Water-use process of two desert shrubs along a precipitation gradient in Horqin Sandy Land
CHEN Ding-Shuai2,3,*, DONG Zheng-Wu1,*, GAO Lei2, CHEN Xiao-Min3, PENG Xin-Hua2, SI Bing-Cheng4, ZHAO Ying1,4,**, 1Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, ürümqi 830011, China;2State Key Laboratory of Soil and Sustainable Agriculture, Nanjing Institute of Soil Science, Chinese Academy of Sciences, Nanjing 210008, China3College of Resources and Environmental Sciences, Nanjing Agricultural University, Nanjing 210095, Chinaand 4Institute of Water Saving Agriculture in Arid Areas of China, Northwest A & F University, Yangling, Shaanxi 712100, China; 通讯作者:** 通信作者Author for correspondence (E-mail: yzhaosoils@gmail.com) 责任编辑: 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家重点基础研究发展计划(973计划)(2013CB429902)、国家“****”新疆项目(Y472241001)和国家自然科学基金(41630860和41371234)
关键词:小叶锦鸡儿;盐蒿;水分来源;模拟降水 Abstract Aims The determination of the source of plant water is an important research on the plant-water relationship in arid and semiarid regions and helps to understand the adaptation strategy of desert species to the dry environment. Plant water use pattern affects plant community composition and ecosystem water budget. This study aims to investigate the water use patterns of Caragana microphylla and Artemisia halodendron, two typical shrub species, under altered rainfall conditions in Horqin Sandy Land. Methods Water treatments include ambient rainfall (natural rainfall), 50% increase in rainfall (enhanced rainfall) and 50% decrease in rainfall (reduced rainfall) by artificially intercepting and redistributing natural rainfall. Stable hydrogen and oxygen isotope ratios (δD and δ18O) were measured for xylem water, rainfall, and soil water in different soil layers (0-120 cm depth). The possible ranges of potential water sources used by C. microphylla and A. halodendron were calculated using the IsoSource model. Important findings 1) Alteration of ambient rainfall mainly affected the soil water condition in the shallow soil (0-30 cm). Increase in rainfall significantly increased the above- and below-ground biomass, and δ18O values of soil water declined with soil depth. 2) Under the enhanced rainfall treatment, A. halodendron mainly used the soil water in the shallow soil (0-40 cm) and C. microphylla was able to extract water from multiple soil layers. Under the reduced rainfall treatment, both species increasingly relied on extracting water from deeper soil layers, 60-80 cm for A. halodendron and 60-120 cm for C. microphylla. 3) For the natural rainfall treatment, in the wet season, the upper soil water was recharged by rainfall, C. microphylla and A. halodendron extract the shallow soil water (0-60 cm). However, in the dry season, soil water content was dramatically reduced, and main water sources for C. microphylla shifted from topsoil to deeper soil, and A. halodendron can use multiple layers of soil water. In summary, A. halodendron is more capable of exploring deeper soil moisture under reduced rainfall in comparison with C. microphylla, and is likely to be more adaptive to this water-limiting desert environment. Keywords:Caragana microphylla; Artemisia halodendron; water sources;simulated rainfall -->0 PDF (1149KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 陈定帅, 董正武, 高磊, 陈效民, 彭新华, 司炳成, 赵英. 不同降水条件下科尔沁沙地小叶锦鸡儿和盐蒿的水分利用动态. 植物生态学报, 2017, 41(12): 1262-1272 https://doi.org/10.17521/cjpe.2017.0219 在干旱半干旱区, 水是限制植物定居、生存和生长的关键因子(Duan et al., 2008; 周海等, 2013; 刘凯等, 2015)。降水是沙地生态系统的主要水分来源, 年降水格局在塑造植物适应性方面起着重要作用(Schwinning & Ehleringer, 2001), 制约着沙地植被的形成和发展(Knapp et al., 2001), 也影响着土壤-植被系统的演变方向和生态功能(李小雁, 2012)。由于沙漠地区降水少, 降水时间不确定, 加上蒸发量大, 表层土壤水分含量波动剧烈。为了维持生长, 植物必须依赖较稳定的水源, 如深层土壤水或地下水(张凯等, 2011; 郑肖然等, 2015)。已有研究发现, 在干旱半干旱区, 植物主要吸收降水补充的浅层土壤水或地下水补充的深层土壤水(Dodd et al., 1998)。在不同的降水条件下, 植物的水分利用效率也不同, 植物可能同时利用多个水源, 或者季节性的转换水源(周雅聃等, 2011)。近年来随着全球气候变化加剧, 极端降水事件及降水量减少表现出波动上升的趋势, 大间隔单次降水的增加势必会导致干旱半干旱地区植物水分利用格局发生变化。因此, 研究降水梯度下区域植物水分利用策略极有必要。其中, 利用增减雨处理, 以“空间代替时间”的方法, 是旱地模拟植物响应未来气候变化的较好手段(赵文智等, 2002; 程徐冰等, 2011)。如果能同时利用水同位素技术, 则可能明晰旱区植物水分利用机制。 氢氧稳定同位素技术是定量研究植物水分利用来源、水分利用格局的较好方法。利用该技术手段, 国内****对干旱半干旱区植物水分来源及不同水源对植物的贡献做了相关研究(赵良菊等, 2008; 余绍文等, 2012; 李亚飞等, 2017; 朱雅娟等, 2017)。小叶锦鸡儿(Caragana microphylla)属豆科锦鸡儿属灌木, 盐蒿(Artemisia halodendron)属菊科蒿属灌木或半灌木。这两种植物均为内蒙古草原的优势植物种, 主要分布于锡林河流域及科尔沁沙地, 在防风固沙、保持水土等方面起着极其重要的作用(曹成有等, 2004; 郑肖然等, 2015)。近年来, 小叶锦鸡儿和盐蒿群落在科尔沁沙地出现退化现象, 而降水和地下水的变化可能与两种植物退化存在密切关系。因此, 了解降水增减过程中小叶锦鸡儿和盐蒿对降水变化的响应机制对制定有效的植被管理对策具有重要意义。 科尔沁沙地位于内蒙古东部西辽河中下游, 是我国最大的沙地, 也是我国沙漠化最为严重的一个地区。在全球气候变化的影响下, 该地区年降水量呈减少趋势, 气候趋于干旱化(张腊梅等, 2014)。作为科尔沁沙地的主要优势种, 小叶锦鸡儿和盐蒿的水分利用对当地生态系统的水分平衡和植物群落组成产生了重要的影响(熊小刚等, 2003; Li et al., 2013)。本研究应用稳定性同位素技术分析模拟降水条件下小叶锦鸡儿和盐蒿水分利用动态以及变化特征, 研究结果对认识干旱半干旱区气候变化下植物的适应性, 以及对植被保护、恢复与重建具有参考价值, 为合理安排当地种植固沙植株的密度及空间分布提供指导。
