
Characteristic and representativeness of the vertical vegetation zonation along the altitudinal gradient in Shennongjia Natural Heritage
MAMing-Zhe通讯作者:
收稿日期:2017-04-07
接受日期:2017-08-29
网络出版日期:2017-11-10
版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
关键词:
Abstract
Methods Based on the community survey data of the six vegetation zonation along the elevational gradients in Shennongjia, the global land use dataset, and the literature data of the communities along the altitudinal gradients of other natural heritage sites and the nominated world natural heritage sites in Oriental Deciduous Forest Biogeographic Province by Udvardy, we explored the outstanding universal value of the zonal vegetation along the altitude gradients by the methods of spatial analysis.
Important findings Shennongjia heritage site preserves the intact vegetation zonation of the typical Oriental Deciduous Forest Biogeographical Province in the Classification of the Biogeographical Provinces of the World by Udvardy, including evergreen broad-leaved forests (South Slope of the Heritage Site), evergreen deciduous broad-leaved mixed forests, deciduous broad-leaved forests, coniferous and broad-leaved mixed forests, coniferous forests and subalpine shrub and meadow along the elevation gradients. The altitudinal zonation of vegetation in the Shennongjia heritage site represented a variety of bio-ecological processes, such as the turnover of the dominant trees along the altitudinal gradients, and is an outstanding example of the ongoing ecological processes occurring in the development of intact subtropical mixed broadleaved evergreen and deciduous forest in the Northern Hemisphere.
Keywords:
-->0
PDF (3798KB)元数据多维度评价相关文章收藏文章
本文引用格式导出EndNoteRisBibtex收藏本文-->
山地是指具有一定海拔高度、相对高度和坡度的地面(王襄平等, 2004)。山地覆盖全球陆地地表近25%, 全球近1/3世界自然遗产地分布于山地(Barthlott et al., 1996; He et al., 2016)。山地因其在较小的空间范围内显著的环境梯度, 浓缩了水平自然带的自然地理和生态学特征(Korner, 2007)。山地因受第四纪冰川和现代人类活动的影响较小而保存了良好的自然生境和植被, 形成了显著的植被垂直带谱, 成为生物多样性的避难所和关键生物生态学过程的集中展示地(Beniston, 2003)。生态学家和生物地理学家很早就认识到山地植被垂直带谱的生物生态学意义和突出价值(Sundqvist et al., 2013)。1914年, Grinnell首次认识到山地海拔梯度对动植物分布的限制作用, 并认识到山地海拔梯度决定动植物进化和适应等生物生态学方面的重要性(Grinnell, 1924)。20世纪Whittaker开展了美国大烟山(Great Smoky Mountains)和锡斯基尤山(Siskiyou Mountains)山地植被垂直带谱的生物生态学研究(Whittaker, 1956, 1960)。近年来, 日趋增多的山地群落多样性(Rahbek, 2005)和生态系统过程等方面的研究, 进一步证实了山地植被垂直带谱在生物生态学过程方面的重要价值(Raich et al., 1997; Rahbek, 2005; Colwell et al., 2008)。
中国是一个山地大国, 国土面积的2/3为山地(中国科学院中国自然地理编委会, 1980), 15处被列入《世界自然遗产名录》(其中4处为世界自然和文化双遗产地)的自然遗产地均位于山地(马克平, 2016)。充分认识并掌握我国自然遗产地山地植被垂直带谱的代表性, 凝练其世界自然遗产地全球突出普遍价值, 对正确地评估自然遗产地的保护价值进而制定相关保护管理政策, 具有重要的科学意义和现实意义。
神农架自然遗产地地处Udvardy全球生物地理区划(Giogeographic Classification System)中的东方落叶林生物地理省, 山体海拔高差达2 700 m, 从低海拔到高海拔依次呈现出亚热带、暖温带、温带、寒温带等气候特点, 海拔垂直分异显著。本研究围绕植被垂直带谱的代表性, 从植被垂直带谱的地带性、完整性以及不同垂直带群落物种更替等角度, 分析论证了神农架自然遗产地植被垂直带谱的突出遗产价值, 以期为神农架自然遗产地的科学保护和管理提供依据。
1 材料和方法
1.1 研究区概况
神农架自然遗产地主体位于湖北神农架国家级自然保护区, 包括部分巴东沿渡河自然保护区, 总面积73 318 hm2, 缓冲区总面积41 536 hm2。遗产地位于中国地势第二阶梯的东部边缘, 为大巴山脉东段组成的中山地貌, 呈近东西方向延伸, 地势西南高东北低, 最高海拔神农顶3 106.2 m, 为华中第一峰, 最低海拔400 m (遗产地南坡)。神农架自然遗产地地跨中亚热带和北亚热带, 年平均气温12.1 ℃, 最冷月(1月)平均气温-8 ℃, 最热月(7月)平均气温26.5 ℃。年降水量800-2 500 mm, 有明显的季节性。1.2 研究方法
1.2.1 群落调查在神农架自然遗产地沿海拔梯度设置14条调查样线, 每条样线按海拔每升高100 m设置1个样地, 或选取特殊的群落类型设置样地。森林样地大小为20 m × 20 m (部分样地为20 m × 30 m或30 m × 30 m), 灌丛样地大小为10 m × 10 m, 草丛样地大小为5 m × 5 m。共调查森林样地166个, 灌丛样地21个, 草丛样地4个。另外, 在常绿阔叶林带、常绿落叶阔叶混交林带、落叶阔叶林带、针叶林带还调查了4个100 m × 100 m的固定样地。群落环境因子记录地名、经纬度、海拔、坡向、坡度、土壤类型、干扰因子等; 森林样地乔木层进行每木调查, 记录种名、高度、胸径和冠幅; 灌木层设置5个2 m × 2 m的样方, 草本层设置5个1 m × 1 m的样方, 分种记录种名、高度、盖度和多度。
1.2.2 文献资料收集整理
基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息。同时, 收集整理《中国植物地理》(应俊生和陈梦玲, 2011)、《湖南植被》(祁承经, 1990)、《湖南森林》(湖南森林编委会, 1991)、《安徽森林》(安徽森林编委会, 1990)、《安徽植被》(安徽植被编委会, 1983)、《江西森林》(江西植被编委会, 1983)、《陕西植被》(雷明德, 1999)、《陕西森林》(陕西森林编委会, 1989)、《湖北森林》(湖北森林编委会, 1990)、《贵州森林》(贵州森林编委会, 1989)、《四川森林》(四川植被编委会, 1983)、《神农架自然保护区科学考察集》(朱兆泉和宋朝枢, 1999)、《神农架地区自然资源综合调查报告》(廖明尧, 2015)等书籍以及相关文献(谭景燊等, 1982; 李晓东, 1985; 吴国芳等, 1988; 田自强, 2002; 何飞, 2006), 整理分析全球生物地理区划(Udvardy)中东方落叶林生物地理省内已有世界自然遗产地和世界自然遗产预备清单中主要山地植被垂直带谱的群落类型、物种组成等资料。
1.2.3 对比分析
世界自然遗产的遴选有4条标准, 即美学、地质、生物生态学和栖息地标准。世界自然遗产地全球突出普遍价值(outstanding universal value)的提炼, 是世界自然遗产地遴选及保护的关键。全球生物地理区划(Udvardy)为国际自然保护联盟(IUCN)和联合国教科文组织(UNESCO)世界遗产中心遴选世界自然遗产地的重要科学依据。依据全球自然地理环境特征和生物群落类型, Udvardy把全球地表划分为8个生物地理区(biogeographical realm)、14个生物群区(biome)和193个生物地理省(biogeographical province)(Udvardy, 1975)。自然遗产地在美学、地质、生物生态学和栖息地等某一方面或几方面的遗产价值在其所在生物地理省具有突出性和唯一性, 且其遗产价值完整, 则该项自然遗产地具有全球突出普遍价值。神农架自然遗产地位于全球生物地理区划193个生物地理省的东方落叶林生物地理省(Oriental Deciduous Forest Biogeographical Province)。本研究收集了东方落叶林生物地理省已有世界自然遗产地中国黄山、三清山、九寨沟, 日本白神山、日本富士山, 以及世界自然遗产地预备清单中的中国太白山、梵净山、八大公山等山地植被垂直带谱组成等数据, 对比分析了神农架自然遗产地与东方落叶林生物地理省内已有世界自然遗产地和预备世界自然遗产地海拔梯度上植被垂直带谱的完整性, 并分析了其植被垂直带谱的保护状态。
2 研究结果
2.1 神农架自然遗产地保存有北半球同纬度少有的地带性常绿落叶阔叶混交林
受副热带高压的控制, 全球30°-35° N大部分地段主要植被类型为疏林、荒漠植被(表1; 图1), 仅中国神农架, 美国东南部大烟山, 地中海沿岸, 以及日本本州、四国、九州和琉球群岛北部等地, 分布有地带性常绿落叶阔叶混交林。神农架自然遗产地地跨中亚热带和北亚热带, 青藏高原的隆起使神农架自然遗产地北坡保存有全球同纬度最为完好的地带性常绿落叶阔叶混交林带, 其既不同于地中海型耐旱热的硬叶常绿林, 也不同于北半球同纬度的亚热带、热带荒漠植被。常绿落叶阔叶混交林是遗产地北坡的基带植被类型, 也是北亚热带的地带性代表类型。神农架自然遗产地常绿落叶阔叶混交林带以壳斗科树种为主, 其中落叶的主要为栎属(Quercus)和水青冈属(Fagus)等, 常绿的则以青冈属(Cyclobalanopsis)、栲属(Castanopsis)和石栎属(Lithocarpus)等为主, 其优势树种完全不同于美国东南部大烟山,地中海沿岸, 以及日本本州、四国、九州和琉球群岛北部等地的优势树种。其中, 日本本州、四国、九州和琉球群岛北部地带性常绿落叶阔叶混交林主要为以Castanopsis cuspidata、C. cuspidate var. sieboldi、Quercus salicina、Q. acuta、Q. sessilifolia、Machilus thunbergii等为优势的栲类、润楠类常绿落叶林(何飞, 2006)。地中海沿岸的地带性植被类型则为在地中海气候下形成的常绿落叶阔叶混交林, 主要为以Q. coccifera、Q. ilex、Q. suber和Pistacia lentiscus等为优势种的硬叶常绿阔叶林(Walter, 1984)。美国东南部大烟山的常绿阔叶落叶混交林主要以Q. borealis、Tilia heterophylla、Betula alleghenicnsis和Acer rubrum等为优势种(Whittaker, 1956)。Table 1
表1
表1神农架及北半球同纬度典型地区概况
Table 1Overview of Shennongjia and its counterparts in the same latitude
| 国家 Country | 地区 Area | 植被类型 Vegetation type | 位置 Location | 海拔 Altitude (m) | 年平均气温 Mean annual temperature (℃) | 年降水量 Mean annual precipitation (mm) | 优势种 Dominant species | 土壤类型 Soil type |
|---|---|---|---|---|---|---|---|---|
| 中国 China | 神农架 Shennongjia | 常绿落叶阔叶混交林 Evergreen deciduous broad-leaved mixed forest | 31.27° N, 110.48° E | 1 670 | 10.6 | 1 200 | 米心水青冈, 青冈 Fagus engleriana, Cyclobalanopsis glauca | 山地黄棕壤 Mountain yellow brown soil |
| 日本 Japan | 本州南部低地, 四国, 九州和琉球群岛的北部 The southern lowlands of Honshu, north part of Shikoku, Kyushu and Ryuryu | 常绿阔叶林 Evergreen broad-leaved forest | 26.58°-35.5° N, 130°- 140° E | 250-1 100 | 13-21 | 1 300-2 500 | Castanopsis cuspidata, C. cuspidate var. sieboldii, Quercus salicina, Q. acuta, Q. sessilifolia, Machilus thunbergii | 棕壤 Brown soil |
| 法国、意大利、西班牙、 希腊 France, Italy, Spain, and Greece, | 地中海沿岸 Mediterranean | 硬叶常绿阔叶林 Sclerophyllous evergreen broad-leaved forest | 30.25°- 45.83° N, 0°-30° E | 300-2 800 | 16-20 | 300-1 000 | Quercus coccifera, Q. ilex, Q. suber, Pistacia lentiscus | 红壤和黄壤 Red soil and yellow soil |
