 , 杨万勤
, 杨万勤Edge effects of forest gap in Pinus massoniana plantations on the ecological stoichiometry of Cinnamomum longepaniculatum
SONGSi-Meng, YANGWan-Qin通讯作者:
版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
展开
摘要
关键词:
Abstract
Keywords:
-->0
PDF (472KB)元数据多维度评价相关文章收藏文章
本文引用格式导出EndNoteRisBibtex收藏本文-->
林窗作为森林中普遍存在的一种重要的干扰方式(Dupuy & Chazdon, 2008), 在微地形、小气候及土壤理化性质等方面有着异于林下的特殊生境(宋小艳等, 2014)。同时, 在林窗与森林结合部位构成林窗边缘区(Restrepo et al., 1999)。由于能量流及物质流会沿林外—林缘—林内的生态环境产生梯度变化(He et al., 2015), 边缘区形成了组成物种的结构、配置和动态等多方面特殊的环境异质性, 即边缘效应(Kneeshaw & Bergeron, 1998; Yamamoto, 2000; 田超等, 2011)。林窗面积是衡量林窗干扰强度的重要指标(Gálhidy et al., 2006), 不同大小林窗边缘影响的范围和程度不同(Goldblum & Beatty, 1999), 影响着整个林分动态、物种多样性更新与维持(崔宁洁等, 2014)及系统养分循环过程(张艳等, 2015)。因此, 研究边缘效应对森林生态系统管理具有重要的意义。
生态化学计量学(ecological stoichiometry)是研究生物系统能量平衡和多重化学元素平衡(特别是碳(C)、氮(N)、磷(P))的一门科学(Elser et al., 2000)。植物结构性元素C和限制性元素N、P耦合形成的分配格局(如C:N、N:P)影响着群落养分利用效率及生产力等(Elser et al., 2010; Yang & Wang, 2011)。林窗边缘生态过渡带迥异的水热环境可影响植被更新、物种侵入与定居, 造成边缘种对土壤养分的激烈争夺, 加速地上和地下养分周转速率(王家华和李建东, 2006; Kern et al., 2013); 同时, 边缘区适宜的温湿度可提高土壤酶活性(Arunachalam & Arunachalam, 2000), 促进土壤动物微生物繁殖及对凋落物中有机质的矿化分解(Gallo et al., 2009; 张艳等, 2015), 为边缘种提供充足的有机质和养分基础, 这些都可能影响林窗边缘的植物化学计量特征。
马尾松(Pinus massoniana)是我国长江上游低山丘陵区退耕还林的主要树种, 面积已超过200万hm2 (崔宁洁等, 2014), 但长期采取的纯林经营模式致使马尾松林地出现生产力下降、地力衰退等多种生态问题(张明锦等, 2016)。通过林窗干扰构建复合林分是改造低效人工林的重要技术手段(Dupuy & Chazdon, 2008)。马尾松-油樟(Cinnamomum longepaniculatum)是四川省宜宾地区的一种混交类型, 油樟是当地一种珍贵的乡土树种, 人为引入在林窗斑块中形成优势种, 构建针叶人工林-阔叶乡土树种镶嵌分布的景观格局。目前, 对天然林或次生林林窗及其边缘效应的研究较多, 如林窗形成时间及方位(王家华和李建东, 2006; Dee et al., 2014)、林窗对物种更新的影响等(廉振民和于广志, 2000; Daoliveira & Ribas, 2011; Haghverdi et al., 2012; Kern et al., 2013), 而对人工林林窗研究报道还相对较少。与天然林相比, 人工林林窗干扰导致的环境异质性及其引起的林下物种元素动态平衡变异性可能更为复杂(王晶苑等, 2011), 可能在养分管理方面存在明显的林窗边缘效应。因此, 本研究在四川宜宾市高县低山丘陵区马尾松人工林7种不同面积的人工林窗边缘, 选取人工更新油樟为研究对象, 研究不同大小林窗边缘效应对油樟化学计量特征的影响, 拟回答以下科学问题: (1)马尾松人工林林窗更新植物油樟在化学计量特征水平上是否存在边缘效应? (2)如果存在边缘效应, 多大林窗面积的边缘效应更加显著, 养分分配格局最优? (3)如果存在边缘效应, 其边缘效应的季节动态如何?
1 研究地区和研究方法
1.1 研究区概况
研究区位于四川省宜宾市高县来复镇(28.60°- 28.61° N, 104.56°-104.57° E, 海拔412-567 m), 地处长江上游和四川盆地南缘, 属中亚热带湿润季风气候。全年日照时间1 148 h, 年平均气温18.1 ℃, 年降水量1 021.8 mm, 雨量充沛, 雨热同季。样地所在的毛巓坳工区经营小班研究阶段降水主要集中在2015年的5-9月(图1)。土壤为酸性山地黄壤, 发育完善, 土层厚度约50 cm。原生植被为亚热带常绿阔叶林, 现已基本垦殖为马尾松人工林, 林分结构简单,郁闭度0.7左右, 林下灌草覆盖度40%, 主要灌木有梨叶悬钩子(Rubus pirifolius)、枹栎(Quercus serrata)、铁仔(Myrsine africana)、茶荚蒾(Viburnum setigerum)等; 主要草本植物有芒(Miscanthus sinensis)、芒萁(Dicranopteris dichotoma)、皱叶狗尾草(Setaria plicata)、淡竹叶(Lophatherum gracile)、荩草(Arthraxon hispidus)等。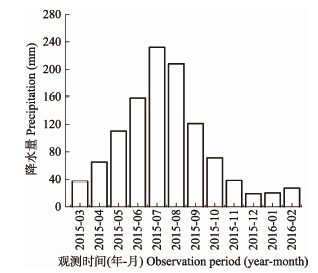 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1马尾松人工林不同观测期的降水量。
-->Fig. 1Precipitation during different observation periods in Pinus massoniana plantations.
-->
1.2 样地设置
2011年, 根据前期样地调查, 在研究区内选择地形地貌、海拔、母岩、土壤类型、坡度、坡位等相同或相近, 林地情况相近、林分密度相似的39年生成熟马尾松人工林作为研究样地, 人工砍伐形成面积分别为100 m2 (G1)、225 m2 (G2)、400 m2 (G3)、625 m2 (G4)、900 m2 (G5)、1 225 m2 (G6)和1 600 m2 (G7)的林窗系列, 形状均近正方形, 各林窗林缘间距不小于20 m。仅在本研究中将这些林窗划分为小型林窗(G1、G2)、中型林窗(G3、G4、G5)和大型林窗(G6、G7)。于次年(2012年)按照2.5 m ′ 2.5 m的造林密度, 在各梯度林窗内补植一年生珍贵乡土树种油樟优质幼苗构建团块状混交改造模式, 其余灌草植被自然更新, 以远离林窗边缘且大于400 m2的成片马尾松纯林林下为对照处理(CK)。不同大小的林窗和林下对照各设置3个重复样地。样地基本概况见表1。Table 1
表1
表1马尾松人工林不同面积林窗样地基本概况
Table 1General characteristics of the sampling plots in forest gaps with different sizes in Pinus massoniana plantations
| 林窗 Gap | 面积 Size (m2) | 经纬度 Longitude and latitude | 海拔 Altitude (m) | 坡度 Slope (°) | 坡向 Aspect | 坡位 Slope position |
|---|---|---|---|---|---|---|
| G1 | 100 | 28.60° N, 104.56° E | 423 | 24.5 | SW | 中坡 Middle slope |
| G2 | 225 | 28.61° N, 104.56° E | 438 | 26.1 | SE | 中坡 Middle slope |
| G3 | 400 | 28.60° N, 104.56° E | 408 | 23.5 | SE | 中坡 Middle slope |
| G4 | 625 | 28.60° N, 104.57° E | 424 | 24.2 | SE | 中坡 Middle slope |
| G5 | 900 | 28.61° N, 104.57° E | 441 | 21.5 | S | 中坡 Middle slope |
| G6 | 1 225 | 28.61° N, 104.56° E | 418 | 27.0 | SE | 中坡 Middle slope |
| G7 | 1 600 | 28.60° N, 104.56° E | 430 | 26.5 | SE | 中坡 Middle slope |
| CK | - | 28.61° N, 104.57° E | 427 | 23.9 | SE | 中坡 Middle slope |
新窗口打开
1.3 样品采集与测定
2015年3月, 林窗形成4年后对林窗内植被和土壤进行调查, 土壤理化特性见表2。自2015年4月(春季)开始, 每隔一个季度采集一次新鲜植物样品, 即采集时间在2015年7月(夏季)、10月(秋季)和2016年1月(冬季), 并监测不同大小林窗边缘及林下环境因子季节动态(表3)。本研究选取各林窗边缘(林窗边缘定义为林冠空隙与周围边界木基部所围成的部分)及林下共有的人工更新优势植物——四年生幼树油樟为研究对象进行样品采集。采样时, 在林窗4条边缘中点及四角位置, 分别采集近等高油樟东南西北四个方位及上中下不同部位当年生健康成熟叶片各5份, 每份不少于10 g。将同一林窗采集的油樟叶片混合后采用四分法取样并装于牛皮纸信封袋中带回实验室处理, 经105 ℃杀青, 85 ℃烘干至恒质量后, 用粉碎机粉碎制样用于测定C、N、P含量。全碳含量采用重铬酸钾氧化-外加热法(GB 7657-87)测定, 全N和全P含量经H2SO4-H2O2消煮后分别采用半微量凯氏定氮法(LY/T 1269-1999)和钼锑抗比色法(LY/T 1270-1999)测定。Table 2
表2
表2马尾松人工林不同面积林窗样地土壤理化特性(平均值±标准误差)
Table 2Soil physical and chemical properties of the sampling plots in forest gaps with different sizes in Pinus massoniana plantations (mean ± SE)
| 林窗 Gap | 容重 Bulk density (g·cm-3) | 含水量 Soil water content (%) | 最大持水量 Maximum field capacity (g·kg-1) | pH值 pH value | 土壤全碳 Soil total carbon (g·kg-1) | 土壤全氮 Soil total nitrogen (g·kg-1) | 土壤全磷 Soil total phosphorus (g·kg-1) |
|---|---|---|---|---|---|---|---|
| G1 | 1.21 ± 0.09c | 22.10 ± 3.09a | 392.87 ± 6.44a | 4.03 ± 0.11b | 11.67 ± 2.99b | 0.63 ± 0.15bc | 0.45 ± 0.11c |
| G2 | 1.22 ± 0.11bc | 21.67 ± 4.31a | 364.43 ± 10.04ab | 4.22 ± 0.17ab | 11.15 ± 1.51b | 0.57 ± 0.12d | 0.44 ± 0.08d |
| G3 | 1.23 ± 0.08ab | 21.39 ± 4.08ab | 373.16 ± 8.57ab | 4.24 ± 0.23ab | 11.90 ± 1.45ab | 0.56 ± 0.06d | 0.49 ± 0.14a |
| G4 | 1.23 ± 0.11ab | 22.15 ± 3.11a | 379.47 ± 8.85ab | 4.24 ± 0.10ab | 12.67 ± 1.52a | 0.64 ± 0.15b | 0.49 ± 0.07a |
| G5 | 1.22 ± 0.12bc | 21.40 ± 3.51ab | 381.48 ± 11.54ab | 4.34 ± 0.21ab | 11.69 ± 1.21b | 0.67 ± 0.12a | 0.47 ± 0.15b |
