Trade-off relationship between vein density and vein diameter of Achnatherum splendens in response to habitat changes in Zhangye wetland
HANLing, ZHAOCheng-Zhang*,, FENGWei, XUTing, ZHENGHui-Ling, DUANBei-Bei College of Geography and Environmental Science, Northwest Normal University, Research Center of Wetland Resources Protection and Industrial Development Engineering of Gansu Province, Lanzhou 730070, China 通讯作者:* 通信作者Author for correspondence (E-mail: zhaocz601@163.com) 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家自然科学基金(41461013和91125014)
关键词:芨芨草;光照;叶脉密度;叶脉直径;权衡;张掖湿地 Abstract Aims The coevolution between vein traits has influences on water use strategies of plant and the formation of leaf economic spectrum, and therefore is important for understanding the trade-off between carbon input in leaf vein construction and the functional feedback from leaf veins. Our aim is to study the allometric relationship between vein density and vein diameter of Achnatherum splendens populations at three natural microhabitats (subcanopy, transitional and open areas) in Zhangye wetland. Methods According to the shade condition of the arbor canopy and the distance to arbor, the A. splendens community were divided into three microenvironments: subcanopy, transitional and open areas. We sampled 10 (4 m × 4 m) A. splendens plots from each microenvironment and investigate the biological characteristics of the plots and leaf traits of the plants within the plots. Then the soil physical and chemical properties, and community photosynthetically active radiation (PAR) were investigated at three gradients. Six individual of A. splendens were selected in each plot and the leaf length, leaf width, vein density and vein diameter of two or three healthy and complete leaves from four directions of each individual were measured in laboratory. The SMA estimation method and correlation analysis were then used to examine the allometric relationship between vein density and vein diameter. Important findings Along the gradient from subcanopy, transitional zone to open areas, soil moisture displayed a pattern of initial decrease of plant community, and soil electric conductivity displayed increase changing trends. Photosynthetically active radiation (PAR), vapor pressure deficit (VPD), vein density (1.28-1.59 mm·mm-2), leaf width and water use efficiency (WUE) increase gradually, while the leaf length, vein diameter (0.21-0.16 mm) of A. splendens decrease. The average value of plasticity indexes of leaf characteristics was 0.19. Leaf net photosynthetic rate (Pn), transpiration rate (Tr) and bundle density increase first and them decrease. The vein density and vein diameter of A. splendens were negatively correlated with each other in subcanopy environment (p < 0.01), transitional and open areas (p < 0.05). The SMA (0.54-1.50) slope of regression equation in the scaling relationships between vein density and vein diameter decrease gradually from subcanopy to open areas.
不同微生境中湿地乔-草群落的生物学特征、PAR、VPD和土壤理化性质均存在显著差异(p < 0.05, 表1)。从冠盖区依次到空旷区, 芨芨草所接收的PAR由冠盖区的521.3 μmol·m-2·s-1增加到空旷区的1 411.8 μmol·m-2·s-1, 增加了1.71倍; VPD呈现逐渐增加的趋势, 增加了16.89%; 土壤含水量和土壤电导率呈相反的变化趋势, 土壤含水量减少了30.19%, 土壤电导率增加了2.68倍。表明冠盖区的土壤盐碱化程度最低, 群落内的光照环境较差, 而在空旷区土壤盐分含量高, 光照条件明显改善, 过渡区的光照环境、土壤盐分和含水量均居中, 环境胁迫强度居中。表明在3种生境中, 过渡区的生境条件更适宜芨芨草株丛的生长。 Table 1 表1 表1不同生境下湿地植物群落的生物学特征和土壤特性(平均值±标准误差, n = 30) Table 1Biological characteristics and soil characteristics of wetland plant communities in the different microenvironments (mean ± SE, n = 30)
样地 Plot
PAR (μmol·m-2·s-1)
VPD (Pa·kPa)
土壤含水量 Soil moisture (%)
土壤电导率 Soil electrical conductivity (ms·cm-1)
冠盖区 Subcanopy areas
636.30 ± 14.18c
27.24 ± 2.68c
40.07 ± 1.24a
314 ± 13.89c
过渡区 Transitional areas
879.20 ± 27.95b
29.79 ± 4.22b
37.64 ± 1.07b
669 ± 15.02b
空旷区区 Open areas
1 205.10 ± 50.75a
31.84 ± 3.34a
31.4 ± 0.67c
1 090 ± 20.45a
PAR, photosynthetically active radiation; VPD, vapor pressure deficit. Different lowercase letters in the same column indicate significant differences among plots (p < 0.05).PAR, 光合有效辐射; VPD, 水汽压亏缺。同列不同小写字母表示样地间差异显著(p < 0.05)。 新窗口打开
2.2 不同生境条件下芨芨草叶性状和光合生理参数
不同生境下芨芨草叶性状和光合生理参数的变化趋势存在显著差异(p < 0.05, 表2)。从冠盖区依次到空旷区, 芨芨草叶片宽度(LW)、叶脉密度和WUE均呈显著增加趋势, 可塑性指数均大于0.1, 三者分别增加了21.43%、24.22%和12.96%; 叶脉直径和叶片长度(LL)均呈显著的减小趋势, 分别减少了23.81%和16.25%, 与叶脉密度和叶脉直径相比, LL和LW的可塑性指数相对较小; 芨芨草种群的Pn、Tr和株丛密度(BD)均呈先增大后减小的倒U型趋势: 过渡区>空旷区>冠盖区, 其中Pn和Tr在3种微生境中的波动较小, 可塑性指数均小于0.1, 从冠盖区到空旷区三者分别增加了5.08%、4.72%和1.76倍。表型可塑性指数在不同功能性状间存在差异, 从大到小依次为: BD >叶脉直径>叶脉密度> LW > LL > WUE > Pn= Tr的趋势, 除芨芨草株丛密度在不同环境间的差异, 叶脉密度和叶脉直径的可塑性指数最大, 分别为0.20和0.24。其次是LW和LL, 分别为0.17和0.16。表明芨芨草株丛密度在3种环境下变化较显著, 且叶性状具有较高的表型可塑性, 可以较好地适应资源异质性的环境。 Table 2 表2 表2不同生境芨芨草叶性状与光合生理参数特征(平均值±标准误差, n = 30) Table 2Leaf traits characteristics and photosynthetic physiological parameters of Achnatherum splendens in the different microenvironments (mean ± SE, n = 30)
样地 Plot
冠盖区 Subcanopy areas
过渡区 Transitional areas
空旷区 Open areas
可塑性指数 Plasticity index
叶脉密度 Vein destiny (mm·mm-2)
1.28 ± 0.14c
1.46 ± 0.15b
1.59 ± 0.18a
0.20
叶脉直径 Vein diameter (mm)
0.21 ± 0.04a
0.18 ± 0.03b
0.16 ± 0.02c
0.24
叶片长度 Leaf length (cm)
58.6 ± 0.39a
51.99 ± 0.28b
49.08 ± 0.22c
0.16
叶片宽度 Leaf width (cm)
0.28 ± 0.04c