| 美国 USA | 大烟山 Great Smoky Mountains | 温带阔叶林 Temperate broad-leaved forest | 35.6° N, 83.48° W | 1 830 | 12 | 1 270-2 030 | Quercus borealis, Tilia heterophylla, Betula alleghenicnsis, Acer rubrum | 山地棕壤 Mountain brown soil |
| 北非 North Africa | 撒哈拉沙漠北部 North of the Sahara Desert | 荒漠 Desert | 25°-35° N, 15°W-50° E | - | >30 | 90-450 | 金合欢属, 蒿属, 三芒草属, 画眉草属, 稷属 Acacia, Artemisia, Aristida, Eragrostis, Panicum | 石漠(岩漠)、砾漠和沙漠 Rock desert (rock desert), gravel desert and desert |
新窗口打开
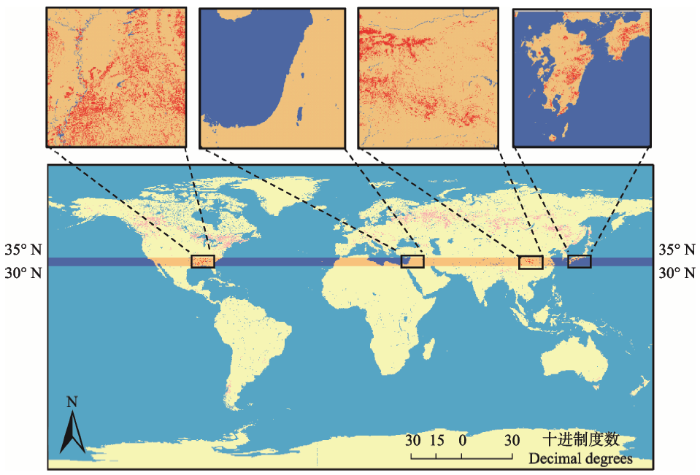 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1全球30°-35° N带上常绿落叶阔叶混交林的分布(红色代表混交林)。
-->Fig. 1The distribution of evergreen deciduous broad-leaved mixed forests (red areas) along the belt of 30°-35° N in the Northern Hemisphere.
-->
2.2 神农架自然遗产地展示了东方落叶林生物地理省植被垂直带谱的物种更替过程
神农架自然遗产地植被垂直带谱在海拔梯度上呈现出随海拔升高优势树种种类和数量逐渐更替的生态学过程: 遗产地南坡为中亚热带北缘, 发育的常绿阔叶林带优势种主要为青冈(Cyclobalanopsis glauca)、曼青冈(C. oxyodon)、小叶青冈(C. myrsinifolia)、巴东栎(Quercus engleriana)等壳斗科的常绿树种。随海拔升高, 部分常绿优势树种逐渐被锐齿槲栎(Q. aliena var. acutiserrata)和米心水青冈(Fagus engleriana)等落叶树种代替, 形成了常绿和落叶树种混交的常绿落叶阔叶混交林带。随着海拔的进一步升高, 常绿优势树种逐渐减少, 锐齿槲栎、米心水青冈等落叶树种成为优势种, 形成了落叶树种为优势种的落叶阔叶林带; 海拔升高到2 200-2 600 m时, 以落叶树种为优势种的落叶阔叶林带中出现针叶树种, 落叶阔叶林逐渐被以巴山冷杉(Abies fargesii)和红桦(Betula albosinensis)为优势种的针、阔叶混交林带所替代。海拔2 600 m以上的地段, 以巴山冷杉和红桦为优势种的针阔混交林带中的红桦基本被针叶树种替代, 形成了以巴山冷杉为优势的寒温带针叶林。海拔3 000 m以上的地段, 优势树种巴山冷杉逐渐被粉红杜鹃(Rhododendron hypoglaucum)、香柏(Sabina squamata var. fargesii)、箭竹(Fargesia spathacea)和野古草(Arundinella hirta)等灌草优势种替代, 形成了高山灌丛草甸植被带(图2, 图3)。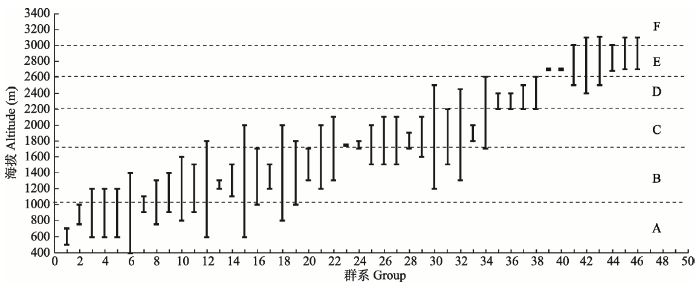 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2神农架海拔梯度上主要植物群系分布图。大写英文字母对应各垂直带: A, 亚热带常绿阔叶林带; B, 北亚热带常绿落叶阔叶混交林带; C, 暖温带落叶阔叶林带; D, 温带针阔混交林带; E, 寒温带针叶林带; F, 亚高山灌丛、草甸带。 横坐标数字对应群系: 1, 蜡梅灌丛; 2, 楠木、小叶青冈为主的常绿阔叶林; 3, 马桑、毛黄栌灌丛; 4, 马尾松、栓皮栎林; 5, 杉木林; 6, 栓皮栎林; 7, 香叶树、小叶青冈、化香树、亮叶桦林; 8, 马尾松林; 9, 尖齿高山栎灌丛; 10, 曼青冈、水丝梨、巴东栎、青冈林; 11, 乌冈栎、岩栎、鹅耳枥、化香树林; 12, 野核桃林; 13, 栓皮栎、锐齿槲栎、茅栗林; 14, 巴东栎、曼青冈、亮叶桦、化香树林; 15, 刺叶栎林; 16, 短柄枹林; 17, 茅栗林; 18, 巴山松林; 19, 亮叶桦、化香树、鹅耳枥林; 20, 华山松、糙皮桦林; 21, 锐齿槲栎林; 22, 秦岭冷杉林; 23, 川榛、鸡树条荚蒾、湖北海棠灌丛; 24, 薹草、地榆、香青、血见愁老鹳草草甸; 25, 野漆树、锐齿槲栎、灯台树、化香树林; 26, 芒、蕨草丛; 27, 美丽胡枝子、绿叶胡枝子灌丛; 28, 薹草、葱状灯芯草、长叶地榆、柳兰沼泽化草甸; 29, 华山松、锐齿槲栎林; 30, 华山松林; 31, 米心水青冈林; 32, 秦岭冷杉、青扦林; 33, 锐齿槲栎、米心水青冈、红桦林; 34, 红桦林; 35, 华山松、山杨、红桦林; 36, 华山松、山杨林; 37, 中华黄花柳、华中山楂、湖北花楸灌丛; 38, 巴山冷杉、红桦、槭类林; 39, 杯腺柳灌丛; 40, 直穗小檗灌丛; 41, 箭竹灌丛; 42, 平枝荀子灌丛; 43, 巴山冷杉林; 44, 粉红杜鹃灌丛; 45, 香柏灌丛; 46, 印度三毛草、紫羊茅、糙野青茅草甸。群系海拔分布信息主要参考田自强(2002)。
-->Fig. 2The distribution of main plant formations in Shennongjia along the elevation gradient. Capital English letters represent vertical vegetation zones: A, subtropical zone of evergreen broad-leaved forest; B, north subtropical zone of Mixed evergreen and deciduous broad-leaved forest; C, warm temperate zone of broadleaved deciduous forest; D, temperate zone of mixed broadleaf-conifer forest; E, cold temperate zone of coniferous forest; F, subalpine zone of shrub meadow. The abscissa represents different formations: 1, Form. Chimonanthus praecox; 2, Form. Phoebe zhennan, Cyclobalanopsis gracilis; 3, Form. Coriaria sinica, Cotinus coggyqria var. pubescens; 4, Form. Pinus massoniana, Quercus variabilis; 5, Form. Cunninghamia lanceolata; 6, Form. Quercus variabilis; 7, Form. Lindera communis, Cyclobalanopsis gracilis, Platycarya strobilacea, Betula luminifera; 8, Form. Pinus massoniana; 9, Form. Quercus acrodonta; 10, Form. Cyclobalanopsis oxyodon, Sycopsis chinense, Quercus engleriana, Cyclobanopsis myrsinaefolia; 11, Form. Quercus philyraeoides, Q. acrodonta, Carpinus sp., Platycarya strobilacea; 12, Form. Juglans cathayensis; 13, Form. Quercus variabilis, Q. aliena var. acuteserrata, Castanea segyinii; 14, Form. Quercus engleriana, Cyclobalanopsis oxyodon, Betula luminifera, Platycarya strobilacea; 15, Form. Quercus spinosa; 16, Form. Quercus glandulifera var. brevipetiolata; 17, Form. Castanea seguinii; 18, Form. Pinus henryi; 19, Form. Betula luminifera, Platycarya strobilacea, Carpinus sp.; 20, Form. Pinups armandii, Betula utilis; 21, Form. Quercus aliena var. acuteserrata; 22, Form. Abies chensiensis; 23, Form. Corylus heterophylla var. sutchuenensis, Viburnum opulus var. calvescens, Malus hupehensis; 24, Form. Carex sp., Sanguisorba officinalis var. longifolia, Anaphalis sinica, Geranium henryi; 25, Form. Rhus verniciflua, Quercus acutidentata, Cornus controversa, Platycarya strobilacea; 26, Form. Miscanthus sinensis, Pteridium aquilinum var. latiusculum; 27, Form. Lespedeza formosa, L. buergeri; 28, Form. Carex sp., Juncus concinus, Sanguisorba officicnalis var. longifolia, Chamaenerion angustifolium; 29, Form. Pinus armandii, Querus aliena var. acuteserrata; 30, Form. Pinus armandii; 31, Form. Fagus engleriana; 32, Form. Abies chensiensis, Picea wilsonii; 33, Form. Quercus aliena var. acuteserrata, Fagus engleriana, Betula albosinensis; 34, Form. Betula albosinensis; 35, Form. Pinus armandii, Populus davidiana, Betula albosinensis; 36, Form. Pinus armandii, Populus davidiana; 37, Form. Salix carprea var. inica, Crataegus wilsonii, Sorbus hupehensis; 38, Form. Abies fargesii, Betula albosinensis, Acer sp.; 39, Form. Salix cupularis; 40, Form. Berberis dasystachya; 41, Form. Sinarundinaria nitida; 42, Form. Cotoneaster horizontalis; 43, Form. Abies fargesii; 44, Form. Rhododendron fargesii; 45, Form. Sabina pingii var. wilsonii; 46, Form. Trisetum clarkei, Festuca rubra, Deyeuxia scabrescens. Formation distribution along the elevation was based on
-->
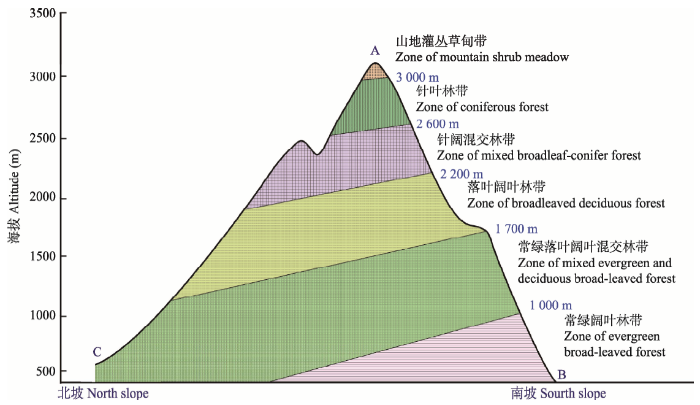 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3神农架山地示意图。A, 神农顶, 神农架遗产地最高点, 海拔3 106.2 m; B, 下谷坪, 神农架遗产地南坡最低点, 海拔400 m; C, 韩家坪, 神农架遗产地北坡最低点, 海拔600 m。
-->Fig. 3Sketch map of Shennongjia mountain. A, Shennongding, the summit of the heritage site with the elevation of 3 106.2 m; B, Xiaguping, the lowest point of the south slope of the heritage site with the elevation of 400 m; C, Hanjiaping, the lowest point of the north slope of the heritage site with the elevation of 600 m.
-->
2.3 神农架自然遗产地很好地保存了东方落叶林生物地理省完整的植被垂直带谱
比较分析神农架自然遗产地植被垂直带谱与东方落叶林生物地理省内世界自然遗产地及预备世界自然遗产地的植被垂直带谱(表2)发现: 神农架自然遗产地垂直高差达2 700 m, 形成了完整的山地植被垂直带谱, 山地自下而上依次发育有常绿阔叶林带(遗产地南坡)、常绿落叶阔叶混交林带、落叶阔叶林带、针阔混交林带、针叶林带及亚高山灌丛和草甸带(图3)。与神农架海拔梯度上植被垂直带相比, 安徽黄山、江西三清山海拔带上无温性针阔混交林带和寒温性针叶林带, 黄山海拔800-1 800 m分布的黄山松林、三清山1 400-1 700 m分布的黄山松林均为暖性至中温性针叶林(蒋木青等, 1982)。湖南八大公山海拔带上无温性针阔混交林带和寒温性针叶林带。梵净山的植被垂直带谱为以梵净山冷杉为优势种的针阔混交林, 分布于较窄的2 100-2 350 m海拔范围内, 植被垂直带谱中缺少寒温性针叶林带。日本白神山地带性植被类型为落叶阔叶林, 但其植被垂直带谱中缺少常绿阔叶林带和常绿落叶阔叶混交林带(表2)。日本富士山地带性植被类型为温带落叶阔叶林, 其垂直带谱缺失常绿阔叶林带、常绿落叶阔叶混交林带以及针阔混交林带(Ohsawa, 1984) (表2)。四川九寨沟地带性植被类型为温性针阔混交林, 其植被垂直带谱中缺少落叶阔叶林带、常绿落叶阔叶混交林带和常绿阔叶林带(表2)。与神农架自然遗产地相比, 秦岭太白山植被垂直带谱也比较完整, 但由于长期的人类活动, 其地带性常绿落叶阔叶混交林带已被严重破坏, 目前无完好的成片的常绿落叶阔叶混交林带分布, 现存植被类型主要为马尾松(Pinus massoniana)林、麻栎(Quercus acutissima)林等(表2)。比较分析结果表明, 神农架自然遗产地在较小的水平距离内浓缩了亚热带、暖温带、中温带和寒温带的典型植被类型, 保存有完整的植被垂直带谱, 在东方落叶林生物地理省中具有唯一性和代表性。Table 2