| G6 | 1.23 ± 0.17a | 21.99 ± 4.17a | 381.24 ± 13.21ab | 4.34 ± 0.27ab | 11.90 ± 1.54ab | 0.62 ± 0.23c | 0.49 ± 0.12a |
| G7 | 1.23 ± 0.26a | 21.33 ± 1.96ab | 359.62 ± 6.14b | 4.19 ± 0.11ab | 11.14 ± 1.37b | 0.54 ± 0.12e | 0.46 ± 0.08bc |
| CK | 1.24 ± 0.14a | 20.16 ± 2.14b | 358.19 ± 15.79b | 4.46 ± 0.32a | 9.48 ± 0.56c | 0.47 ± 0.62f | 0.43 ± 0.12d |
新窗口打开
Table 3
表3
表3不同季节林窗边缘及林下环境因子概况
Table 3Environmental factors in the sampling plots in the edges of forest gaps with different sizes in different seasons
| 林窗 Gap | 月平均气温 Mean monthly air temperature (℃) | 月平均空气湿度 Mean monthly air humidity (%) | 相对光强 Relative light intensity (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 春 Spring | 夏 Summer | 秋 Autumn | 冬 Winter | 春 Spring | 夏 Summer | 秋 Autumn | 冬 Winter | 春 Spring | 夏 Summer | 秋 Autumn | 冬 Winter | |
| G1 | 19.43 | 28.41 | 21.03 | 6.45 | 71.9 | 84.7 | 94.6 | 83.3 | 61.7 | 87.8 | 55.3 | 58.9 |
| G2 | 19.23 | 28.51 | 22.05 | 6.21 | 72.4 | 87.2 | 94.5 | 86.1 | 63.6 | 90.5 | 57.4 | 59.2 |
| G3 | 19.67 | 29.73 | 21.67 | 5.98 | 69.8 | 89.6 | 94.7 | 86.4 | 64.7 | 94.8 | 60.4 | 60.1 |
| G4 | 19.76 | 29.88 | 22.22 | 9.03 | 72.2 | 90.0 | 96.3 | 89.1 | 67.9 | 95.4 | 65.2 | 66.3 |
| G5 | 19.27 | 30.42 | 22.63 | 8.43 | 69.1 | 88.3 | 93.4 | 84.7 | 68.3 | 95.0 | 66.9 | 66.6 |
| G6 | 19.92 | 30.63 | 21.91 | 7.11 | 71.3 | 86.1 | 92.6 | 85.2 | 71.0 | 96.4 | 69.4 | 68.8 |
| G7 | 19.80 | 30.74 | 22.55 | 7.10 | 69.7 | 86.3 | 91.7 | 85.3 | 71.8 | 98.2 | 70.4 | 71.4 |
| CK | 18.32 | 27.66 | 20.09 | 3.12 | 72.5 | 90.3 | 93.6 | 87.3 | 18.6 | 16.4 | 16.2 | 17.2 |
新窗口打开
1.4 数据处理与统计分析
数据的前期处理、统计分析及绘图分别在Microsoft Office Excel 2007、SPSS 20.0和Origin 9.0中完成。计算林下及不同大小林窗边缘油樟叶片C、N、P含量及C:N、C:P、N:P的算术平均值和标准误差, 采用双因素方差分析法分析林窗大小和采样季节对油樟化学计量特征的影响, 用Tukey HSD法分别检验不同面积及不同季节林窗边缘油樟化学计量特征的差异(α = 0.05)。以11个环境因子为自变量, 以油樟叶片C、N、P含量及C:N、C:P、N:P为因变量在Canoco for Windows 4.5中进行主成分分析(PCA)排序, 结果经Monte Carlo Test检验。
2 结果
2.1 不同季节林窗边缘及林下油樟叶片C、N、P含量
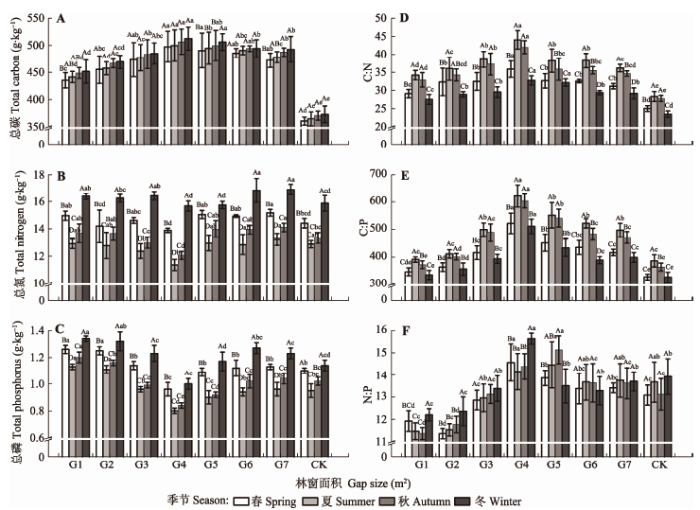
由表4可知, 油樟叶片C含量受林窗面积的显著影响, 叶片N含量受季节动态的显著影响, 叶片P含量受林窗面积和季节动态的显著影响。各林窗边缘油樟叶片C含量在不同季节里均显著高于林下(图2A); 叶片N含量在春季和秋季仅G7林窗边缘显著高于林下, 冬季G6和G7林窗边缘显著高于林下, 在夏季各林窗边缘与林下无显著差异(图2B); 叶片P含量在春、夏、秋季里G1和G2林窗边缘显著高于林下, 在冬季除G4和G5外其余林窗边缘均显著高于林下(图2C)。Table 4
表4
表4林窗面积和季节及交互作用对油樟化学计量特征影响的双因素方差分析(F值)
Table 4Two-way ANOVA on the effects of gap size, season, and their interaction on C, N, P and their stoichiometry of Cinnamomum longepaniculatum
| 变异来源 Source of variation | C | N | P | C:N | C:P | N:P |
|---|---|---|---|---|---|---|
| 林窗面积 Gap size (G) | 198.314*** | 2.072 | 5.789* | 126.905*** | 342.956*** | 78.855*** |
| 季节 Season (S) | 2.848 | 670.556*** | 540.694*** | 257.959*** | 278.434*** | 4.560 |
| 林窗面积×季节 Gap size × Season (G × S) | 3.594 | 3.084 | 4.701 | 4.454 | 3.020 | 9.219** |
新窗口打开
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2马尾松人工林不同季节不同林窗边缘油樟化学计量特征(平均值±标准误差)。不同大写字母代表季节间差异显著; 不同小写字母代表林窗间差异显著。G1、G2、G3、G4、G5、G6、G7分别代表面积为100 m2、225 m2、400 m2、625 m2、900 m2、1 225 m2、1 600 m2的林窗; CK, 林下对照。
-->Fig. 2Carbon, nitrogen, phosphorus and their stoichiometry of Cinnamomum longepaniculatum in the edges of forest gaps with different sizes in different seasons in Pinus massoniana plantations (mean ± SE). Different capital letters indicate significant differences among seasons, and different lowercase letters indicate significant differences among forest gaps. G1, G2, G3, G4, G5, G6, and G7 represent gaps with the size of 100 m2, 225 m2, 400 m2, 625 m2, 900 m2, 1 225 m2, and 1 600 m2, respectively; CK, closed canopy as control.
-->
不同季节里油樟在不同林窗边缘的叶片C含量变化趋势相同, 随林窗面积增大均先升后降, 且在G4林窗边缘有最大值。在油樟叶片N含量中, 秋季变化最显著, 随林窗面积增大先降后升, 其余季节里无显著变化。不同季节里油樟叶片P含量随林窗面积增大均呈现出先降后升的变化趋势, 在G4林窗边缘出现最小值。就季节变化来看, 春季到冬季, 各林窗边缘及林下油樟叶片C含量整体呈逐渐升高的趋势, 但不同季节差异不显著; 叶片N、P含量均呈先降后升的变化趋势, 且在夏季有最小值。
2.2 不同季节林窗边缘及林下油樟叶片化学计量比
油樟叶片C:N及C:P受林窗面积和季节动态的显著影响, 叶片N:P受林窗面积和面积与季节交互作用的显著影响。各林窗边缘油樟叶片C:N在四季均显著高于林下(图2D); 叶片C:P在春、秋、冬季除G1外其余林窗边缘均显著高于林下, 在夏季除G1和G2外其余林窗边缘均显著高于林下(图2E); 叶片N:P在春季和秋季G4和G5林窗边缘显著高于林下, 在冬季仅G4林窗边缘显著高于林下, 在夏季各林窗边缘与林下无显著差异(图2F)。不同季节油樟在不同林窗边缘叶片C:N变化趋势相同, 随林窗面积增大均先升后降, 且在G4林窗边缘有最大值。油樟叶片C:P与C:N在不同季节有一致的变化趋势, 随林窗面积增大先升后降, G4林窗
边缘有最大值。不同季节油樟叶片N:P随林窗面积增大均呈现出先显著上升后平缓下降的变化趋势, 在G4和G5林窗边缘有最大值。就季节变化来看, 春季到冬季, 各林窗边缘及林下油樟叶片C:N和C:P均呈先升后降的变化趋势, 且在夏季有最大值; 叶片N:P无规律变化。
2.3 各环境因子对油樟化学计量特征的相对贡献
选择11种环境因子与油樟叶片C含量、N含量、P含量、C:N、C:P及N:P进行PCA排序, 前两个排序轴对油樟化学计量特征的累积贡献率(累计值占特征值总和)达98.4%, 可见, 第1、2排序轴成分(环境因子)对油樟化学计量特征起主要控制作用。其中第1轴与林窗面积、土壤全P、月平均湿度显著正相关, 林窗面积对第1排序轴贡献最大; 第2轴与含水量、土壤全C、全N、月平均气温及相对光强极显著负相关, 相对光强对第2排序轴贡献最大, 其次为月平均气温(表5)。由图3可知, 林窗面积、含水量、土壤全C、全N、全P、月平均气温及相对光强对油樟叶片N含量影响较小, 对其余计量特征影响较大, 特别是叶片C含量及N:P; 月平均湿度对油樟叶片C含量及N:P影响较小, 对其余计量特征影响较大, 特别是叶片N含量。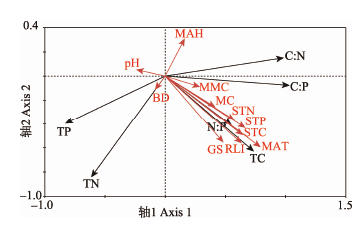 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3马尾松人工林不同林窗边缘油樟化学计量特征与环境因子的主成分分析(PCA)双序图。TC, 叶片C含量; TN, 叶片N含量; TP, 叶片P含量; C:N, 叶片C:N; C:P, 叶片C:P; N:P, 叶片N:P。BD, 容重; GS, 林窗面积; MAH, 月平均湿度; MAT, 月平均气温; MC, 含水量; MMC, 最大持水量; RLI, 相对光强; STC, 土壤全碳; STN, 土壤全氮; STP, 土壤全磷。
-->Fig. 3The principal component analysis (PCA) on carbon, nitrogen, phosphorus and their stoichiometry of Cinnamomum longepaniculatum and environmental variables in the edges of forest gaps with different sizes in Pinus massoniana plantations. TC, total C in leaf; TN, total N in leaf; TP, total P in leaf; C:N, C:N in leaf; C:P, C:P in leaf; N:P, N:P in leaf. BD, bulk density; GS, gap size; MAH, monthly average humidity; MAT, monthly average air temperature; MC, moisture content; MMC, maximum moisture capacity; RLI, relative light intensity; STC, total C in soil; STN, total N in soil; STP, total P in soil.
