厚雪被处理下最大, 出现这一现象的原因是雪岭云杉凋落叶中碳氮比的初始值为50.278, 远远大于氮元素固定和释放的临界值25 (Chapin et al., 2002), 不足以满足凋落叶中微生物分解代谢的需求, 因此微生物开始从外界土壤吸收氮元素, 大量的氮元素从土壤向凋落叶转移, 从而导致凋落叶氮表现为富集现象。在融冻期和生长季时期凋落叶氮发生富集现象, 可能是冻融期和深冻期强烈的冻融循环作用以及土壤动物破碎作用使微生物更易附着在物理结构受到破坏的凋落叶上(Cragg & Bardgett, 2001), 从而加强氮元素的固持。凋落叶磷在分解过程中不仅表现出释放规律, 而且亦被微生物所固持, 甚至从土壤中富集, 呈现释放和富集两种模式(Hobbie et al., 2012), 可能因为微生物需求和土壤养分不同。本研究中, 凋落叶磷在冻融期呈现富集状态, 这表明在早期分解者数量和活性受到冻融破碎和土壤动物取食的影响, 导致外界环境中的磷元素被分解者固持(何洁等, 2014); 而后期凋落叶磷表现为释放状态可能因为雪被具有绝热保温的功能, 为微生物活动提供了相对稳定的分解环境(Wu et al., 2010), 同时新鲜凋落叶也为微生物提供分解原料。在生长季前期中雪被和厚雪被下磷元素表现为富集状态, 可能因为林窗内强烈的阳光辐射蒸发了凋落叶表面的水分, 而水分作为凋落物分解的关键因子, 对微生物赖以生存的环境起着决定性作用, 从而抑制了凋落叶磷的释放。
综上所述, 本研究初步表明季节性雪被对中高山森林生态系统凋落叶分解过程具有显著影响。雪岭云杉凋落叶分解速率随雪被厚度的增加而增加, 由于受到不同雪被覆盖时期特有作用特征的影响, 不同雪被厚度下凋落叶碳、氮、磷含量变化特征明显不同。在全球气候变暖的背景下, 雪被厚度减小使得凋落物分解过程氮元素释放得到促进, 而磷元素释放却被抑制。这些结果为深入了解物质循环等关键生态过程在全球变化情景下对冬季环境条件改变的响应提供了一定的基础数据。

扫码加入读者圈
听语音, 看问答
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 1]

1 Decomposition of plant litter, a key component of the global carbon budget, is hierarchically controlled by the triad: climate > litter quality > soil organisms. Given the sensitivity of decomposition to temperature, especially in cold biomes, it has been hypothesized that global warming will lead to increased litter decomposition rates, both through direct temperature effects and through indirect effects on litter quality and soil organisms. 2 A meta-analysis of experimental warming studies in cold biomes (34 site-species combinations) showed that warming resulted in slightly increased decomposition rates. However, this response was strongly dependent on the method used: open top chambers reduced decomposition rates, whereas heating lamps stimulated decomposition rates. The low responsiveness was mainly due to moisture-limited decomposition rates in the warming treatments, especially at mesic and xeric sites. This control of litter decomposition by both temperature and moisture was corroborated by natural gradient studies. 3 Interspecific differences in litter quality and decomposability are substantially larger than warming-induced phenotypic responses. Thus, the changes in the species composition and structure of plant communities that have been observed in medium-term warming studies in cold biomes will have a considerably greater impact on ecosystem litter decomposition than phenotypic responses. 4 Soil fauna communities in cold biomes are responsive to climate warming. Moreover, temperature-driven migration of the, hitherto absent, large comminuters to high-latitude sites may significantly increase decomposition rates. However, we do not know how far-reaching the consequences of changes in the species composition and structure of the soil community are for litter decomposition, as there is a lack of data on functional species redundancy and the species' dispersal ability. 5 Global warming will lead to increased litter decomposition rates only if there is sufficient soil moisture. Hence, climate scenario and experimental studies should focus more on both factors and their interaction. As interspecific differences in potential decomposability and litter chemistry are substantially larger than phenotypic responses to warming, the focus of future research should be on the former. In addition, more light should be shed on the below-ground 'darkness' to evaluate the ecological significance of warming-induced soil fauna community changes for litter decomposition processes in cold biomes.
DOIURLPMID [本文引用: 2]
Litter decomposition and nutrient mineralization in high-latitude peatlands are constrained by low temperatures. So far, little is known about the effects of seasonal components of climate change (higher spring and summer temperatures, more snow which leads to higher winter soil temperatures) on these processes. In a 4-year field experiment, we manipulated these seasonal components in a sub-arctic bog and studied the effects on the decomposition and N and P dynamics of leaf litter ofCalamagrostis lapponica,Betula nana, andRubus chamaemorus, incubated both in a common ambient environment and in the treatment plots. Mass loss in the controls increased in the orderCalamagrostis<Betula<Rubus. After 4/years, overall mass loss in the climate-treatment plots was 10/% higher compared to the ambient incubation environment. Litter chemistry showed within each incubation environment only a few and species-specific responses. Compared to the interspecific differences, they resulted in only moderate climate treatment effects on mass loss and these differed among seasons and species. Neither N nor P mineralization in the litter were affected by the incubation environment. Remarkably, for all species, no net N mineralization had occurred in any of the treatments during 4/years. Species differed in P-release patterns, and summer warming strongly stimulated P release for all species. Thus, moderate changes in summer temperatures and/or winter snow addition have limited effects on litter decomposition rates and N dynamics, but summer warming does stimulate litter P release. As a result, N-limitation of plant growth in this sub-arctic bog may be sustained or even further promoted.
DOIURL [本文引用: 1]
We assessed direct and indirect effects of snow cover on litter decomposition and litter nitrogen release in alpine tundra. Direct effects are driven by the direct influence of snow cover on edaphoclimatic conditions, whereas indirect effects result from the filtering effect of snow cover on species abundance and traits. We compared the in situ decomposition of leaf litter from four dominant plant species (two graminoids, two shrubs) at early and late snowmelt locations using a two-year litter-bag experiment. A seasonal experiment was also performed to estimate the relative importance of winter and summer decomposition. We found that growth form (graminoids vs. shrubs) are the main determinants of decomposition rate. Direct effect of snow cover exerted only a secondary influence. Whatever the species, early snowmelt locations showed consistently reduced decomposition rates and delayed final stages of N mineralization. This lower decomposition rate was associated with freezing soil temperatures during winter. The results suggest that a reduced snow cover may have a weak and immediate direct effect on litter decomposition rates and N availability in alpine tundra. A much larger impact on nutrient cycling is likely to be mediated by longer term changes in the relative abundance of lignin-rich dwarf shrubs.
URL [本文引用: 1]
This dissertation addresses carbon uptake and storage by terrestrial vegetation in the Southwest Brazilian Amazon. The three-article dissertation is composed of independent, self-authored articles that are linked in their relevance to fluxes of carbon from the terrestrial biosphere.
[本文引用: 1]
DOIURL [本文引用: 1]
Disappearance of plant material from fine-mesh nylon bags occurred during the late fall, winter, or early spring when mountain ranges in central Utah are generally snow covered. Quantitative loss of matter from leaves and stems of two grasses and two broad-leaved forbs was mostly attributed to decomposition by fungi and bacteria and to leaching. Loss of material from bags in direct contact with the snow cover during two consecutive winter periods averaged 30% for the relatively coarse Agropyron trachycaulum, 39% for Bromus inermis, 48% for Lupinus alpestris, and 51% for Mertensia arizonica var, leonardi. The ratio of leaf to stem weights decreased with all aspects.
DOIURL
A "semi-empirical" model for describing the nitrogen dynamics of decomposing forest litter in a medium-long time perspective is presented. Derived results indicate the strong regulating influence of decomposition rate and initial nitrogen concentration of the litter on its retention and release of N. An increased decomposition rate tends to reduce the rate of N release per unit of carbon mineralized. This makes the "critical" C/N ratio of a litter, below which a net N mineralization takes place, lower the higher the decomposition rate. For a litter of low initial N concentration a faster decomposition generally also means a greater immobilized amount of N but a shortened immobilization phase. The peak of mineralized N is more marked and comes earlier the lower the C/N ratio and the higher the decomposition rate. Furthermore, in all litter types the C/N ratio becomes narrower as decomposition proceeds, but the change levels out earlier and on a lower level the higher the decomposition rate. The possible role of these features as regulating factors for N distribution in soil and in secondary succession of forest ecosystems is discussed. /// Разработана полуэмпирическая модель для описания динамики азота в разлагающейся лесной подстилке в течение срока средней продолжительности. Полученные результаты показывают большое регулирующее влияние скорости разложения и исходного содержания азота на его связывание и высвобождение. Повышение скорости разложения снижает скорость высвобождения азота на единицу минерализованного углерода. Это снижает критическое отношение C/N в подстилке, ниже которого имеет место минерализация азота при повышении скорости разложения. Для подстилки с низкими исходным содержанием азота более быстрые темпы разложения также означают увеличение количества иммобилизованного азота, но с укороченной фазой иммобилизации. Пик минерализованного азота более заметен и наступает тем раньше, чем ниже значение C/N и чем выше скорость разложения. Более того, во всех типах подстилки отношение C/N сужается по мере разложения, но эти изменения проявляются раньше и на более низком уровне, чем при повышении скорости разложения. Обсуждается возможная раль этих особенностей как факторов, регулирующих распределение азота в почве и значеие их для вторичной сукцессии лесных экосистем.
DOIURL [本文引用: 1]
土壤微生物多样性是指土壤生态系统中所有的微生物种类,当前研究主要集中在物种多样性、遗传多样性、结构多样性及功能多样性等4个方面。综述了土壤微生物多样性的主要研究方法及高通量测序技术的应用,分析了影响土壤微生物环境多样性的环境因子,并分析展望了土壤微生物多样性研究在生态系统功能和气候变化条件下应用的潜力。
DOIURL [本文引用: 1]
土壤微生物多样性是指土壤生态系统中所有的微生物种类,当前研究主要集中在物种多样性、遗传多样性、结构多样性及功能多样性等4个方面。综述了土壤微生物多样性的主要研究方法及高通量测序技术的应用,分析了影响土壤微生物环境多样性的环境因子,并分析展望了土壤微生物多样性研究在生态系统功能和气候变化条件下应用的潜力。
DOIURL [本文引用: 1]
Photosynthesis by plants provides the carbon and energy that drive most biological processes in ecosystems. This chapter describes the controls over this carbon input.
DOIURL [本文引用: 2]
Hekala and Buell, in their first response, argued that using data from more than one era is a grave error. They no longer raise this issue, apparently recognizing that sociobiological principles, if true, should operate across cultures and eras. They examine the conflicts described in Hen-Thorir's saga and conclude that kinship had no influence in individuals' involvements in the conflict described in this saga, and generalize to sagas in general. Hen-Thorir's saga is a very short minor saga. Another short saga is described herein and yields contradictory results. More sagas might well have been examined, and if analysis was limited to one saga, the saga chosen should have been much longer. It would have been appropriate to have examined the Orkneyinga saga as one of those that formed the content of the original paper. Issues having to do with the accuracy of those English-language versions of the sagas available to most readers are discussed.