0.31 ± 0.05b
0.34 ± 0.07a
0.17
株丛密度 Bundle density (bundle·m-2)
4.25 ± 0.32c
13.50 ± 0.82b
11.75 ± 1.02a
0.64
Pn (μmol·m-2·s-1)
13.2 ± 0.12c
14.01 ± 0.18a
13.87 ± 0.13b
0.05
Tr (mmol·m-2·s-1)
6.35 ± 0.07c
6.83 ± 0.11a
6.65 ± 0.09b
0.05
WUE (μmol·mmol-1)
1.93 ± 0.01c
2.12 ± 0.02b
2.18 ± 0.03a
0.11
Pn, net photosynthetic rate; Tr, transpiration rate; WUE, water use efficiency. Different lowercase letters in the same line indicate significant differences among plots (p < 0.05).Pn, 净光合速率; Tr, 蒸腾速率; WUE, 水分利用效率。同行不同小写字母表示样地间差异显著(p < 0.05)。 新窗口打开
BlonderB, ViolleC, BentleyLP (2011). Venation networks and the origin of the leaf economics spectrum. Ecology Letters, 14, 91-100.https://doi.org/10.1111/j.1461-0248.2010.01554.xURLPMID:21073643 [本文引用: 1]摘要 Abstract The leaf economics spectrum describes biome-invariant scaling functions for leaf functional traits that relate to global primary productivity and nutrient cycling. Here, we develop a comprehensive framework for the origin of this leaf economics spectrum based on venation-mediated economic strategies. We define a standardized set of traits - density, distance and loopiness - that provides a common language for the study of venation. We develop a novel quantitative model that uses these venation traits to model leaf-level physiology, and show that selection to optimize the venation network predicts the mean global trait-trait scaling relationships across 2548 species. Furthermore, using empirical venation data for 25 plant species, we test our model by predicting four key leaf functional traits related to leaf economics: net carbon assimilation rate, life span, leaf mass per area ratio and nitrogen content. Together, these results indicate that selection on venation geometry is a fundamental basis for understanding the diversity of leaf form and function, and the carbon balance of leaves. The model and associated predictions have broad implications for integrating venation network geometry with pattern and process in ecophysiology, ecology and palaeobotany. 脗漏 2010 Blackwell Publishing Ltd/CNRS.
[2]
BrodribbTJ, BowmanDJMS, NicholsS, DelzonS, BurlettR (2010). Xylem function and growth rate interact to determine recovery rates after exposure to extreme water deficit. New Phytologist, 188, 533-542.https://doi.org/10.1111/j.1469-8137.2010.03393.xURLPMID:20673281 [本文引用: 2]摘要 61 Motivated by the urgent need to understand how water stress-induced embolism limits the survival and recovery of plants during drought, the linkage between water-stress tolerance and xylem cavitation resistance was examined in one of the world's most drought resistant conifer genera, Callitris. 61 Four species were subjected to drought treatments of -5, -8 and -10 MPa for a period of 3-4 wk, after which plants were rewatered. Transpiration, basal growth and leaf water potential were monitored during and after drought. 61 Lethal water potential was correlated with the tension producing a 50% loss of stem hydraulic conductivity. The most resilient species suffered minimal embolism and recovered gas exchange within days of rewatering from -10 MPa, while the most sensitive species suffered major embolism and recovered very slowly. The rate of repair of water transport in the latter case was equal to the rate of basal area growth, indicating xylem reiteration as the primary means of hydraulic repair. 61 The survival of, and recovery from, water stress in Callitris are accurately predicted by the physiology of the stem water-transport system. As the only apparent means of xylem repair after embolism, basal area growth is a critical part of this equation.
[3]
BrodribbTJ, JordanGJ (2008). Internal coordination between hydraulics and stomatal control in leaves. Plant, Cell & Environment, 31, 1557-1564.https://doi.org/10.1111/j.1365-3040.2008.01865.xURLPMID:18684244 [本文引用: 1]摘要 Abstract The stomatal response to changing leaf-atmospheric vapour pressure gradient (D(l)) is a crucial yet enigmatic process that defines the daily course of leaf gas exchange. Changes in the hydration of epidermal cells are thought to drive this response, mediated by the transpiration rate and hydraulic conductance of the leaf. Here, we examine whether species-specific variation in the sensitivity of leaves to perturbation of D(l) is related to the efficiency of water transport in the leaf (leaf hydraulic conductivity, K(leaf)). We found good correlation between maximum liquid (K(leaf)) and gas phase conductances (g(max)) in leaves, but there was no direct correlation between normalized D(l) sensitivity and K(leaf). The impact of K(leaf) on D(l) sensitivity in our diverse sample of eight species was important only after accounting for the strong relationship between K(leaf) and g(max). Thus, the ratio of g(max)/K(leaf) was strongly correlated with stomatal sensitivity to D(l). This ratio is an index of the degree of hydraulic buffering of the stomata against changes in D(l), and species with high g(max) relative to K(leaf) were the most sensitive to D(l) perturbation. Despite the potentially high adaptive significance of this phenomenon, we found no significant phylogenetic or ecological trend in our species.