1 材料和方法
1.1 研究区概况
研究区位于科尔沁沙地中南部内蒙古自治区通辽市奈曼旗境内(120.32°-121.52° E, 42.23°-43.53° N, 平均海拔约为360 m)。该区域属温带半干旱大陆性季风气候, 年平均气温6.5 ℃, 最热月(7月)平均气温为23.5 ℃, 全年≥10 ℃的有效积温在3 000 ℃以上, 无霜期约150天, 年降水量364.6 mm, 主要集中在6-8月, 年蒸发量1β972.8 mm, 年平均风速3.5 m·s-1。地貌类型以沙丘和面积不等的平缓沙地及甸子地交错分布为特征, 土壤多为风沙土或沙质草甸土。研究区天然植物群落以中旱生植物为主, 其主要植物种有盐蒿、小叶锦鸡儿、糙隐子草(Cleistogenes squarrosa)、砂蓝刺头(Echinops gmelini)、猪毛菜(Salsola collina)、长穗虫实(Corispermum elongatum)和冷蒿(Artemisia frigida)等。
1.2 样地布设
试验场地于2014年6月建于中国科学院奈曼沙漠化研究站周围围封的平缓固定沙地, 在样区内分别选择当地天然生长的生长良好、形态相近的10年生小叶锦鸡儿和盐蒿植株, 分别在两种植物生长较为均匀的样地布设增减雨装置各3个处理, 每个增减雨处理下有植株4-6株(其中增雨和减雨处理下各2-3株)。根据当地降水分布特征和增减雨装置的特性, 将增减雨量设计为降水单因素的重复对比实验, 即增雨、减雨和对照。以正常降水为对照, 增雨处理比对照增加降水50%, 减雨处理比对照减少降水50%。增减雨设置的样地为2 m × 4 m (增雨和减雨样地分别为2 m × 2 m)(张腊梅等, 2014), 在增减雨装置交界处, 用防水布将增雨和减雨样地完全隔离, 并将增减雨样地交界处地下1 m深土体用防水布包裹。每套装置周围留出5 m的缓冲带, 以避免相互干扰。对增减雨装置边缘的地下部分1 m深土体用防水布包裹, 防水布高出地表10 cm, 防止其土壤水分受相邻样地地表径流和侧向水分的影响。本研究中所采用的增减雨装置是由支撑架截雨槽、集雨箱和增雨管组成(张腊梅等, 2014)。截雨槽由透明有机玻璃板热压成直角角槽, 对光照的影响小。减雨功能通过截雨槽将自然降水的50%截留, 达到减雨样地降低50%的降水量, 同时这部分所截留的降水流进集雨箱内, 然后通过增雨管将截留的降水均匀引入前方增水样方内, 实现同步增雨50%的目的(张腊梅等, 2014)。
1.3 样品采集与分析
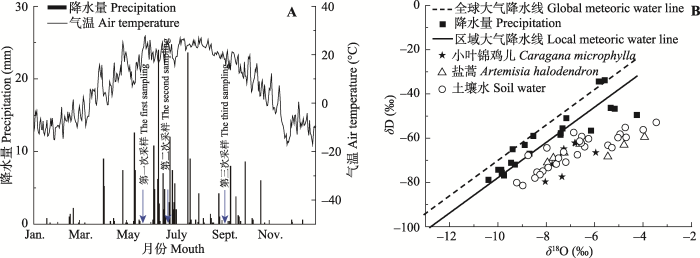
在样区分别选取4株生长良好、形态相近、具有代表性的小叶锦鸡儿、盐蒿植株。于2015年5月17日(生长前期)、 6月23日(生长中期)和9月2日(生长后期)分别采集增雨、减雨和对照处理的两种植物木质部样品, 每株每次取1个样, 共4个重复。采样方法: 剪取非绿色的栓化小枝(直径0.1-0.3 cm, 长3-5 cm), 去皮后迅速装入采样瓶并用封口膜密封, 然后放入随身携带的冰盒中, 带回实验室冷冻保存。在每株植物的下方用土钻取0-120 cm土层的土壤样品, 取样时间同植物的取样时间一致。每20 cm作为一个土层取土样(盐蒿由于根系长度关系土样只采到80 cm), 每层所取土样迅速装入样品瓶, 用封口膜密封后放入携带的冰盒, 带回实验室冷冻保存, 用于分析土壤水的稳定氢同位素比率(δD)、稳定氧同位素比率(δ18O)值; 同时采用时域反射仪(TDR, IMKO, Ettlingen, Germany)测定植株附近土壤含水量。降水样品来自奈曼站气象观测站, 每次降水后收集雨水, 密封于专用玻璃瓶, 然后用封口膜密封, 放置在-2 ℃冰箱冷藏以备分析使用。2015年降水总量为230.2 mm, 试验期(5-9月)降水量为183.4 mm, 占全年降水量的80% (图1)。降水主要集中在5-7月(155.2 mm), 而8-9月降水总量仅为28.2 mm, 样区地下水埋深为7.5 m。采用低温真空抽提法提取土壤和植物样品中的水分, 将抽提的植物小枝木质部水(简称木质部水)和土壤水分别装入小玻璃瓶, 土壤水样品采用LGR液态水同位素分析仪(908-0008, Los Gatos Research, Mountain View, USA )测量, 为避免植物水样品中醇及其他挥发性有机物干扰同位素, 植物水样品采用质谱仪(Finnigan MAT-253, Thermo finnigan, Bremen, Germany)测定。 显示原图|下载原图ZIP|生成PPT 图1研究区降水和气温(2015) (A)及降水、土壤水和植物木质部水稳定氢、氧同位素比率(δD、δ18O) (B)。 -->Fig. 1The precipitation and air temperature (2015) (A), and the stable hydrogen and oxygen isotope ratios (δD and δ18O) of precipitation, soil water and plant water (B) in the study area. -->
利用SPSS 17.0统计软件进行单因素方差分析, 多重比较采用最小显著差异法。用Origin 8.0软件进行绘图。 显示原图|下载原图ZIP|生成PPT 图2不同降水处理下小叶锦鸡儿(A、B、C)和盐蒿(D、E、F)的土壤含水量(平均值±标准误差, n = 4)。 -->Fig. 2Soli water content under Caragana microphylla (A, B, C) or Artemisia halodendron (D, E, F) as affected by enhanced or reduced rainfall (mean ± SE, n = 4). -->
对降水中δD、δ18O值进行分析, 通过回归分析得出该区域大气降水线方程(LMWL, δDLMWL = 6.36δ18OLMWL - 10.126, p < 0.001), 与全球大气降水线方程(GMWL, δDGMWL = 8δ18O + 10)相比, 斜率和截距明显偏小, 且小叶锦鸡儿木质部和盐蒿木质部水、雨水和土壤水氢氧稳定同位素值都位于全球大气降水线右侧, 表明在该区域的降水过程中氢氧稳定同位素受到了二次蒸发的影响(图1)。 小叶锦鸡儿木质部水和土壤水δ18O值的变化趋势见图5和图6。5月17日增雨与对照处理下土壤水δ18O值差异不显著(p > 0.05), 减雨处理下δ18O值在0-20 cm土层高于增雨和对照处理, 也高于其他土层; 小叶锦鸡儿木质部水δ18O值表现为: 增雨(-7.18‰) <对照(-6.95‰) <减雨(-4.87‰); 6月23日由于降水增多和气温升高, 蒸散作用加剧, 不同深度土壤水δ18O在各处理之间差异显著(p < 0.05): 减雨处理>增雨处理>对照处理, 但各处理下小叶锦鸡儿木质部水的δ18O值差异不明显(-6.39‰至-5.84‰);经过了夏天干旱高温, 9月2日各处理下浅层土壤δ18O值富集明显, 甚至达到正值, 而在60 cm以下土层δ18O值保持相对稳定。 显示原图|下载原图ZIP|生成PPT 图3不同降水处理下小叶锦鸡儿(A)和盐蒿(B)的地上和地下生物量(平均值±标准误差, n = 4)。不同小写字母表示处理间差异显著(p < 0.05)。 -->Fig. 3The aboveground and underground biomass of Caragana microphylla (A) and Artemisia halodendron (B) as affected by enhanced or reduced precipitation (mean ± SE, n = 4). Different lowercase letters indicate significant differences among treatments (p < 0.05). -->
显示原图|下载原图ZIP|生成PPT 图4小叶锦鸡儿(A、B)和盐蒿(C、D)吸收根与输导根的垂直分布特征(平均值±标准误差, n = 4)。 -->Fig. 4The distribution of absorptive-conductive root biomass for Caragana microphylla (A, B) and Artemisia halodendron (C, D) as affected by enhanced or reduced precipitation (mean ± SE, n = 4). -->
显示原图|下载原图ZIP|生成PPT 图5不同降水处理下土壤水氧稳定同位素(δ18O)值(平均值±标准误差, n = 4)。A、B、C为不同月份小叶锦鸡儿土壤水δ18O值, D、E为不同月份盐蒿土壤水δ18O值。 -->Fig. 5Oxygen stable isotope ratio (δ18O) of soil water as affected by enhanced or reduced precipitation (mean ± SE, n = 4). A, B and C show the δ18O values of Caragana microphylla in different months, D and E show the δ18O values of Artemisia halodendron in different months. -->