表2
表2Udvardy东方落叶林生物地理省内主要山地的现存植被垂直带谱
Table 2The altitudinal vegetation zonation of the Eastern Deciduous Forest Biogeographic Province in the Classification of the Biogeographical Provinces of the World by Udvardy
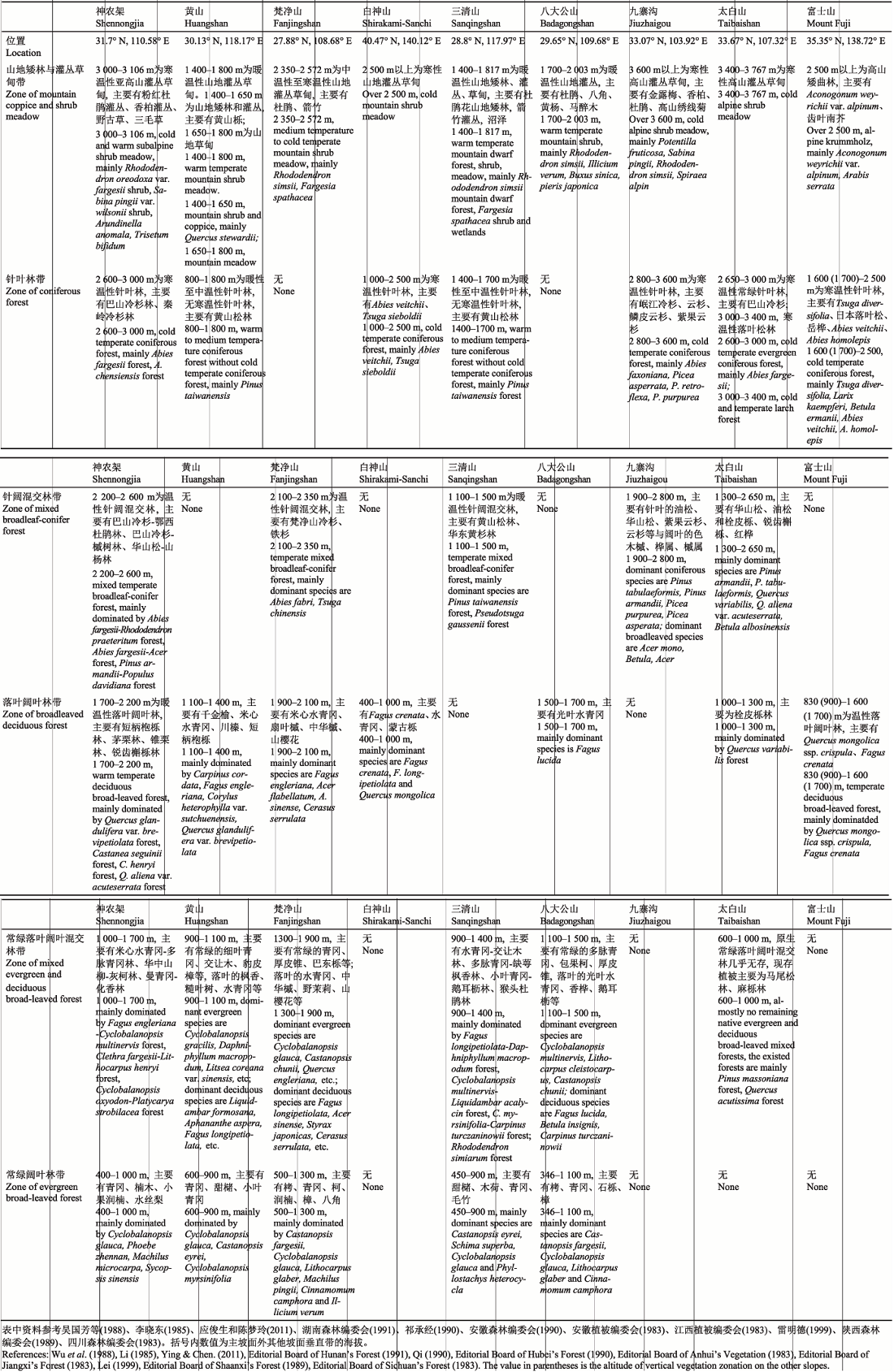 |
新窗口打开
3 讨论
随着海拔升高, 山地气温、气压和地表面积下降, 山地环境条件发生显著变化(Korner, 2007)。植被垂直带谱是山地海拔梯度上生物和非生物因素综合作用的结果(He et al., 2016)。海拔梯度以较小的地理尺度, 在相似的环境条件下呈现出较大的气候梯度, 浓缩了不同的植被类型, 是众多生物生态学过程的集中展示场所, 也是研究生物生态学过程随气候变化的理想场所(Malhi et al., 2010)。青藏高原的隆起, 使受副热带高压控制下的东亚亚热带形成全球独一无二的东亚季风型(夏季湿润、冬季干冷)亚热带常绿阔叶林, 其既不同于地中海型耐旱热的硬叶常绿林, 也不同于北半球同纬度的亚热带、热带荒漠植被(中国科学院中国植被图编辑委员会, 2007)。神农架自然遗产地地跨中亚热带和北亚热带, 孕育了北半球典型的地带性常绿落叶阔叶混交林。同时, 神农架自然遗产地的海拔梯度在较小的水平距离范围内浓缩了中亚热带、北亚热带、暖温带、温带和寒温带等气候类型, 孕育了全球生物地理区划东方落叶林生物地理省内完整的植被垂直带谱, 形成了从低海拔到高海拔的常绿阔叶林、常绿落叶阔叶混交林、落叶阔叶林、针阔混交林、针叶林及亚高山灌丛和草甸等植被类型, 展示了东方落叶林生物地理省典型的生物生态学过程。神农架自然遗产地海拔梯度上完好的植被垂直带谱, 展示了全球生物地理区划东方落叶林生物地理省的植物群落优势种和功能群随海拔梯度的更替过程。研究发现: 山地海拔梯度上的植被垂直带谱展示了植物群落优势种和功能群随海拔梯度变化的更替过程(Whittaker, 1956; Sundqvist et al., 2013), 是研究功能性状随气候变化的理想场所。神农架自然遗产地低海拔分布的常绿阔叶林群落, 其优势种主要为青冈、曼青冈、小叶青冈和巴东栎等壳斗科的常绿树种, 比叶面积高, 叶氮含量高, 单位叶面积营养元素含量高。随海拔升高, 群落优势种逐渐被比叶面积低、叶氮含量低、单位叶面积营养元素含量低(nutrient amounts per leaf area)的物种所替代(Vitousek et al., 1988; Tanner et al., 1998; Salinas et al., 2011), 如锐齿槲栎、米心水青冈、红桦等落叶树种以及巴山冷杉等寒温带针叶树种。高海拔地段植被群落主要为矮化优势种组成, 如粉红杜鹃、香柏、箭竹和野古草等, 其功能群生长率低, 适应高海拔胁迫环境。
神农架自然遗产地海拔梯度上不同植被带内的动物类群, 展示了东方落叶林生物地理省不同功能动物类群随海拔梯度变化的更替过程。已有研究发现, 海拔垂直梯度上动物群落主要通过改变其多度及物种更替适应海拔梯度上的不同环境条件(Hodkinson, 2005; Sanders et al., 2007; McCain, 2009)。不同功能群的动物分布于不同的海拔带, 每种动物占据一定的海拔范围并与其他类群的物种以一定多度共存(Hodkinson, 2005)。神农架自然遗产地植被垂直带谱中, 常绿阔叶林内的常见动物有猕猴(Macaca mulatta)、麝鼩(Crocidura spp.)、喜鹊(Pica pica)、家燕(Hirundo rustica)等。随着海拔升高,一些适应高海拔寒冷气候条件的动物逐渐出现, 如寒温带针叶林巴山冷杉林和高山灌丛和草甸内的主要动物类群为金丝猴(Rhinopithecus roxellana)、秦岭鼠兔(Ochotona huangensis)等兽类以及普通朱雀(Carpodacus spp.)、褐冠山雀(Parus dichrous)、灰头灰雀(Pyrrhula erythaca)等鸟类(李义明等, 2003; 章波, 2014)。
神农架自然遗产地海拔梯度上不同植被带内的土壤动物和微生物的种类和数量, 展示了东方落叶林生物地理省的土壤动物和微生物随海拔变化的生物生态学过程。研究发现, 山地植被垂直带谱的土壤动物和微生物的种类和数量也随海拔梯度变化发生相应的变化(Olson, 1994; Gonzalez et al., 2007; Bahram et al., 2012)。某些特定的微生物群落只分布于一定的海拔范围(Gomez-Hernandez et al., 2012)。土壤白蚁群落的物种组成随海拔梯度变化更替不明显(Palin et al., 2011), 但其凋落物分解者的种类更替随海拔变化较显著, 土壤微生物功能群的生物量和多度随海拔梯度的变化也比较显著(Olson, 1994)。研究发现, 真菌与细菌的生物量比随海拔升高而变大(Wagai et al., 2011), 这主要是因为高海拔环境胁迫压力大, 以真菌为主的微生物群落的土壤养分循环能较好地适应不良的环境条件(Wardle, 2002)。
神农架自然遗产地完整的植被垂直带谱, 充分代表并展示了全球生物地理区划中东方落叶林生物地理省的山地植被垂直带谱的完整性及其孕育的生物生态学过程, 是东方落叶林生物地理省山地生态系统垂直分异生态学过程的天然实验场所。为此,神农架自然遗产地完整的植被垂直带谱也成为神农架自然遗产地遗产价值的重要组成部分。进一步掌握神农架山地植被垂直带谱的生物生态学过程及其驱动机制是今后神农架自然遗产地研究和保护的重要内容, 也是神农架自然遗产地科学管理急需解决的重要问题。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | . 61 Altitudinal gradients strongly affect the diversity of plants and animals, yet little is known about the altitudinal effects on the distribution of microorganisms, including ectomycorrhizal fungi. 61 By combining morphological and molecular identification methods, we addressed the relative effects of altitude, temperature, precipitation, host community and soil nutrient concentrations on species richness and community composition of ectomycorrhizal fungi in one of the last remaining temperate old-growth forests in Eurasia. 61 Molecular analyses revealed 367 species of ectomycorrhizal fungi along three altitudinal transects. Species richness declined monotonically with increasing altitude. Host species and altitude were the main drivers of the ectomycorrhizal fungal community composition at both the local and regional scales. The mean annual temperature and precipitation were strongly correlated with altitude and accounted for the observed patterns of richness and community. 61 The decline of ectomycorrhizal fungal richness with increasing altitude is consistent with the general altitudinal richness patterns of macroorganisms. Low environmental energy reduces the competitive ability of rare species and thus has a negative effect on the richness of ectomycorrhizal fungi. Because of multicollinearity with altitude, the direct effects of climatic variables and their seasonality warrant further investigation at the regional and continental scales. |
| [2] | . |
| [3] | . |
| [4] | . Many studies suggest that global warming is driving species ranges poleward and toward higher elevations at temperate latitudes, but evidence for range shifts is scarce for the tropics, where the shallow latitudinal temperature gradient makes upslope shifts more likely than poleward shifts. Based on new data for plants and insects on an elevational transect in Costa Rica, we assess the potential for lowland biotic attrition, range-shift gaps, and mountaintop extinctions under projected warming. We conclude that tropical lowland biotas may face a level of net lowland biotic attrition without parallel at higher latitudes (where range shifts may be compensated for by species from lower latitudes) and that a high proportion of tropical species soon faces gaps between current and projected elevational ranges. |
| [5] | |
| [6] | |
| [7] | |
| [8] | |
| [9] | |
| [10] | |
| [11] | |
| [12] | |
| [13] | |
| [14] | |
| [15] | . Gradient analysis is rarely used in studies of fungal communities. Data on macromycetes from eight sites along an elevation gradient in central Veracruz, Mexico, were used to demonstrate methods for gradient analysis that can be applied to studies of communities of fungi. Selected sites from 100 to 3,50002m altitude represent tropical dry forest, tropical montane cloud forest, conifer forest, and their ecotones. From May to October 2010, macromycetes were collected monthly within ten 1002×021002m permanent plots per site. In total, 672 individuals of 213 species of macromycetes were recorded. Models for richness and diversity for all macromycete and ectomycorrhizal communities displayed peaks in the mid-part of the gradient, and a tendency to increase with elevation, whereas xylophagous fungi displayed a peak in the mid-lower part but tended to decrease with elevation. Cluster and Maximum Likelihood analyses distinguished four communities for both macromycetes and trees, but plant and fungal communities were only partly concordant. Canonical correspondence analysis indicated that macromycete distribution along the gradient is related to slope, relative humidity, soil temperature, soil water content, canopy openness, and litter depth. Spearman’s correlation and regression trees suggested that air and soil temperature, relative humidity, soil water content, canopy openness, vegetation structure and tree species richness were most strongly related to macrofungal functional groups, but these environmental variables were often correlated to the forest type and may not be causal. Variation in the environment along the elevation gradient differentially affected macromycete functional groups. Results from the different methods used in this work were concordant and showed significant patterns. |