CaiJ, TyreeMT (2010). The impact of vessel size on vulnerability curves: Data and models for within-species variability in saplings of aspen, Populus tremuloides Michx. Plant, Cell & Environment, 33, 1059-1069. [本文引用: 1]
[6]
CaiJ, ZhangSX, TyreeMT (2010). A computational algorithm addressing how vessel length might depend on vessel diameter. Plant, Cell & Environment, 33, 1234-1238.https://doi.org/10.1111/j.1365-3040.2010.02142.xURLPMID:20199614 [本文引用: 1]摘要 ABSTRACT Top of page ABSTRACT INTRODUCTION METHODS RESULTS DISCUSSION ACKNOWLEDGMENTS REFERENCES The objective of this method paper was to examine a computational algorithm that may reveal how vessel length might depend on vessel diameter within any given stem or species. The computational method requires the assumption that vessels remain approximately constant in diameter over their entire length. When this method is applied to three species or hybrids in the genus Populus , vessel length is sometimes a linear function of vessel diameter and sometimes an exponential function of vessel diameter within a stem, based on R 2 values. Our results give within-species variation of vessel length versus diameter, and we compare this to between-species variation of mean diameter versus mean length.
[7]
Cavender-BaresJ, CortesP, RambalS, JoffreR, MilesB, RocheteauA (2005). Summer and winter sensitivity of leaves and xylem to minimum freezing temperatures: A comparison of co-occurring Mediterranean oaks that differ in leaf lifespan. New Phytologist, 168, 597-612.https://doi.org/10.1111/j.1469-8137.2005.01555.xURLPMID:16313643 [本文引用: 1]摘要 Abstract Freezing sensitivity of leaves and xylem was examined in four co-occurring Mediterranean oaks (Quercus spp.) grown in a common garden to determine whether freezing responses of leaves and xylem were coordinated and could be predicted by leaf lifespan. Freezing-induced embolism and loss of photosynthetic function were measured after overnight exposure to a range of subzero temperatures in both summer and winter. Both measures were found to be dependent on minimum freezing temperature and were correlated with leaf lifespan and vessel diameter. The dependence of xylem embolism on minimum freezing temperature may result from the decline in water potential with ice temperature that influences the redistribution of water during freezing and leads to an increase in xylem tension. Winter acclimatization had a relatively small effect on the vulnerability to freezing-induced embolism, although leaf photosynthetic function showed a strong acclimatization response, particularly in the two evergreen species. Quercus ilex, the species with the longest leaf lifespan and narrowest vessel diameters, showed the highest freezing tolerance. This helps explain its ability to inhabit a broad range throughout the Mediterranean region. By contrast, the inability of the deciduous oaks to maintain photosynthetic and vascular function throughout the winter indicates a competitive disadvantage that may prevent them from expanding their ranges.
[8]
DangJJ, ZhaoCZ, LiY, HouZJ, DongXG (2014). Variations with slope in stem and leaf traits of Melica przewalskyi in alpine grassland. Chinese Journal of Plant Ecology, 38, 1307-1314.(in English with Chinese abstract) [党晶晶, 赵成章, 李钰, 侯兆疆, 董小刚 (2014). 高寒草地甘肃臭草茎-叶性状的坡度差异性. 植物生态学报, 38, 1307-1314.] [本文引用: 1]
[9]
DreznerTD (2007). An analysis of winter temperature and dew point under the canopy of a common Sonoran Desert nurse and the implications for positive plant interactions. Journal of Arid Environments, 69, 554-568.https://doi.org/10.1016/j.jaridenv.2006.11.003URL [本文引用: 1]摘要 Many studies have focused on positive-plant interactions such as nurse plants, which provide a sheltered subcanopy environment that benefits the nursed species. Most of these studies have focused on plant distribution and association patterns, while the microclimatic benefits are often assumed. This study quantifies 5 a.m. subcanopy temperatures as well as dew points beneath a common nurse tree of the Sonoran Desert, Cercidium microphyllum (palo verde, Fabaceae). Data are collected over 35 days in the winter (January and February) at six locations (at the base of the trunk, midway to the canopy edge, canopy edge, all to the north and to the south of the base of the tree) under each of two trees, as well as a control in the open. It is warmer beneath the tree than in the open, but also, it is warmer in the interior than at the canopy edge, and warmer to the south. Furthermore, differences in temperature between the subcanopy and the open site are greater on colder nights, and less pronounced on warmer winter nights, possibly due to the effect of cloud cover which often results in warmer overnight lows. In addition, variation in 5 a.m. temperature is greatest at the canopy edge and open, and temperature varies less in the interior where temperatures are also warmer. Subcanopy cover was quantified using fish-eye imagery. Results show that there is a significant relationship between 5 a.m. temperature and overlying cover. Dew point temperatures, surprisingly, were lower under the tree than at the canopy edge and in the open. That is, it is relatively dry under the nurse. This pattern can similarly be seen by distance and direction from the base of the tree (drier in the south). This may have to do with the nurse's roots and other vegetation growing beneath the nurse's canopy that compete for water in the ground, which leaves less available water to evaporate into the subcanopy air.