显示原图|下载原图ZIP|生成PPT 图6不同降水处理下小叶锦鸡儿(A)和盐蒿(B)木质部水氧稳定同位素(δ18O)值(平均值±标准误差, n = 4)。 -->Fig. 6The xylem water δ18O values of Caragana microphylla (A) and Artemisia halodendron (B) as affected by enhanced or reduced precipitation (mean ± SE, n = 4). -->
BarnesCJ, AllisonGB (1998). Tracing of water movement in the unsaturated zone using stable isotopes of hydrogen and oxygen .Journal of Hydrology, 100, 143-176.https://doi.org/10.1016/0022-1694(88)90184-9URL [本文引用: 1]摘要 The development of an analytical model for movement of the stable isotopic species of water in unsaturated soils is presented by means of a review of recent literature on the subject. The model adequately represents experimental observations of isotope profiles during evaporation from saturated or unsaturated soils, under both nonisothermal and nonsteady conditions. Interpretation of field isotope profiles using the model is discussed, and indications are made of areas where further work is desirable.
[2]
CaoCY, JiangDM, LuoYM, KouZW (2004). Stability ofCaragana microphylla plantation for wind protection and sand fixation . Acta Ecologica Sinica, 24, 1178-1186.(in Chinese with English abstract) [曹成有, 蒋德明, 骆永明, 寇振武 (2004). 小叶锦鸡儿防风固沙林稳定性研究 . 生态学报, 24, 1178-1186.]https://doi.org/10.3321/j.issn:1000-0933.2004.06.013URL [本文引用: 1]摘要 针对科尔沁沙地小叶锦鸡儿固沙林在持续高温干旱条件下出现稳定性波动的问题,选择2、5、15、35a人 工群落和天然群落进行对比研究,探讨了人工小叶锦鸡儿防风固沙林物种多样性在35a间的变化以及对大气持续干旱的抵抗力和对外界干扰的恢复能力.流动沙丘 固定后,固沙林内草本植物迅速侵入,其后则缓慢发展并处于相对稳定状态,植物种类数保持在20~23种.但在人工固沙群落中1年生植物在种类数和数量上始 终处于优势地位,而天然群落则是多年生植物占优势.人工固沙群落物种多样性指数逐年提高,但明显低于天然群落.在受到大气持续高温干旱的干扰时,人工群落 土壤条件极度恶化,出现了明显的干沙层,最深达140cm,叶片含水量极度降低,植物生长停止.沙地植物群落对牲畜取食均具有一定的补偿的能力,在啃食较 轻时表现为超补偿现象.通过建立数学模型分析,人工群落个体的补偿点和超补偿点明显低于天然群落.采用冗余理论对植物群落稳定性机制进行了论述,认为冗余 结构的变化是导致群落稳定性变化的内在原因.对现有人工小叶锦鸡儿固沙林的稳定性进行了评价,并认为15a左右人工固沙林正处于相对稳定并缓慢发展状态. 对于稳定的人工固沙群落应具备以下条件:(1)适于当地环境条件;(2)对外界干扰具有一定的抵抗性和恢复性;(3)具有正常生长发育和自我更新能力; (4)有较为完善的种群结构.
[3]
ChengXB, WuJ, HanSJ, ZhouYM, WangXX (2011). Effects of decreased rainfall on Quercus mongolica leaf eco- physiological characteristics. Chinese Journal of Ecology, 30, 1908-1914.(in Chinese with English abstract) [程徐冰, 吴军, 韩士杰, 周玉梅, 王秀秀 (2011). 减少降水对长白山蒙古栎叶片生理生态特性的影响 . 生态学杂志, 30, 1908-1914.] [本文引用: 1]
[4]
ChimnerRA, CooperDJ (2004). Using stable oxygen isotopes to quantify the water source used for transpiration by native shrubs in the San Luis Valley, Colorado USA .Plant and Soil, 260, 225-236.https://doi.org/10.1023/B:PLSO.0000030190.70085.e9URL [本文引用: 1]
[5]
DaiY, ZhengXJ, TangLS, LiY (2014). Dynamics of water usage in Haloxylon ammodendron in the southern edge of the Gurbantünggüt Desert. Chinese Journal of Plant Ecology, 38, 1214-1225.(in Chinese with English abstract) [戴岳, 郑新军, 唐立松, 李彦 (2014). 古尔班通古特沙漠南缘梭梭水分利用动态 . 植物生态学报, 38, 1214-1225.] [本文引用: 1]
[6]
DawsonTE, MambelliS, PlamboeckAH (2002). Stable isotopes in plant ecology .Annual Review of Ecology and Systematics, 33, 507-559.https://doi.org/10.1146/annurev.ecolsys.33.020602.095451URL [本文引用: 1]摘要 The use of stable isotope techniques in plant ecological research has grown steadily during the past two decades. This trend will continue as investigators realize that stable isotopes can serve as valuable nonradioactive tracers and nondestructive integrators of how plants today and in the past have interacted with and responded to their abiotic and biotic environments. At the center of nearly all plant ecological research which has made use of stable isotope methods are the notions of interactions and the resources that mediate or influence them. Our review, therefore, highlights recent advances in plant ecology that have embraced these notions, particularly at different spatial and temporal scales. Specifically, we review how isotope measurements associated with the critical plant resources carbon, water, and nitrogen have helped deepen our understanding of plant-resource acquisition, plant interactions with other organisms, and the role of plants in ecosystem studies. Where possible we also introduce how stable isotope information has provided insights into plant ecological research being done in a paleontological context. Progress in our understanding of plants in natural environments has shown that the future of plant ecological research will continue to see some of its greatest advances when stable isotope methods are applied.