| [16] | . Abstract Thermal pasteurization is a well known and old technique for reducing the microbial count of foods. Traditional thermal processing, however, can destroy heat-sensitive nutrients and food product qualities such as flavor, color and texture. For more than 2 decades now, the use of high-pressure carbon dioxide (HPCD) has been proposed as an alternative cold pasteurization technique for foods. This method presents some fundamental advantages related to the mild conditions employed, particularly because it allows processing at much lower temperature than the ones used in thermal pasteurization. In spite of intensified research efforts the last couple of years, the HPCD preservation technique has not yet been implemented on a large scale by the food industry until now. This review presents a survey of published knowledge concerning the HPCD technique for microbial inactivation, and addresses issues of the technology such as the mechanism of carbon dioxide bactericidal action, the potential for inactivating vegetative cells and bacterial spores, and the regulatory hurdles which need to be overcome. In addition, the review also reflects on the opportunities and especially the current drawbacks of the HPCD technique for the food industry. |
| [17] | . |
| [18] | . . |
| [19] | . Altitude is a determining factor of ecosystem properties and processes in mountains. This study investigated the changes in the concentrations of carbon (C), nitrogen (N), and phosphorus (P) and their ratios in four key ecosystem components (forest floor litter, fine roots, soil, and soil microorganisms) along an altitudinal gradient (from 50 to 950 a.s.l.) in subtropical China. The results showed that soil organic C and microbial biomass C concentrations increased linearly with increasing altitude. Similar trends were observed for concentrations of total soil N and microbial biomass N. In contrast, the N concentration of litter and fine roots decreased linearly with altitude. With increasing altitude, litter, fine roots, and soil C:N ratios increased linearly, while the C:N ratio of soil microbial biomass did not change significantly. Phosphorus concentration and C:P and N:P ratios of all ecosystem components generally had nonlinear relationships with altitude. Our results indicate that the altitudinal pattern of plant and soil nutrient status differs among ecosystem components and that the relative importance of P vs. N limitation for ecosystem functions and processes shifts along altitudinal gradients. |
| [20] | . |
| [21] | . . |
| [22] | . |
| [23] | |
| [24] | . . |
| [25] | . . 于1999~2001年调查了神农架自然保护区6个地点不同栖息地的非飞行哺乳动物的物种丰富度.栖息地分为8类:原始林、择伐林(采伐枯立木)、次生林、灌木林、草地、常年性河流水溪、农田和人居住区.小型非飞行哺乳动物调查用捕鼠夹;大型非飞行哺乳动物调查主要根据皮张收购资料以及样线法和痕迹法;用10 m×10 m的样方调查林地树种丰富度.调查发现,神农架自然保护区有非飞行哺乳动物59种[不包括引进种梅花鹿(Cervus nippon)].在同一海拔高度,原始林通常比择伐林和次生林的物种丰富度高,说明采伐严重降低了物种丰富度.对比同一栖息地不同海拔高度的物种丰富度,我们发现,在中海拔地段(800~1700 m)物种丰富度最高:如在原始林和次生林,海拔1700 m的东溪物种丰富度最高;在择伐林,海拔800 m的九冲物种丰富度最高.聚类分析显示,6个地点的哺乳动物物种组成可以分为两组:高海拔组(2100 m以上)和中低海拔组(1700 m以下).各地点的哺乳动物物种组成与植被的垂直分布是一致的.各地点的物种丰富度与单位面积(100 m2)树种平均丰富度、栖息地类型数和海拔高度相关.3个环境变量间也是相关的:海拔高度对单位面积树种平均丰富度和栖息地类型数有重要影响.根据研究结果提出两点保护建议:第一,保护区的移民迁出和退耕还林工程应首先在物种丰富度最高的九冲进行,而后是东溪和下谷;第二,为了增加个体流和基因流,保护区东西两片相间的非保护区地带应划入保护区,建立栖息地廊道. |
| [26] | |
| [27] | . . 第40届世界遗产大会于2016年7月在土耳其伊斯坦布尔召开,湖北神农架和广西花山岩画被正式列入世界自然和文化遗产,使得我国的世界遗产地达到50处,位列世界第二,与排在第一位的意大利仅差一处。截至目前,全球的世界遗产地已达1,052处,其中自然遗产地203处,文化遗产地814处,双遗产地35处,分布在165个缔约方(http://whc.unesco org/en/list/)。 |
| [28] | . Tropical forests have a major influence on global patterns of biodiversity, ecosystem ecology, productivity and biogeochemical cycles, but they remain relatively understudied. Moreover, our understanding of many global patterns (e.g. of how biodiversity, ecophysiology or ecosystem function vary with latitude) are often influenced by a handful of data points from tropical latitudes (in contrast to swarms of data points from temperate regions). The result is that the wet tropics are often treated as a warm, wet ‘end-point’ of most global analyses. However, comparison of tropical with extra-tropical regions is not straightforward, because of the vast geographical separations involved that lead to complications resulting from both biogeography and climate. The sheer size of the tropical zone implies further that comparison is necessary among tropical biomes, not just between climate zones. |
| [29] | . ABSTRACT Aim68 Elevational gradients distributed across the globe are a powerful test system for understanding biodiversity. Here I use a comprehensive set of bird elevational gradients to test the main drivers of diversity, including sampling, area, mid-domain effect, temperature, temperature and water availability, and hypotheses of evolutionary history. Location68 Seventy-eight elevational gradients of bird diversity from mountains in both hemispheres spanning 24.5°S to 48.2°N, including gradients from various climates, biogeographical regions and habitat types. Methods68 Data on bird elevational diversity were taken from the literature. Of the 150 datasets found or compiled, only those with a high, unbiased sampling effort were used in analyses. Datasets sampled all birds, all breeding birds or all forest birds; a few studies detailed seasonal, elevational shifts. Eighteen predictions of diversity theory were tested, including three sets of interactions. Results68 Birds display four distinct diversity patterns in nearly equal frequency on mountains: decreasing diversity, low-elevation plateaus, low-elevation plateaus with mid-peaks, and unimodal mid-elevational peaks. Bird elevational diversity strongly supports current climate as the main driver of diversity, particularly combined trends in temperature and water availability. Bird diversity on humid mountains is either decreasing or shows a low-elevation plateau in diversity, while on dry mountains it is unimodal or a broad, low-elevation plateau usually with a mid-elevation maximum. The predictions of sampling, area and mid-domain effect were not consistently supported globally. The only evolutionary hypothesis with preliminary support was niche conservatism. Main conclusions68 Both water and temperature variables are needed to comprehensively predict elevational diversity patterns for birds. This result is consistent for breeding and forest birds, for both hemispheres, and for local- or regional-scale montane gradients. More analyses are needed to discern whether the mechanism underlying these relationships is ecological, based on direct physiological limitations or indirect food resource limitations, or historical, based on phylogenetic niche conservation or other evolutionary trends related to climate. The species–area and mid-domain effects are not supported as primary drivers of elevational diversity in birds. |