FunkJL, CornwellWK (2013). Leaf traits within communities: Context may affect the mapping of traits to function. Ecology, 94, 1893-1897.https://doi.org/10.1890/12-1602.1URLPMID:24279259 [本文引用: 1]摘要 The leaf economics spectrum (LES) has revolutionized the way many ecologists think about quantifying plant ecological trade-offs. In particular, the LES has connected a clear functional trade-off (long-lived leaves with slow carbon capture vs. short-lived leaves with fast carbon capture) to a handful of easily measured leaf traits. Building on this work, community ecologists are now able to quickly assess species carbon-capture strategies, which may have implications for community-level patterns such as competition or succession. However, there are a number of steps in this logic that require careful examination, and a potential danger arises when interpreting leaf-trait variation among species within communities where trait relationships are weak. Using data from 22 diverse communities, we show that relationships among three common functional traits (photosynthetic rate, leaf nitrogen concentration per mass, leaf mass per area) are weak in communities with low variation in leaf life span (LLS), especially communities dominated by herbaceous or deciduous woody species. However, globally there are few LLS data sets for communities dominated by herbaceous or deciduous species, and more data are needed to confirm this pattern. The context-dependent nature of trait relationships at the community level suggests that leaf-trait variation within communities, especially those dominated by herbaceous and deciduous woody species, should be interpreted with caution.
[12]
GivnishTJ (1987). Comparative studies of leaf form: Assessing the relative roles of selective pressures and phylogenetic constraints. New Phytologist, 106, 131-160.https://doi.org/10.1111/j.1469-8137.1987.tb04687.xURL [本文引用: 1]摘要 Leaves or their functional analogues provide outstanding opportunities for comparative studies. Here, I use leaves to illustrate the crucial role of ecological, biogeographic and phylogenetic comparisons in generating and testing hypotheses regarding the adaptive significance of morphological variation, the relative importance of selective pressures vs phylogenetic constraints, and the rise of adaptations within lineages. The complementary roles of comparative studies and optimality models are stressed throughout. The first section of this paper reviews 23 ecological patterns in leaf form, physiology and arrangement which have been uncovered by comparative studies. Three general sets of energetic trade-offs, involving the economics of gas exchange, support, and biotic interactions, appear likely to influence the evolution of leaves and underlie these trends. The first of these trade-offs is illustrated with an analysis of the adaptive significance of leaf size, in both terrestrial and aquatic plants. The resulting predictions are compared with the actual trends observed, and the relative strengths and limitations of the approach are discussed. The second section addresses the role of selective pressures and phylogenetic constraints in determining features of leaf form and phenology in forest herbs. Ecological comparisons of 74 species from a site in the Virginia Piedmont show that members of each temporal photosynthetic guild display evolutionary convergence in several aspects of leaf form and arrangement. These convergences can each be understood in terms of models that assume that selection favours plants whose form and physiology tend to maximize whole-plant growth. Phylogenetic comparisons indicate that congeners of guild members share the same leaf phenology as the guild members themselves. This remarkable finding suggests that phenology is evolutionary rather non-labile within genera but that, within guilds, species in several different genera and families converge strongly in other leaf traits. In this case, phylogenetic constraints appear to be important mainly in determining which lineages evolve particular phenologies and leaf adaptations, not whether they arise. The section concludes with a critical discussion of the capacity of two classes of evolutionary models, based primarily on functional considerations or phylogenetic constraints, to produce truly deductive predictions. The third section briefly reviews an analysis of adaptive radiation in leaf shape among violets of eastern North America. Each ecological group of species displays the leaf shape expected on functional grounds and separate lineages (as judged by means of phylogeny independent of leaf shape) show parallel trends. Ecological and phylogenetic comparisons, in combination with optimality models, provide insights into the sequence of habitats invaded and leaf forms evolved. I conclude with comments on the advantages and limitations of comparative studies, and speculate on avenues for future research on leaf form. An integrated approach, involving comparisons at ecological, phylogenetic and biogeographic levels, and complemented by optimality analyses and detailed populational studies involving biomechanics, physiological ecology and ecological genetics, is strongly advocated.
[13]
GongR, GaoQ (2015). Research progress in the effects of leaf hydraulic characteristics on plant physiological functions. Chinese Journal of Plant Ecology, 39, 300-308.(in Chinese with English abstract) [龚容, 高琼 (2015). 叶片结构的水力学特性对植物生理功能影响的研究进展. 植物生态学报, 39, 300-308.] [本文引用: 1]
[14]
HaleBK, HermsDA, HansenRC, ClausenTP, ArnoldD (2005). Effects of drought stress and nutrient availability on dry matter allocation, phenolic glycosides, and rapid induced resistance of poplar to two Lymantriid defoliators. Journal of Chemical Ecology, 31, 2601-2620.https://doi.org/10.1007/s10886-005-7616-8URLPMID:16273431 [本文引用: 4]摘要 The growth鈥揹ifferentiation balance hypothesis (GDBH) postulates that variation in resource availability can increase or decrease allocation to secondary metabolism, depending on how growth is affected relative to carbon assimilation. Growth and leaf area of black poplar ( Populus nigra ) increased substantially in response to increased nutrient availability, while net assimilation rate and photosynthesis were less strongly affected. In response, total phenolic glycoside concentrations declined, which is consistent with GDBH. Drought stress decreased net assimilation rate and photosynthesis as well as growth, while increasing total phenolic glycoside concentrations. This pattern does not follow GDBH, which predicts lower secondary metabolism when resource limitation decreases both growth and carbon assimilation. However, there was a strong negative correlation between growth and total phenolic glycoside concentration consistent with a trade-off between primary and secondary metabolism, a key premise of GDBH. Drought decreased the growth of gypsy moth ( Lymantria dispar ) larvae but had no effect on whitemarked tussock moth ( Orgyia leucostigma ). Increased nutrient availability had a positive linear effect on growth of whitemarked tussock moth, but no effect on gypsy moth. Treatment effects on gypsy moth corresponded closely with effects on total phenolic glycosides, whereas effects on whitemarked tussock moth more closely tracked changes in nutritional quality. Localized gypsy moth herbivory elicited rapid induced resistance to gypsy moth, with the effect being independent of water and nutrient availability, but did not affect whitemarked tussock moth, indicating that the effects of biotic and abiotic stress on insect resistance of trees can be species-specific.