DOIURL [本文引用: 1]
The growth of ice particles in dry seasonal snow is caused by vapor diffusion among particles due to temperature gradients imposed on the snow cover. The diffusion is calculated by using the potential field solutions for electrostatically charged particles. The stereography of snow is represented by using a log-normal distribution function for a geometrical enhancement factor defined here. Reasonable crystal growth rates and supersaturations are found. The transition between the growth of highly faceted crystals and the growth of highly rounded crystals is determined by the critical supersaturation for the onset of dislocation aided growth. Thermal convection and the continuous movement of vapor around the particles due to the imposed temperature gradient are accommodated in the theory, although both have little effect relative to the interparticle diffusion. The growth of a layer of faceted crystals just below semi-permeable crusts is explained by showing that crusts can cause the local supersaturation to exceed the critical value. Faceted crystals are shown to grow most rapidly in the lower, warmer portions of the snow cover because of temperature effects on growth rate.
DOIURL [本文引用: 1]
There are few experimental data on the consequence of varying the composition and diversity of soil animals communities, or soil food-webs, on ecosystem properties. Here, we tested the hypothesis that varying the diversity and composition of soil animals within a trophic group, the microbial-feeders, affects litter decomposition and nutrient flux in grassland. Microcosms containing grassland plant litter were inoculated with individual species of Collembola Folsomia candida, Pseudosinella alba, and Protaphorura armata,and all possible two and three species combinations of these species. Our data show that towards the end of the experiment individual species of Collembola, and especially F. candida, had markedly different, but positive, effects on measures of litter mass loss, microbial activity (CO 2 respiration) and the leaching of dissolved organic carbon (DOC) and nitrate-N. Two and three species combinations of Collembola revealed that effects of fauna on ecosystem processes were due to differences in the composition of the collembolan community, rather than the number of species present. In comparison to a treatment that had no fauna, significantly higher rates of litter mass loss, microbial activity, and DOC and nitrate release were detected only in microcosms that contained F. candida. There was no evidence of effects of F. candida in combination with other species, relative to effects of F. candida alone, on the above properties. These findings support the notion that changes in the diversity of microbivorous fauna may not have a predictable effect on decomposition processes rates and that the functioning of the microbial-feeding trophic group is influenced mainly by the physiological attributes of the dominant animal species present, in this case F. candida.
URL [本文引用: 2]
采用凋落物网袋法,研究冬季不 同关键时期雪被斑块对川西高山森林6种代表性树种凋落物分解过程中N和P释放的影响.结果表明:整个雪被覆盖季节凋落物N表现为富集,P表现为释放,且雪 被融化期P释放速率最大.厚型和中型雪被斑块下凋落物P释放速率大于薄型和无雪被斑块,而薄型和无雪被斑块下凋落物的N释放速率明显较高.6种凋落物N释 放率和释放速率与日均温呈显著负相关;除岷江冷杉外,其他树种凋落物P释放率和释放速率与日均温表现为正相关.气候变暖情景下冬季雪被覆盖的减小将促进高 山森林冬季凋落物分解过程中N释放,抑制P释放.
URL [本文引用: 2]
采用凋落物网袋法,研究冬季不 同关键时期雪被斑块对川西高山森林6种代表性树种凋落物分解过程中N和P释放的影响.结果表明:整个雪被覆盖季节凋落物N表现为富集,P表现为释放,且雪 被融化期P释放速率最大.厚型和中型雪被斑块下凋落物P释放速率大于薄型和无雪被斑块,而薄型和无雪被斑块下凋落物的N释放速率明显较高.6种凋落物N释 放率和释放速率与日均温呈显著负相关;除岷江冷杉外,其他树种凋落物P释放率和释放速率与日均温表现为正相关.气候变暖情景下冬季雪被覆盖的减小将促进高 山森林冬季凋落物分解过程中N释放,抑制P释放.
DOIURL [本文引用: 1]
To understand the effects of forest gaps on lignin degradation during shrub foliar litter decomposition, a field litterbag experiment was conducted in an alpine fir (Abies faxoniana) forest of the eastern Tibet Plateau. Dwarf bamboo (Fargesia nitida) and willow (Salix paraplesia) foliar litterbags were placed on the forest floor from the gap center to the closed canopy. The litterbags were sampled during snow formation, snow coverage, snow melting and the growing season from October 2010 to October 2012. The lignin concentrations and loss in the litter were measured. Over 2/years, lignin loss was lower in the bamboo litter (34.64-43.89%) than in the willow litter (38.91-55.10%). In the bamboo litter, lignin loss mainly occurred during the first decomposition year, whereas it occurred during the second decomposition year in the willow litter. Both bamboo and willow litter lignin loss decreased from the gap center to the closed canopy during the first year and over the entire 2-year decomposition period. Compared with the closed canopy, the gap center showed higher lignin loss for both bamboo and willow litter during the two winters, but lower lignin loss during the early growing period. Additionally, the dynamics of microbial biomass carbon during litter decomposition followed the same trend as litter lignin loss during the two winters and growing period. These results indicated that alpine forest gaps had significant effects on shrub litter lignin loss and that reduced snow cover during winter warming would inhibit shrub lignin degradation in this alpine forest.
DOIURL [本文引用: 3]
高山/亚高山森林灌木层植物凋落物的分解对于系统物质循环等过程具有重要意义,并可能受到冬季不同厚度雪被斑块下冻融格局的影响。该文采用凋落物分解袋法,研究了高山森林典型灌层植物华西箭竹(Fargesia nitida)和康定柳(Salix paraplesia)凋落物在沿林窗-林下形成的冬季雪被厚度梯度(厚型雪被斑块、较厚型雪被斑块、中型雪被斑块、薄型雪被斑块、无雪被斑块)上在第一年不同关键时期(冻结初期、冻结期、融化期、生长季节初期和生长季节后期)的质量损失特征。在整个冻融季节,华西箭竹和康定柳凋落叶的平均质量损失分别占全年的(48.78±2.35)%和(46.60±5.02)%。冻融季节雪被覆盖斑块下凋落叶的失重率表现出厚型雪被斑块大于薄型雪被斑块的趋势,而生长季节无雪被斑块的失重率明显较高。尽管如此,华西箭竹凋落物第一年分解表现出随冬季雪被厚度增加而增加的趋势,但康定柳凋落物第一年失重率以薄型雪被斑块最高,而无雪被斑块最低。同时,相关分析表明冻融季节凋落叶的失重率与平均温度和负积温呈极显著正相关,生长季节凋落叶的失重率与所调查的温度因子并无显著相关关系,但全年凋落物失重率与平均温度和正/负积温均显著相关。这些结果清晰地表明,未来冬季变暖情境下高山森林冬季雪被格局的改变将显著影响灌层植物凋落物分解,影响趋势随着物种的差异具有明显差异。
DOIURL [本文引用: 3]
高山/亚高山森林灌木层植物凋落物的分解对于系统物质循环等过程具有重要意义,并可能受到冬季不同厚度雪被斑块下冻融格局的影响。该文采用凋落物分解袋法,研究了高山森林典型灌层植物华西箭竹(Fargesia nitida)和康定柳(Salix paraplesia)凋落物在沿林窗-林下形成的冬季雪被厚度梯度(厚型雪被斑块、较厚型雪被斑块、中型雪被斑块、薄型雪被斑块、无雪被斑块)上在第一年不同关键时期(冻结初期、冻结期、融化期、生长季节初期和生长季节后期)的质量损失特征。在整个冻融季节,华西箭竹和康定柳凋落叶的平均质量损失分别占全年的(48.78±2.35)%和(46.60±5.02)%。冻融季节雪被覆盖斑块下凋落叶的失重率表现出厚型雪被斑块大于薄型雪被斑块的趋势,而生长季节无雪被斑块的失重率明显较高。尽管如此,华西箭竹凋落物第一年分解表现出随冬季雪被厚度增加而增加的趋势,但康定柳凋落物第一年失重率以薄型雪被斑块最高,而无雪被斑块最低。同时,相关分析表明冻融季节凋落叶的失重率与平均温度和负积温呈极显著正相关,生长季节凋落叶的失重率与所调查的温度因子并无显著相关关系,但全年凋落物失重率与平均温度和正/负积温均显著相关。这些结果清晰地表明,未来冬季变暖情境下高山森林冬季雪被格局的改变将显著影响灌层植物凋落物分解,影响趋势随着物种的差异具有明显差异。
DOIURL [本文引用: 2]
Despite the importance of litter decomposition for ecosystem fertility and carbon balance, key uncertainties remain about how this fundamental process is affected by nitrogen (N) availability. Resolving such uncertainties is critical for predicting the ecosystem consequences of increased anthropogenic N deposition. Toward that end, we decomposed green leaves and senesced litter of northern pin oak (Quercus ellipsoidalis) in three forested stands dominated by northern pin oak or white pine (Pinus strobus) to compare effects of substrate N (as it differed between leaves and litter) and externally supplied N (inorganic or organic forms) on decomposition and decomposer community structure and function over four years. Asymptotic decomposition models fit the data equally well as single exponential models and allowed us to compare effects of N on both the initial decomposition rate (k a ) and the level of asymptotic mass remaining (A, proportion of mass remaining at which decomposition approaches zero, i.e., the fraction of slowly decomposing litter). In all sites, both substrate N and externally supplied N (regardless of form) accelerated the initial decomposition rate. Faster initial decomposition rates corresponded to higher activity of polysaccharide-degrading enzymes associated with externally supplied N and greater relative abundances of Gram-negative and Gram-positive bacteria associated with green leaves and externally supplied organic N (assessed using phospholipid fatty acid analysis, PLFA). By contrast, later in decomposition, externally supplied N slowed decomposition, increasing the fraction of slowly decomposing litter (A) and reducing lignin-degrading enzyme activity and relative abundances of Gram-negative and Gram-positive bacteria. Higher-N green leaves, on the other hand, had lower levels of A (a smaller slow fraction) than lower-N litter. Contrasting effects of substrate and externally supplied N during later stages of decomposition likely occurred because higher-N leaves also had considerably lower lignin, causing them to decompose more quickly throughout decomposition. In conclusion, elevated atmospheric N deposition in forest ecosystems may have contrasting effects on the dynamics of different soil carbon pools, decreasing mean residence times of active fractions in fresh litter (which would be further reduced if deposition increased litter N concentrations), while increasing those of more slowly decomposing fractions, including more processed litter.