| [30] | . The floristic and structural differentiation of vegetation along the altitudinal gradient in four subalpine forests of different developmental stages on Mt. Fuji has been studied. Near the forest limit a micropattern of vegetation corresponding to the altitudinal zonation has been observed which elucidated the mechanisms of development of the vegetation zonation.As to early stages of vegetation development only two types can be distinguished: the volcanic desert above 1500 m and the pioneer forests below. As to later stages a differentiation of subzones includes from higher to lower altitudes: the Alnus maximowiczii, Betula ermanii, Abies veitchii and Tsuga diversifolia forests. Larix leptolepis and Sorbus americana ssp. japonica , appear as co-dominants in ecotonal communities between the principal subzones and are also important pioneers in early stages. Similarity analyses reveal that the upper subalpine Alnus-Betula forests can be regarded as early successional phases of the climax Abies-Tsuga forests of the lower subalpine zone.The regular arrangement of A. maximowiczii-B. ermanii-A. veitchii is studied along the gradient from the margin to the interior of the forest growing near the forest limit where locally favourable conditions prevail. Growth form, height growth, photosynthetic activity, seed supply, and seedling distribution of the three principal species have been compared, as well as biomass and production relations in contiguous forests of these species. The marginal Alnus type community is productive and disturbance-tolerant, and has a wide ecological and sociological amplitude along the gradient, while the central Abies community is accumulative and disturbance-intolerant, and has a narrower tolerance range, but is superior in competition under stable habitat conditions. A vegetation organization, emporal multi-storeyed structure , is suggested which means that a zonal pattern of vegetation within a climax region develops by successive replacement of successional species along an environmental gradient. |
| [31] | . |
| [32] | . Through their role as 090004ecosystem engineers090005, termites provide a range of ecosystem services including decomposition, and carbon and nitrogen cycling. Although termite diversity levels differ between regions as a result of variation in regional species pool size, in general, termite diversity is thought to decline with elevation. This study (1) investigated how termite species density, abundance, functional group diversity and termite attack on dead wood vary with altitude along an Amazon090009Andes altitudinal gradient in Peru; (2) identified likely environmental causes of this pattern; and (3) explored the implications of termite presence for ecosystem functioning (notably for decomposition). Termites were sampled with a standardized 100 0103 2 m straight-belt transect at five undisturbed forest sites along a gradient 190 to 3025 m, as were environmental variables and termite and fungus attack on dead wood. Termite diversity was similar to that found at comparable sites in South America, and there was little turnover of assemblage composition with elevation suggesting that montane specialists are not present. Termite diversity declined with increased elevation, though the upper distribution limit for termites was at a lower elevation than anticipated. We suggest that key drivers of this elevation pattern are reduced temperature with altitude and mid-elevation peaks in soil water content. Also, attack on dead wood diminished with decreasing termite indirect absolute abundance, while the depth of the soil humic layer increased. We hypothesize that termite abundance is a major accelerant of decomposition rates (and associated mineralization) in Amazonian forests.Abstract in Spanish is available at http://www.blackwell-synergy.com/loi/btp. |
| [33] | |
| [34] | . |
| [35] | . We measured aboveground plant biomass, aboveground net primary productivity (ANPP), detritus accumulation, and nitrogen and phosphorus uptake by above-ground vegetation in six Metrosideros polymorpha stands on the windward slopes of Mauna Loa, Hawai`i, USA. Our objective was to quantify the effects of elevation (primarily temperature) on ecosystem properties during primary succession, as a key to understanding ecosystem-climate interactions. Four study sites were on 111- to 136-yr-old lava flows at elevations of 290, 700, 1130, and 1660 m. Two additional sites on 3400-yr-old lava were at 700 and 1660 m elevations. All sites were on solid pahoehoe (smooth or ropy-textured) lava substrates with gentle relief, were free of significant human disturbance, received abundant precipitation, and had similar vegetation composition. Total aboveground biomass, soil organic matter mass, and aboveground net primary production (ANPP) were all greater in the old sites than in young sites. Differences between young and old sites in aboveground live biomass, detrital mass, and ANPP all supported the conclusion that ecosystem development proceeded relatively faster at 700 m elevation than at 1660 m. However, above-ground biomass in the old sites (81 Mg/ha at 1660 m elevation and 123 Mg/ha at 700 m) was low in comparison with other wet tropical forests. Accumulations of N and P in live biomass and detritus followed the same trends as were observed for organic matter. Rates of soil carbon accumulation over the first 3400 yr of succession averaged 2.1 g.m-2. yr-1, similar to other reported soil chronosequences. Observed rates of N accumulation ranged from 0.1 to 0.6 g.m-2. yr-1 over the first 136 yr of succession. There were no monotonic elevational trends among young sites with respect to live biomass, detritus mass, or total N or P accumulation. Foliar nitrogen concentrations in the young sites were among the lowest reported from any tropical forests and tended to decline with increasing elevation. The growth and biomass of individual plant species varied in distinctive ways along the elevational gradient. Nevertheless, among young sites there was a direct, linear relationship between total ANPP and mean annual temperature of the site, with a similar pattern in the two old sites. For each 1 increase in mean annual temperature, total ANPP increased by 54 g.m-2. yr-1. Community-level ANPP also was directly correlated with rates of N and P uptake by the vegetation, regardless of site age or elevation. |