[15]
HanL, ZhaoCZ, XuT, FengW, DuanBB, ZhengHL (2016). Trade-off between leaf size and vein density of Achnatherum splendens in Zhangye wetland. Chinese Journal of Plant Ecology, 40, 788-797.(in Chinese with English abstract) [韩玲, 赵成章, 徐婷, 冯威, 段贝贝, 郑慧玲 (2016). 张掖湿地芨芨草叶大小和叶脉密度的权衡关系. 植物生态学报, 40, 788-797.] [本文引用: 1]
[16]
HaoCS, WangQK, SunXL (2016). Effects of light heterogeneity on leaf anatomical structure in Buchloe dactyloides. Chinese Journal of Plant Ecology, 40, 246-254.(in Chinese with English abstract) [郝晨淞, 王庆凯, 孙小玲 (2016). 异质性光对野牛草叶片解剖结构的影响. 植物生态学报, 40, 246-254.] [本文引用: 1]
[17]
HaoGY, HoffmannWA, ScholzFG, BucciSJ, MeinzerFC, FrancoAC, CaoKF, GoldsteinG (2008). Stem and leaf hydraulics of congeneric tree species from adjacent tropical savanna and forest ecosystems. Oecologia, 155, 405-415.https://doi.org/10.1007/s00442-007-0918-5URLPMID:18049826摘要 Leaf and stem functional traits related to plant water relations were studied for six congeneric species pairs, each composed of one tree species typical of savanna habitats and another typical of adjacent forest habitats, to determine whether there were intrinsic differences in plant hydraulics between these two functional types. Only individuals growing in savanna habitats were studied. Most stem traits, including wood density, the xylem water potential at 50% loss of hydraulic conductivity, sapwood area specific conductivity, and leaf area specific conductivity did not differ significantly between savanna and forest species. However, maximum leaf hydraulic conductance ( K leaf ) and leaf capacitance tended to be higher in savanna species. Predawn leaf water potential and leaf mass per area were also higher in savanna species in all congeneric pairs. Hydraulic vulnerability curves of stems and leaves indicated that leaves were more vulnerable to drought-induced cavitation than terminal branches regardless of genus. The midday K leaf values estimated from leaf vulnerability curves were very low implying that daily embolism repair may occur in leaves. An electric circuit analog model predicted that, compared to forest species, savanna species took longer for their leaf water potentials to drop from predawn values to values corresponding to 50% loss of K leaf or to the turgor loss points, suggesting that savanna species were more buffered from changes in leaf water potential. The results of this study suggest that the relative success of savanna over forest species in savanna is related in part to their ability to cope with drought, which is determined more by leaf than by stem hydraulic traits. Variation among genera accounted for a large proportion of the total variance in most traits, which indicates that, despite different selective pressures in savanna and forest habitats, phylogeny has a stronger effect than habitat in determining most hydraulic traits.
[18]
HarveyPH, PagelMD (1991). The Comparative Method in Evolutionary Biology. Oxford University Press, Oxford, UK.Heberling JM, Fridley JD (2012). Biogeographic constraints on the world-wide leaf economics spectrum. Global Ecology and Biogeography, 21, 1137-1146. [本文引用: 3]
[19]
JamesSA, BellDT (2000). Influence of light availability on leaf structure and growth of two Eucalyptus globulus ssp. globulus provenances. Tree Physiology, 20, 1007-1018.https://doi.org/10.1093/treephys/20.15.1007URLPMID:11305455 [本文引用: 1]摘要 Light availability strongly affects leaf structure of the distinctive ontogenetic leaf forms of Eucalyptus globulus subsp. globulus. Plants of a late-maturing provenance from St. Marys, Tasmania and an early maturing provenance from Wilsons Promontory (Wilsons Prom.), Victoria, Australia, were grown for 9 months in 100, 50 or 10% sunlight. Growth, biomass and leaf area were significantly reduce...
[20]
LiL, ZengH, GuoDL (2013). Leaf venation functional traits and their ecological significance. Chinese Journal of Plant Ecology, 37, 691-698.(in Chinese with English abstract) [李乐, 曾辉, 郭大立 (2013). 叶脉网络功能性状及其生态学意义. 植物生态学报,37, 691-698.]https://doi.org/10.3724/SP.J.1258.2013.00072URL [本文引用: 1]摘要 叶脉网络结构是叶脉系统在叶片里的分布和排列样式。早期叶脉网络结构研究主要集中在其分类学意义上;近年来叶脉网络功能性状及其在植物水分利用上的意义己成为植物生态学研究的热点。该文介绍了叶脉网络功能性状的指标体系(包括叶脉密度、叶脉直径、叶脉之间的距离、叶脉闭合度等),综述了叶脉网络功能性状与叶脉系统功能(包括水分、养分和光合产物等物质运输、机械支撑和虫害防御等)的关系,叶脉网络功能性状与叶片其他功能性状(包括比叶重、叶寿命、光合速率、叶片大小、气孔密度等)的协同变异和权衡关系,以及叶脉网络功能性状随环境因子(包括水分、温度、光照等)的变化规律等方面的最新研究进展。此外,叶脉网络功能性状的研究成果也被应用于古环境重建、城市交通规划、流域规划及全球变化研究中。由于叶脉网络功能性状是环境因子与系统发育共同作用的结果,未来开展分子-叶片-植物-生态系统等多尺度的叶脉网络功能性状研究,理清叶脉网络功能性状与气孔失水-茎干导水-根系吸水等植物水分利用的关系,将为预测植物及生态系统对全球变化的响应提供新的启示。
[21]
LiR, DangW, CaiJ, ZhangSX, JiangZM (2016). Relationships between xylem structure and embolism vulnerability in six species of drought tolerance trees. Chinese Journal of Plant Ecology, 40, 255-263.(in Chinese with English abstract) [李荣, 党维, 蔡靖, 张硕新, 姜在民 (2016). 6个耐旱树种木质部结构与栓塞脆弱性的关系. 植物生态学报,40, 255-263.]URL [本文引用: 1]
[22]
Matthew-OgburnR, EdwardsEJ (2013). Repeated origin of three-dimensional leaf venation releases constraints on the evolution of succulence in plants. Current Biology, 23, 722-726.https://doi.org/10.1016/j.cub.2013.03.029URLPMID:23583553 [本文引用: 1]摘要 Succulent water storage is a prominent feature among plants adapted to arid zones, but we know little about how succulence evolves and how it is integrated into organs already tasked with multiple functions. Increased volume in succulent leaves, for example, may result in longer transport distances between veins and the cells that they supply, which in turn could negatively impact photosynthesis. We quantified water storage in a group of 83 closely related species to examine the evolutionary dynamics of succulence and leaf venation. In most leaves, vein density decreased with increasing succulence, resulting in significant increases in the path length of water from veins to evaporative surfaces. The most succulent leaves, however, had a distinct three-dimensional (3D) venation pattern, which evolved 11-12 times within this small lineage, likely via multiple developmental pathways. 3D venation "resets" internal leaf distances, maintaining moderate vein density in extremely succulent tissues and suggesting that the evolution of extreme succulence is constrained by the need to maintain an efficient leaf hydraulic system. The repeated evolution of 3D venation decouples leaf water storage from hydraulic path length, facilitating the evolutionary exploration of novel phenotypic space.