URL [本文引用: 1]
高山和高纬度地区,氮素是植物生命活动的主要限制元素之一。这类区域冬季往往被长时间的季节性雪被覆盖着。研究证实,寒冷而漫长的冬季雪被下土壤氮素在维持年际土壤氮循环中起着重要的作用,然而目前对气候变化极其敏感的青藏高原东缘雪下土壤物质转化过程的研究却很少。为了探索青藏高原东缘季节性雪被覆盖地区,冬季凋落物输入对土壤氮素转化过程的影响,2010年1—5月在青藏高原东缘(松潘卡卡沟地区)采用PVC原位培养管培养土壤,并对培养土壤进行不同的雪厚度(0、30、100 cm)处理和不同水平的凋落物添加(0、5、20 g鲜卑花叶片)处理,从实验开始后,每隔1个月采集各个处理的土壤,测定其无机氮(NH4+-N和NO3--N)含量,并计算净氮矿化率,以探讨冬季季节性雪被覆盖下不同碳供应水平对高山土壤氮转化过程的动态影响。研究发现,雪被覆盖能有效地绝缘大气和土壤,减少冻融交替的幅度和频次,并加速了土壤的净氮矿化。说明对于雪被覆盖的高山土壤而言冷季是氮素循环的关键时期。冬季一定厚度的积雪覆盖可通过调节整个土壤氮素的矿化水平,从而为来年春季高山植物的生长提供一个巨大的潜在氮库。添加大量凋落物显著增加了NO3--N含量,降低了NH4+-N含量,加速了土壤净氮矿化。暗示在具有高有机质含量的青藏高原东部地区,土壤微生物的生长和活性极有可能仍然受到低水平可利用碳的限制。
URL [本文引用: 1]
高山和高纬度地区,氮素是植物生命活动的主要限制元素之一。这类区域冬季往往被长时间的季节性雪被覆盖着。研究证实,寒冷而漫长的冬季雪被下土壤氮素在维持年际土壤氮循环中起着重要的作用,然而目前对气候变化极其敏感的青藏高原东缘雪下土壤物质转化过程的研究却很少。为了探索青藏高原东缘季节性雪被覆盖地区,冬季凋落物输入对土壤氮素转化过程的影响,2010年1—5月在青藏高原东缘(松潘卡卡沟地区)采用PVC原位培养管培养土壤,并对培养土壤进行不同的雪厚度(0、30、100 cm)处理和不同水平的凋落物添加(0、5、20 g鲜卑花叶片)处理,从实验开始后,每隔1个月采集各个处理的土壤,测定其无机氮(NH4+-N和NO3--N)含量,并计算净氮矿化率,以探讨冬季季节性雪被覆盖下不同碳供应水平对高山土壤氮转化过程的动态影响。研究发现,雪被覆盖能有效地绝缘大气和土壤,减少冻融交替的幅度和频次,并加速了土壤的净氮矿化。说明对于雪被覆盖的高山土壤而言冷季是氮素循环的关键时期。冬季一定厚度的积雪覆盖可通过调节整个土壤氮素的矿化水平,从而为来年春季高山植物的生长提供一个巨大的潜在氮库。添加大量凋落物显著增加了NO3--N含量,降低了NH4+-N含量,加速了土壤净氮矿化。暗示在具有高有机质含量的青藏高原东部地区,土壤微生物的生长和活性极有可能仍然受到低水平可利用碳的限制。
URL [本文引用: 5]
2010年1—5月在川西高原采用人工雪厚度梯度试验(0、30和100cm),应用网袋分解法对窄叶鲜卑花叶片凋落物进行分解试验,测定了凋落物的分解速率及其养分动态.结果表明:在无雪被覆盖的样地上分解5个月后的凋落物质量损失率为29.9%,而中雪和深雪样地的凋落物质量损失率分别为33.8%和35.2%.分解过程中,凋落物氮存在一定的富集现象,磷处于波动的富集状态,碳质量分数和碳氮比均呈现前期急剧下降后期逐渐上升的趋势.雪被覆盖显著增加了凋落物的质量损失率和氮含量,而对碳和磷含量无显著影响.在川西高原地区,30cm以上的持续雪被覆盖能够改变凋落物的分解过程,从而可能对土壤营养物质转化和植物群落构建产生实质性的影响.
URL [本文引用: 5]
2010年1—5月在川西高原采用人工雪厚度梯度试验(0、30和100cm),应用网袋分解法对窄叶鲜卑花叶片凋落物进行分解试验,测定了凋落物的分解速率及其养分动态.结果表明:在无雪被覆盖的样地上分解5个月后的凋落物质量损失率为29.9%,而中雪和深雪样地的凋落物质量损失率分别为33.8%和35.2%.分解过程中,凋落物氮存在一定的富集现象,磷处于波动的富集状态,碳质量分数和碳氮比均呈现前期急剧下降后期逐渐上升的趋势.雪被覆盖显著增加了凋落物的质量损失率和氮含量,而对碳和磷含量无显著影响.在川西高原地区,30cm以上的持续雪被覆盖能够改变凋落物的分解过程,从而可能对土壤营养物质转化和植物群落构建产生实质性的影响.
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
于2013年11月至2014年4月,采用微生物磷酸脂肪酸(PLFA)分析方法,以川西高山草甸代表物种黄花亚菊(Ajania nubigena)和黑褐苔草(Carex atrofusca)凋落物为研究对象,在雪被形成期、雪被稳定期和雪被融化期研究了凋落物分解过程中土壤动物对微生物群落结构及其多样性的影响.结果表明:(1)整个冬季,土壤动物增加了黄花亚菊凋落物分解过程中微生物总P LFA含量、真菌PLFA含量以及细菌PLFA含量,提高了其真菌/细菌比例,减少了黑褐苔草中微生物总PLFA含量、真菌PLFA含量以及细菌PLFA含量,降低了其真菌/细菌比例;(2)在雪被形成期,黄花亚菊凋落物中土壤动物显著增加了真菌表征脂肪酸18:2ω6,9c的含量,但降低了细菌表征脂肪酸16:0、16:1ω9c、18:1ω7c的含量.黑褐苔草凋落物中土壤动物显著增加了细菌表征脂肪酸18:1ω7c的含量,但降低了真菌表征脂肪酸18:2ω6,9c的含量.雪被融化期,土壤动物显著增加了黑褐苔草凋落物中细菌表征脂肪酸16:1ω9c的含量,降低了细菌表征脂肪酸a15:0和微真核生物表征脂肪酸18:3和20:4的含量;(3)土壤动物对凋落物中微生物多样性的影响主要在雪被融化期,增加了黄花亚菊凋落物中微生物多样性、均匀性、丰富度和优势度指数,降低了黑褐苔草凋落物中微生物多样性和均匀性指数,增加了黑褐苔草凋落物中微生物丰富度和优势度指数.这些结果表明,冬季恶劣环境条件下高山草甸土壤动物仍然对微生物的数量、群落结构和多样性具有显著的影响.
DOIURL [本文引用: 1]
于2013年11月至2014年4月,采用微生物磷酸脂肪酸(PLFA)分析方法,以川西高山草甸代表物种黄花亚菊(Ajania nubigena)和黑褐苔草(Carex atrofusca)凋落物为研究对象,在雪被形成期、雪被稳定期和雪被融化期研究了凋落物分解过程中土壤动物对微生物群落结构及其多样性的影响.结果表明:(1)整个冬季,土壤动物增加了黄花亚菊凋落物分解过程中微生物总P LFA含量、真菌PLFA含量以及细菌PLFA含量,提高了其真菌/细菌比例,减少了黑褐苔草中微生物总PLFA含量、真菌PLFA含量以及细菌PLFA含量,降低了其真菌/细菌比例;(2)在雪被形成期,黄花亚菊凋落物中土壤动物显著增加了真菌表征脂肪酸18:2ω6,9c的含量,但降低了细菌表征脂肪酸16:0、16:1ω9c、18:1ω7c的含量.黑褐苔草凋落物中土壤动物显著增加了细菌表征脂肪酸18:1ω7c的含量,但降低了真菌表征脂肪酸18:2ω6,9c的含量.雪被融化期,土壤动物显著增加了黑褐苔草凋落物中细菌表征脂肪酸16:1ω9c的含量,降低了细菌表征脂肪酸a15:0和微真核生物表征脂肪酸18:3和20:4的含量;(3)土壤动物对凋落物中微生物多样性的影响主要在雪被融化期,增加了黄花亚菊凋落物中微生物多样性、均匀性、丰富度和优势度指数,降低了黑褐苔草凋落物中微生物多样性和均匀性指数,增加了黑褐苔草凋落物中微生物丰富度和优势度指数.这些结果表明,冬季恶劣环境条件下高山草甸土壤动物仍然对微生物的数量、群落结构和多样性具有显著的影响.
[本文引用: 3]
[本文引用: 3]
DOIURL [本文引用: 1]
The winter climate is changing in many parts of the world, and it is predicted that winter climate change will modify the structure and function of plant–soil systems. An understanding of these changes and their consequences in terrestrial ecosystems requires knowledge of the linkage between above- and below-ground components as well as the species interactions found in plant–soil systems, which have important implications for biogeochemical cycles. However, winter climate-change studies have focused on only a part of the ecosystem or ecological process. We summarize here recent findings related to the effects of winter climate and its changes on soil nitrogen (N) dynamics, greenhouse gas (N 2 O) emissions from the soil, N use by individual plants, vegetation development, and interactions between vegetation and pollinators to generate an integrative understanding of the response of the plant–soil system to winter climate change. This review indicates that the net effects on plants, soil microbes, pollinators, and the associated biogeochemical cycles are balanced among several processes and are highly variable depending on the context, such as the target species/functional group, original winter condition of the habitat, and type of climate change. The consequences of winter climate change for species interactions among plants, associated animals, and biogeochemical cycles are largely unknown. For further research, a large-scale comparative study to measure ecosystem-level functions is important, especially in less-cold ecosystems.
DOIURL [本文引用: 2]
In the Zona da Mata Mineira of Southeastern Brazil the development of sustainable land requires the integration of crops with trees. The objectives of this study then were to (i) characterize prunings from the main tree species in an agroforestry system; (ii) determine the effects of the physical and chemical characteristics of the prunings on their decomposition patterns in the laboratory; (iii) assess the effect of mixing leaves of different species on decomposition rates; and (iv) propose a decomposition index for the residues studied. The study was carried out with pruning residues from Cajanus cajan, Solanum variable, Cassia ferruginea, Piptadenia gonoacantha, Croton urucurana , and Melinis multiflora . The materials were characterized for total C, N, P, Ca, Mg and K contents; lignin, cellulose, hemicellulose and soluble polyphenols contents. The pruning residues had high polyphenols and lignin contents, high C:N and C:P ratios, and low contents of Ca, Mg, and K. The low decomposition rates of the prunings were related to the P, K, hemicellulose and polyphenol contents. The rates of N mineralization from most of the residues indicate that there is a potential to supply the needs of a crop of maize. The residues of some species, if decomposed alone, would not supply sufficient nutrients, and need to be mixed with leaves of other species.
DOIURL [本文引用: 3]
See full-text article at JSTOR
DOIURLPMID [本文引用: 2]
Litter decomposition provides the primary source of mineral nitrogen (N) for biological activity in most terrestrial ecosystems. A 10-year decomposition experiment in 21 sites from seven biomes found that net N release from leaf litter is dominantly driven by the initial tissue N concentration and mass remaining regardless of climate, edaphic conditions, or biota. Arid grasslands exposed to high ultraviolet radiation were an exception, where net N release was insensitive to initial N. Roots released N linearly with decomposition and exhibited little net N immobilization. We suggest that fundamental constraints on decomposer physiologies lead to predictable global-scale patterns in net N release during decomposition.