| [36] | . 61 We present the results from a litter translocation experiment along a 2800-m elevation gradient in Peruvian tropical forests. The understanding of the environmental factors controlling litter decomposition is important in the description of the carbon and nutrient cycles of tropical ecosystems, and in predicting their response to long-term increases in temperature. 61 Samples of litter from 15 species were transplanted across all five sites in the study, and decomposition was tracked over 448 d. 61 Species' type had a large influence on the decomposition rate (k), most probably through its influence on leaf quality and morphology. When samples were pooled across species and elevations, soil temperature explained 95% of the variation in the decomposition rate, but no direct relationship was observed with either soil moisture or rainfall. The sensitivity of the decay rate to temperature (κ(T)) varied seven-fold across species, between 0.024 and 0.169 °C6301, with a mean value of 0.118 ± 0.009 °C6301 (SE). This is equivalent to a temperature sensitivity parameter (Q6968) for litter decay of 3.06 ± 0.28, higher than that frequently assumed for heterotrophic processes. 61 Our results suggest that the warming of approx. 0.9 °C experienced in the region in recent decades may have increased decomposition and nutrient mineralization rates by c. 10%. |
| [37] | . ABSTRACT Aim68 This research aims to understand the factors that shape elevational diversity gradients and how those factors vary with spatial grain. Specifically, we test the predictions of the species–productivity hypothesis, species–temperature hypothesis, the metabolic theory of ecology and the mid-domain effects null model. We also examine how the effects of productivity and temperature on richness depend on spatial grain. Location68 Deciduous forests along an elevational gradient in Great Smoky Mountains National Park, USA. Methods68 We sampled 22 leaf litter ant assemblages at three spatial grains, from 1-m 2 quadrats to 50 × 50 m plots using Winkler samplers. Results68 Across spatial grains, warmer sites had more species than did cooler sites, and primary productivity did not predict ant species richness. We found some support for the predictions of the metabolic theory of ecology, but no support for the mid-domain effects null model. Thus, our data are best explained by some version of a species–temperature hypothesis. Main conclusions68 Our results suggest that temperature indirectly affects ant species diversity across spatial grains, perhaps by limiting access to resources. Warmer sites support more species because they support more individuals, thereby reducing the probability of local extinction. Many of our results from this elevational gradient agree with studies at more global scales, suggesting that some mechanisms shaping ant diversity gradients are common across scales. |
| [38] | . Community structure and ecosystem processes often vary along elevational gradients. Their responses to elevation are commonly driven by changes in temperature, and many community- and ecosystem-level variables therefore frequently respond similarly to elevation across contrasting gradients. There are also many exceptions, sometimes because other factors such as precipitation can also vary with elevation. Given this complexity, our capacity to predict when and why the same variable responds differently among disparate elevational gradients is often limited. Furthermore, there is utility in using elevational gradients for understanding community and ecosystem responses to global climate change at much larger spatial and temporal scales than is possible through conventional ecological experiments. However, future studies that integrate elevational gradient approaches with experimental manipulations will provide powerful information that can improve predictions of climate change impacts within and across ecosystems. |
| [39] | . . |
| [40] | . |
| [41] | . . 神农架位于湖北省西部,长江以北、汉水以南的广阔地带,属北亚热 带向暖温带的过渡区域。本文依据该地区所处地理位置的植被分布规律等资料,绘制了1:20万的植被复原图。并在此基础上,运用 ERDASimagine8.4和MapInfoprofessional6.0软件,分别对神农架地区的TM影像(5、4、3波段)进行监督分类及目视 解译,同时结合野外的样方调查,绘制了神农架地区1:20万的植被类型图,并建立了相应的属性数据库。最后,根据野外的GPS定位点对制图精度进行了 Kappa检验。 制图结果表明,制图区总面积3476.67km2,共计504个斑块。据统计,林地面积2607.45km2,森林覆盖率75%;山地... |
| [42] | |
| [43] | . Soils and plants were sampled along an elevational gradient from 265–1675 m on a 133-and a 3100-year-old lava flow on Mauna Loa, Hawai'i. Soil organic matter and nutrients accumulated more rapidly at low elevation on the young flow, but reached higher levels at higher elevation on the old flow. Foliar nitrogen and phosphorus concentrations were less and specific leaf weight greater for Metrosideros polymorpha leaves collected at high versus low elevations and on the young versus the old flow. Foliar δ 13 C was strongly correlated with specific leaf weight across the range of sites sampled. |
| [44] | . Climate and parent material strongly control vegetation structure and function, yet their control over the belowground microbial community is poorly understood. We assessed variation in microbial lipid profiles in undisturbed forest soils (organic and surface mineral horizons) along an altitudinal gradient (700, 1,700, and 2,70002m a.s.l. mean annual temperature of 12–24°C) on two contrasting parent materials (acidic metasedimentary vs. ultrabasic igneous rock) in Mt. Kinabalu, Borneo. Soil organic carbon and nitrogen concentrations were generally higher at higher altitudes and, within a site, at upper soil horizons. Soil pH ranged from 3.9 to 5.3, with higher values for the ultrabasic soils especially at higher altitudes. The major shifts in microbial community structure observed were the decline in the ratio of fungal to bacterial lipid markers both with increasing soil depth and decreasing altitude. The positive correlation between this ratio with soil C and N concentrations suggested a strong substrate control in accord with the literature from mid to high-latitude ecosystems. Principal component analysis using seven groups of signature lipids suggested a significant altitude by parent material interaction—the significant difference in microbial community structure between the two rock types found at 2,700-m sites developed on weakly weathered soils diminished with decreasing altitude towards 700-m sites where soils were strongly weathered. These results are consistent with the hypothesis that parent material effect on soil microbial community (either directly via soil geochemistry or indirectly via floristic composition) is stronger at an earlier stage of ecosystem development. |