[23]
McCullohKA, JohnsonDM, PetitmermetJ, McNellisB, MeinzerFC, LachenbruchB (2015). A comparison of hydraulic architecture in three similarly sized woody species differing in their maximum potential height. Tree Physiology, 35, 723-731.https://doi.org/10.1093/treephys/tpv035URLPMID:25972291 [本文引用: 1]摘要 Abstract The physiological mechanisms underlying the short maximum height of shrubs are not understood. One possible explanation is that differences in the hydraulic architecture of shrubs compared with co-occurring taller trees prevent the shrubs from growing taller. To explore this hypothesis, we examined various hydraulic parameters, including vessel lumen diameter, hydraulic conductivity and vulnerability to drought-induced embolism, of three co-occurring species that differed in their maximum potential height. We examined one species of shrub, one short-statured tree and one taller tree. We worked with individuals that were approximately the same age and height, which was near the maximum for the shrub species. A number of variables correlated with the maximum potential height of the species. For example, vessel diameter and vulnerability to embolism both increased while wood density declined with maximum potential height. The difference between the pressure causing 50% reduction in hydraulic conductance in the leaves and the midday leaf water potential (the leaf's hydraulic safety margin) was much larger in the shrub than the other two species. In general, trends were consistent with understory shrubs having a more conservative life history strategy than co-occurring taller species. 脗漏 The Author 2015. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
[24]
MencucciniM (2003). The ecological significance of long-distance water transport: Short-term regulation, long- term acclimation and the hydraulic costs of stature across plant life forms. Plant, Cell & Environment, 26, 163-182.URL [本文引用: 1]
[25]
MiliosE, PipinisE, PetrouP, AkritidouS, SmirisP, AslanidouM (2007). Structure and regeneration patterns of the Juniperus excelsa Bieb. stands in the central part of the Nestos valley in the northeast of Greece, in the context of anthropogenic disturbances and nurse plant facilitation. Ecological Research, 22, 713-723.https://doi.org/10.1007/s11284-006-0310-7URL [本文引用: 1]
[26]
NardiniA, LuglioJ (2014). Leaf hydraulic capacity and drought vulnerability: Possible trade-offs and correlations with climate across three major biomes. Functional Ecology, 28, 810-818.https://doi.org/10.1111/1365-2435.12246URL [本文引用: 1]摘要 SummaryLeaf hydraulic capacity and vulnerability to drought stress are key determinants of plant competitive ability and productivity. Yet, it is not clear how these traits correlate to climatic variables across biomes and whether a trade-off exists between them.We collected leaf hydraulics data for 130 woody angiosperms from selected published articles. Species-specific values of leaf hydraulic capacity on a leaf area (Kleaf_area) and dry mass (Kleaf_mass) basis, leaf water potential inducing 50% loss of Kleaf (P50), as well as climatic variables (mean annual temperature, MAT and precipitation, MAP) for study sites were collected. Species were classified as belonging to three major biomes, that is dry sclerophyllous forests (DSF), temperate forests (TMF) and tropical forests (TRF).Significant differences were observed between biomes, with DSF species displaying the lowest hydraulic efficiency (low Kleaf) and the highest resistance to drought stress (low P50). P50 was correlated with both MAP and MAT, with species from low precipitation habitats having the lowest P50 values. Both Kleaf_area and Kleaf_mass were positively correlated with MAP and MAT. Leaf gas exchange rates were positively correlated with both Kleaf_area and Kleaf_mass. Although no correlation was found between P50 and Kleaf_area, a weak trade-off between leaf hydraulic safety and capacity emerged when P50 was plotted versus Kleaf_mass.Leaf hydraulics emerge as an important functional trait underlying plant adaptation to different habitats and contributing to shape vegetation features in different biomes.