DOIURL [本文引用: 1]
Abstract Deforestation and forest degradation are driving unprecedented declines in biodiversity across the tropics, and understanding the consequences of these changes for ecosystem functioning is essential for human well-being. Forest degradation and loss alter ecosystem functioning through changes in species composition and abiotic conditions. However, the consequences of these changes for heterospecific processes are often poorly understood. Leaf litter decomposition is a major source of atmospheric carbon and critical for carbon and nutrient cycling. Through a highly replicated litter-bag experiment (3360 bags), we quantified the effects of litter quality, decomposer functional diversity and seasonal precipitation regime on litter decomposition along a tropical disturbance gradient in SW China. In addition, using soil and litter from sites selected from across the disturbance gradient, we established replicated litter-bed treatments and exposed these to a gradient of simulated canopy cover in a shade-house. Across the landscape, mass loss from litter-bags after 12 months varied from 7% to 98%. Even after 12 months, litter-bags installed at the beginning of the dry season had much lower mass loss than those installed at the beginning of the wet season. As expected, litter quality and faunal exclusion had substantial effects on decomposition rates. Decomposition rates declined along the disturbance gradient from mature forest, through regenerating forest to open land, although the effect size was strongly dependent on installation season. The effect of excluding meso- and macro-invertebrates increased with increasing forest degradation, whereas the effect of litter quality declined. Results from the shade-house experiment strongly suggested that forest degradation effects were driven predominantly by changes in micro-climatic conditions resulting from increased canopy openness. To better model the impacts of anthropogenic global change on litter decomposition rates, it will be important to consider landscape scale processes, such as forest degradation.
DOIURL [本文引用: 2]
DOIURL [本文引用: 1]
AbstractAimsClimate-induced changes in snow cover are likely to affect cold arctic and alpine ecosystems functioning and major processes such as wintertime plant litter decomposition. However, it remains poorly studied in subalpine systems where the snowpack may be irregular. In this paper we explored the dynamic of the winter plant litter decomposition process, its magnitude and its relationship with the snowpack properties.MethodsIn subalpine grasslands of the Central French Alps, we performed a litter bag experiment monitoring over a whole winter the litter decomposition from the exploitative ResultsLitter decomposition rates were stable during winter and 3-fold higher under deeper and permanent snowpack with higher thermal resistance. Litter quality appeared only significant under thinner snowpack with higher decomposition rates for the exploitative species. A snowpack with higher thermal resistance created an insulating layer promoting the decomposition process.ConclusionThese results suggest that the temporal (permanence vs. intermittency) and physical (depth and thermal resistance) characteristics of the snowpack should be considered when studying the response of winter ecosystems functioning to global changes.
DOIURL
DOIURL [本文引用: 1]
凋落物分解是陆地生态系统物质循环和能量转换的主要途径。凋落物分解主要受到基质质量和气候因素多因子的综合影响。目前国内尚缺乏关于凋落物分解同气候因子和基质质量关系的多元统计和综合分析。应用分解袋法,对亚热带8个主要树种的凋落叶沿中国东部气候带5个地点历时2a的分解试验研究表明,年均降水量是影响中国东部凋落叶分解速率的首要气候因子,其次是实际蒸散和年均温度。凋落叶的初始N含量是决定分解快慢的首要基质因子,其次是P含量和Lignin∶N比和C∶N比。
DOIURL [本文引用: 1]
凋落物分解是陆地生态系统物质循环和能量转换的主要途径。凋落物分解主要受到基质质量和气候因素多因子的综合影响。目前国内尚缺乏关于凋落物分解同气候因子和基质质量关系的多元统计和综合分析。应用分解袋法,对亚热带8个主要树种的凋落叶沿中国东部气候带5个地点历时2a的分解试验研究表明,年均降水量是影响中国东部凋落叶分解速率的首要气候因子,其次是实际蒸散和年均温度。凋落叶的初始N含量是决定分解快慢的首要基质因子,其次是P含量和Lignin∶N比和C∶N比。
DOIURL [本文引用: 1]
全球环境变化将对森林生态系统凋落物的分解和养分循环产生直接和间接的多重影响。就全球环境变化如全球变暖、大气CO2浓度升高、UV-B辐射增强、氮沉降等对凋落物分解影响的研究进展进行了综合述评。影响凋落物分解的内部因素为凋落物基质质量,外部因素包括生物因素(微生物和动物)和非生物因素(温度、水分和土壤性质等)。全球变暖对凋落物分解的非生物作用有正效应,也有负效应。全球变暖对凋落物化学组成虽然只有轻微的影响,但可以通过影响植被的物种组成来间接改变凋落物的产量、化学性质和分解。全球变暖对凋落物分解生物作用的主要影响是增强土壤微生物活性,从而加速凋落物的分解。CO2浓度上升将增加凋落物产量,并通过影响凋落物质量(提高C/N比、木质素/N比等)和生物环境(微生物的数量和活性)而影响分解过程。UV-B辐射和大气N沉降的增加亦对凋落物分解产生直接和间接的影响,但影响效果尚不很清楚,有待进一步的研究。总起来看,全球环境变化将通过影响凋落物的分解速率而对全球碳循环产生重要影响,但由于气候变化和凋落物分解响应的复杂性以及各因子之间的相互作用,气候变化对凋落物分解的总效应尚需更深入的研究来定量化。
DOIURL [本文引用: 1]
全球环境变化将对森林生态系统凋落物的分解和养分循环产生直接和间接的多重影响。就全球环境变化如全球变暖、大气CO2浓度升高、UV-B辐射增强、氮沉降等对凋落物分解影响的研究进展进行了综合述评。影响凋落物分解的内部因素为凋落物基质质量,外部因素包括生物因素(微生物和动物)和非生物因素(温度、水分和土壤性质等)。全球变暖对凋落物分解的非生物作用有正效应,也有负效应。全球变暖对凋落物化学组成虽然只有轻微的影响,但可以通过影响植被的物种组成来间接改变凋落物的产量、化学性质和分解。全球变暖对凋落物分解生物作用的主要影响是增强土壤微生物活性,从而加速凋落物的分解。CO2浓度上升将增加凋落物产量,并通过影响凋落物质量(提高C/N比、木质素/N比等)和生物环境(微生物的数量和活性)而影响分解过程。UV-B辐射和大气N沉降的增加亦对凋落物分解产生直接和间接的影响,但影响效果尚不很清楚,有待进一步的研究。总起来看,全球环境变化将通过影响凋落物的分解速率而对全球碳循环产生重要影响,但由于气候变化和凋落物分解响应的复杂性以及各因子之间的相互作用,气候变化对凋落物分解的总效应尚需更深入的研究来定量化。
DOIURL [本文引用: 1]
Projected future decreases in snow cover associated with global warming in alpine ecosystems could affect soil biochemical cycling. To address the objectives how an altered snow removal could affect soil microbial biomass and enzyme activity related to soil carbon and nitrogen cycling and pools, plastic film coverage and returning of melt snow water were applied to simulate the absence of snow cover in a Tibetan alpine forest of western China. Soil temperature and moisture, nutrient availability, microbial biomass and enzyme activity were measured at different periods (before snow cover, early snow cover, deep snow cover, snow cover melting and early growing season) over the entire 2009/2010 winter. Snow removal increased the daily variation of soil temperature, frequency of freeze haw cycle, soil frost depth, and advanced the dates of soil freezing and melting, and the peak release of inorganic N. Snow removal significantly decreased soil gravimetric water, ammonium and inorganic N, and activity of soil invertase and urease, but increased soil nitrate, dissolve organic C (DOC) and N (DON), and soil microbial biomass C (MBC) and N (MBN). Our results suggest that a decreased snow cover associated with global warming may advance the timing of soil freezing and thawing as well as the peak of releases of nutrients, leading to an enhanced nutrient leaching before plant become active. These results demonstrate that an absence of snow cover under global warming scenarios will alter soil microbial activities and hence element biogeochemical cycling in alpine forest ecosystems.
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
Repeated cycles of freezing and thawing of leaf litter could accelerate decomposition if they caused physical damage to leaves or chemical changes which improve access or render leaf material more easily degradable by decomposers. Moistened leaf litter of aspen ( Populus tremuloides Michx.) and pine ( Pinus contorta Loud. × P. banksiana Lamb.), which had been frozen and thawed 14 times, absorbed water faster than litter frozen for the same total duration, before thawing once. Aspen, but not pine, began leaching sooner. Total solubles content of aspen leaves was reduced 8% by repeated freezing and thawing, but leaching loss from pine needles doubled from 4 to 8% of total mass. Previously frozen and thawed litter (14 cycles) decomposing at 10° C in laboratory microcosms lost more mass initially than litter frozen only once, but the difference was not maintained after 2 or 3 months decay. Litter exposed in microcosms to + 10°C (days) and 616°C (nights) for 3 months lost mass no faster or slower than litter maintained at the same mean temperature (+2° C) without variation, once differences in moisture content were allowed for. However, mass losses from frozen and thawed litter were substantially greater than expected if losses are prorated for the fact that these leaves were frozen 12 h each day. This difference reflects either the acceleration of decomposition due to freezing-thawing, possibly arising from weakening of the cuticle, or continuing decomposition at below-zero temperatures. For aspen litter, this effect was much smaller after 1 month of nightly freezing, followed by 2 months at 10° C. Frequency of freeze-thaw cycles in the field was estimated at 2–11 cycles month 611 during spring or fall at the aspen field site, and 0–8 cycles month 611 at the pine site. Evidence indicates that simple freezing (as opposed to freezing-thawing) and decomposer activity beneath snow may be more important factors in winter time litter decomposition than freeze-thaw cycles.
DOIURL [本文引用: 1]
The snow-bed vegetation in the Northern Apennines was studied by the Braun-Blanquet method. The phytosociological relev茅s were classified by numerical methods. Four main vegetation types were identified: Salicetum herbaceae, community of Carex foetida, Poo-Cerastietum cerastioidis and Oligotricho-Gnaphalietum supini. The latter was described as a new association. The vegetation types were ecologically characterized by an indirect gradient analysis based on principal component analysis. The results were interpreted by means of: i) geomorphological observations, ii) soil profiles, iii) measurements of the length of the snow-free period.
DOIURL [本文引用: 2]
Winter CO 2 emissions from soil and snow-covered surfaces can represent a significant fraction of the annual carbon cycle in cool-temperate broad-leaved deciduous forests. The sources of this CO 2 production are, however, poorly understood. To clarify the role of saprophytic microorganisms in the carbon cycle during the winter (December–April), mass loss rates of organic substrates (leaf litter and cellulose sheets), and microbial activity and fungal biomass in the litter layer of a cool-temperate broad-leaved deciduous forest in Japan were investigated. The mass loss rate of leaves reached 13%, which accounted for 26% of the annual mass loss. Microbial respiration in the leaf litter, measured in the laboratory, was detected even at 612 °C. The ergosterol content (an indicator of fungal biomass) of the cellulose sheets increased significantly in the snow-covered season. Seven genera of fungi including Varicosporium elodeae, Epicoccum purpurascens and Cylindrocarpon destructans were isolated from leaf litter collected from under snow cover. Microbial respiration during the winter was estimated based on the relationships between the amount of leaf litter available, the temperature dependency of microbial respiration and the temperature in the litter layer. The microorganisms in the litter layer emitted 31 gCO 2-C m 612, which accounts for 38% of the total CO 2 efflux during the winter. These results suggest that through soil respiration, microorganisms in the litter layer play an important role in the carbon cycle during the winter.