-->
Table 5
表5
表5环境变量与排序轴的相关系数
Table 5Correlation coefficients of environmental variables with ordination axes
| 环境变量 Environmental variables | 排序轴 Ordination axis | |
|---|---|---|
| 第1轴 Axis 1 | 第2轴 Axis 2 | |
| 林窗面积 Gap size | 0.506 4** | -0.245 3 |
| 容重 Bulk density | 0.134 5 | 0.333 5 |
| 含水量 Moisture content | 0.038 0 | -0.544 2** |
| 最大持水量 Maximum moisture capacity | 0.075 0 | -0.381 7 |
| pH值 pH value | -0.027 0 | 0.381 1 |
| 土壤全碳 Total C in soil | 0.251 1 | -0.510 5** |
| 土壤全氮 Total N in soil | 0.210 3 | -0.552 8** |
| 土壤全磷 Total P in soil | 0.425 4* | -0.356 6 |
| 月平均气温 Monthly average air temperature | 0.371 6 | -0.659 5*** |
| 月平均湿度 Monthly average humidity | 0.446 6* | 0.210 0 |
| 相对光强 Relative light intensity | 0.099 7 | -0.811 4*** |
新窗口打开
3 讨论
3.1 油樟叶片C、N、P元素含量分析
C、N、P元素是细胞结构与功能最重要的生命元素(王振南和杨惠敏, 2013), 相同生活型植物往往趋于分享一套关键的功能性状来实现对环境的趋同适应(李家湘等, 2017)。因此, 与相关常绿阔叶林研究结果相比, 本研究中油樟平均叶片C含量(466.04 g·kg-1)与广东鼎湖山(472.46 g·kg-1)(王晶苑等, 2011)及云南普洱地区(469.30 g·kg-1)(刘万德等, 2015)植物叶片C含量相似, 研究区油樟叶片具有较强的C存储能力; 而油樟平均叶片P含量低于云南普洱地区(1.30 g·kg-1)(刘万德等, 2015)植物叶片P含量, 平均叶片N含量远低于广东鼎湖山(19.82 g·kg-1)(王晶苑等, 2011)、云南普洱(23.30 g·kg-1)(刘万德等, 2015)及浙江天童山(16.06 g·kg-1)(阎恩荣等, 2010)地区植物叶片N含量。这可能是因为研究区高温多雨能够导致N、P元素淋溶(杜满义等, 2016), 使油樟可吸收利用的有效氮或有效磷减少; 低的P含量还可能与研究区所属酸性山地黄壤对P有很强的固定作用有关, 也与全国土壤P含量普遍较低的结果(李慧等, 2016)一致。C含量在一定程度上可以表征植物存储C积累有机物质的能力(Elser et al., 2010), N、P含量则是植物在一定生境下吸收N、P等营养元素的能力(李家湘等, 2017)。本研究中, 各林窗边缘油樟叶片C含量四季均显著高于林下, 平均叶片N、P含量在林窗边缘亦高于林下, 具有明显的边缘效应。林窗与森林结合的部位构成林窗边缘, 与林下相比, 林窗边缘能更好地接受阳光直射从而形成适宜植物生长代谢的温度与光照条件(田超等, 2011)。PCA分析表明, 月平均气温和相对光强与油樟叶片C含量呈极显著正相关关系, 与P含量相关性较N含量更强, 叶N含量比P含量具有更高的自我调控能力, 变异性和环境依赖性较小, 表现出较强的化学内稳态(Chen et al., 2013; Pan et al., 2015; 李家湘等, 2017)。光合固C是植物体内主要的C来源, 光照、温度及H2O的变化都会引起光合效率的改变(刘辉等, 2015)。林窗边缘湿度(84.5%)与林下湿度(85.9%)相近, 但边缘区光照强度(55.3%-98.2%)远高于林下(16.2%-18.6%), 特别在夏季更加明显, 充足的光照及适宜的温湿度均有利于边缘区油樟进行光合作用, 进而增加对有机物质的合成与积累, 同时还能加快养分周转, 增加土壤养分的可获得性。
此外, 因林窗空间尺度不同, 使得不同面积林窗边缘拥有光照、热量和水分各异的区域小气候(Gálhidy et al., 2006)。本研究发现, 林窗大小对油樟叶片C、P含量有显著影响, 在不同季节, C含量随林窗面积增大先升后降, 且均为中型林窗G3-G5(特别是G4)有机质积累量最高, 而P含量具有相反的变化趋势, 在一定程度上支持了温度-植物生理假说(Reich & Oleksyn, 2004)。PCA分析表明, 油樟叶片C含量、P含量与月平均气温、月平均湿度、光强及土壤含水量有显著相关性。王丽霞等(2013)的研究表明中林隙土壤含水量最丰富, 其次为大林隙和小林隙, 中型林窗边缘更有利于植物的生长。林窗形成后疏开了上层林冠, 影响着空间及资源的组合状况, 光照强度和气流的改变影响了不同林窗边缘温湿度的变化(Gálhidy et al., 2006)。小型林窗边缘因林冠遮阴及截流导致光照较弱且水分含量较低, 大型林窗边缘因中心所接受的阳光辐射太强使得地面温度较高且水分蒸发较快, 它们均不能保持适宜的温湿度及降水、蒸腾的良好平衡, 不利于油樟的生长。而中型林窗边缘温度低于大型林窗, 适宜的温湿度有利于土壤理化过程(宋小艳等, 2014)、凋落物中有机质的分解矿化(Arunachalam & Arunachalam, 2000)和土壤动物微生物的繁殖(Sylvain & Wall, 2011), 这些要素对于油樟的光合作用、生长发育、高养分吸收利用效率的维持有着重要的作用。
3.2 油樟叶片C、N、P化学计量比分析
叶片C:N和C:P可揭示生物量(C)和养分比值关系, 即养分利用效率, 较高的C:N和C:P代表植物对N、P的利用率较高(Elser et al., 2010)。本研究中, 林窗大小对油樟叶片C:N和C:P有极显著影响, 且不同季节各林窗边缘均显著高于林下。在辐射强烈的林窗中心与林木郁闭的林下之间, 光、温、水的再分配改变了地表水热动态(Kern et al., 2013), 干燥与湿润的气流交替循环(刘辉等, 2015), 使林窗边缘形成异于邻近林下的温和的区域微环境, 可影响物种更新与分解者群落结构, 进一步影响林窗边缘植被养分利用和周转效率(Schliemann, 2011; 张艳等, 2015), 深刻作用于植物养分分配格局。林窗边缘区土壤动物多样性和丰富度高于林下(张明锦等, 2016), 土壤动物与植物根系相互作用, 对营养物质的转化、储存和释放发挥着积极作用。同时, 林窗边缘物种丰富度指数也大于林下(崔宁洁等, 2014), 植物间竞争压力较大, 对土壤养分的激烈争夺促进了地上/地下养分周转速率, 进而提高了优势种对资源的利用效率。张艳等(2015)的研究表明, 林窗边缘凋落物叶难降解物质降解率高于林下, 凋落物分解可为植物根系提供充足的有机质, 促进植物养分吸收利用。Foereid等(2010)也认为光照时间越长, 辐射通量越大, 光降解作用使养分得到快速分解和释放, 从而为植物生长代谢提供更好的养分基础, 可见林窗边缘油樟对养分吸收利用具有明显的边缘效应。更为重要的是, 林窗面积不同会导致林窗边缘地表接收不同强度和不同时长的阳光直射及降水分配而拥有迥异的水热模式, 进而影响油樟养分周转及分配格局。本研究中, 不同季节油樟叶片C:N和C:P随林窗面积增大先升后降, 且均在G4林窗边缘呈现最大值, 养分利用效率最高, G4林窗边缘C、N、P元素分配相对平衡, 边缘效应更显著。在植物群落中, 非生物环境的C、N和P化学计量比值一旦不相匹配, 就会导致C、N和P循环途径的不同(Elser et al., 2000; Fan et al., 2015), 任一元素稀缺或过量必将导致另外两种元素积累或消耗。从G1到G4林窗边缘, C:N和C:P上升, 表征油樟朝着N、P相对稀缺而C相对过量的方向生长; 从G4到G7林窗边缘, C:N和C:P下降, 表征油樟朝着N、P相对充裕而C相对不足的方向生长。C:N和C:P变化趋势均在G4林窗边缘出现高值拐点, 这在一定程度上反映了G4林窗边缘油樟C、N、P元素的相对平衡状态以及对N、P营养元素的高的利用效率, 边缘效应更显著。一方面, 蒸腾作用是养分元素从根系运输到叶片的主导驱动力, 且养分元素溶于水以离子态被植物吸收利用(刘辉等, 2015), 水分过低将影响到植物对养分元素的吸收, G4林窗边缘土壤含水量、全C及全P显著高于其余林窗, 充足的水分及养分环境为油樟提供了良好的生长条件; 另一方面, 大型林窗边缘中午的强光照也会导致植物叶片气孔变小或关闭, 气孔阻力增加, 蒸腾作用减弱, 而G4林窗边缘适宜的光温环境保障了油樟蒸腾作用的正常进行及对N、P元素的吸收利用。
叶片或生物量中N:P可作为生态系统限制性营养元素的一个重要指标(Han et al., 2005)。目前大量研究认为这种限制的阈值在不同地域、不同物种上表现不一致(Wassen et al., 1995; Koerselman & Meuleman, 1996; 杜满义等, 2016)。本文采用Güsewell (2004)得出的陆地生态系统阈值: 当植被N:P小于10时, 植物生长受N限制; N:P大于20时, 植物生长受P限制; 介于两者之间时, 植物受N、P共同限制或不受二者限制。本研究中油樟在4个季节不同林窗边缘中N:P均在10-20之间, 根据这一标准可知研究区油樟生长与N、P关系暂不明显, 接下来可实施增肥实验来确定该地区N:P临界值或N、P限制作用。
植物叶片元素含量及比例与自身结构特点及生长节律有密切联系(刘万德等, 2015)。本研究表明, 夏秋季油樟叶片对养分元素的吸收利用优于春冬季, 林窗边缘效应更显著。一方面, 春季林窗气温开始回升, 植物叶片细胞快速分裂需大量的蛋白质和核酸(李家湘等, 2017), 因此叶片选择性吸收N、P元素, 二者浓度较高, C:N、C:P较低; 在夏季, 林窗内光照强度增加, 降雨充足, 植物生长旺盛, 根系对营养元素的吸收速度不及细胞膨胀生长速度(李慧等, 2016), 因此N、P元素浓度逐渐被稀释而剧烈下降, C:N、C:P升高; 在秋季, 林窗内光照减弱, 温湿度适宜, 植物稳定生长, 根系吸收能力加强, 且植物代谢强度增加, N、P元素浓度略升高, C:N、C:P开始降低; 在冬季, 植物代谢强度降低, 部分衰老叶片凋落之前, 营养元素回收至植物体内及其他健康成熟叶片当中, N、P元素升高, C:N、C:P显著降低。另一方面, 凋落物的归还也是C和N等元素的主要来源(Arunachalam & Arunachalam, 2000; 张明锦等, 2016)。有研究表明夏季温暖湿润的林窗环境更有利于难降解物质中酸不溶性组分在微生物作用下进行降解, 释放出更多的养分供给植物利用(张艳等, 2015)。研究区域位于亚热带季风气候带, 研究期间降雨多集中在5-9月, 夏秋季较高的温湿度环境可提高林窗边缘土壤酶活性、促进土壤动物和微生物对凋落物的分解, 进而提高养分可利用性。
4 结论
综上所述, 本研究区马尾松人工林各林窗边缘油樟叶片C含量及N、P养分利用效率显著高于林下, 在化学计量特征水平上具有明显的边缘效应, 且夏秋季更显著。在不同季节里, 均为G4 (625 m2)林窗边缘油樟叶片有机质含量最高, C、N、P元素最趋于动态平衡, 边缘效应最显著。这将为开展马尾松人工林养分管理提供科学依据。致谢 感谢四川农业大学生态林业研究所谭波老师在野外采样、室内分析工作中给予的帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | We examined the effects of treefall gap size and soil properties on microbial biomass dynamics in an undisturbed mature-phase humid subtropical broadleaved forest in north-east India. Canopy gaps had low soil moisture and low microbial biomass suggesting that belowground dynamics accompanied changes in light resources after canopy opening. High rainfall in the region causes excessive erosion/leaching of top soil and eventually soil fertility declines in treefall gaps compared to understorey. Soil microbial population was less during periods when temperature and moisture conditions are low, while it peaked during rainy season when the litter decomposition rate is at its peak on the forest floor. Greater demand for nutrients by plants during rainy season (the peak vegetative growth period) limited the availability of nutrients to soil microbes and, therefore, low microbial C, N and P. Weak correlations were also obtained for the relationships between microbial C, N and P and soil physico-chemical properties. Gap size did influence the microbial nutrients and their contribution to soil organic carbon, total Kjeldhal nitrogen and available-P. Contribution of microbial C to soil organic carbon, microbial N to total nitrogen were similar in both treefall gaps and understorey plots, while the contribution of microbial P to soil available-P was lower in gap compared to the understorey. These results indicate that any fluctuation in microbial biomass related nutrient cycling processes in conjunction with the associated microclimate variation may affect the pattern of regeneration of tree seedlings in the gaps and hence be related with their size. |
| [2] | Leaf chemistry is important in predicting the functioning and dynamics of ecosystems. As two key traits, leaf nitrogen (N) and phosphorus (P) concentrations set the limits for plant growth, and leaf N:P ratios indicate the shift between N- and P-limitation. To understand the responses of leaf chemistry to their potential drivers, we measured leaf N and P concentrations of 386 woody species at 14 forest sites across eastern China, and explored the effects of climate, soil, and plant growth form on leaf N, P and N:P ratios. In general, leaf N and P were both negatively related to mean annual temperature and precipitation, and positively related to soil N and P concentrations. Leaf N:P ratios showed opposite trends. General linear models showed that variation in leaf N was mainly determined by a shift in plant growth form (from evergreen broadleaved to deciduous broadleaved to conifer species) along the latitudinal gradient, while variations in leaf P and N:P were driven by climate, plant growth form, and their interaction. These differences may reflect differences in nutrient cycling and physiological regulations of P and N. Our results should help understand the ecological patterns of leaf chemical traits and modeling ecosystem nutrient cycling. |
| [3] | . 人工林目前存在结构单一、土壤退化、生物多样性降低等人类普遍关注的生态问题。马尾松(Pinus massoniana)是长江上游低山丘陵区退耕还林的主要人工林树种。研究采伐林窗对植物物种组成和更新的影响,对马尾松低效人工林的改造,提升其生态服务功能具有重要的意义。该文以采伐39年生的马尾松人工林形成的7种不同大小的林窗为研究对象,分析了不同季节林窗内的植物生活型组成及多样性变化。结果表明:1)马尾松人工林林下植物以高位芽植物居多,其次是地面、地下芽植物,一年生植物较少而缺少地上芽植物。在林窗形成初期,林窗的高位芽植物比例明显低于林下,大林窗的高位芽植物比例稍高于小林窗,地下芽和一年生植物的比例低于小林窗。2)林下的物种丰富度和物种多样性指数显著低于大林窗。不同林窗下植物的丰富度指数、优势度指数、多样性指数也存在显著差异。3)夏季林窗下植物多样性最高,其次是秋季,春季多样性最低。1225-1600 m2的大林窗能够促进马尾松人工林植物多样性恢复和植被更新。 |