[7]
DoddMB, LauenrothWK, WelkerJM (1998). Differential water resource use by herbaceous and woody plant life forms in a shortgrass steppe community .Oecologia, 117, 504-512.https://doi.org/10.1007/s004420050686URLPMID:28307675 [本文引用: 1]摘要 We conducted a study to test the predictions of Walter's two-layer model in the shortgrass steppe of northeastern Colorado. The model suggests that grasses and woody plants use water resources from different layers of the soil profile. Four plant removal treatments were applied in the spring of 1996 within a plant community codominated by Atriplex canescens (a C 4 shrub) and Bouteloua gracilis (a C 4 grass). During the subsequent growing season, soil water content was monitored to a depth of 18065cm. In addition, stem and leaf tissue of Atriplex , Bouteloua and the streamside tree Populus sargentii were collected monthly during the growing seasons of 1995 and 1996 for analysis of the δ 18 O value of plant stem water (for comparison with potential water sources) and the δ 13 C value of leaves (as an indicator of plant water status). Selective removal of shrubs did not significantly increase water storage at any depth in the measured soil profile. Selective removal of the herbaceous understory (mainly grasses) increased water storage in the top 6065cm of the soil. Some of this water gradually percolated to lower layers, where it was utilized by the shrubs. Based on stem water δ 18 O values, grasses were exclusively using spring and summer rain extracted from the uppermost soil layers. In contrast, trees were exclusively using groundwater, and the consistent δ 13 C values of tree leaves over the course of the summer indicated no seasonal changes in gas exchange and therefore minimal water stress in this life-form. Based on anecdotal rooting-depth information and initial measurements of stem water δ 18 O, shrubs may have also had access to groundwater. However, their overall δ 18 O values indicated that they mainly used water from spring and summer precipitation events, extracted from subsurface soil layers. These findings indicate that the diversity of life-forms found in this shortgrass steppe community may be a function of the spatial partitioning of soil water resources, and their differential use by grasses, shrubs, and trees. Consequently, our findings support the two-layer model in a broad sense, but indicate a relatively flexible strategy of water acquisition by shrubs.
[8]
DuXL, WangSJ (2011). Recent advances of stable hydrogen and oxygen isotopic techniques in plant water use strategy .Chinese Agricultural Science Bulletin, 27(22), 5-10.(in Chinese with English abstract) [杜雪莲, 王世杰 (2011). 稳定性氢氧同位素在植物用水策略中的研究进展 . 中国农学通报,27(22), 5-10.]
[9]
DuanDY, OuyangH, SongMH (2008). Water sources of dominant species in three alpine ecosystems on the Tibetan Plateau, China .Journal of Integrative Plant Biology, 50, 257-264.https://doi.org/10.1111/jipb.2008.50.issue-3URL [本文引用: 2]
EhleringerJR, DawsonTE (1992). Water uptake by plants: Perspectives from stable isotope composition .Plant, Cell & Environment, 15, 1073-1082.https://doi.org/10.1111/j.1365-3040.1992.tb01657.xURL [本文引用: 2]摘要 ABSTRACT Stable isotope studies of hydrogen and oxygen stable isotope ratios of water within plants are providing new information on water sources, competitive interactions and water use patterns under natural conditions. Variation in the utilization of summer rain by aridland species and limited use of stream water by mature riparian trees are two examples of how stable isotope studies have modified our understanding of plant water relations. Analyses of xylem sap and tree rings have the potential of providing both short-term and long-term information on plant water use patterns.
[12]
FlanaganLB, EhleringerJR (1992). Differential uptake of summer precipitation among co-occurring trees and shrubs in a pinyon-juniper woodland .Plant, Cell & Environment, 15, 831-836.URL [本文引用: 2]
[13]
GroismanPY, KarlTR, EasterlingDR (1999). Changes in the probability of heavy precipitation: Important indicators of climatic change . Climatic Change, 42, 243-283.https://doi.org/10.1023/A:1005432803188URL [本文引用: 2]
[14]
KnappAK, BriggsJM, KoellikerJK (2001). Frequency and extent of water limitation to primary production in a mesic temperate grassland .Ecosystems, 4, 19-28.https://doi.org/10.1007/s100210000057URL [本文引用: 1]
[15]
LiKF, LuoYY, ZhangHL, SheHY (2012). The relations between root distribution of Artemisia halodendron and soil water in Horqin . Journal of Arid Land Resources and Environment, 26(8), 167-171.(in Chinese with English abstract) [李凯锋, 罗于洋, 张海龙, 折红燕 (2012). 科尔沁差巴嘎蒿根系分布规律与土壤水分关系的研究 . 干旱区资源与环境,26(8), 167-171.] [本文引用: 1]
[16]
LiXY (2012). Coupling, respond and adaptation of soil-plant water in arid areas in arid area .Science China: Earth Science, 41, 1721-1730.(in Chinese) [李小雁 (2012). 干旱地区土壤-植被-水文耦合, 响应与适应机制 . 中国科学: 地球科学, 41, 1721-1730.] [本文引用: 1]
[17]
LiXY, ZhangSY, PengHY (2013). Soil water and temperature dynamics in shrub-encroached grasslands and climatic implications: Results from Inner Mongolia steppe ecosystem of north China . Agricultural and Forest Meteorology, 171-172, 20-30. [本文引用: 1]
[18]
LiYF, YuJJ, LuK, WangP, ZhangYC (2017). Water sources of Populus euphratica and Tamarix ramosissima in Ejina Delta, the lower reaches of the Heihe River, China. Chinese Journal of Plant Ecology, 41, 519-528.(in Chinese with English abstract) [李亚飞, 于静洁, 陆凯, 王平, 张一驰 (2017). 额济纳三角洲胡杨和多枝柽柳水分来源解析 . 植物生态学报, 41, 519-528.]https://doi.org/10.17521/cjpe.2016.0381URL [本文引用: 1]摘要 为研究黑河下游额济纳三角洲典型河岸带植物胡杨(Populus euphratica)和多枝柽柳(Tamarix ramosissima)的水分来源及其时空变化特征,于2015–2016年植物生长期在额济纳东、西河沿岸选取8处样点,分别采集胡杨和多枝柽柳木质部、土壤以及降水、河水、地下水样品,分析测试获取各样品的氧稳定同位素比率(δ18O),并利用氧同位素直接对比法确定植物主要吸水层位,利用Iso Source线性混合模型确定胡杨和多枝柽柳水分来源构成及比例。研究表明:河水和地下水为胡杨和多枝柽柳的主要补给水源,降水补给因雨水少且入渗浅可忽略不计;胡杨和多枝柽柳吸收土壤水的层位因地下水位波动、土壤物理特性、河水对土壤水的侧向补给及漫溢补给等存在较大的空间差异,但其利用河水和地下水的比例未见明显空间差异;胡杨更多地利用河水(68%),而多枝柽柳更多地利用地下水(65%);植物水分来源对生态输水工程响应敏感,河水贡献率在输水期增大至84%和48%,非输水期下降至63%和30%,地下水贡献率在输水期下降至16%和52%,非输水期增大至37%和70%。值得指出的是,河岸带地下水与河水交互作用强烈,导致地下水与河水的δ18O较为接近,使得源分解析定量具有不确定性。
[19]
LinGH, PhillipsSL, EhleringerJR (1996). Monsoonal precipitation responses of shrubs in a cold desert community on the Colorado Plateau .Oecologia, 106, 8-17.https://doi.org/10.1007/BF00334402URL [本文引用: 1]
[20]
LiuK, GaoL, PengXH, ZhangZB (2015). Spatio-temporal variability of soil moisture in Horqin sandy land .Soils, 47, 765-772.(in Chinese with English abstract) [刘凯, 高磊, 彭新华, 张中彬 (2015). 半干旱区科尔沁沙地土壤水分时空特征研究 . 土壤, 47, 765-772.] [本文引用: 2]
[21]