[27]
NavasML, RoumetC, BellmannA, LaurentG, GarnierE (2010). Suites of plant traits in species from different stages of a Mediterranean secondary succession. Plant Biology, 12, 183-196.https://doi.org/10.1111/(ISSN)1438-8677URL [本文引用: 1]
PingXY, ZhouGS, SunJS (2010). Advances in the study of photosynthate allocation and its controls. Chinese Journal of Plant Ecology, 34, 100-111.(in Chinese with English abstract) [平晓燕, 周广胜, 孙敬松 (2010). 植物光合产物分配及其影响因子研究进展, 植物生态学报,34, 100-111.https://doi.org/10.3773/j.issn.1005-264x.2010.01.013URL摘要 植物光合产物分配受环境因子和生物因子的共同影响。为增进对植物对全球变化响应的理解,从植物个体水平与群落/生态系统水平综述了植物光合产物分配的影响因子与影响机理的最新研究进展。植物个体在光照增强及受水分和养分胁迫时,会将光合产物更多地分配到根系;CO2浓度升高对植物光合产物分配的影响受土壤氮素的制约,植物受氮素胁迫时,CO2浓度升高会促进光合产物更多地分配到根系;反之,对植物光合产物分配没有影响。植物群落/生态系统的光合产物分配对环境因子的响应不敏感;光合产物向根系的分配比例随其生长阶段逐渐降低。功能平衡假说、源汇关系假说和相关生长关系假说分别从环境因子、个体发育和环境因子与个体发育协同作用方面阐述了植物光合产物分配的影响机理。在此基础上,指出了未来拟重点加强的研究方向:1)生态系统尺度的光合产物向呼吸部分的分配研究:2)地下净初级生产力(belowground net primary productivity,BNPP)研究;3)温室和野外条件下及幼苗和成熟林光合产物分配对环境因子响应的比较研究;4)生态系统尺度的多因子控制试验;5)整合环境因子和个体发育对植物光合产物分配格局的影响研究。
[30]
PitmanEJG (1939). A note on normal correlation. Biometrika, 31, 9-12.https://doi.org/10.2307/2334971URL [本文引用: 4]摘要 Abstract 1." In a recent paper Finney discussed the distribution of the ratio of estimates of the two variances in a sample from a normal bi-variate population, and showed how a test for significance could be applied when the population correlation coefficient is known. He
[31]
QinFF, LiQ, CuiZM, LiHP, YangZR (2012). Leaf anatomical structures and ecological adaptabilities to light of three alfalfa cultivars with different fall dormancies under shading during overwintering. Chinese Journal of Plant Ecology, 36, 333-345.(in Chinese with English abstract) [覃凤飞, 李强, 崔棹茗, 李洪萍, 杨智然 (2012). 越冬期遮阴条件下3个不同秋眠型紫花苜蓿品种叶片解剖结构与其光生态适应性. 植物生态学报,36, 333-345.]https://doi.org/10.3724/SP.J.1258.2012.00333URL [本文引用: 1]摘要 在我国南北气候过渡地区,采用遮阴试验和石蜡切片法,研究越冬期不同光强对3个不同秋眠型紫花苜蓿(Medicago sativa)品种(‘维多利亚’、‘巨人201’和‘游客’)叶片解剖结构的影响.结果表明:随着光强减弱,各紫花苜蓿品种表皮结构中上、下表皮角质层厚度,气孔密度和气孔开度明显下降;上、下表皮厚度呈上升趋势.随着遮阴强度增加,叶肉组织中海绵组织细胞宽度显著上升,栅栏组织厚度、栅栏组织细胞层数、栅栏组织厚度/海绵组织厚度显著下降;品种间海绵组织厚度和栅栏组织细胞宽度变化趋势不一致.叶片结构整体特征中叶片厚度、叶肉厚度、中脉厚度、组织结构紧密度随光强减弱而显著下降,组织结构疏松度明显上升,叶脉突起度变化不明显.品种间各叶片解剖性状变幅及可塑性指数具有明显的差异,表明其对弱光适应方式不同.Pearson相关分析表明,各紫花苜蓿品种叶片气孔密度、栅栏组织厚度、叶肉厚度、叶片厚度及栅栏组织厚度/海绵组织厚度与光强呈显著正相关,可能是紫花苜蓿叶片解剖结构光强敏感特征参数,其中,‘维多利亚’叶片敏感特征参数与光强相关程度较低,与光强相关的性状较少.综合各项分析结果,初步确定越冬期紫花苜蓿耐阴性与其秋眠性相关,半秋眠型品种‘维多利亚’>秋眠型品种‘巨人201’≥非秋眠型品种‘游客’.
[32]
ReichPB, CornelissenH (2014). The world-wide ‘fast-slow’ plant economics spectrum: A traits manifesto. Journal of Ecology, 102, 275-301.https://doi.org/10.1111/1365-2745.12211URL [本文引用: 1]摘要 ABSTRACT The leaf economics spectrum (LES) provides a useful framework for examining species strategies as shaped by their evolutionary history. However, that spectrum, as originally described, involved only two key resources (carbon and nutrients) and one of three economically important plant organs. Herein, I evaluate whether the economics spectrum idea can be broadly extended to water – the third key resource –stems, roots and entire plants and to individual, community and ecosystem scales. My overarching hypothesis is that strong selection along trait trade-off axes, in tandem with biophysical constraints, results in convergence for any taxon on a uniformly fast, medium or slow strategy (i.e. rates of resource acquisition and processing) for all organs and all resources. Evidence for economic trait spectra exists for stems and roots as well as leaves, and for traits related to water as well as carbon and nutrients. These apply generally within and across scales (within and across communities, climate zones, biomes and lineages). There are linkages across organs and coupling among resources, resulting in an integrated whole-plant economics spectrum. Species capable of moving water rapidly have low tissue density, short tissue life span and high rates of resource acquisition and flux at organ and individual scales. The reverse is true for species with the slow strategy. Different traits may be important in different conditions, but as being fast in one respect generally requires being fast in others, being fast or slow is a general feature of species. Economic traits influence performance and fitness consistent with trait-based theory about underlying adaptive mechanisms. Traits help explain differences in growth and survival across resource gradients and thus help explain the distribution of species and the assembly of communities across light, water and nutrient gradients. Traits scale up – fast traits are associated with faster rates of ecosystem processes such as decomposition or primary productivity, and slow traits with slow process rates. Synthesis. Traits matter. A single ‘fast–slow’ plant economics spectrum that integrates across leaves, stems and roots is a key feature of the plant universe and helps to explain individual ecological strategies, community assembly processes and the functioning of ecosystems.