| [4] | The main objectives were to study the effect of gap size and canopy openness on the natural regeneration dynamics considering the parameters of sapling growth, recruitment, mortality, density, species composition and above-ground biomass accumulation. The study was carried out in 32 artificial gaps with sizes varying from 100 to 120002m 2 and canopy openness from 10 to 45%, from the second to the twelfth year after gap creation. The gap size was measured using the vertical projection of the tree crowns on the ground (Brokaw's definition), and the canopy openness measurement by hemispherical photography. In the first five years, mean sapling growth (0.5402cm02year 611 ), mortality (3.9%02year 611 ) and AGB (26.202Mg02ha 611 or 8.702Mg02ha 611 02year 611 ) were significantly higher in the gaps than in the forest understorey (0.1702cm02year 611 , 1.5%02year 611 and 610.5902Mg02ha 611 02year 611 respectively) and positively correlated with gap size and canopy openness. In the same period, recruitment was also significantly higher in the gaps (5.8%02year 611 ) than in the forest understorey (0.4%02year 611 ) but decreased with gap size and negatively correlated with canopy openness. In the first five years, the relative density of pioneer species was higher in the gaps but not significantly correlated with gap size or canopy openness. AGB increased linearly since canopy opening, and twelve years after gap creation it was still higher in larger (121.202Mg02ha 611 or 10.102Mg02ha 611 02year 611 ) rather than smaller (62.502ha 611 or 5.202ha 611 02year 611 ) gaps. Twelve years after gap creation there were no significant differences in the parameters of sapling growth, recruitment, and mortality which could be attributed to the original gap size and canopy openness. |
| [5] | AbstractQuestionsThe ecology of landscape gaps is important for the maintenance of species diversity in a variety of plant ecosystems. In the scrubby flatwoods community type of the pyrogenic Florida scrub, we aimed to identify the relationship between time-since-fire, gap area, quality (microhabitat) and aggregation (structural connectivity) with neighbouring gaps on gap species diversity and occupancy.LocationArchbold Biological Station (ABS), Lake Wales Ridge, south-central Florida, USA.MethodsWe measured species diversity, gap area, aggregation and microhabitat (percentage bare sand) in 120 gaps divided evenly amongst 1, 2, 4, 8, 16 and 26 yrs since fire in the scrubby flatwoods of ABS (typical fire return interval of 5 20 yrs) throughout the summer of 2011.ResultsGap area and aggregation were significantly highest in the first 4 yrs after fire, but decreased and remained fairly constant at longer times-since-fire. Percentage bare sand gradually decreased with time-since-fire until a strong decline at the 26-yr mark, where litter cover was significantly higher than all previous years. Gap area strongly and positively predicted species richness (total and herb spp.) and total species diversity (H'). Bare sand content was positively correlated with the richness of species that are obligate seeders post-fire. Gap aggregation was generally a poor predictor of diversity components, although it positively influenced the occurrence of Stipulicida setacea, a seeder species that occasionally resprouts after fire. Overall, species richness and diversity peaked at intermediate times-since-fire, probably because long-unburned gaps were too small and inundated with litter to sustain higher richness of herbs and obligate seeders.ConclusionIn a prior gap ecology study, in neighbouring more xeric rosemary scrub, gaps were larger, more persistent and had higher bare sand cover, resulting in higher levels of total species richness and diversity for decades longer after fire than gaps in this study. In both these types of Florida scrub, the threat of fire suppression and its adverse effects on gap area and microhabitat may be detrimental for several endemic species that depend on gaps. |
| [6] | . 碳(C)、氮(N)、磷(P)生态化学计量比是生态系统过程与功能的重要特征,开展种群生态化学计量学研究可以细化植物种群化学计量学内容,确定限制植物生长的元素类型,同时为大尺度模型的发展提供数据基础。为阐明我国毛竹(Phyllostachys edulis)林C、N、P化学计量学特征,通过对毛竹主要产区文献数据的搜集整理与分析,探索我国毛竹林“植物-土壤-凋落物”系统C、N、P及C:N、C:P、N:P生态化学计量特征,以及不同组分生态化学计量特征与经纬度之间的关系。结果表明:1)我国毛竹林叶片C含量为478.30 mg·g(–1),N含量为22.20 mg·g(–1),P含量为1.90 mg·g(–1),C:N为26.80,C:P为299.60,N:P为14.40;毛竹林0–20 cm土层C含量为21.53 mg·g(–1),N含量为1.66 mg·g(–1),P含量0.41 mg·g(–1),C:N为14.20,C:P为66.74,N:P为4.28;毛竹凋落物C含量为438.49 mg·g(–1),N含量为13.39 mg·g(–1),P含量为0.86 mg·g(–1),C:N为22.53,C:P为665.67,N:P为22.55。2)毛竹林“植物-土壤-凋落物”系统中,C:N表现为:叶片>凋落物>土壤,C:P和N:P均表现为:凋落物>叶片>土壤,叶片N、P再吸收率分别为39.68%和54.74%,我国毛竹林生长发育总体上可能受到P限制或者N和P两种元素的双重限制。3)纬度梯度:叶片N含量、N:P随纬度增加而增加,C:N随纬度增加而降低。经度梯度:叶片N:P随经度增加而增加,P含量、C:N随经度增加而降低;土壤C:N随经度增加而增加,N含量随经度增加而降低;凋落物N含量随经度增加而降低。4)叶片N含量与年平均气温和年降水量均存在明显负相关关系,但对温度的响应比降水更敏感,叶片N含量与纬度呈正相关关系,支持“温度-植物生理假说”,反映了植物对自然环境的适应。 |
| [7] | We experimentally investigated interacting effects of canopy gaps, understory vegetation and leaf litter on recruitment and mortality of tree seedlings at the community level in a 20-year-old lowland forest in Costa Rica, and tested several predictions based on results of previous studies. We predicted that experimental canopy gaps would greatly enhance tree seedling recruitment, and that leaf litter removal would further enhance recruitment of small-seeded, shade-intolerant seedlings in gaps. We created a large (320–540 m 2) gap in the center of 5 out of 10 40 m × 40 m experimental plots, and applied the following treatments bimonthly over a 14-month-period in a factorial, split–split plot design: clipping of understory vegetation (cut, uncut), and leaf litter manipulations (removal, addition, control). As expected, experimental gaps dramatically increased tree seedling recruitment, but gap effects varied among litter treatments. Litter addition reduced recruitment in gaps, but enhanced recruitment under intact canopy. Species composition of recruits also differed markedly between gap treatments: several small-seeded pioneer and long-lived pioneer species recruited almost exclusively in gaps. In contrast, a few medium-to-large-seeded shade-tolerant species recruited predominantly under intact canopy. Leaf litter represents a major barrier for seedling emergence and establishment of small-seeded, shade-intolerant species, but enhances emergence and establishment of large-seeded, shade-tolerant species, possibly through increased humidity and reduced detection by predators. Periodic clipping of the understory vegetation marginally reduced tree seedling mortality, but only in experimental gaps, where understory vegetation cover was greatly enhanced compared to intact canopy conditions. Successful regeneration of commercially valuable long-lived pioneer trees that dominate the forest canopy may require clear-cutting, as well as weeding and site preparation (litter removal) treatments in felling clearings. Management systems that mimic natural canopy gaps (reduced-impact selective logging) could favor the regeneration of shade-tolerant tree species, potentially accelerating convergence to old-growth forest composition. In contrast, systems that produce large canopy openings (clear-cutting) may re-initiate succession, potentially leading to less diverse but perhaps more easily managed “natural plantations” of long-lived pioneer tree species. |
| [8] | Shows both similarities and differences in the carbon: nitrogen: phosphorus ratios of primary producers (autotrophs) and invertebrate primary consumers (herbivores) across habitats. Ratios in autotrophs, which are nutrient-poor, in both terrestrial and lake food webs; Nutrient-rich terrestrial herbivores (insects) and freshwater herbivores (zooplankton), which have identical C:N:P stoichiometry; Stoichiometric constraints on herbivore growth, which appear to be qualitatively similar and widespread in both environments. |