| [45] | |
| [46] | . . |
| [47] | |
| [48] | . |
| [49] | . |
| [50] | . . |
| [51] | |
| [52] | . (in Chinese with English abstract) . 神农架国家级自然保护区,地处湖北省的西北部。它有着优越的自然环境和气候条件,再加上其特殊的地理位置,使得这里的生物多样性非常丰富,备受****们的关注。但自从保护区成立后近几十年,关于神农架保护区内的野生动物资源状况,尤其是保护区内的鸟类资源,还没有一次较全面的调查统计。另外,对整个神农架保护区的鸟类多样性研究还尚未见报道。所以通过“神农架本底资源综合调查”这一项目,我们着手调查整个神农架保护区的鸟类资源状况,获取其鸟类多样性状况的第一手资料。从而为保护区内的鸟类种群变化等研究,以及保护区的发展建设提供有力的支持和依据。 笔者分别于2011年7月-8月、2011年10月、2012年1月、2012年3-5月、2012年7月-8月、2013年3月和2013年6月-7月,采用样线法和样点法对神农架鸟类资源进行实地调查,并结合访问调查和文献查阅,结果如下: 神农架保护区共有鸟类389种,隶属于18目67科。其中留鸟所占比例最大,为45.244%,夏候鸟为25.193%,冬候鸟为18.252%,旅鸟为11.311%;在繁殖鸟类(274种)中,东洋界种185种,占67.518%,古北界种60种,占21.898%,广布种29种,占10.584%。神农架保护区389种鸟类中,有国家一级保护鸟类5种,国家二级保护鸟类65种;中国特有种鸟类26种;湖北省保护鸟类120种;被列于“三有”名录的鸟类共233种;被收录于《中国濒危动物红皮书》的共34种;被列入CITES附录的鸟类共计62种;被列入《中华人民共和国政府和日本国政府保护候鸟及其栖息环境的协定》的鸟类共111种,被列入《中华人民共和国政府和澳大利亚政府保护候鸟及其栖息环境的协定》的鸟类共22种。 此次调查统计发现神农架地区鸟类新记录73种,其中有13种为湖北省鸟类新纪录,它们分别是:乌灰鹞(Circus pypargus)、棕尾鵟(Buteo rufinus)、灰腹角雉(Tragopan blythii)、黑腹燕鸥(Sterna acuticauda)、毛脚渔鴞(Ketupa blakistoni)、花头鸺鶹(Glaucidium passerinum)、赤红山椒鸟(Pericrocotus flammenss)、纹喉鹎(Pycnonotus finlaysoni)、栗背伯劳(Lanius collurioides)、灰树鹊(Dendrocitta vagabunda)、蓝翅希鹛(Minla cyanouroptera)、斑胸鸦雀(Paradoxornis flavirostris)、大树莺(Cettia major)。 神农架自然保护区的G指数为4.910,F指数为35.621,故G-F指数为:0.862。四个季节的G-F多样性指数由大到小依次是:夏季(0.777)、秋季(0.697)、春季(0.600)和冬季(0.202)。 采用频率指数法估算不同季节的鸟类优势种,根据RB值计算可知:春季中优势种鸟类有9种,如:白鹊鸰(Motacilla alba)、橙翅噪鹛(Garrulax elliotii)矛纹草鹛(Babax lanceolatus)等等;夏季中优势种鸟类有18种,如是:白鹊鸽、红尾水鸲(Rhyacornis fuliginosa)、领雀嘴鹎(Spizixos semitorques)等等;秋季中优势种鸟类有13种,分别是:燕雀(Fringilla montifringilla)、白鹊鸰、领雀嘴鹎等等;冬季中优势种鸟类有11种,如:小鸊鷉(Tachybaprtus ruficollis)、松鸦(Garrulus glandarius)、雉鸡(Phasianus colchicus)等等。 在四个季节中,鸟类物种丰富度由高到低依次是夏季(148种)、秋季(77种)、春季(72种)、冬季(28种)。六种生境的鸟类群落物种丰富度依次为:阔叶林(146种)、针阔混交林(69种)、水域湿地(45种)、农田村落(42种)、灌丛竹林草地(19种)、针叶林(8种)。 分别用Pielou指数和Alatalo指数分析鸟类群落的均匀性。就四个季节的鸟类群落均匀性而言,Pielou指数和Alatalo指数均显示——冬季、夏季和秋季的鸟类群落均匀性依次递减,而对春季的均匀性,Pielou指数测得春季的鸟类群落均匀性最高,Alatalo指数测得春季的鸟类群落均匀性位居第三;就六种生境的鸟类群落均匀性而言,二种均匀性指数均测得灌丛竹林草地、水域湿地、农田村落、针叶林的均匀性依次递减,但对于针阔混交林和阔叶林的均匀性二种指数的测量结果有差异。 对不同鸟类群落的多样性,采用Shannon-Wiener指数和辛普森指数2种多样性指数来分析。就四个季节的鸟类群落多样性而言,Shannon-Wiener指数和辛普森指数均显示——夏季、春季和冬季的鸟类群落多样性依次递减,但对秋季的多样性,辛普森指数测得秋季的鸟类群落多样性在四个季节中最小,而Shannon-Wiener指数测得秋季的鸟类群落多样性四个季节中排倒数第二;就六种生境的鸟类群落多样性而言,Shannon-Wiener指数和辛普森指数均显示阔叶林、针阔混交林、水域湿地、农田村落、针叶林的多样性依次递减,但对灌丛竹林草地的鸟类群落多样性,Shannon-Wiener指数测得其多样性居于第五位,辛普森指数测得其多样性居于第四位。 由于鸟类群落多样性与其物种丰富度和均匀性是密切相关的,故用Shannon-Wiener指数和辛普森指数分别与丰富度、Pielou指数和Alatalo指数做相关性分析,相关性分析表明:①就本实验而言,Pielou均匀性指数和Alatalo均匀性指数中前者更适合;②Shannon-Wiener指数和辛普森指数在本实验中均有一定的适合度,二者没有显著的区分。 分别利用Sorenson指数和Bray-Curtis指数分析不同鸟类群落间相似性,两种相似性系数均测得阔叶林鸟类群落与针阔混交林鸟类群落相似性最高。 本研究从鸟类物种数、鸟类数量、多样性以及均匀性四个方面证明了:神农架鸟类垂直海拔分布符合“中域效应”假说。 |
| [53] |
1
2012
... 神农架自然遗产地海拔梯度上不同植被带内的土壤动物和微生物的种类和数量, 展示了东方落叶林生物地理省的土壤动物和微生物随海拔变化的生物生态学过程.研究发现, 山地植被垂直带谱的土壤动物和微生物的种类和数量也随海拔梯度变化发生相应的变化(
Global distribution of species diversity in vascular Plants: Towards a world map of phytodiversity
1
1996
... 山地是指具有一定海拔高度、相对高度和坡度的地面(
Climatic change in mountain regions: A review of possible impacts
1
2003
... 山地是指具有一定海拔高度、相对高度和坡度的地面(
Global warming, elevational range shifts, and lowland biotic attrition in the wet tropics
1
2008
... 山地是指具有一定海拔高度、相对高度和坡度的地面(
1
1990
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(
1
1983
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(
1
1989
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(
1
1990
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(
1
1991
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(
1
1983
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(
1
1980
... 中国是一个山地大国, 国土面积的2/3为山地(
1
1989
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(
1
1983
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(
1
2007
... 随着海拔升高, 山地气温、气压和地表面积下降, 山地环境条件发生显著变化(
Patterns of macromycete community assemblage along an elevation gradient: Options for fungal gradient and metacommunity analyses
1
2012
... 神农架自然遗产地海拔梯度上不同植被带内的土壤动物和微生物的种类和数量, 展示了东方落叶林生物地理省的土壤动物和微生物随海拔变化的生物生态学过程.研究发现, 山地植被垂直带谱的土壤动物和微生物的种类和数量也随海拔梯度变化发生相应的变化(
High pressure carbon dioxide inactivation of microorganisms in foods: The past, the present and the future
1
2007
... 神农架自然遗产地海拔梯度上不同植被带内的土壤动物和微生物的种类和数量, 展示了东方落叶林生物地理省的土壤动物和微生物随海拔变化的生物生态学过程.研究发现, 山地植被垂直带谱的土壤动物和微生物的种类和数量也随海拔梯度变化发生相应的变化(
Geography and evolution
1
1924
... 山地是指具有一定海拔高度、相对高度和坡度的地面(
日本的森林植被
4
2006
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(
... 受副热带高压的控制, 全球30°-35° N大部分地段主要植被类型为疏林、荒漠植被(
... Information in the table is from UNESCO official website (http://whc.unesco.org) and related published literature, e.g.,
... 表中资料参考: UNESCO官方网站(http://whc.unesco.org)数据库以及相关文献, 如
Altitudinal patterns and controls of plant and soil nutrient concentrations and stoichiometry in subtropical China
2
2016
... 山地是指具有一定海拔高度、相对高度和坡度的地面(
... 随着海拔升高, 山地气温、气压和地表面积下降, 山地环境条件发生显著变化(
Terrestrial insects along elevation gradients: Species and community responses to altitude
2
2005
... 神农架自然遗产地海拔梯度上不同植被带内的动物类群, 展示了东方落叶林生物地理省不同功能动物类群随海拔梯度变化的更替过程.已有研究发现, 海拔垂直梯度上动物群落主要通过改变其多度及物种更替适应海拔梯度上的不同环境条件(
... ).不同功能群的动物分布于不同的海拔带, 每种动物占据一定的海拔范围并与其他类群的物种以一定多度共存(
黄山的植被
1
1982
... 比较分析神农架自然遗产地植被垂直带谱与东方落叶林生物地理省内世界自然遗产地及预备世界自然遗产地的植被垂直带谱(
The use of “altitude” in ecological research
2
2007
... 山地是指具有一定海拔高度、相对高度和坡度的地面(
... 随着海拔升高, 山地气温、气压和地表面积下降, 山地环境条件发生显著变化(
1
1999
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(
对陕西秦岭西段南坡植被垂直带划分问题的一点认识
1
1985
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(
神农架自然保护区非飞行哺乳动物的物种丰富度: 沿海拔梯度的分布格局
1
2003
... 神农架自然遗产地海拔梯度上不同植被带内的动物类群, 展示了东方落叶林生物地理省不同功能动物类群随海拔梯度变化的更替过程.已有研究发现, 海拔垂直梯度上动物群落主要通过改变其多度及物种更替适应海拔梯度上的不同环境条件(
1
2015
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(
世界自然遗产既要加强保护也要适度利用
1
2016
... 中国是一个山地大国, 国土面积的2/3为山地(
Introduction: Elevation gradients in the tropics: Laboratories for ecosystem ecology and global change research
1
2010
... 随着海拔升高, 山地气温、气压和地表面积下降, 山地环境条件发生显著变化(
Global analysis of bird elevational diversity
1
2009
... 神农架自然遗产地海拔梯度上不同植被带内的动物类群, 展示了东方落叶林生物地理省不同功能动物类群随海拔梯度变化的更替过程.已有研究发现, 海拔垂直梯度上动物群落主要通过改变其多度及物种更替适应海拔梯度上的不同环境条件(
Differentiation of vegetation zones and species strategies in the subalpine region of Mt. Fuji
1
1984
... 比较分析神农架自然遗产地植被垂直带谱与东方落叶林生物地理省内世界自然遗产地及预备世界自然遗产地的植被垂直带谱(
The distribution of leaf litter invertebrates along a Neotropical altitudinal gradient
2
1994