LiuSG, PiaoSJ, AnMZ, LiuF (2003). Distribution dynamics of Artemisia halodendron absorbent roots in different kinds of sandy land . Acta Phytoecologica Sinica ,27, 684-689.(in Chinese with English abstract) [刘士刚, 朴顺姬, 安卯柱, 刘芳 (2003). 不同类型沙地上差不嘎蒿细根的分布状态 . 植物生态学报, 27, 684-689.] [本文引用: 1]
[22]
MaCC, GaoYB, JiangFQ, WangJL, GuoHY, WuJB, SuD (2004). The comparison studies of ecological and water regulation characteristics of Caragana microphylla and Caragana stenophylla . Acta Ecologica Sinica, 24, 1442-1451.(in Chinese with English abstract) [马成仓, 高玉葆, 蒋福全, 王金龙, 郭宏宇, 吴建波, 苏丹 (2004). 小叶锦鸡儿和狭叶锦鸡儿的生态和水分调节特性比较研究 . 生态学报, 24, 1442-1451.]https://doi.org/10.3321/j.issn:1000-0933.2004.07.019URL [本文引用: 1]摘要 对锦鸡儿属羽状叶类群的代表植物——小叶锦鸡儿和假掌状叶类群的代表植物——狭叶锦鸡儿的地理分布、生长发育、形态结构、渗透调节和水分代谢进行了比较研究。小叶锦鸡儿在内蒙古高原分布于东起呼伦贝尔 (1 2 4 .0 0°E)西至达茂旗(1 1 0 .4 3°E)的半湿润至半干旱地区 ,生长发育最适地区是锡林浩特 ;狭叶锦鸡儿在内蒙古高原分布于东起锡林浩特 (1 1 6 .0 7°E)西至阿拉善 (98°E)的半干旱至极干旱地区 ,生长发育最适地区是阿拉善。这种分布格局和生长发育适应性说明 ,狭叶锦鸡儿比小叶锦鸡儿更适宜干旱环境。小叶锦鸡儿叶片平展、被绿色柔毛 ,有时上面近无毛 ,狭叶锦鸡儿叶片呈瓦状、被粉白色柔毛 ;狭叶锦鸡儿叶片厚度 /面积比值和长 /宽比值大于小叶锦鸡儿 ,叶片生物量小于小叶锦鸡儿。这些特性使狭叶锦鸡儿比小叶锦鸡儿保水能力强。由于狭叶锦鸡儿的渗透调节物质浓度大于小叶锦鸡儿 ,导致细胞渗透势低于小叶锦鸡儿 ;狭叶锦鸡儿叶含水量低于小叶锦鸡儿 ,束缚水 /自由水比值大于小叶锦鸡儿 ;狭叶锦鸡儿叶水势低于小叶锦鸡儿。这些特性表明狭叶锦鸡儿比小叶锦鸡儿渗透调节能力强。狭叶锦鸡儿的蒸腾速率和日蒸腾积累值均小于小叶锦鸡儿 ,水分利用效率高于小叶锦鸡儿。由于叶形态变异、良好的渗透调节功
[23]
MeehlGA, ArblasterJM, TebaldiC (2005). Understanding future patterns of increased precipitation intensity in climate model simulations .Geophysical Research Letters, 32, 109-127. [本文引用: 1]
[24]
NiuH, LiHP, ZhaoML, HanX, DongXH (2008). Relationship between soil water content and vertical distribution of root system under different ground water gradients in Maowusu Sandy Land .Journal of Arid Land Resources and Environment, 22, 157-163.(in Chinese with English abstract) [牛海, 李和平, 赵萌莉, 韩雄, 董晓红 (2008). 毛乌素沙地不同水分梯度根系垂直分布与土壤水分关系的研究 . 干旱区资源与环境, 22, 157-163.] [本文引用: 1]
[25]
PhillipsDL, GreggJW (2003). Source partitioning using stable isotopes: Coping with too many sources . Oecologia, 136, 261-269.https://doi.org/10.1007/s00442-003-1218-3URLPMID:12759813 [本文引用: 1]摘要 Abstract Stable isotopes are increasingly being used as tracers in environmental studies. One application is to use isotopic ratios to quantitatively determine the proportional contribution of several sources to a mixture, such as the proportion of various pollution sources in a waste stream. In general, the proportional contributions of n+1 different sources can be uniquely determined by the use of n different isotope system tracers (e.g., delta13C, delta15N, delta18O) with linear mixing models based on mass balance equations. Often, however, the number of potential sources exceeds n+1, which prevents finding a unique solution of source proportions. What can be done in these situations? While no definitive solution exists, we propose a method that is informative in determining bounds for the contributions of each source. In this method, all possible combinations of each source contribution (0-100%) are examined in small increments (e.g., 1%). Combinations that sum to the observed mixture isotopic signatures within a small tolerance (e.g., +/-0.1 per thousand ) are considered to be feasible solutions, from which the frequency and range of potential source contributions can be determined. To avoid misrepresenting the results, users of this procedure should report the distribution of feasible solutions rather than focusing on a single value such as the mean. We applied this method to a variety of environmental studies in which stable isotope tracers were used to quantify the relative magnitude of multiple sources, including (1) plant water use, (2) geochemistry, (3) air pollution, and (4) dietary analysis. This method gives the range of isotopically determined source contributions; additional non-isotopic constraints specific to each study may be used to further restrict this range. The breadth of the isotopically determined ranges depends on the geometry of the mixing space and the similarity of source and mixture isotopic signatures. A sensitivity analysis indicated that the estimated ranges vary only modestly with different choices of source increment and mass balance tolerance parameter values. A computer program (IsoSource) to perform these calculations for user-specified data is available at http://www.epa.gov/wed/pages/models.htm.
[26]
PhillipsDL, NewsomeSD, GreggJW (2005). Combining sources in stable isotope mixing models: Alternative methods .Oecologia, 144, 520-527.https://doi.org/10.1007/s00442-004-1816-8URLPMID:15711995 [本文引用: 1]摘要 Stable isotope mixing models are often used to quantify source contributions to a mixture. Examples include pollution source identification; trophic web studies; analysis of water sources for soils, plants; or water bodies, and many others. A common problem is having too many sources to allow a unique solution. We discuss two alternative procedures for addressing this problem. One option is a priori to combine sources with similar signatures so the number of sources is small enough to provide a unique solution. Aggregation should be considered only when isotopic signatures of clustered sources are not significantly different, and sources are related so the combined source group has some functional significance. For example, in a food web analysis, lumping several species within a trophic guild allows more interpretable results than lumping disparate food sources, even if they have similar isotopic signatures. One result of combining mixing model sources is increased uncertainty of the combined end-member isotopic signatures and consequently the source contribution estimates; this effect can be quantified using the IsoError model (http://www.epa.gov/wed/pages/models/isotopes/isoerror1_04.htm). As an alternative to lumping sources before a mixing analysis, the IsoSource mixing model (http://www.epa.gov/wed/pages/models/ isosource/isosource.htm) can be used to find all feasible solutions of source contributions consistent with isotopic mass balance. While ranges of feasible contributions for each individual source can often be quite broad, contributions from functionally related groups of sources can be summed a posteriori, producing a range of solutions for the aggregate source that may be considerably narrower. A paleohuman dietary analysis example illustrates this method, which involves a terrestrial meat food source, a combination of three terrestrial plant foods, and a combination of three marine foods. In this case, a posteriori aggregation of sources allowed str
[27]
SaxenaRK (1984). Seasonal variations of oxygen-18 in soil moisture and estimation of recharge in esker and moraine formations .Nordic Hydrology, 15, 235-242.