| [9] | Abstract Contents Summary 593 I. Introduction 594 II. Variation in plant C:N:P ratios: how much and what are the sources? 595 III. The growth rate hypothesis in terrestrial plants and the scaling of whole-plant N:P stoichiometry and production 597 IV. Scaling from tissues to whole plants 599 V. Applications: large-scale patterns and processes associated with plant stoichiometry 601 VI. Global change and plants: a stoichiometric scaling perspective 603 VII. Synthesis and summary 604 Acknowledgements 605 References 605 Summary Biological stoichiometry theory considers the balance of multiple chemical elements in living systems, whereas metabolic scaling theory considers how size affects metabolic properties from cells to ecosystems. We review recent developments integrating biological stoichiometry and metabolic scaling theories in the context of plant ecology and global change. Although vascular plants exhibit wide variation in foliar carbon:nitrogen:phosphorus ratios, they exhibit a higher degree of toichiometric homeostasis than previously appreciated. Thus, terrestrial carbon:nitrogen:phosphorus stoichiometry will reflect the effects of adjustment to local growth conditions as well as species replacements. Plant stoichiometry exhibits size scaling, as foliar nutrient concentration decreases with increasing plant size, especially for phosphorus. Thus, small plants have lower nitrogen:phosphorus ratios. Furthermore, foliar nutrient concentration is reflected in other tissues (root, reproductive, support), permitting the development of empirical models of production that scale from tissue to whole-plant levels. Plant stoichiometry exhibits large-scale macroecological patterns, including stronger latitudinal trends and environmental correlations for phosphorus concentration (relative to nitrogen) and a positive correlation between nutrient concentrations and geographic range size. Given this emerging knowledge of how plant nutrients respond to environmental variables and are connected to size, the effects of global change factors (such as carbon dioxide, temperature, nitrogen deposition) can be better understood. |
| [10] | Ecological stoichiometry plays important roles in ecosystem dynamics and functioning, but relationships between above- and belowground stoichiometry and stoichiometric effects on the growth of differe |
| [11] | Many field experiments have indicated that litter decomposition in semi-arid areas may be partly or fully controlled by photodegradation. We devised a study to test our hypothesis that light exposure makes plant litter more degradable. Dry, senescent, aboveground plant litter from Miscanthus x giganteus was exposed to light including ultraviolet (UV) radiation for various lengths of time from 0 to 289days. Weight loss was measured after exposure and appeared to be modest and did not increase with time of exposure. The litter of the longest and shortest exposure time as well as controls were then incubated with soil and moisture for 35days and CO2 and N2O production were measured. The longest exposed litter degraded much faster than any other treatment during incubation with moisture, about twice as fast as the unexposed control. The shortest exposed however, degraded only slightly faster than the unexposed control. This suggests that increasing litter degradability is a more important mechanism for photodegradation than direct light-induced mass loss. N2O production from decomposition of the exposed litter was high in the beginning, suggesting that nitrogen may be released quickly. The mechanism is probably that light exposure leaves the nitrogen in plant litter easily available to microbial utilisation upon wetting. Such a mechanism might play an important role for nutrient cycling in semi-arid areas. |
| [12] | |
| [13] | In arid ecosystems, abiotic processes facilitate the physical and chemical degradation of plant litter to the extent that decomposition models that use climatic and litter composition variables as surrogates for microbial activity are not predictive. The purpose of this study was to estimate the potential contribution of photodegradation to the decomposition of plant litters that varies in architecture and chemical composition. Litter of Pinus edulis , Juniperus monosperma and Populus deltoides were exposed to ambient and attenuated sunlight, with and without supplemental water additions, at a riparian forest site along the Middle Rio Grande (New Mexico, USA). Mass loss, elemental composition, and microbial extracellular enzyme activities (EEA) were measured over 639 days. The composition of the fungal communities associated with the decomposing litters was compared by analyses of fungal ITS nrDNA sequences. Litter exposed to ambient sunlight had greater mass loss rates than shaded litter, independent of the water treatment: Populus increased by 100%, Pinus by 86% and Juniperus by 46%. The increases were proportional to exposed litter surface area per g dry mass. EEA potentials, particularly oxidative activities, were low in comparison to those measured in mesic ecosystems. For Populus litter, the principal driver of photoacceleration appeared to be photodegradation of cellulose; for Pinus , it was photodegradation of polyphenols; for Juniperus accelerated mass loss was associated with photodegradation of both polysaccharides and polyphenols. Fungal community composition varied by litter type, but the dominant colonizers were yeasts and dark-septate hyphal taxa; a finding consistent with the low enzymatic oxidation potential. This study shows that photochemical oxidation can supplement enzymatic oxidation and increase decomposition rates. As a result, organic matter decomposition in arid ecosystems is not restricted to periods of high moisture availability as is plant production. This decoupling may partly account for the low soil organic matter content of these ecosystems. |
| [14] | We examined edge effects within a forest along old field boundaries in a mixed deciduous/hemlock forest community in upstate New York. Species composition and community structure of both forest understory and treefall gap vegetation were analyzed in two ways: first, we used belt transects to quantify changes in the understory community with distance from the forest edge, and second, we compared species composition of treefall gaps both near and distant from the edge. Exotic species and locally rare species were significantly more abundant at the forest edges than in the forest interior, while tree seedlings were less common at the forest edge. Species composition of treefall gaps near the old field/forest edge differed from composition of treefall gaps in the forest interior during most of the growing season. Treefall gaps at the forest edge, as a group, were compositionally more heterogeneous than treefall gaps distant from the edge. |
| [15] | |
| [16] | |
| [17] | $\bullet$ Leaf nitrogen and phosphorus stoichiometry of Chinese terrestrial plants was studied based on a national data set including 753 species across the country. $\bullet$ Geometric means were calculated for functional groups based on life form, phylogeny and photosynthetic pathway, as well as for all 753 species. The relationships between leaf N and P stoichiometric traits and latitude (and temperature) were analysed. $\bullet$ The geometric means of leaf N, P, and N : P ratio for the 753 species were 18.6 and $1.21 mg g^{-1}$ and 14.4, respectively. With increasing latitude (decreasing mean annual temperature, MAT), leaf N and P increased, but the N : P ratio did not show significant changes. $\bullet$ Although patterns of leaf N, P and N : P ratios across the functional groups were generally consistent with those reported previously, the overall N : P ratio of China's flora was considerably higher than the global averages, probably caused by a greater shortage of soil P in China than elsewhere. The relationships between leaf N, P and N : P ratio and latitude (and MAT) also suggested the existence of broad biogeographical patterns of these leaf traits in Chinese flora. |
| [18] | |
| [19] | The Gap Partitioning Hypothesis (GPH) posits that gaps create heterogeneity in resources crucial for tree regeneration in closed-canopy forests, allowing trees with contrasting strategies to coexist along resource gradients. Few studies have examined gap partitioning of temperate, ground-layer vascular plants. We used a ground-layer plant community of a temperate deciduous forest in northern Wisconsin, USA, as a model system to test whether the GPH extends to the relatively species-rich ground layer. We used a well-replicated experimental approach that included a gap opening gradient (five gap sizes, 6, 10, 20, 30 and 46 m diameter, and undisturbed reference areas), a within-gap location gradient (gap edge to center), and a temporal gradient (0, 2, 6 and 13 years after gap creation). The data were observations of ground-layer plant abundance, published plant traits, and a modeled index of understory light environments. |