DOIURL [本文引用: 1]
We determined controls on litter decomposition and nutrient release for the widespread native tree Metrosideros polymorpha in 11 sites arrayed on gradients of elevation, precipitation, and substrate age on Hawaiian lava flows. The effects of site characteristics were evaluated using three common substrates (Metrosideros leaf litter from one of the sites, wood dowels, and filter paper) decomposed in each of the sites, and the inherent decomposability of tissue (substrate quality) was evaluated using Metrosideros leaf litter from each of the sites decomposed in a common site. Site characteristics were responsible for most of the variation in rates of decomposition in the range of sites and substrates examined. Common substrates decomposed much more rapidly in warm, low elevation sites; apparent Q"1"0 values, calculated on the basis of variation in mean annual temperature with elevation on individual lava flows, ranged from 4 to 11. Litter decomposed slowly in the dry sites, but leaf litter produced in the dry sites decomposed more than twice as rapidly as litter from wet sites when both were measured in the same site. The higher substrate quality of litter from dry sites could be due to trade-offs among nutrient-use efficiency, water-use efficiency, and carbon gain by water-limited Metrosideros. We used these results to test a revision of the CENTURY soil organic matter model that had been designed to simulate the decomposition of surface litter. Simulations accurately matched the pattern but underestimated the magnitude of among-site differences in the decomposition of common substrates in a range of sites. Analyses of both field and simulation results suggested that the decomposition of Metrosideros leaf litter could be limited by nitrogen availability.
DOIURL [本文引用: 2]
Mass loss and nutrient release of forest litter during the freeze–thaw season could play an essential role in C and nutrient cycling in cold regions, but few studies in some key ecosystems have been available. In order to characterize litter decomposition during the freeze–thaw season in a subalpine forest region of western China, a field experiment using the litterbag method was conducted on the decomposition of foliar litter of two dominant species, fir ( Abies faxoniana) and birch ( Betula platyphylla) under their respective forests. Over the freeze–thaw season following leaf-fall, about 18% and 20% of mass, 13% and 14% of lignin, 30% and 26% of cellulose, 14% and 21% of C, 30% and 27% of N, 17% and 15% of P, and 17% and 13% of K were lost from fir and birch litters, respectively. The lost mass and components accounted for more than 64% and 65% of mass, 72% and 69% of lignin, 75% and 60% of cellulose, 49% and 59% of C, 56% and 71% of N, 62% and 37% of P, and 38% and 37% of K in 1 year net loss rate of fir and birch litter, respectively. In addition, the loss of mass, lignin, cellulose and component bio-elements during the freeze–thaw season correlated closely with the initial substrate type and the levels of the individual bio-elements. The results demonstrated that litter decomposition during the freeze–thaw season contributes significantly to the first year decomposition in these subalpine forests.
DOIURL [本文引用: 4]
AimMethodsImportant findings values from the Olson decomposition constant for three coniferous litters (fir, larch and cypress) were highest under thick snow cover and lowest under no snow cover. However, the value for birch, a broad-leaved species, showed the ranked order of no snow cover﹥thin snow cover﹥thicker snow cover﹥thick snow cover﹥medium snow cover. Although snow cover did not significantly promote decomposition of birch litter during growing season in the second year, snow cover significantly promoted decomposition of fir, larch and cypress litters at all investigated stages in two years. Additionally, mass loss during snow cover period in the first year accounted for 42.5%–65.5% of the entire first year decomposition, indicating that seasonal snow cover dramatically changed the decomposition of leaf litters in winter, especially at the deep frozen stage. In conclusion, litter decomposition in this alpine forest would be delayed by the decrease of winter snow cover predicted with climate change. Compared with broad-leaved litter, coniferous litter could display stronger responses to such changes of snow cover.
DOIURL [本文引用: 4]
AimMethodsImportant findings values from the Olson decomposition constant for three coniferous litters (fir, larch and cypress) were highest under thick snow cover and lowest under no snow cover. However, the value for birch, a broad-leaved species, showed the ranked order of no snow cover﹥thin snow cover﹥thicker snow cover﹥thick snow cover﹥medium snow cover. Although snow cover did not significantly promote decomposition of birch litter during growing season in the second year, snow cover significantly promoted decomposition of fir, larch and cypress litters at all investigated stages in two years. Additionally, mass loss during snow cover period in the first year accounted for 42.5%–65.5% of the entire first year decomposition, indicating that seasonal snow cover dramatically changed the decomposition of leaf litters in winter, especially at the deep frozen stage. In conclusion, litter decomposition in this alpine forest would be delayed by the decrease of winter snow cover predicted with climate change. Compared with broad-leaved litter, coniferous litter could display stronger responses to such changes of snow cover.
DOIURL [本文引用: 1]
雪被是影响高海拔森林凋落物分解的重要生态因子,其是否影响到生长季节与非生长季节凋落物中的P元素释放,尚未量化。为了量化季节性雪被对高海拔森林凋落物分解过程中P元素释放的影响,于2010年10月至2012年10月间,在青藏高原东缘川西高海拔森林不同厚度冬季雪被斑块下,设置凋落物分解袋实验。检测该地区代表性树种岷江冷杉(Abies faxoniana)、红桦(Betula albo-sinensis)、四川红杉(Larix mastersiana)和方枝柏(Sabina saltuaria)凋落叶在雪被覆盖不同关键时期(雪被形成前期、完全覆盖期和消融期)以及生长季节的P元素动态。结果表明,凋落物质量与雪被厚度均显著影响了P元素的释放过程。雪被覆盖时期凋落物P元素释放率表现为有雪被覆盖大于无雪被覆盖,而生长季节中除岷江冷杉外的其他3种凋落物P元素释放率均为无雪被覆盖下最大。相对于无雪被覆盖斑块,冬季雪被的存在提供了保护绝缘层,促进凋落物P元素释放,提高了各物种冬季P元素释放贡献率。这些结果表明,全球变化情景下的雪被减少可能减缓高海拔森林凋落物P元素的释放过程,改变森林土壤P元素水平。所以在研究高寒、高海拔地区全球气候变化下生态系统功能的工作中,应注重雪被这一异质性环境因子对生态系统功能的影响。
DOIURL [本文引用: 1]
雪被是影响高海拔森林凋落物分解的重要生态因子,其是否影响到生长季节与非生长季节凋落物中的P元素释放,尚未量化。为了量化季节性雪被对高海拔森林凋落物分解过程中P元素释放的影响,于2010年10月至2012年10月间,在青藏高原东缘川西高海拔森林不同厚度冬季雪被斑块下,设置凋落物分解袋实验。检测该地区代表性树种岷江冷杉(Abies faxoniana)、红桦(Betula albo-sinensis)、四川红杉(Larix mastersiana)和方枝柏(Sabina saltuaria)凋落叶在雪被覆盖不同关键时期(雪被形成前期、完全覆盖期和消融期)以及生长季节的P元素动态。结果表明,凋落物质量与雪被厚度均显著影响了P元素的释放过程。雪被覆盖时期凋落物P元素释放率表现为有雪被覆盖大于无雪被覆盖,而生长季节中除岷江冷杉外的其他3种凋落物P元素释放率均为无雪被覆盖下最大。相对于无雪被覆盖斑块,冬季雪被的存在提供了保护绝缘层,促进凋落物P元素释放,提高了各物种冬季P元素释放贡献率。这些结果表明,全球变化情景下的雪被减少可能减缓高海拔森林凋落物P元素的释放过程,改变森林土壤P元素水平。所以在研究高寒、高海拔地区全球气候变化下生态系统功能的工作中,应注重雪被这一异质性环境因子对生态系统功能的影响。
DOIURL [本文引用: 1]
冬季凋落物的质量损失是中高纬度和高海拔地区凋落物分解的关键,但冬季凋落物分解是否与土壤动物的贡献有关,不同冻融时期(冻融初期、深冻期和融化期)的土壤动物对凋落物分解的贡献是否存在差异?对这两个问题仍缺乏必要的关注。为了解季节性冻融期间土壤动物对岷江冷杉(Abies faxoniana)凋落物分解的贡献,采用凋落物分解袋法,调查了季节性冻融期间(2010年10月底至2011年4月中旬),不同网孔(0.020mm、0.125mm、1.000mm和3.000mm)凋落物分解袋内的岷江冷杉凋落叶质量损失,分析了微型、中型和大型土壤动物对岷江冷杉凋落叶分解的贡献。在季节性冻融期间,0.020mm、0.125mm、1.000mm和3.000mm分解袋内的岷江冷杉凋落叶质量损失率分别为12.13%、13.07%、14.95%和18.74%。不同体径的土壤动物对季节性冻融期间岷江冷杉凋落叶质量损失的贡献率总共为35.28%;不同孔径凋落物袋内土壤动物的类群和个体相对密度与凋落叶的质量损失率呈现相对一致的变化趋势。在季节性冻融的3个阶段中,土壤动物对岷江冷杉凋落叶质量损失的贡献率均为:微型土壤动物中型土壤动物大型土壤动物。其中,微型、中型和大型土壤动物分别在深冻期、冻融初期和融化期表现出最高的贡献率,分别为6.56%、11.77%和21.94%。然而相对于其他冻融时期,深冻期中型和大型土壤动物对岷江冷杉凋落叶质量损失的贡献率最低。这些结果清晰地表明了川西高山季节性冻融期间土壤动物调控着凋落物分解的生态过程,是高山冬季凋落物分解的重要因素之一。
DOIURL [本文引用: 1]
冬季凋落物的质量损失是中高纬度和高海拔地区凋落物分解的关键,但冬季凋落物分解是否与土壤动物的贡献有关,不同冻融时期(冻融初期、深冻期和融化期)的土壤动物对凋落物分解的贡献是否存在差异?对这两个问题仍缺乏必要的关注。为了解季节性冻融期间土壤动物对岷江冷杉(Abies faxoniana)凋落物分解的贡献,采用凋落物分解袋法,调查了季节性冻融期间(2010年10月底至2011年4月中旬),不同网孔(0.020mm、0.125mm、1.000mm和3.000mm)凋落物分解袋内的岷江冷杉凋落叶质量损失,分析了微型、中型和大型土壤动物对岷江冷杉凋落叶分解的贡献。在季节性冻融期间,0.020mm、0.125mm、1.000mm和3.000mm分解袋内的岷江冷杉凋落叶质量损失率分别为12.13%、13.07%、14.95%和18.74%。不同体径的土壤动物对季节性冻融期间岷江冷杉凋落叶质量损失的贡献率总共为35.28%;不同孔径凋落物袋内土壤动物的类群和个体相对密度与凋落叶的质量损失率呈现相对一致的变化趋势。在季节性冻融的3个阶段中,土壤动物对岷江冷杉凋落叶质量损失的贡献率均为:微型土壤动物中型土壤动物大型土壤动物。其中,微型、中型和大型土壤动物分别在深冻期、冻融初期和融化期表现出最高的贡献率,分别为6.56%、11.77%和21.94%。然而相对于其他冻融时期,深冻期中型和大型土壤动物对岷江冷杉凋落叶质量损失的贡献率最低。这些结果清晰地表明了川西高山季节性冻融期间土壤动物调控着凋落物分解的生态过程,是高山冬季凋落物分解的重要因素之一。
DOIURL [本文引用: 1]
Soluble carbon during foliar litter decomposition in alpine forests plays an important role in carbon cycling in ecosystems, which is also closely related to the material and energy flow in downstream ecosystems. However, forest gaps could regulate soluble carbon dynamics during litter decomposition by changing the frequency of soil freeze-thaw cycles in winter and hydrothermal environment in growing seasons, but little information has been available. Therefore, a field litterbag experiment was conduct in an alpine forest in western Sichuan Province, China from 15 November 2012 to 31 October 2013. Foliar litter of typical arbor species (birch: Betula albo-sinensis, cypress: Sabina saltuaria, larch: Larix mastersiana and fir: Abies faxoniana) and shrub species (willow: Salix paraplesia, azalea: Rhododendron lapponicum) were selected. Samples of air-dried foliar litter were filled in nylon litterbags and placed on the forest floor with 2 cm spacing between litterbags from the gap center, canopy gap edge, extended gap edge to closed canopy in the alpine forest. The litterbags were sampled at snow formation stage, snow cover stage, snow melt stage and growing season in the first year of decomposition. The results indicated that the content of soluble carbon and soluble organic carbon from the foliar litter of the six species showed a similar dynamical tendency at the first year of litter decomposition. Both contents of soluble carbon and soluble organic carbon increased in winter, and decreased in growing season, though the content of soluble inorganic carbon decreased in the first year of litter decomposition. The maximum contents of soluble carbon and soluble organic carbon in closed canopy were observed at snow formation stage regardless of litter species, but those in gap center and extended gap edge were observed at snow cover stage. In contrast, the contents of soluble carbon, soluble organic carbon and soluble inorganic carbon decreased significantly in growing season, even less than the initial contents. Multivariate analysis displayed that the contents of soluble carbon and soluble inorganic carbon were significantly affected by forest gap in the first year of litter decomposition, but the content of soluble organic carbon only affected by forest gap in the winter. Moreover, the contents of soluble carbon and soluble organic carbon were influenced particularly by litter quality in the winter. These results suggested that the ongoing winter warming would decrease the content of soluble carbon during foliar litter decomposition by decreasing the cover-time and thickness of snow in the winter, although the decrease degree could be controlled by litter quality.