[28]
SchwinningS, EhleringerJR (2001). Water use trade-offs and optimal adaptations to pulse-driven arid ecosystems .Journal of Ecology, 89, 464-480.https://doi.org/10.1046/j.1365-2745.2001.00576.xURL [本文引用: 1]摘要 Summary Top of page Summary Introduction Methods Results Discussion Acknowledgements References 1 We introduce a hydraulic soil-plant model with water uptake from two soil layers; one a pulse-dominated shallow soil layer, the other a deeper soil layer with continuous, but generally less than saturated soil moisture. Water uptake is linked to photosynthetic carbon assimilation through a photosynthesis model for C 3 plants. 2 A genetic algorithm is used to identify character suites that maximize photosynthetic carbon gain for plants that experience a particular soil moisture pattern. The character suites include allocation fraction to stem, leaves and shallow root, stem capacitance and stem water storage capacity, maximal leaf conductance and sensitivity of leaf conductance to plant water potential, and a critical soil water potential at which shallow roots cease to transfer water. 3 We find that if pulse water is a more important water source than deeper soil water in the environment, optimal phenotypes lean towards adaptations that maximize pulse water use (small root:shoot ratio, predominantly shallow root system, high leaf conductance with high stomatal sensitivity to plant water status). If deeper soil water is more important, phenotypes lean towards adaptations that maximize deeper soil water use (large root:shoot ratio, predominantly deep root system, lower leaf conductance with low stomatal sensitivity). Stem succulence is adaptive only when deeper soil water is unavailable. 4 From among the continuum of derived phenotypes, four phenotypes are selected that resemble the character suites of winter annuals, drought-deciduous perennials, evergreen perennials and stem succulents. Under common conditions, these phenotypes reproduce many of the responses to drought and water pulse observed in their respective life-form counterparts. The comparison also highlights the differences in plant life-form sensitivity to summer and winter drought conditions. 5 Based on these results, we discuss the possible role of annual precipitation patterns in shaping plant adaptations and determining the plant composition of arid and semi-arid environments.
[29]
SchwinningS, StarrBI, EhleringerJR (2005). Summer and winter drought in a cold desert ecosystem (Colorado Plateau) part I: Effects on soil water and plant water uptake .Journal of Arid Environments, 60, 547-566.https://doi.org/10.1016/j.jaridenv.2004.07.003URL [本文引用: 1]摘要 We investigated the effects of winter and summer drought on plants of the Colorado Plateau in western North America. This winter-cold, summer-hot desert region receives both winter and summer precipitation. Droughts were imposed for two consecutive years using rainout shelters. Here, we examine drought effects on the hydrologic interactions between plants and soil. We chose three perennial species for this study, representing different rooting patterns and responsiveness to precipitation pulses: Oryzopsis hymenoides, a perennial bunch grass with shallow roots; Gutierrezia sarothrae, a subshrub with dimorphic roots; and Ceratoides lanata, a predominantly deep-rooted woody shrub. Drought effects on plant water status were qualitatively similar among species, despite morphological differences. Summer drought affected the water status of all species more negatively than winter drought. Isotopic analysis of stem water revealed that all three species took up deeper soil water under drought conditions and shallow soil water after a large rainfall event in summer. Thus all three species appeared to use the same water sources most of the time. However, after a particularly dry summer, only the deepest-rooted species continued to take up soil water, while the more shallow-rooted species were either dead or dormant. Our study suggests therefore that increased occurrence of summer drought could favor the most deep-rooted species in ecosystem.
[30]
VitousekPM (1994). Beyond global warming: Ecology and global change .Ecology, 75, 1862-1876. [本文引用: 1]
[31]
WilliamsDG, EhleringerJR (2000). Intra and interspecific variation for summer precipitation use in pinyon-juniper woodlands .Ecological Monographs, 70, 517-537. [本文引用: 2]
[32]
XiongXG, HanXG, BaiYF, PanQM (2003). Increased distribution of Caragana microphylla in rangelands and its causes and consequences in Xilin River Basin . Acta Prataculturae Sinica, 12(3), 57-62.(in Chinese with English abstract) [熊小刚, 韩兴国, 白永飞, 潘庆民 (2003). 锡林河流域草原小叶锦鸡儿分布增加的趋势、原因和结局 . 草业学报, 12(3), 57-62.] [本文引用: 1]
[33]
XuQ, LiH, ChenJ (2011). Water use patterns of three species in subalpine forest, southwest China: The deuterium isotope approach .Ecohydrology, 4, 236-244.https://doi.org/10.1002/eco.179URL [本文引用: 1]
[34]
YuSW, SunZY, ZhouAG, ZhangX, DuanLJ (2012). Determination of water sources of Gobi plants by δD and δ18O stable isotopes in middle reaches of the Heihe River . Journal of Desert Research, 32, 717-723.(in Chinese with English abstract) [余绍文, 孙自永, 周爱国, 张溪, 段丽军 (2012). 用δD、δ18O同位素确定黑河中游戈壁地区植物水分来源 . 中国沙漠, 32, 717-723.] [本文引用: 2]