| [20] | This study identifies patterns in the gap disturbance regime along a successional gradient in the southern boreal forest and uses this information to investigate canopy composition changes. Gaps were characterized in hardwood, mixed-forest, and conifer stands surrounding Lake Duparquet in northwestern Quebec. From 39 to 80 gaps were evaluated along transects established in each of these stands. The abundance of gap makers and gap fillers and total regeneration was evaluated by species, as well as the size of each gap encountered along the transects. The percentage of the forest in canopy gap was calculated directly from the proportion of the transect in gap and by using gap area and line-intercept techniques. Changes in composition were evaluated from gap-maker and gap-filler distributions and by using transition matrices based on species mortality and regeneration in canopy gaps. The percentage of the forest in canopy gap ranges from 7.1% in a 50-yr-old forest dominated primarily by aspen to 40.4% in a 234-yr-old fir-dominated forest. Gap events are due to individual or small-group tree mortality in the early successional forest but become species-specific events controlled by spruce budworm outbreaks in the later stages of succession. Due to the high latitude, direct light only reaches the forest floor in the very largest gaps of the conifer-dominated stands. However, these gaps form slowly as budworm-caused mortality occurs over a number of years, whereas in aspen-dominated stands gaps are formed quickly by the snapping of tree stems. Balsam fir is the most abundant gap-filling species; however, its abundance is negatively correlated to gap size in all stand types. Markovian transition matrices suggest that in the young aspen-dominated forests small gaps lead to species replacment by more shade-tolerant conifers but that in the oldest forests the larger gaps will result in maintenance of the intolerant species and an increase in the abundance of cedar. |
| [21] | 1. Nutrient limitation (mostly N or P) is a driving force in ecosystem development. Current techniques to determine the nature of nutrient limitation use laborious fertilization experiments. 2. It was hypothesized that the N:P ratio of the vegetation directly indicates the nature of nutrient limitation on a community level (N vs. P limitation). This hypothesis was tested by reviewing data on fertilization studies in a variety of European freshwater wetland ecosystems (bogs, fens, wet heathlands, dune slacks, wet grasslands). In a subset of the data (dune slacks) between-site intraspecific variation and within-site interspecific variation in nutrient content and N:P ratio was studied in five plant species. 3. A review of 40 fertilization studies reveals that an N:P ratio >16 indicates P limitation on a community level, while an N:P ratio <14 is indicative of N limitation. At N:P ratios between 14 and 16, either N or P can be limiting or plant growth is colimited by N and P together. In only one out of 40 fertilization studies, the N:P ratio gave a false indication of the nature of nutrient limitation. Measuring the N:P ratio of the vegetation is a simple and cheap alternative to fertilization studies. The method can only be used under conditions where either N or P controls plant growth. 4. The dataset contains a large variety of vegetation types and plant species, and 11 of the 40 sites were near-monocultures. This suggests that interspecific differences in critical N:P ratios among species may be insignificant. However, a rigorous test of this hypothesis is required. 5. A survey in 18 dune slacks showed large within-site variation in N:P ratio among five species (Calamagrostis epigejos, Phragmites australis, Lycopus europaeus, Mentha aquatica and Eupatorium cannabinum). The N:P ratios of the five species suggested that within plant communities species can be differentially limited by N or P. Moreover, species with an N:P ratio that suggested P-limitation were found at sites where N controlled community biomass production, and vice versa. Between-site intraspecific variation in N and P contents and N:P ratios was also large, and about equal for the five species. This illustrates the plasticity of plant species with respect to N and P contents, probably in response to differences in N and P supply ratios. 6. The vegetation N:P ratio is of diagnostic value and its use may increase our understanding of numerous facets of physiological, population, community and ecosystem ecology. |
| [22] | . [目的]探究晋西黄土区不同树种营养器官(叶、枝、干、根)及其土壤的养分含量变化特征。[方法]通过野外样地调查和取样分析方法,对3种典型森林树种(刺槐、侧柏、辽东栎)不同器官及其林地土壤有机碳、全氮、全磷含量进行了研究,并探讨了不同树种叶片与其土壤 C、N、P 化学计量的关系。[结果]辽东栎、刺槐和侧柏的叶片有机碳含量分别为468.43、454.96、438.53 g·kg -1,刺槐、辽东栎和侧柏叶片的全氮含量分别为27.52、20.74、12.73 g· kg -1,辽东栎、侧柏和刺槐叶片的全磷含量分别为2.73、2.15、1.35 g·kg -1,叶片 C∶N 值为侧柏>辽东栎>刺槐, C∶P、N∶P 值分别为刺槐>侧柏>辽东栎、刺槐>辽东栎>侧柏;3个树种树枝的有机碳含量均最大,树叶的全氮含量均最大,而树干的全氮含量均最小。侧柏、辽东栎、刺槐树枝的全磷含量分别为3.07、3.07、1.87 g·kg -1,显著比其它器官的大;3种森林土壤的 C、N、P 含量均随土层的加深而降低,且010 cm 土层的含量最大,C∶N、C∶P、N∶P值随土层深度的变化不一致;刺槐叶片的有机碳、全磷含量与土壤 C∶N 值显著或极显著负相关;侧柏叶片的有机碳含量均与土壤 C∶N、C∶P、N∶P 值极显著负相关,叶片 N∶P 值与土壤 C∶N、C∶P、N∶P 值均极显著正相关;辽东栎叶片的有机碳含量、C∶P 值与土壤有机碳含量极显著负相关。[结论]辽东栎、刺槐和侧柏的有机碳含量均较高,分别为468.43、454.96、438.53 g·kg -1;辽东栎和刺槐对晋西黄土区干旱环境的防御能力比侧柏强,当地环境较适宜辽东栎的生长发育,而刺槐和侧柏的生长分别受土壤 P 和 N 的限制。 |
| [23] | . 探索植物器官中氮(N)、磷(P)含量沿环境梯度的分异规律,有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制。该文采用分层随机抽样法,在中国南方12个省市区布设462个灌丛样点,对其中193种优势木本植物叶N、P含量进行调查取样。结果表明:1)南方灌丛优势木本植物叶的N、P含量几何均值分别为16.57 mg·g~(–1)和1.02 mg·g~(–1);其中,落叶木本植物(17.91 mg·g~(–1)、1.14 mg·g~(–1))显著高于常绿木本植物(15.19 mg·g~(–1)、0.89 mg·g~(–1));叶P含量较N含量具有更大的变异性和环境依赖性。2)随年平均气温(MAT)的升高,常绿木本植物叶N、P含量降低,落叶木本植物叶N、P含量呈逐渐增加的趋势;随年降水量(MAP)的增加,常绿木本植物叶N含量降低,落叶木本植物叶N含量增加,两者P含量下降。3)土壤N含量的增加对两者叶的N含量无显著影响;但随土壤P含量的增加,常绿和落叶木本植物叶P含量均显著增加。4)广义线性回归显示,植物生活型分别可解释叶N、P变异的7.6%和14.4%,MAP和土壤P含量分别解释了0.8%和16.4%的叶P变异。结果表明,中国南方灌丛中优势植物叶的N含量主要受不同生活型植物生长需求所决定,而P含量则受气候、土壤和植物生活型共同决定。 |
| [24] | . |
| [25] | . 为了解马尾松人工林窗对伴生树种的影响,为马尾松人工纯林的团块状混交提供科学依据,研究了10 m×10 m(T1)、15 m×15 m(T2)、20 m×20 m(T3)、25 m×25 m(T4)、30 m×30 m(T5)、35 m×35 m(T6)和40 m×40 m(T7)马尾松人工林窗中油樟(Cinnamomum longepaniculatum)幼苗叶片形态和光合生理特征的变化,探讨马尾松林窗斑块对混生树种生长的影响。结果表明:1)林窗面积低于20 m×20 m时,油樟幼苗叶片最大净光合速率显著低于旷地对照;2)叶片比叶重随着林窗面积的增大显著升高;林窗内油樟幼苗叶氮含量在小林窗中(10 m×10 m)显著低于旷地对照,但在大林窗中(如20 m×20 m)显著高于对照;林窗内幼苗叶磷含量则与旷地无显著性差异;3)叶氮在光合组分中的总分配系数随着林窗面积的增加而增大,其中叶氮在羧化组分中的分配系数升高尤为明显,而捕光组分的分配系数在林窗面积10 m×10 m—20 m×20 m范围内随林窗面积的增加而显著降低。可见,当马尾松林窗面积低于20 m×20 m时,林窗环境会显著影响油樟幼苗的光合能力,油樟幼苗可以通过调节比叶面积、叶氮含量以及叶氮在光合组分中的分配等形态、生理适应特征来适应林窗环境的变化。 |
| [26] | . 为探索植物叶片氮(N)、磷(P)、碳(C)生态化学计量特征随植物生长发育的变化规律,在普洱季风常绿阔叶林中,选取6种优势植物种(红锥(Castanopsis hystrix)、短刺锥(Castanopsis echidnocarpa)、泥柯(Lithocarpus fenestratus)、截果柯(Lithocarpus truncatus)、西南木荷(Schima wallichii)、茶梨(Anneslea fragrans))采集叶片,分析其N、P、C含量及化学计量比随植物生长发育的变化。结果显示:6种植物在不同生长阶段的N含量变化范围为7.90–17.72 mg·g^–1,P为0.34–1.39 mg·g^–1,C为458.48–516.87 mg·g^–1,C:N为28.04–65.70,N:P为11.41–63.50,C:P为355.23–1 878.17,且不同生长阶段6种植物及总体叶片N、P、C含量及其化学计量比变化趋势各异。在变异系数上,N:P比整体变异最大,为36.46%(变化范围19.19%–91.65%),其次为C:P,为34.80%(变化范围15.99%–91.60%),C的整体变异最小,为3.12%(变化范围1.61%–5.89%)。变异来源分析结果显示,N含量、C含量、C:N、N:P及C:P均主要受植物生长阶段的影响,而P含量主要受物种与生长阶段的交互作用影响。 |
| [27] | Soil nitrogenand phosphorus are the most important factors influenced the leaf N:P ratios across plant functional groups in the karst region of southwestern China, and plant diversity are also importa |
| [28] | A global data set including 5,087 observations of leaf nitrogen (N) and phosphorus (P) for 1,280 plant species at 452 sites and of associated mean climate indices demonstrates broad biogeographic patterns. In general, leaf N and P decline and the N/P ratio increases toward the equator as average temperature and growing season length increase. These patterns are similar for five dominant plant groups, coniferous trees and four angiosperm groups (grasses, herbs, shrubs, and trees). These results support the hypotheses that (i) leaf N and P increase from the tropics to the cooler and drier midlatitudes because of temperature-related plant physiological stoichiometry and biogeographical gradients in soil substrate age and then plateau or decrease at high latitudes because of cold temperature effects on biogeochemistry and (ii) the N/P ratio increases with mean temperature and toward the equator, because P is a major limiting nutrient in older tropical soils and N is the major limiting nutrient in younger temperate and high-latitude soils. |