DOIURL [本文引用: 1]
Soluble carbon during foliar litter decomposition in alpine forests plays an important role in carbon cycling in ecosystems, which is also closely related to the material and energy flow in downstream ecosystems. However, forest gaps could regulate soluble carbon dynamics during litter decomposition by changing the frequency of soil freeze-thaw cycles in winter and hydrothermal environment in growing seasons, but little information has been available. Therefore, a field litterbag experiment was conduct in an alpine forest in western Sichuan Province, China from 15 November 2012 to 31 October 2013. Foliar litter of typical arbor species (birch: Betula albo-sinensis, cypress: Sabina saltuaria, larch: Larix mastersiana and fir: Abies faxoniana) and shrub species (willow: Salix paraplesia, azalea: Rhododendron lapponicum) were selected. Samples of air-dried foliar litter were filled in nylon litterbags and placed on the forest floor with 2 cm spacing between litterbags from the gap center, canopy gap edge, extended gap edge to closed canopy in the alpine forest. The litterbags were sampled at snow formation stage, snow cover stage, snow melt stage and growing season in the first year of decomposition. The results indicated that the content of soluble carbon and soluble organic carbon from the foliar litter of the six species showed a similar dynamical tendency at the first year of litter decomposition. Both contents of soluble carbon and soluble organic carbon increased in winter, and decreased in growing season, though the content of soluble inorganic carbon decreased in the first year of litter decomposition. The maximum contents of soluble carbon and soluble organic carbon in closed canopy were observed at snow formation stage regardless of litter species, but those in gap center and extended gap edge were observed at snow cover stage. In contrast, the contents of soluble carbon, soluble organic carbon and soluble inorganic carbon decreased significantly in growing season, even less than the initial contents. Multivariate analysis displayed that the contents of soluble carbon and soluble inorganic carbon were significantly affected by forest gap in the first year of litter decomposition, but the content of soluble organic carbon only affected by forest gap in the winter. Moreover, the contents of soluble carbon and soluble organic carbon were influenced particularly by litter quality in the winter. These results suggested that the ongoing winter warming would decrease the content of soluble carbon during foliar litter decomposition by decreasing the cover-time and thickness of snow in the winter, although the decrease degree could be controlled by litter quality.
DOIURL [本文引用: 1]
科学地估算亚洲中部天山雪岭杉(Picea schrenkiana)生态系统碳密度与碳储量是评价新疆森林碳汇潜力、评估森林在减缓大气CO2浓度上升、应对气候变化等方面功能的关键,对干旱区森林生态系统的保育和可持续发展具有重要意义.该文基于在天山雪岭杉林区布设的70个野外样地调查数据,结合新疆森林资源连续清查数据,全面估算了天山雪岭杉生态系统的碳密度和碳储量,分析了其分布格局与影响因素.结果表明:天山雪岭杉不同龄组叶、枝、干和根的含碳率变化不显著,其乔木层平均含碳率为49%,而林下植被(凋落物、草本等)平均含碳率仅为42%.雪岭杉森林生态系统单位面积生物量为187.98Mg·hm2,其中乔木层生物量占生态系统总生物量的98.93%.乔木层各组分生物量大小为:干>根>枝>叶,而各龄组生物量排序为:成熟林>中龄林>近熟林>过熟林>幼龄林.雪岭杉生态系统碳密度为544.57Mg·hrm-2,碳储量为290.84 Tg C,其中植被碳密度为92.57 Mg·hm-2,植被碳储量为53.14 Tg C,土壤碳密度为452.00Mg·hm-2,土壤碳储量为237.70 Tg C.天山雪岭杉生态系统碳密度分异与不同林区林带垂直宽度变化具有很高的相关性,其生态系统碳密度西高东低的分布格局和它所处的环境因子西优东劣的变异是相一致的,即不同的环境因素组合是造成天山雪岭杉生态系统碳密度差异的主要原因.
DOIURL [本文引用: 1]
科学地估算亚洲中部天山雪岭杉(Picea schrenkiana)生态系统碳密度与碳储量是评价新疆森林碳汇潜力、评估森林在减缓大气CO2浓度上升、应对气候变化等方面功能的关键,对干旱区森林生态系统的保育和可持续发展具有重要意义.该文基于在天山雪岭杉林区布设的70个野外样地调查数据,结合新疆森林资源连续清查数据,全面估算了天山雪岭杉生态系统的碳密度和碳储量,分析了其分布格局与影响因素.结果表明:天山雪岭杉不同龄组叶、枝、干和根的含碳率变化不显著,其乔木层平均含碳率为49%,而林下植被(凋落物、草本等)平均含碳率仅为42%.雪岭杉森林生态系统单位面积生物量为187.98Mg·hm2,其中乔木层生物量占生态系统总生物量的98.93%.乔木层各组分生物量大小为:干>根>枝>叶,而各龄组生物量排序为:成熟林>中龄林>近熟林>过熟林>幼龄林.雪岭杉生态系统碳密度为544.57Mg·hrm-2,碳储量为290.84 Tg C,其中植被碳密度为92.57 Mg·hm-2,植被碳储量为53.14 Tg C,土壤碳密度为452.00Mg·hm-2,土壤碳储量为237.70 Tg C.天山雪岭杉生态系统碳密度分异与不同林区林带垂直宽度变化具有很高的相关性,其生态系统碳密度西高东低的分布格局和它所处的环境因子西优东劣的变异是相一致的,即不同的环境因素组合是造成天山雪岭杉生态系统碳密度差异的主要原因.
DOIURL [本文引用: 1]
根据对小兴安岭凉水国家级自然保护区红松阔叶混交林的凋落物、土壤动物和土壤2a的连续采样 及测定主要营养元素N、P、K的含量,研究凋落物-土壤动物-土壤系统中主要营养元素在各分室的动态变化,并通过比较主要营养元素在不同分室中的分异,进 而分析了土壤动物在该系统营养循环中的作用。结果表明,研究区内不同凋落叶分解过程中元素含量的动态变化比较复杂,阔叶落叶中营养元素含量的变化大于针叶 落叶,但不同凋落叶在分解过程中的元素损失量之间的差异不显著。在研究时段,腐殖土层各种营养元素的含量高于土壤层;蚯蚓、蜈蚣和马陆大型土壤动物体内的 营养元素含量之间差别较大,其中蚯蚓体内全N含量最高,而马陆体内全P含量最高,蜈蚣则全K含量最高。土壤动物和土壤中营养元素含量的动态变化和凋落叶中 的变化趋势不同。N和P在土壤动物分室中表现出一定的富集,而K的富集不明显。土壤动物可以通过其新陈代谢活动加速凋落物-土壤动物-土壤系统中营养元素 的循环速率。不同凋落物分解过程中养分含量变化复杂可能是由于不同凋落物种类特性差异造成的。土壤动物在营养元素循环,尤其是N、P元素循环中具有重要意 义。
DOIURL [本文引用: 1]
根据对小兴安岭凉水国家级自然保护区红松阔叶混交林的凋落物、土壤动物和土壤2a的连续采样 及测定主要营养元素N、P、K的含量,研究凋落物-土壤动物-土壤系统中主要营养元素在各分室的动态变化,并通过比较主要营养元素在不同分室中的分异,进 而分析了土壤动物在该系统营养循环中的作用。结果表明,研究区内不同凋落叶分解过程中元素含量的动态变化比较复杂,阔叶落叶中营养元素含量的变化大于针叶 落叶,但不同凋落叶在分解过程中的元素损失量之间的差异不显著。在研究时段,腐殖土层各种营养元素的含量高于土壤层;蚯蚓、蜈蚣和马陆大型土壤动物体内的 营养元素含量之间差别较大,其中蚯蚓体内全N含量最高,而马陆体内全P含量最高,蜈蚣则全K含量最高。土壤动物和土壤中营养元素含量的动态变化和凋落叶中 的变化趋势不同。N和P在土壤动物分室中表现出一定的富集,而K的富集不明显。土壤动物可以通过其新陈代谢活动加速凋落物-土壤动物-土壤系统中营养元素 的循环速率。不同凋落物分解过程中养分含量变化复杂可能是由于不同凋落物种类特性差异造成的。土壤动物在营养元素循环,尤其是N、P元素循环中具有重要意 义。
DOIURLPMID [本文引用: 1]
Climate and initial litter quality are the major factors influencing decomposition rates on large scales. We established a comprehensive database of terrestrial leaf litter decomposition, including 785 datasets, to examine the relationship between climate and litter quality and evaluate the factors controlling decomposition on a global scale, the arid and semi-arid (AS) zone, the humid middle and humid low (HL) latitude zones. Initial litter nitrogen (N) and phosphorus (P) concentration only increased with mean annual temperature (MAT) in the AS zone and decreased with mean annual precipitation (MAP) in the HL zone. Compared with nutrient content, MAT imposed less effect on initial litter lignin content than MAP. MAT were the most important decomposition driving factors on a global scale as well as in different climatic zones. MAP only significantly affected decomposition constants in AS zone. Although litter quality parameters also showed significant influence on decomposition, their importance was less than the climatic factors. Besides, different litter quality parameters exerted significant influence on decomposition in different climatic zones. Our results emphasized that climate consistently exerted important effects on decomposition constants across different climatic zones.