[35]
ZhangK, FengQ, LüYQ, ZhangB, SiJH (2011). Study on spatial heterogeneity of soil water contents in oasis-desert belt of Minqin .Journal of Desert Research, 31, 1149-1155.(in Chinese with English abstract) [张凯, 冯起, 吕永清, 张勃, 司建华 (2011). 民勤绿洲荒漠带土壤水分的空间分异研究 . 中国沙漠, 31, 1149-1155.]URL [本文引用: 1]摘要 通过野外试验和室内分析,对民勤绿洲荒漠带土壤水分的空间分异进行了研究。结果表明,在水平方向上,土壤水分的平均最大值基本出现在绿洲区,最小值出现在绿洲荒漠过渡带。在绿洲区,土壤含水量由表层向下逐渐递增;过渡带土壤含水量呈现出表层和底层低,中间层高的特点;荒漠区土壤含水量呈现出表层和底层高,中层低的特点。土壤水分特性的变异系数最大的是过渡带,最小的是绿洲。土壤水分含量各层最佳拟合模型不尽相同。水分含量的自相关性不强,受随机因素的影响较大。
[36]
ZhangLM, LiuXP, ZhaoXY, ZhangTH, YueXF (2014). Response of sandy vegetation characteristics to precipitation change in Horqin Sandy Land .Acta Ecologica Sinica, 34, 2737-2745.(in Chinese with English abstract) [张腊梅, 刘新平, 赵学勇, 张铜会, 岳祥飞 (2014). 科尔沁固定沙地植被特征对降雨变化的响应 . 生态学报, 34, 2737-2745.]https://doi.org/10.5846/stxb201306191738URL [本文引用: 4]摘要 选择科尔沁固定沙地利用一种野外增减雨试验装置研究了沙地植被生长特征对降雨增减变化的响应。结果表明:(1)在6月,降雨增减变化对植物群落高度有显著影响(P0.05)。减雨60%和30%时,植物群落平均高度比对照分别降低8.8%和2.3%,增雨60%和30%时,则分别增加6.8%和1.4%;相比增雨60%,增雨30%更能促进群落盖度的增大;降雨量变化影响群落植株密度。(2)1a的降雨增减变化对沙地植被的多样性和均匀度均没有显著影响,但减雨可显著增加7月物种的丰富度(P0.05)。(3)随着降雨量的增加,地上生物量逐渐增大,在增雨30%时达到最大值;而地下生物量会随着降雨量增加而显著增大,同时,减雨60%也使地下生物量增加。此外,降雨量的增加和减少都会使地下与地上生物量的比值增加。(4)固定沙地地下生物量主要分布在0—20 cm之间,占总地下生物量的52.7%;降雨量的增加显著增加20—40 cm土壤中根系的分布,当降雨量减少60%时,20—40 cm土壤中根系的分布也略有增加,增雨60%和减雨60%对地下生物量在40—60 cm土层的分布具有明显的促进作用。
[37]
ZhaoLJ, XiaoHL, ChengGD (2008). A preliminary study of water sources of riparian plants in the lower reaches of the Heihe Basin .Acta Geoscientica Sinica, 29, 709-718.(in Chinese with English abstract) [赵良菊, 肖洪浪, 程国栋 (2008). 黑河下游河岸林植物水分来源初步研究 . 地球学报, 29, 709-718.] [本文引用: 1]
[38]
ZhaoWZ, LiuZM, ChangXL (2002). Skewness and inequality of height distribution of young Pinus sylvestris var. mongolica stands introduced on sandy soil with lower limited precipitation for tree survival and normal growth . Chinese Journal of Applied Ecology, 13, 6-10.(in Chinese with English abstract) [赵文智, 刘志民, 常学礼 (2002). 降水量下限引种区沙地樟子松幼林种群树高分布偏斜度和不整齐性 . 应用生态学报, 13, 6-10.]URL [本文引用: 2]摘要 对降水量下限引种区沙地樟子松自然稀疏前人工林种群树高分布偏斜度和不整齐性的研究表明,种群高生长的不 整齐性和分布的不对称性在林分自疏前随着林分发育而下降;造成高生长不整齐性变化的因素主要有年龄、初始生长状况、种群密度、对土壤水分的竞争和沙丘部位 的差异;沙丘部位对林分不整齐性的影响作用要大于密度.
[39]
ZhengXR, ZhaoGQ, LiXY, LiL, WuHW, ZhangSY, ZhangZH (2015). Application of stable hydrogen isotope in study of water sources for Caragana microphylla bush land in Nei Mongol . Chinese Journal of Plant Ecology, 39, 184-196.(in Chinese with English abstract) [郑肖然, 赵国琴, 李小雁, 李柳, 吴华武, 张思毅, 张志华 (2015). 氢同位素在内蒙古小叶锦鸡儿灌丛水分来源研究中的应用 . 植物生态学报,39, 184-196.]https://doi.org/10.17521/cjpe.2015.0018URL [本文引用: 2]摘要 小叶锦鸡儿(Caragana microphylla)是我国内蒙古草原广泛分布的优势灌木,目前国内外有关小叶锦鸡儿水分利用来源的定量研究较少.该文运用氢稳定同位素法,分析了内蒙古小叶锦鸡儿木质部水和潜在水源(夏季降雨、冬季降雪和土壤水)的氢稳定同位素比率(δD)的季节变化,发现小叶锦鸡儿在遮雨处理和自然状态下δD季节变化差异不明显,分别为-101.36‰±13.02%和-101.18%±12.71‰.在遮雨处理条件下,小叶锦鸡儿主要利用0-20 cm土壤层水分,占其所利用水分的73.30%±16.14%;自然状态下,小叶锦鸡儿对0-20、20-60和60-100 cm不同土壤层土壤的水分利用较为均衡,分别为34.66%±7.83%、32.44%±7.42%和32.90%±4.14%.该结果表明不同生境下灌木与草本的水分竞争可能是小叶锦鸡儿对不同土层水分利用差异的原因.结合两源模型分析了降雨对各土壤层的贡献率,并用IsoSource多源混合模型分析了各土壤层对小叶锦鸡儿用水的贡献率,发现降雨对小叶锦鸡儿的贡献率为42.65%-63.92%.该结果反映了小叶锦鸡儿对夏季降雨和冬季融雪的利用情况.
[40]
ZhouH, ZhengXJ, TangLS, LiY (2013). Differences and similarities between water sources of Tamarix ramosissima, Nitraria sibirica and Reaumuria soongorica in the southeastern Junggar Basin . Chinese Journal of Plant Ecology, 37, 665-673.(in Chinese with English abstract) [周海, 郑新军, 唐立松, 李彦 (2013). 准噶尔盆地东南缘多枝柽柳、白刺和红砂水分来源的异同 . 植物生态学报, 37, 665-673.]https://doi.org/10.3724/SP.J.1258.2013.00069URL [本文引用: 1]摘要 荒漠生态系统中,水是植物生长最主要的限制因子。为了比较同一生境下不同荒漠植物的水分来源特征,选取了同一生境下的多枝柽柳(Tamarix ramosissima)、白刺(Nitraria sibirica)和红砂(Reaumuria soongorica),测定了这3种植物茎水和各潜在水源(降水、土壤水和地下水)的氢、氧稳定同位素比率(δD和δ18O)值,并利用IsoSource软件计算了3种植物对潜在水源的利用比例。结果表明:红砂和白刺的茎水δD和δ18O值及其水分来源有明显的季节波动特征。其中,红砂为浅根系植物,春季(3-5月)以表层土壤水为主要水源,夏秋季节(6-10月)表层土壤含水量显著降低,其主要的水分来源逐渐偏向于较深层的土壤水;白刺的根系分布范围介于红砂和多枝柽柳之间,在春季能够较多地利用表层土壤水,而到了夏秋季节,所利用的水分更多地来源于深层土壤水或地下水;多枝柽柳为深根系植物,其90%以上的水分来源于深层土壤水和地下水,而且茎水δD和δ18O值及其水分来源没有季节波动特征。3种植物水分来源特征的差异与其水分利用策略密切相关,同时,也说明荒漠灌木可以通过自身调节向着最优(最有利)表现型发展,从而最大限度地获取水分。
[41]
ZhouYD, ChenSP, SongWM, LuQ, LinGH (2011). Water- use strategies of two desert plants along a precipitation gradient in northwestern China .Chinese Journal of Plant Ecology, 35, 789-800.(in Chinese with English abstract) [周雅聃, 陈世苹, 宋维民, 卢琦, 林光辉 (2011). 不同降水条件下两种荒漠植物的水分利用策略 . 植物生态学报, 35, 789-800.] [本文引用: 1]
[42]
ZhuYJ, ZhaoXB, LiuYS, LiY, FanWX (2017). Water use process of Salix psammophila and Salix cheilophila in Gonghe Basin, Qinghai Province . Journal of Desert Research, 37, 281-287.(in Chinese with English abstract) [朱雅娟, 赵雪彬, 刘艳书, 李蕴, 范文秀 (2017). 青海共和盆地沙柳(Salix psammophila)和乌柳(Salix cheilophila)的水分利用过程 . 中国沙漠, 37, 281-287.] [本文引用: 1]
Tracing of water movement in the unsaturated zone using stable isotopes of hydrogen and oxygen 1 1998
Soil water and temperature dynamics in shrub-encroached grasslands and climatic implications: Results from Inner Mongolia steppe ecosystem of north China 1 2013
... 科尔沁沙地位于内蒙古东部西辽河中下游, 是我国最大的沙地, 也是我国沙漠化最为严重的一个地区.在全球气候变化的影响下, 该地区年降水量呈减少趋势, 气候趋于干旱化(张腊梅等, 2014).作为科尔沁沙地的主要优势种, 小叶锦鸡儿和盐蒿的水分利用对当地生态系统的水分平衡和植物群落组成产生了重要的影响(熊小刚等, 2003; Li et al., 2013).本研究应用稳定性同位素技术分析模拟降水条件下小叶锦鸡儿和盐蒿水分利用动态以及变化特征, 研究结果对认识干旱半干旱区气候变化下植物的适应性, 以及对植被保护、恢复与重建具有参考价值, 为合理安排当地种植固沙植株的密度及空间分布提供指导. ...