| [29] | In a montane tropical forest in southwestern Colombia, we investigated how anthropogenic edges may alter bird-mediated seed dispersal from edge to forest interior as a function of edge age and presence of treefall gaps. We estimated fruit abundance and mist-netted birds at four distances from edge to forest interior (0-10, 30-40, 60-70, and 190-200 m) in three young (<12 yr) and three old (>40 yr) edges. Fruit-sampling plots (50-m2 plots) at each of the four distances were classified into gap and intact forest. Fruit abundance and frugivore capture rates varied from edge to forest interior, but such changes depended on edge age. At new edges, the total number of fruits was higher at the forest edge than at the forest interior, whereas bird captures showed the opposite trend. At old edges, the total number of fruits and bird capture rates did not vary among the four distances. In a first group of 12 plant and four bird species, the distribution of individuals in fruit (7 species) and captures (3 species) from edge to forest interior differed between old and new edges. In a second group of 18 plant and five bird species, which included those that were not amenable for a comparison between old and new edges and those that were not influenced by edge age, the distribution of individuals in fruit (12 species) and captures (3 species) was not uniform from forest edge to forest interior. Lastly, 124 plant and 19 bird species with <20 individuals in fruit and captures, respectively, were classified into very sparse and sparse species. We found that all but the sparse frugivores were more abundant at the forest edge than in the forest interior. Because very sparse and sparse plant species showed such a clear trend, we used seeds retrieved from mist-netted birds to assess potential seed movement of these species from edge to forest interior. Seeds of very sparse and sparse plant species were found both at forest "edge" (0-10 m) and at forest "interior" (the three other distances combined). Our results suggest that birds are not responding to changes in fruit abundance (resourcebase-driven mechanism). Instead, they indicate that frugivore capture rates reflect either a direct edge effect or a non-edge induced effect on birds. The apparent uncoupling of processes generating the observed patterns in fruit and frugivore abundance may affect seed dispersal in important ways. Furthermore, our results indicate that, as edges age, "edge effects" (i.e., maximum distance at which changes induced by edge creation are apparent within forest stands) change. |
| [30] | |
| [31] | . 以四川宜宾39年生马尾松人工林人工采伐形成的不同大小林窗为对象,研究林窗对土壤团聚体的组成、有机碳及活性有机碳含量和储量的影响.结果表明:土壤团聚体组成以〉2mm团聚体为主,其含量占团聚体总量的51.7%~78.7%.〉5mm土壤团聚体有机碳和活性有机碳含量与土壤总有机碳和总活性有机碳含量相关性最高,且有机碳及活性有机碳含量和储量均较高,是该地区土壤有机碳固定的特征团聚体,马尾松林窗形成后,土壤总有机碳及各团聚体有机碳含量普遍降低,但1225m^2林窗有机碳储量略高于林下;总活性有机碳含量仅225和400m^2林窗较马尾松林下高,总活性有机碳储量225、400、900和1225m^2林窗较马尾松林下高,其余面积林窗低于林下,这表明合适的林窗面积可以增加土壤有机碳及活性有机碳积累,林窗大小显著影响到团聚体的组成、有机碳及活性有机碳含量和储量,其中,1225m^2林窗土壤有机碳含量和储量均最高,活性有机碳储量也较高,且团聚体组成较好,是比较适宜的林窗面积。 |
| [32] | Soil biota are intimately tied to plant communities through herbivory and symbiosis and indirectly by the decomposition of dead organic plant material. Through both roots and aboveground organic material (e.g., leaves and wood), plants provide substantial inputs of organic matter to soil systems. Plants are the basis for most biotic soil food webs that comprise an enormous diversity of species whose multiple interactions function to help regulate nutrient cycling, which in turn influences plant growth. Many factors govern the biogeography of soil biota, including the physical and chemical properties of soil, climate, the composition and type of vegetation, and interactions with other soil biota. Despite awareness of factors influencing soil communities, no single factor allows predictions of soil animal diversity or distribution. However, research is showing that plants can have unique soil biotic communities. Degradation of soil, which removes predators and biotic regulation that occurs in less managed ecosystems, can result in increased pathogens and pests that affect humans, other animals and plants. Global changes such as land use, desertification, and soil pollution all have been shown to alter soil animal diversity and abundance. Because of our dependence on soils and plant production, studies linking soil biotic communities to primary productivity are needed to assure long-term soil sustainability. |
| [33] | . 边缘效应是生态学和生物保护的重要概念之一,它在研究生态系统尺度和景观生态系统尺度的能量流和物质流等生态过程中具有重要作用.本文对边缘效应的内涵、特征、定量评价(包括定量分析基础、强度、影响区、模型等)、应用研究等方面进行阐述,分析了边缘效应研究中存在的不足,总结了边缘效应对森林生态系统的影响及其研究方向,以期为森林经营、保护区管理等生产实践提供借鉴. |
| [34] | . |
| [35] | . |
| [36] | . |
| [37] | . ?在不同的环境条件下,植物会进行不同的生理生化反应,通过植物调节碳(C)、氮(N)、磷(P)的代谢和循环,使其具有不同的含量和分布,最终表现出特定的元素生态化学计量学特征。因此,植物主要元素的生态化学计量学特征从另一个方面体现了其对环境变化的响应和适应性。本研究对影响植物生长的主要非生物因子(包括土壤营养、CO2、水分、光照和温度)变化下植物的C、N、P生态化学计量学特征及其可能的机制进行了综合分析,并讨论了其空间和时间变异性。 |
| [38] | The above-ground standing crop and nutrient concentrations in plant material were examined in 45 stands of mire vegetation in the Biebrza peatland, Poland. The stands included flood-plains, rich fens, transitional fens and bogs. The pattern in nutrient concentrations in the above-ground plant material resembled the pattern in nutrient concentrations in peatwater and peat which had been investigated in an earlier study. Concentrations of N were quite uniform along the gradient. P-concentrations were highest in the transitional fen. Critical nutrient concentrations were defined on the basis of a review of nutrient concentrations in plant material from peatlands in which a fertilization experiment had been carried out. Defined critical values for phanerogams were: 13-14 and 0.7 mg/g dry wt for N and P respectively. Concentrations lower than these values indicate deficiency. P/N ratios ≥ 0.07 indicate N-deficiency and P/N ratios ≤ 0.04 - 0.05 indicate Pdeficiency. According to these values the Biebrza fens and bogs appear to be primarily deficient in N. The growth of the flood-plain vegetation does not appear to be restricted by nutrients. |
| [39] | |
| [40] | . 以浙江天童常绿阔叶林、常绿针叶林和落叶阔叶林为对象,通过对叶片和凋落物C:N:P比率与N、P重吸收的研究,揭示3种植被类型N、P养分限制和N、P重吸收的内在联系。结果显示:1)叶片C:N:P在常绿阔叶林为758:18:1,在常绿针叶林为678:14:1,在落叶阔叶林为338:11:1;凋落物C:N:P在常绿阔叶林为777:13:1,常绿针叶林为691:14:1,落叶阔叶林为567:14:1;2)常绿阔叶林和常绿针叶林叶片与凋落物C:N均显著高于落叶阔叶林;叶片C:P在常绿阔叶林最高,常绿针叶林中等,落叶阔叶林最低,常绿阔叶林和常绿针叶林凋落物C:P显著高于落叶阔叶林;叶片N:P比也是常绿阔叶林最高、常绿针叶林次之,落叶阔叶林最低,但常绿阔叶林凋落物N:P最低;3)植被叶片N、P含量间(N为x,P为y)的II类线性回归斜率显著大于1(p0.05),表明叶片P含量的增加可显著提高叶片N含量;凋落物N、P含量的回归斜率约等于1,反映了凋落物中单位P含量与单位N含量间的等速损耗关系;4)常绿阔叶林N重吸收率显著高于常绿针叶林与落叶阔叶林,落叶阔叶林P重吸收率显著高于常绿阔叶林和常绿针叶林。虽然植被的N:P指示常绿阔叶林受P限制,落叶阔叶林受N限制,常绿针叶林受N、P的共同限制,但是N、P重吸收研究结果表明:受N素限制的常绿阔叶林具有高的N重吸收率,受P限制的落叶阔叶林并不具有高的P重吸收率。可见,较高的N、P养分转移率可能不是植物对N、P养分胁迫的一种重要适应机制,是物种固有的特征。 |
| [41] | Ecological stoichiometry is the science studying the balance of energy and elements(i.e.C,N and P) in biological systems,providing an integrative approach to investigate the stoichiometric relationships and rules in the biogeochemical cycling and ecological processes.During the recent decade,progress has been achieved in the study on the ecological stoichiometry in terrestrial systems.Lots of efforts have been put in the research on the measurement of stoichiometric ratios in grassland,forest,wetland and arable land,etc.Concerns have been focused on the comparison of stoichiometric traits among different systems,functional groups and species,and on the coupling of the traits with environmental factors.In this review article,the advances in the study on ecological stoichiometry in grass-environment system were summarized,focusing on the stoichiometric traits in grassland system and its response to environmental factors. |
| [42] | . 选择长江上游人工采伐形成的7种不同大小的马尾松人工林林窗(G1:100 m~2;G2:225 m~2;G3:400 m~2;G4:625 m~2;G5:900 m~2;G6:1 225 m~2;G7:1 600 m~2),探讨不同林窗大小、位置和凋落物分解时间下土壤动物对凋落物微生物生物量的影响.结果显示:中等林窗(G4和G5)内,土壤动物显著影响了凋落物的微生物生物量氮(MBN)(P0.05),MBN分别增加了28.16%和26.18%.同时,林窗边缘的土壤动物使凋落物的MBN增加了29.06%(P0.05).此外,分解30 d,土壤动物使凋落物的MBN增加了26.52%(P0.05);分解90 d,土壤动物使凋落物中微生物生物量碳(MBC)显著增加了49.10%(P0.05);但在180 d时,土壤动物显著降低了MBC(P0.05).这些结果说明,在马尾松人工林林窗内马尾松凋落物分解初期,土壤动物对微生物生物量的增加有一定的促进作用,但其作用大小受到林窗大小、林窗位置和凋落物分解时间的影响而表现出一定的差异. |
| [43] | . 林窗通过改变森林微环境及土壤环境而影响凋落物难降解物质的降解, 目前关于人工林林窗对凋落物分解过程中难降解物质影响的研究较少。该文采用凋落物分解袋法, 以马尾松()人工林人工砍伐形成的7个不同面积的林窗(G1: 100 m、G2: 225 m、G3: 400 m、G4: 625 m、G5: 900 m、G6: 1 225 m、G7: 1 600 m)为研究对象, 以林下为对照, 研究了林窗大小对两种乡土树种——樟()和红椿()凋落叶分解过程中难降解物质(木质素、纤维素、总酚、缩合单宁)含量的影响。结果表明: 1)林窗大小对林窗中心红椿凋落叶缩合单宁、总酚、木质素的含量有显著影响, 对其纤维素含量和樟凋落叶中4种难降解物质含量均无显著影响。随着林窗面积的增大, 红椿凋落叶中除纤维素含量外的其余3种难降解物质含量, 中小型林窗(G1–G5, G1: 100 m, G2: 225 m, G3:400 m, G4: 625 m, G5: 900 m)低于大型林窗(G6、G7, G6: 1 225 m, G7: 1 600 m)。2)林窗不同位置, 只有红椿凋落叶中缩合单宁含量林窗中心显著低于边缘, 其余难降解物质含量和樟凋落叶中4种难降解物质含量均无显著差异。樟凋落叶的木质素含量在G3林窗显著低于林下; 红椿凋落叶除纤维素含量外的其余难降解物质含量, 中小型林窗从林窗中心到边缘均显著低于林下。3)随着分解时间的延长, 两种凋落叶都表现出缩合单宁、纤维素含量降低, 木质素含量升高, 总酚含量先升高后降低的变化趋势。研究结果表明: 中小型林窗(100–900 m)较大面积林窗干扰更有利于凋落叶中难降解物质的降解, 而林窗内的环境异质性应该是凋落物中难分解物质分解动态的主要调控因子, 并且这种效应依赖于初始凋落物质量。 |
3
2000
... 生态化学计量学(ecological stoichiometry)是研究生物系统能量平衡和多重化学元素平衡(特别是碳(C)、氮(N)、磷(P))的一门科学(
... 此外, 因林窗空间尺度不同, 使得不同面积林窗边缘拥有光照、热量和水分各异的区域小气候(
... 植物叶片元素含量及比例与自身结构特点及生长节律有密切联系(
Leaf nitrogen and phosphorus concentrations of woody plants differ in responses to climate, soil and plant growth from.
1
2013
... C含量在一定程度上可以表征植物存储C积累有机物质的能力(
马尾松人工林不同大小林窗植物多样性及其季节动态
3
2014
... 林窗作为森林中普遍存在的一种重要的干扰方式(
... 马尾松(Pinus massoniana)是我国长江上游低山丘陵区退耕还林的主要树种, 面积已超过200万hm2 (
... 叶片C:N和C:P可揭示生物量(C)和养分比值关系, 即养分利用效率, 较高的C:N和C:P代表植物对N、P的利用率较高(
Forest regeneration in artificial gaps twelve years after canopy opening in Acre State Western Amazon.
1
2011
... 马尾松(Pinus massoniana)是我国长江上游低山丘陵区退耕还林的主要树种, 面积已超过200万hm2 (
Gap ecology in the Florida scrubby flatwoods: Effects of time-since-fire, gap area, gap aggregation and microhabitat on gap species diversity.