DOIURL [本文引用: 2]
Freeze–thaw events in winter may affect litter decomposition in cold biomes but few reports are available. We characterized the fir (Abies faxoniana) litter decomposition over a whole winter (November 2008 to April 2009) during the late autumn, deep winter, and early spring stages. The mass loss, nutrient release, and quality change of fir litter were determined using the litterbag method at 2700, 3000, 3300, and 3600 m altitude in southwest China. Over the winter an average of 18% mass, 27% C, 50% N, 40% P, 36% K, 30% cellulose, and 14% lignin were lost. Of these total losses, a majority loss of mass (70%), C (65%), N (50%), P (58%), K (42%), cellulose (70%), and lignin (68%) occurred during the deep winter stage. The highest loss rate of mass (19.2%) and lignin (16.4%) but the lowest N loss (47.9%) was at the highest 3600 m altitude. Soil freeze–thaw cycle resulted in significant losses of mass, while mass loss rate did not increase under the higher mean soil temperature during each stage. Our results confirmed that the physical process seemed to be the most important process for cold season decomposition in the cold biome.
The freezer defrosting: Global warming and litter decomposition rates in cold biomes.
1
2006
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
Seasonal climate manipulations have only minor effects on litter decomposition rates and N dynamics but strong effects on litter P dynamics of sub-arctic bog species.
2
2012
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(
... 天山地区频繁的降雪、融化以及冻融循环作用可能对凋落叶质量产生一定的影响, 继而影响整个分解过程.本研究表明, 雪岭云杉凋落叶碳、氮、磷含量对不同雪被厚度均产生了各自的响应.在冻融期, 不同雪被厚度下凋落叶碳含量表现为上升趋势且各雪被厚度间无明显差异, 可能因为冬季初期不同雪被厚度下雪岭云杉凋落叶质量完全相同, 且环境条件基本一致, 雪被厚度对凋落叶碳含量无明显影响.而凋落叶碳含量有升高趋势说明在这一时期发生了大量的质量损失, 导致不同雪被厚度下凋落叶碳含量表现出略微增加的现象.进入深冻期, 各雪被处理下凋落叶碳含量达到最低且存在显著性差异, 这是因为土壤中的分解者随雪被厚度的增加而变得更为活跃(
Direct and indirect control by snow cover over decomposition in alpine tundra along a snowmelt gradient.
1
2010
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
Carbon Accumulation and Storage in Amazonian Ecosystems
1
2008
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
Plant Litter. Decomposition, Humus Formation, Carbon Sequestration, 2nd edn
1
2008
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
Disappearance of plant material under a winter snow cover.
1
1970
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
The control of nitrogen turn-over in forest litter.
1982
土壤微生物多样性及其影响因素研究进展
1
2015
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(
土壤微生物多样性及其影响因素研究进展
1
2015
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(
Carbon input to terrestrial ecosystems.
1
2002
... 凋落叶分解过程中氮的变化状态分为3个过程: 淋溶(释放)、固定(N吸收)、矿化(N释放)(
Microbial activity of tundra and taiga soils at sub-zero temperatures.
2
1995
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(
... 雪被下不同时期对凋落叶分解具有不同的作用机制, 因此不同凋落物分解阶段或许对雪被覆盖表现出不同的响应机制.本研究中, 在冬季3个关键时期, 厚雪被处理下凋落叶分解贡献率总是大于无雪被处理下凋落叶分解贡献率, 冬季雪被覆盖显著提高了凋落叶的分解贡献率.可能由于地表雪被为土壤动物以及微生物活性提供了隔热保温且相对稳定的环境(
Theory of metamorphism of dry snow.
1
1983
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(
How changes in soil faunal diversity and composition within a trophic group influence decomposition processes.
1
2001
... 凋落叶分解过程中氮的变化状态分为3个过程: 淋溶(释放)、固定(N吸收)、矿化(N释放)(
雪被斑块对川西高山森林凋落叶N和P释放的影响
2
2014
... 凋落叶分解过程中氮、磷元素的释放通常被认为是为分解者提供良好底物的过程, 并从营养方面对凋落物分解过程进行调控.虽然其释放特征能够根据土壤营养情况分别表现为富集和释放, 但释放出来的氮、磷元素常被看作是凋落物对于分解者的可食性指标(
... 凋落叶分解过程中氮的变化状态分为3个过程: 淋溶(释放)、固定(N吸收)、矿化(N释放)(
雪被斑块对川西高山森林凋落叶N和P释放的影响
2
2014
... 凋落叶分解过程中氮、磷元素的释放通常被认为是为分解者提供良好底物的过程, 并从营养方面对凋落物分解过程进行调控.虽然其释放特征能够根据土壤营养情况分别表现为富集和释放, 但释放出来的氮、磷元素常被看作是凋落物对于分解者的可食性指标(
... 凋落叶分解过程中氮的变化状态分为3个过程: 淋溶(释放)、固定(N吸收)、矿化(N释放)(
Lignin degradation in foliar litter of two shrub species from the gap center to the closed canopy in an alpine fir forest.
1
2016
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(
雪被斑块对高山森林两种灌木凋落叶质量损失的影响
3
2013
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(
... 天山地区频繁的降雪、融化以及冻融循环作用可能对凋落叶质量产生一定的影响, 继而影响整个分解过程.本研究表明, 雪岭云杉凋落叶碳、氮、磷含量对不同雪被厚度均产生了各自的响应.在冻融期, 不同雪被厚度下凋落叶碳含量表现为上升趋势且各雪被厚度间无明显差异, 可能因为冬季初期不同雪被厚度下雪岭云杉凋落叶质量完全相同, 且环境条件基本一致, 雪被厚度对凋落叶碳含量无明显影响.而凋落叶碳含量有升高趋势说明在这一时期发生了大量的质量损失, 导致不同雪被厚度下凋落叶碳含量表现出略微增加的现象.进入深冻期, 各雪被处理下凋落叶碳含量达到最低且存在显著性差异, 这是因为土壤中的分解者随雪被厚度的增加而变得更为活跃(
... 凋落叶分解过程中碳释放是区域内碳循环中重要的组成过程.不同时期影响凋落叶碳释放的因素不同.以往很多****认为冬季冻结情况下凋落叶分解处于“停滞”状态, 然而近年来, 已有不少研究指出冬季冻结环境中凋落叶分解仍在进行(
雪被斑块对高山森林两种灌木凋落叶质量损失的影响
3
2013
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(
... 天山地区频繁的降雪、融化以及冻融循环作用可能对凋落叶质量产生一定的影响, 继而影响整个分解过程.本研究表明, 雪岭云杉凋落叶碳、氮、磷含量对不同雪被厚度均产生了各自的响应.在冻融期, 不同雪被厚度下凋落叶碳含量表现为上升趋势且各雪被厚度间无明显差异, 可能因为冬季初期不同雪被厚度下雪岭云杉凋落叶质量完全相同, 且环境条件基本一致, 雪被厚度对凋落叶碳含量无明显影响.而凋落叶碳含量有升高趋势说明在这一时期发生了大量的质量损失, 导致不同雪被厚度下凋落叶碳含量表现出略微增加的现象.进入深冻期, 各雪被处理下凋落叶碳含量达到最低且存在显著性差异, 这是因为土壤中的分解者随雪被厚度的增加而变得更为活跃(
... 凋落叶分解过程中碳释放是区域内碳循环中重要的组成过程.不同时期影响凋落叶碳释放的因素不同.以往很多****认为冬季冻结情况下凋落叶分解处于“停滞”状态, 然而近年来, 已有不少研究指出冬季冻结环境中凋落叶分解仍在进行(
Response of decomposing litter and its microbial community to multiple forms of nitrogen enrichment.
2
2012
... 凋落叶分解过程中氮、磷元素的释放通常被认为是为分解者提供良好底物的过程, 并从营养方面对凋落物分解过程进行调控.虽然其释放特征能够根据土壤营养情况分别表现为富集和释放, 但释放出来的氮、磷元素常被看作是凋落物对于分解者的可食性指标(
... 凋落叶分解过程中氮的变化状态分为3个过程: 淋溶(释放)、固定(N吸收)、矿化(N释放)(
青藏高原东缘雪被覆盖和凋落物添加对土壤氮素动态的影响
1
2012a
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(
青藏高原东缘雪被覆盖和凋落物添加对土壤氮素动态的影响
1
2012a
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(
川西高原季节性雪被覆盖对窄叶鲜卑花凋落物分解和养分动态的影响
5
2012b
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
... ), 因此, 冬季雪被斑块下凋落物质量损失在年际分解率中可能占较大比例.当前关于季节性雪被下凋落物分解研究主要集中在青藏高原中高海拔山地森林生态系统(
... 凋落叶分解过程中氮、磷元素的释放通常被认为是为分解者提供良好底物的过程, 并从营养方面对凋落物分解过程进行调控.虽然其释放特征能够根据土壤营养情况分别表现为富集和释放, 但释放出来的氮、磷元素常被看作是凋落物对于分解者的可食性指标(
... 凋落叶分解过程中碳释放是区域内碳循环中重要的组成过程.不同时期影响凋落叶碳释放的因素不同.以往很多****认为冬季冻结情况下凋落叶分解处于“停滞”状态, 然而近年来, 已有不少研究指出冬季冻结环境中凋落叶分解仍在进行(
... 凋落叶分解过程中氮的变化状态分为3个过程: 淋溶(释放)、固定(N吸收)、矿化(N释放)(
川西高原季节性雪被覆盖对窄叶鲜卑花凋落物分解和养分动态的影响
5
2012b
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
... ), 因此, 冬季雪被斑块下凋落物质量损失在年际分解率中可能占较大比例.当前关于季节性雪被下凋落物分解研究主要集中在青藏高原中高海拔山地森林生态系统(
... 凋落叶分解过程中氮、磷元素的释放通常被认为是为分解者提供良好底物的过程, 并从营养方面对凋落物分解过程进行调控.虽然其释放特征能够根据土壤营养情况分别表现为富集和释放, 但释放出来的氮、磷元素常被看作是凋落物对于分解者的可食性指标(
... 凋落叶分解过程中碳释放是区域内碳循环中重要的组成过程.不同时期影响凋落叶碳释放的因素不同.以往很多****认为冬季冻结情况下凋落叶分解处于“停滞”状态, 然而近年来, 已有不少研究指出冬季冻结环境中凋落叶分解仍在进行(
... 凋落叶分解过程中氮的变化状态分为3个过程: 淋溶(释放)、固定(N吸收)、矿化(N释放)(
中国天然林凋落物量的空间分布及其影响因子分析
1
2016
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
中国天然林凋落物量的空间分布及其影响因子分析
1
2016
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
高山草甸冬季凋落物分解过程中土壤动物对微生物群落结构的影响
1
2016
... 凋落叶分解过程中碳释放是区域内碳循环中重要的组成过程.不同时期影响凋落叶碳释放的因素不同.以往很多****认为冬季冻结情况下凋落叶分解处于“停滞”状态, 然而近年来, 已有不少研究指出冬季冻结环境中凋落叶分解仍在进行(
高山草甸冬季凋落物分解过程中土壤动物对微生物群落结构的影响
1
2016
... 凋落叶分解过程中碳释放是区域内碳循环中重要的组成过程.不同时期影响凋落叶碳释放的因素不同.以往很多****认为冬季冻结情况下凋落叶分解处于“停滞”状态, 然而近年来, 已有不少研究指出冬季冻结环境中凋落叶分解仍在进行(
川西亚高山生态系统三种典型植物凋落物分解动态特征
3
2017
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
... ;
... 凋落叶分解过程中碳释放是区域内碳循环中重要的组成过程.不同时期影响凋落叶碳释放的因素不同.以往很多****认为冬季冻结情况下凋落叶分解处于“停滞”状态, 然而近年来, 已有不少研究指出冬季冻结环境中凋落叶分解仍在进行(
川西亚高山生态系统三种典型植物凋落物分解动态特征
3
2017
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
... ;
... 凋落叶分解过程中碳释放是区域内碳循环中重要的组成过程.不同时期影响凋落叶碳释放的因素不同.以往很多****认为冬季冻结情况下凋落叶分解处于“停滞”状态, 然而近年来, 已有不少研究指出冬季冻结环境中凋落叶分解仍在进行(
Winter climate change in plant-soil systems: Summary of recent findings and future perspectives.