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT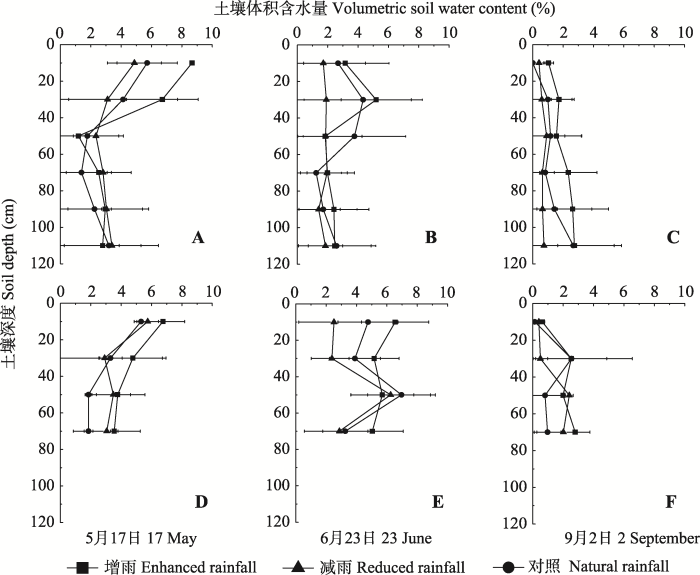 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT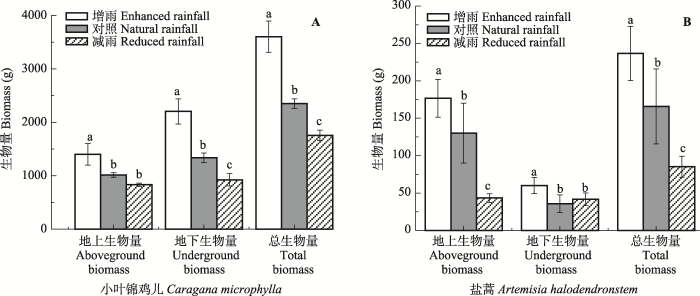 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT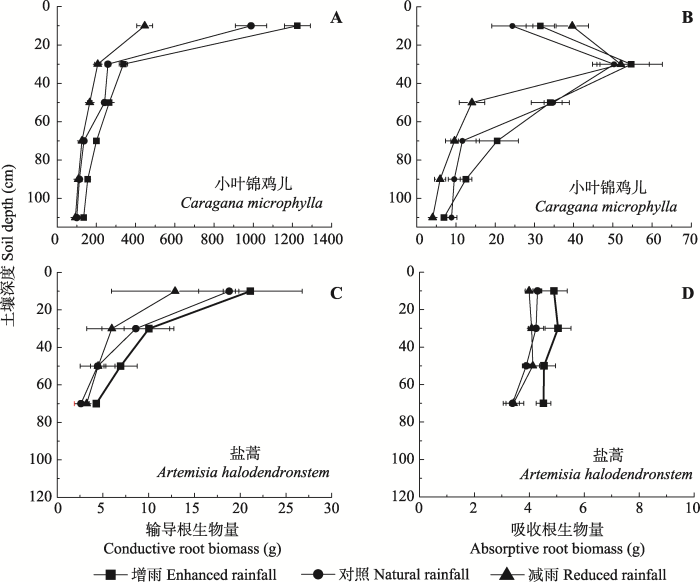 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT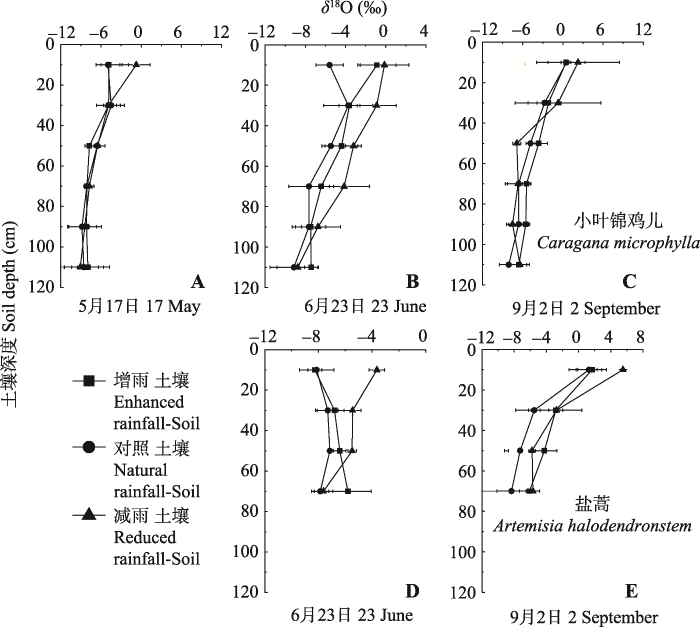 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT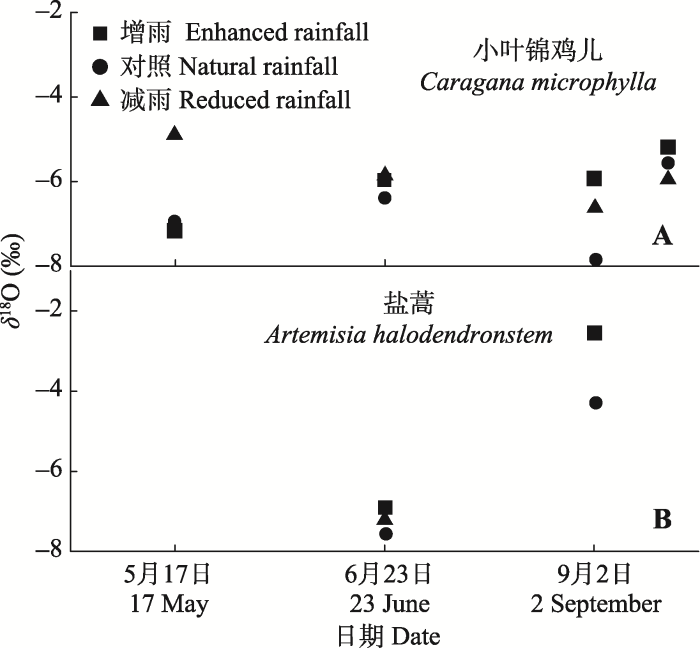 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}