1
2014
... 马尾松(Pinus massoniana)是我国长江上游低山丘陵区退耕还林的主要树种, 面积已超过200万hm2 (
中国毛竹林碳氮磷生态化学计量特征
2
2016
... C、N、P元素是细胞结构与功能最重要的生命元素(
... 叶片或生物量中N:P可作为生态系统限制性营养元素的一个重要指标(
Interacting effects of canopy gap, understory vegetation and leaf litter on tree seedling recruitment and composition in tropical secondary forests.
2
2008
... 林窗作为森林中普遍存在的一种重要的干扰方式(
... 马尾松(Pinus massoniana)是我国长江上游低山丘陵区退耕还林的主要树种, 面积已超过200万hm2 (
Nutritional constraints in terrestrial and freshwater food webs.
2
2000
... 生态化学计量学(ecological stoichiometry)是研究生物系统能量平衡和多重化学元素平衡(特别是碳(C)、氮(N)、磷(P))的一门科学(
... 更为重要的是, 林窗面积不同会导致林窗边缘地表接收不同强度和不同时长的阳光直射及降水分配而拥有迥异的水热模式, 进而影响油樟养分周转及分配格局.本研究中, 不同季节油樟叶片C:N和C:P随林窗面积增大先升后降, 且均在G4林窗边缘呈现最大值, 养分利用效率最高, G4林窗边缘C、N、P元素分配相对平衡, 边缘效应更显著.在植物群落中, 非生物环境的C、N和P化学计量比值一旦不相匹配, 就会导致C、N和P循环途径的不同(
Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change.
3
2010
... 生态化学计量学(ecological stoichiometry)是研究生物系统能量平衡和多重化学元素平衡(特别是碳(C)、氮(N)、磷(P))的一门科学(
... C含量在一定程度上可以表征植物存储C积累有机物质的能力(
... 叶片C:N和C:P可揭示生物量(C)和养分比值关系, 即养分利用效率, 较高的C:N和C:P代表植物对N、P的利用率较高(
Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations.
1
2015
... 更为重要的是, 林窗面积不同会导致林窗边缘地表接收不同强度和不同时长的阳光直射及降水分配而拥有迥异的水热模式, 进而影响油樟养分周转及分配格局.本研究中, 不同季节油樟叶片C:N和C:P随林窗面积增大先升后降, 且均在G4林窗边缘呈现最大值, 养分利用效率最高, G4林窗边缘C、N、P元素分配相对平衡, 边缘效应更显著.在植物群落中, 非生物环境的C、N和P化学计量比值一旦不相匹配, 就会导致C、N和P循环途径的不同(
Does light exposure make plant litter more degradable?
2010
Effects of gap size and associated changes in light and soil moisture on the understory vegetation of a Hungarian beech forest.
3
2006
... 林窗作为森林中普遍存在的一种重要的干扰方式(
... 此外, 因林窗空间尺度不同, 使得不同面积林窗边缘拥有光照、热量和水分各异的区域小气候(
... ).PCA分析表明, 油樟叶片C含量、P含量与月平均气温、月平均湿度、光强及土壤含水量有显著相关性.王丽霞等(2013)的研究表明中林隙土壤含水量最丰富, 其次为大林隙和小林隙, 中型林窗边缘更有利于植物的生长.林窗形成后疏开了上层林冠, 影响着空间及资源的组合状况, 光照强度和气流的改变影响了不同林窗边缘温湿度的变化(
Photoacceleration of plant litter decomposition in an arid environment.
1
2009
... 生态化学计量学(ecological stoichiometry)是研究生物系统能量平衡和多重化学元素平衡(特别是碳(C)、氮(N)、磷(P))的一门科学(
Influence of an old field/forest edge on a northeastern United States deciduous forest understory community.
1
1999
... 林窗作为森林中普遍存在的一种重要的干扰方式(
N:P ratios in terrestrial plants: Variation and functional significance.
2004
Variability of plant diversity and soil features following gap creation in Caspian Beech Forests of Iran.
1
2012
... 马尾松(Pinus massoniana)是我国长江上游低山丘陵区退耕还林的主要树种, 面积已超过200万hm2 (
Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China.
1
2005
... 叶片或生物量中N:P可作为生态系统限制性营养元素的一个重要指标(
Effects of forest gaps on soil properties in
1
2015
... 林窗作为森林中普遍存在的一种重要的干扰方式(
Canopy gap size influences niche partitioning of the ground- layer plant community in a northern temperate forest.
3
2013
... 生态化学计量学(ecological stoichiometry)是研究生物系统能量平衡和多重化学元素平衡(特别是碳(C)、氮(N)、磷(P))的一门科学(
... 马尾松(Pinus massoniana)是我国长江上游低山丘陵区退耕还林的主要树种, 面积已超过200万hm2 (
... 叶片C:N和C:P可揭示生物量(C)和养分比值关系, 即养分利用效率, 较高的C:N和C:P代表植物对N、P的利用率较高(
Canopy gap characteristics and tree replacement in the southeastern boreal forest.
1
1998
... 林窗作为森林中普遍存在的一种重要的干扰方式(
The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation.
1
1996
... 叶片或生物量中N:P可作为生态系统限制性营养元素的一个重要指标(
晋西黄土区不同森林树种及其林地土壤养分含量的变化
2
2016
... C、N、P元素是细胞结构与功能最重要的生命元素(
... 植物叶片元素含量及比例与自身结构特点及生长节律有密切联系(
中国南方灌丛优势木本植物叶的氮、磷含量及其影响因素
4
2017
... C、N、P元素是细胞结构与功能最重要的生命元素(
... C含量在一定程度上可以表征植物存储C积累有机物质的能力(
... ;
... 植物叶片元素含量及比例与自身结构特点及生长节律有密切联系(
边缘效应与生物多样性
1
2000
... 马尾松(Pinus massoniana)是我国长江上游低山丘陵区退耕还林的主要树种, 面积已超过200万hm2 (
油樟幼苗对马尾松林窗面积的光合响应特征
3
2015
... C含量在一定程度上可以表征植物存储C积累有机物质的能力(
... 叶片C:N和C:P可揭示生物量(C)和养分比值关系, 即养分利用效率, 较高的C:N和C:P代表植物对N、P的利用率较高(
... 更为重要的是, 林窗面积不同会导致林窗边缘地表接收不同强度和不同时长的阳光直射及降水分配而拥有迥异的水热模式, 进而影响油樟养分周转及分配格局.本研究中, 不同季节油樟叶片C:N和C:P随林窗面积增大先升后降, 且均在G4林窗边缘呈现最大值, 养分利用效率最高, G4林窗边缘C、N、P元素分配相对平衡, 边缘效应更显著.在植物群落中, 非生物环境的C、N和P化学计量比值一旦不相匹配, 就会导致C、N和P循环途径的不同(
云南普洱季风常绿阔叶林优势物种不同生长阶段叶片碳、氮、磷化学计量特征
4
2015
... C、N、P元素是细胞结构与功能最重要的生命元素(
... )(
... )(
... 植物叶片元素含量及比例与自身结构特点及生长节律有密切联系(
Leaf N:P stoichiometry across plant functional groups in the karst region of southwestern China.
1
2015
... C含量在一定程度上可以表征植物存储C积累有机物质的能力(
Global patterns of plant leaf N and P in relation to temperature and latitude.
1
2004
... 此外, 因林窗空间尺度不同, 使得不同面积林窗边缘拥有光照、热量和水分各异的区域小气候(
Anthropogenic edges, treefall gaps, and fruit-frugivore interactions in a neotropical montane forest.
1
1999
... 林窗作为森林中普遍存在的一种重要的干扰方式(
Effects of treefall gap size and age on carbon and nitrogen biogeochemical cycling in Northern hardwood-hemlock forests.
1
2011
... 叶片C:N和C:P可揭示生物量(C)和养分比值关系, 即养分利用效率, 较高的C:N和C:P代表植物对N、P的利用率较高(
马尾松人工林林窗对土壤团聚体及有机碳分布的影响
2
2014
... 林窗作为森林中普遍存在的一种重要的干扰方式(
... 此外, 因林窗空间尺度不同, 使得不同面积林窗边缘拥有光照、热量和水分各异的区域小气候(
Linking soil biodiversity and vegetation: Implications for a changing planet.
1
2011
... 此外, 因林窗空间尺度不同, 使得不同面积林窗边缘拥有光照、热量和水分各异的区域小气候(
边缘效应及其对森林生态系统影响的研究进展
2
2011
... 林窗作为森林中普遍存在的一种重要的干扰方式(
... C含量在一定程度上可以表征植物存储C积累有机物质的能力(
林窗研究进展
2
2006
... 生态化学计量学(ecological stoichiometry)是研究生物系统能量平衡和多重化学元素平衡(特别是碳(C)、氮(N)、磷(P))的一门科学(
... 马尾松(Pinus massoniana)是我国长江上游低山丘陵区退耕还林的主要树种, 面积已超过200万hm2 (
中国四种森林类型主要优势植物的C:N:P化学计量学特征
3
2011
... 马尾松(Pinus massoniana)是我国长江上游低山丘陵区退耕还林的主要树种, 面积已超过200万hm2 (
... C、N、P元素是细胞结构与功能最重要的生命元素(
... )(
红松阔叶混交林林隙大小对土壤水分空间异质性的影响
2013
植物碳氮磷生态化学计量对非生物因子的响应
1
2013
... C、N、P元素是细胞结构与功能最重要的生命元素(
Nutrient concentrations in mire vegetation as a measure of nutrient limitation in mire ecosystems.
1
1995
... 叶片或生物量中N:P可作为生态系统限制性营养元素的一个重要指标(
Forest gap dynamics and tree regeneration.
1
2000
... 林窗作为森林中普遍存在的一种重要的干扰方式(
浙江天童常绿阔叶林、常绿针叶林与落叶阔叶林的C:N:P化学计量特征
1
2010
... C、N、P元素是细胞结构与功能最重要的生命元素(
Advances in the study on ecological stoichiometry in grass-environment system and its response to environmental factors.
1
2011
... 生态化学计量学(ecological stoichiometry)是研究生物系统能量平衡和多重化学元素平衡(特别是碳(C)、氮(N)、磷(P))的一门科学(
马尾松人工林林窗内土壤动物对凋落物微生物生物量的影响
3
2016
... 马尾松(Pinus massoniana)是我国长江上游低山丘陵区退耕还林的主要树种, 面积已超过200万hm2 (
... 叶片C:N和C:P可揭示生物量(C)和养分比值关系, 即养分利用效率, 较高的C:N和C:P代表植物对N、P的利用率较高(
... 植物叶片元素含量及比例与自身结构特点及生长节律有密切联系(
马尾松人工林林窗大小对两种凋落叶难降解物质含量的影响
4
2015
... 林窗作为森林中普遍存在的一种重要的干扰方式(
... 生态化学计量学(ecological stoichiometry)是研究生物系统能量平衡和多重化学元素平衡(特别是碳(C)、氮(N)、磷(P))的一门科学(
... 叶片C:N和C:P可揭示生物量(C)和养分比值关系, 即养分利用效率, 较高的C:N和C:P代表植物对N、P的利用率较高(
... 植物叶片元素含量及比例与自身结构特点及生长节律有密切联系(


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}