... 神农架自然遗产地海拔梯度上不同植被带内的土壤动物和微生物的种类和数量, 展示了东方落叶林生物地理省的土壤动物和微生物随海拔变化的生物生态学过程.研究发现, 山地植被垂直带谱的土壤动物和微生物的种类和数量也随海拔梯度变化发生相应的变化(
... ), 但其凋落物分解者的种类更替随海拔变化较显著, 土壤微生物功能群的生物量和多度随海拔梯度的变化也比较显著(
Termite diversity along an Amazon- Andes elevation gradient, Peru
1
2011
... 神农架自然遗产地海拔梯度上不同植被带内的土壤动物和微生物的种类和数量, 展示了东方落叶林生物地理省的土壤动物和微生物随海拔变化的生物生态学过程.研究发现, 山地植被垂直带谱的土壤动物和微生物的种类和数量也随海拔梯度变化发生相应的变化(
1
1990
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(
The role of spatial scale and the perception of large-scale species-richness patterns
2
2005
... 山地是指具有一定海拔高度、相对高度和坡度的地面(
... ;
Primary productivity and ecosystem development along an elevational gradient on Mauna Loa, Hawaii
1
1997
... 山地是指具有一定海拔高度、相对高度和坡度的地面(
The sensitivity of tropical leaf litter decomposition to temperature: Results from a large-scale leaf translocation experiment along an elevation gradient in Peruvian forests
1
2011
... 神农架自然遗产地海拔梯度上完好的植被垂直带谱, 展示了全球生物地理区划东方落叶林生物地理省的植物群落优势种和功能群随海拔梯度的更替过程.研究发现: 山地海拔梯度上的植被垂直带谱展示了植物群落优势种和功能群随海拔梯度变化的更替过程(
Temperature, but not productivity or geometry, predicts elevational diversity gradients in ants across spatial grains
1
2007
... 神农架自然遗产地海拔梯度上不同植被带内的动物类群, 展示了东方落叶林生物地理省不同功能动物类群随海拔梯度变化的更替过程.已有研究发现, 海拔垂直梯度上动物群落主要通过改变其多度及物种更替适应海拔梯度上的不同环境条件(
Community and ecosystem responses to elevational gradients: Processes, mechanisms, and insights for global change
2
2013
... 山地是指具有一定海拔高度、相对高度和坡度的地面(
... 神农架自然遗产地海拔梯度上完好的植被垂直带谱, 展示了全球生物地理区划东方落叶林生物地理省的植物群落优势种和功能群随海拔梯度的更替过程.研究发现: 山地海拔梯度上的植被垂直带谱展示了植物群落优势种和功能群随海拔梯度变化的更替过程(
湖北植被区划
1
1982
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(
Experimental investigation of nutrient limitation of forest growth on wet tropical mountains
1
1998
... 神农架自然遗产地海拔梯度上完好的植被垂直带谱, 展示了全球生物地理区划东方落叶林生物地理省的植物群落优势种和功能群随海拔梯度的更替过程.研究发现: 山地海拔梯度上的植被垂直带谱展示了植物群落优势种和功能群随海拔梯度变化的更替过程(
神农架的植被及其1:200, 000植被图的编制
2
2002
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(
... The distribution of main plant formations in Shennongjia along the elevation gradient. Capital English letters represent vertical vegetation zones: A, subtropical zone of evergreen broad-leaved forest; B, north subtropical zone of Mixed evergreen and deciduous broad-leaved forest; C, warm temperate zone of broadleaved deciduous forest; D, temperate zone of mixed broadleaf-conifer forest; E, cold temperate zone of coniferous forest; F, subalpine zone of shrub meadow. The abscissa represents different formations: 1, Form. Chimonanthus praecox; 2, Form. Phoebe zhennan, Cyclobalanopsis gracilis; 3, Form. Coriaria sinica, Cotinus coggyqria var. pubescens; 4, Form. Pinus massoniana, Quercus variabilis; 5, Form. Cunninghamia lanceolata; 6, Form. Quercus variabilis; 7, Form. Lindera communis, Cyclobalanopsis gracilis, Platycarya strobilacea, Betula luminifera; 8, Form. Pinus massoniana; 9, Form. Quercus acrodonta; 10, Form. Cyclobalanopsis oxyodon, Sycopsis chinense, Quercus engleriana, Cyclobanopsis myrsinaefolia; 11, Form. Quercus philyraeoides, Q. acrodonta, Carpinus sp., Platycarya strobilacea; 12, Form. Juglans cathayensis; 13, Form. Quercus variabilis, Q. aliena var. acuteserrata, Castanea segyinii; 14, Form. Quercus engleriana, Cyclobalanopsis oxyodon, Betula luminifera, Platycarya strobilacea; 15, Form. Quercus spinosa; 16, Form. Quercus glandulifera var. brevipetiolata; 17, Form. Castanea seguinii; 18, Form. Pinus henryi; 19, Form. Betula luminifera, Platycarya strobilacea, Carpinus sp.; 20, Form. Pinups armandii, Betula utilis; 21, Form. Quercus aliena var. acuteserrata; 22, Form. Abies chensiensis; 23, Form. Corylus heterophylla var. sutchuenensis, Viburnum opulus var. calvescens, Malus hupehensis; 24, Form. Carex sp., Sanguisorba officinalis var. longifolia, Anaphalis sinica, Geranium henryi; 25, Form. Rhus verniciflua, Quercus acutidentata, Cornus controversa, Platycarya strobilacea; 26, Form. Miscanthus sinensis, Pteridium aquilinum var. latiusculum; 27, Form. Lespedeza formosa, L. buergeri; 28, Form. Carex sp., Juncus concinus, Sanguisorba officicnalis var. longifolia, Chamaenerion angustifolium; 29, Form. Pinus armandii, Querus aliena var. acuteserrata; 30, Form. Pinus armandii; 31, Form. Fagus engleriana; 32, Form. Abies chensiensis, Picea wilsonii; 33, Form. Quercus aliena var. acuteserrata, Fagus engleriana, Betula albosinensis; 34, Form. Betula albosinensis; 35, Form. Pinus armandii, Populus davidiana, Betula albosinensis; 36, Form. Pinus armandii, Populus davidiana; 37, Form. Salix carprea var. inica, Crataegus wilsonii, Sorbus hupehensis; 38, Form. Abies fargesii, Betula albosinensis, Acer sp.; 39, Form. Salix cupularis; 40, Form. Berberis dasystachya; 41, Form. Sinarundinaria nitida; 42, Form. Cotoneaster horizontalis; 43, Form. Abies fargesii; 44, Form. Rhododendron fargesii; 45, Form. Sabina pingii var. wilsonii; 46, Form. Trisetum clarkei, Festuca rubra, Deyeuxia scabrescens. Formation distribution along the elevation was based on
2
... 世界自然遗产的遴选有4条标准, 即美学、地质、生物生态学和栖息地标准.世界自然遗产地全球突出普遍价值(outstanding universal value)的提炼, 是世界自然遗产地遴选及保护的关键.全球生物地理区划(Udvardy)为国际自然保护联盟(IUCN)和联合国教科文组织(UNESCO)世界遗产中心遴选世界自然遗产地的重要科学依据.依据全球自然地理环境特征和生物群落类型, Udvardy把全球地表划分为8个生物地理区(biogeographical realm)、14个生物群区(biome)和193个生物地理省(biogeographical province)(
... 神农架自然遗产地海拔梯度上完好的植被垂直带谱, 展示了全球生物地理区划东方落叶林生物地理省的植物群落优势种和功能群随海拔梯度的更替过程.研究发现: 山地海拔梯度上的植被垂直带谱展示了植物群落优势种和功能群随海拔梯度变化的更替过程(
Elevational and age gradients in Hawaiian montane rainforest: Foliar and soil nutrients
1
1988
... 神农架自然遗产地海拔梯度上不同植被带内的土壤动物和微生物的种类和数量, 展示了东方落叶林生物地理省的土壤动物和微生物随海拔变化的生物生态学过程.研究发现, 山地植被垂直带谱的土壤动物和微生物的种类和数量也随海拔梯度变化发生相应的变化(
Interactive influences of climate and parent material on soil microbial community structure in Bornean tropical forest ecosystems
2011
2
1984
... 山地是指具有一定海拔高度、相对高度和坡度的地面(
... 受副热带高压的控制, 全球30°-35° N大部分地段主要植被类型为疏林、荒漠植被(
中国的主要山脉和山峰
2004
4
... 山地是指具有一定海拔高度、相对高度和坡度的地面(
... 受副热带高压的控制, 全球30°-35° N大部分地段主要植被类型为疏林、荒漠植被(
... 神农架自然遗产地海拔梯度上完好的植被垂直带谱, 展示了全球生物地理区划东方落叶林生物地理省的植物群落优势种和功能群随海拔梯度的更替过程.研究发现: 山地海拔梯度上的植被垂直带谱展示了植物群落优势种和功能群随海拔梯度变化的更替过程(
... 神农架自然遗产地海拔梯度上不同植被带内的土壤动物和微生物的种类和数量, 展示了东方落叶林生物地理省的土壤动物和微生物随海拔变化的生物生态学过程.研究发现, 山地植被垂直带谱的土壤动物和微生物的种类和数量也随海拔梯度变化发生相应的变化(
Vegetation of the Great Smokey Mountains
1
1956
... 山地是指具有一定海拔高度、相对高度和坡度的地面(
Vegetation of the Siskiyou Mountains, Oregon and California
1
1960
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(
江西省三清山的植被类型及其分布
1988
2
2011
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(
... 神农架自然遗产地海拔梯度上不同植被带内的动物类群, 展示了东方落叶林生物地理省不同功能动物类群随海拔梯度变化的更替过程.已有研究发现, 海拔垂直梯度上动物群落主要通过改变其多度及物种更替适应海拔梯度上的不同环境条件(
神农架国家级自然保护区鸟类群落多样性研究
2014
1
1999
... 基于全球1 km的土地利用数据(AVHRR_1 km_ LANDCOVER_1981_1994.GLOBAL), 选取与神农架自然遗产地同纬度的30°-35° N带, 提取30°-35° N带土地利用类型中的常绿落叶阔叶林信息.同时, 收集整理《中国植物地理》(


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}