1
2014
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(
Characteristics and decomposition rates of pruning residues from a shaded coffee system in Southeastern Brazil.
2
2003
... 凋落叶分解过程中碳释放是区域内碳循环中重要的组成过程.不同时期影响凋落叶碳释放的因素不同.以往很多****认为冬季冻结情况下凋落叶分解处于“停滞”状态, 然而近年来, 已有不少研究指出冬季冻结环境中凋落叶分解仍在进行(
... ).深冻期的温度相对较低, 淋溶作用比较弱, 因此土壤动物、微生物等分解者的活动是这一时期凋落叶碳释放的主要贡献者(
Energy storage and the balance of producers and decomposers in ecological systems.
3
1963
... 其中y为t时间回收凋落叶袋的残留率(%), k为分解系数, t为回收凋落叶袋时间(a) (
... 利用Olson指数衰减模型对雪岭云杉凋落叶分解过程的残留率(1-质量损失率%)进行拟合(
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(
Global-scale similarities in nitrogen release patterns during long-term decomposition.
2
2007
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
... ), 研究对象包括不同气候带和植被类型, 所涉及的环境因素有光照、大气氮沉降、土壤温湿度、土壤类型以及雪被斑块等(
Quantifying the factors affecting leaf litter decomposition across a tropical forest disturbance gradient.
1
2016
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
Chemical changes during 6 years of decomposition of 11 litters in some Canadian forest sites. Part 2. 13C abundance, solid-state 13C NMR spectroscopy and the meaning of “lignin”.
2
2009
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(
... 凋落叶分解过程中氮、磷元素的释放通常被认为是为分解者提供良好底物的过程, 并从营养方面对凋落物分解过程进行调控.虽然其释放特征能够根据土壤营养情况分别表现为富集和释放, 但释放出来的氮、磷元素常被看作是凋落物对于分解者的可食性指标(
The effects of snowpack properties and plant strategies on litter decomposition during winter in subalpine meadows.
1
2013
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
Litterfall, litter decomposition and nutrient release patterns in four native tree species raised on coal mine spoil at Singrauli, India.
1999
中国东部气候带凋落物分解特征——气候和基质质量的综合影响
1
2009
... 完全分解时间(95%分解): t0.95 = ln0.5/(-k) (
中国东部气候带凋落物分解特征——气候和基质质量的综合影响
1
2009
... 完全分解时间(95%分解): t0.95 = ln0.5/(-k) (
全球环境变化对森林凋落物分解的影响
1
2008
... 天山森林在涵养水源、保育土壤、固碳释氧以及保护生物多样性、维持绿洲稳定性等方面发挥着不可替代的作用(
全球环境变化对森林凋落物分解的影响
1
2008
... 天山森林在涵养水源、保育土壤、固碳释氧以及保护生物多样性、维持绿洲稳定性等方面发挥着不可替代的作用(
Snow removal alters soil microbial biomass and enzyme activity in a Tibetan alpine forest.
1
2014
... 雪被下不同时期对凋落叶分解具有不同的作用机制, 因此不同凋落物分解阶段或许对雪被覆盖表现出不同的响应机制.本研究中, 在冬季3个关键时期, 厚雪被处理下凋落叶分解贡献率总是大于无雪被处理下凋落叶分解贡献率, 冬季雪被覆盖显著提高了凋落叶的分解贡献率.可能由于地表雪被为土壤动物以及微生物活性提供了隔热保温且相对稳定的环境(
Characteristics of soil animal community in the subalpine/alpine forests of western Sichuan during onset of freezing.
1
2010
... 天山森林在涵养水源、保育土壤、固碳释氧以及保护生物多样性、维持绿洲稳定性等方面发挥着不可替代的作用(
Does repeated freezing and thawing accelerate decay of leaf litter?
1
1988
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
The snow-bed vegetation in the Northern Apennines.
1
1991
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(
Microbial activity and litter decomposition under snow cover in a cool-temperate broad-leaved deciduous forest.
2
2005
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(
... ;
Litter decomposition on the Mauna Loa environmental matrix, Hawaii: Patterns, mechanisms, and models.
1
1994
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
Litter decomposition in two subalpine forests during the freeze-thaw season.
2
2010
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
... 凋落叶分解过程中氮的变化状态分为3个过程: 淋溶(释放)、固定(N吸收)、矿化(N释放)(
季节性雪被对高山森林凋落物分解的影响
4
2013
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
... ;
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(
... ), 厚雪被具有隔热保温作用, 所提供的稳定环境更益于雪岭云杉凋落叶分解(
季节性雪被对高山森林凋落物分解的影响
4
2013
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
... ;
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(
... ), 厚雪被具有隔热保温作用, 所提供的稳定环境更益于雪岭云杉凋落叶分解(
冬季雪被对青藏高原东缘高海拔森林凋落叶P元素释放的影响
1
2015
... 凋落叶分解过程中氮、磷元素的释放通常被认为是为分解者提供良好底物的过程, 并从营养方面对凋落物分解过程进行调控.虽然其释放特征能够根据土壤营养情况分别表现为富集和释放, 但释放出来的氮、磷元素常被看作是凋落物对于分解者的可食性指标(
冬季雪被对青藏高原东缘高海拔森林凋落叶P元素释放的影响
1
2015
... 凋落叶分解过程中氮、磷元素的释放通常被认为是为分解者提供良好底物的过程, 并从营养方面对凋落物分解过程进行调控.虽然其释放特征能够根据土壤营养情况分别表现为富集和释放, 但释放出来的氮、磷元素常被看作是凋落物对于分解者的可食性指标(
季节性冻融期间土壤动物对岷江冷杉凋落叶质量损失的贡献
1
2011
... 雪被下不同时期对凋落叶分解具有不同的作用机制, 因此不同凋落物分解阶段或许对雪被覆盖表现出不同的响应机制.本研究中, 在冬季3个关键时期, 厚雪被处理下凋落叶分解贡献率总是大于无雪被处理下凋落叶分解贡献率, 冬季雪被覆盖显著提高了凋落叶的分解贡献率.可能由于地表雪被为土壤动物以及微生物活性提供了隔热保温且相对稳定的环境(
季节性冻融期间土壤动物对岷江冷杉凋落叶质量损失的贡献
1
2011
... 雪被下不同时期对凋落叶分解具有不同的作用机制, 因此不同凋落物分解阶段或许对雪被覆盖表现出不同的响应机制.本研究中, 在冬季3个关键时期, 厚雪被处理下凋落叶分解贡献率总是大于无雪被处理下凋落叶分解贡献率, 冬季雪被覆盖显著提高了凋落叶的分解贡献率.可能由于地表雪被为土壤动物以及微生物活性提供了隔热保温且相对稳定的环境(
长江上游高山森林林窗对凋落物分解过程中可溶性碳的影响
1
2015
... 雪被下不同时期对凋落叶分解具有不同的作用机制, 因此不同凋落物分解阶段或许对雪被覆盖表现出不同的响应机制.本研究中, 在冬季3个关键时期, 厚雪被处理下凋落叶分解贡献率总是大于无雪被处理下凋落叶分解贡献率, 冬季雪被覆盖显著提高了凋落叶的分解贡献率.可能由于地表雪被为土壤动物以及微生物活性提供了隔热保温且相对稳定的环境(
长江上游高山森林林窗对凋落物分解过程中可溶性碳的影响
1
2015
... 雪被下不同时期对凋落叶分解具有不同的作用机制, 因此不同凋落物分解阶段或许对雪被覆盖表现出不同的响应机制.本研究中, 在冬季3个关键时期, 厚雪被处理下凋落叶分解贡献率总是大于无雪被处理下凋落叶分解贡献率, 冬季雪被覆盖显著提高了凋落叶的分解贡献率.可能由于地表雪被为土壤动物以及微生物活性提供了隔热保温且相对稳定的环境(
天山森林生态系统碳储量格局及其影响因素
1
2016
... 天山森林在涵养水源、保育土壤、固碳释氧以及保护生物多样性、维持绿洲稳定性等方面发挥着不可替代的作用(
天山森林生态系统碳储量格局及其影响因素
1
2016
... 天山森林在涵养水源、保育土壤、固碳释氧以及保护生物多样性、维持绿洲稳定性等方面发挥着不可替代的作用(
红松阔叶混交林凋落物-土壤动物-土壤系统中N、P、K的动态特征
1
2007
... 凋落叶分解过程中碳释放是区域内碳循环中重要的组成过程.不同时期影响凋落叶碳释放的因素不同.以往很多****认为冬季冻结情况下凋落叶分解处于“停滞”状态, 然而近年来, 已有不少研究指出冬季冻结环境中凋落叶分解仍在进行(
红松阔叶混交林凋落物-土壤动物-土壤系统中N、P、K的动态特征
1
2007
... 凋落叶分解过程中碳释放是区域内碳循环中重要的组成过程.不同时期影响凋落叶碳释放的因素不同.以往很多****认为冬季冻结情况下凋落叶分解处于“停滞”状态, 然而近年来, 已有不少研究指出冬季冻结环境中凋落叶分解仍在进行(
Control of climate and litter quality on leaf litter decomposition in different climatic zones.
1
2015
... 凋落物分解阶段是森林生态系统物质循环和能量流动的关键阶段, 对森林生态系统的碳循环和氮周转具有决定性作用(
Decomposition of Abies faxoniana litter varies with freeze-thaw stages and altitudes in subalpine/alpine forests of southwest China.
2
2012
... 其中G为质量损失率(%), M0为凋落叶初始质量(g), Mi为第i次回收凋落叶袋时凋落叶烘干质量(g), MI为最后一次回收凋落叶袋时凋落叶烘干质量(g) (
... 以冬季变暖和极端气候事件等为主要特征的全球气候变化可能显著影响高纬度地区森林凋落叶分解等关键生态学过程(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}