[33]
RenQJ, LiHL, BuHY (2015). Comparison of physiological and leaf morphological traits for photosynthesis of the 51 plant species in the Maqu alpine swamp meadow. Chinese Journal of Plant Ecology, 39, 593-603.(in Chinese with English abstract) [任青吉, 李宏林, 卜海燕 (2015). 玛曲高寒沼泽化草甸51种植物光合生理和叶片形态特征的比较. 植物生态学报,39, 593-603.]https://doi.org/10.17521/cjpe.2015.0057URL [本文引用: 2]摘要 基于功能性状的研究方法广泛地应用于生态学研究,用于解释不同层次的复杂的生态学过程,而绿色植物叶片的功能性状长期被认为对植物的生存、生长和繁殖具有重要的影响。该研究对玛曲高寒沼泽化草甸51个植物种(分属于14科)的叶片形态和光合性状进行测量,比较不同物种和不同功能群(莎草科、禾本科和双子叶类杂草)的差异,分析叶片形态特征和叶片光合性状之间的相关性。结果表明:1)不同物种、不同功能群之间在比叶面积、净光合速率和水分利用效率等叶片形态和光合特征方面有着显著的差异,例如禾本科植物具有较高的比叶面积和水分利用效率,双子叶类杂草具有较大的叶面积,而莎草科植物具有较高的净光合速率。2)相关性分析结果显示,无论在物种水平还是功能群水平,叶片形态和叶片光合性状之间都具有显著的相关关系。该研究揭示了高寒沼泽化草甸植物物种在叶片功能性状上的显著分化,进而使得这些物种能在同一个草地群落中共存,而群落中不同功能群物种的组成差异将会对群落的结构、功能和资源利用产生显著的影响。该研究将为进一步研究高寒沼泽化草甸提供基础研究数据并为其保护和恢复提供生理生态学依据。
[34]
SackL, ScoffoniC (2013). Leaf venation: Structure, function, development, evolution, ecology and applications in the past, present and future. New Phytologist, 198, 983-1000.https://doi.org/10.1111/nph.12253URLPMID:23600478 [本文引用: 4]摘要 The design and function of leaf venation are important to plant performance, with key implications for the distribution and productivity of ecosystems, and applications in paleobiology, agriculture and technology. We synthesize classical concepts and the recent literature on a wide range of aspects of leaf venation. We describe 10 major structural features that contribute to multiple key functions, and scale up to leaf and plant performance. We describe the development and plasticity of leaf venation and its adaptation across environments globally, and a new global data compilation indicating trends relating vein length per unit area to climate, growth form and habitat worldwide. We synthesize the evolution of vein traits in the major plant lineages throughout paleohistory, highlighting the multiple origins of individual traits. We summarize the strikingly diverse current applications of leaf vein research in multiple fields of science and industry. A unified core understanding will enable an increasing range of plant biologists to incorporate leaf venation into their research.
[35]
SackL, ScoffoniC, McKownAD (2012). Developmentally based scaling of leaf venation architecture explains global ecological patterns. Nature Communications, 3, 837.https://doi.org/10.1038/ncomms1835URLPMID:22588299 [本文引用: 1]摘要 Leaf size and venation show remarkable diversity across , and are key determinants of plant adaptation in ecosystems past and present. Here we present global scaling relationships of venation traits with leaf size. Across a new database for 485 globally distributed species, larger leaves had major veins of larger diameter, but lower length per leaf area, whereas minor vein traits were independent of leaf size. These scaling relationships allow estimation of intact leaf size from fragments, to improve hindcasting of past climate and biodiversity from fossil remains. The vein scaling relationships can be explained by a uniquely synthetic model for leaf anatomy and derived from published data for numerous species. Vein scaling relationships can explain the global biogeographical trend for smaller leaves in drier areas, the greater construction cost of larger leaves and the ability of to develop larger and more densely vascularised lamina to outcompete earlier-evolved plant lineages.
[36]
SellinA, ?unapuuE, KupperP (2008). Effects of light intensity and duration on leaf hydraulic conductance and distribution of resistance in shoots of silver birch (Betula pendula). Physiologia Plantarum, 134, 412-420.https://doi.org/10.1111/ppl.2008.134.issue-3URL [本文引用: 2]
van KleunenM, FischerM (2007). Progress in the detection of costs of phenotypic plasticity in plants. New Phytologist, 176, 727-730.https://doi.org/10.1111/j.1469-8137.2007.02296.xURLPMID:17997755 [本文引用: 1]摘要 ABSTRACT New Phytologist (2007) 176: 727–730 08 The Authors (2007). Journal compilation 08New Phytologist (2007)doi: 10.1111/j.1469-8137.2007.02296.x
[39]
VillagraM, CampanelloPI, BucciSJ, GoldsteinG (2013). Functional relationships between leaf hydraulics and leaf economic traits in response to nutrient addition in subtropical tree species. Tree Physiology, 33, 1308-1318.https://doi.org/10.1093/treephys/tpt098URLPMID:24284866摘要 Leaves can be both a hydraulic bottleneck and a safety valve against hydraulic catastrophic dysfunctions, and thus changes in traits related to water movement in leaves and associated costs may be critical for the success of plant growth. A 4-year fertilization experiment with nitrogen (N) and phosphorus (P) addition was done in a semideciduous Atlantic forest in northeastern Argentina. Saplings of five dominant canopy species were grown in similar gaps inside the forests (five control and five N鈥+鈥塒 addition plots). Leaf lifespan (LL), leaf mass per unit area (LMA), leaf and stem vulnerability to cavitation, leaf hydraulic conductance (K(leaf_area) and K(leaf_mass)) and leaf turgor loss point (TLP) were measured in the five species and in both treatments. Leaf lifespan tended to decrease with the addition of fertilizers, and LMA was significantly higher in plants with nutrient addition compared with individuals in control plots. The vulnerability to cavitation of leaves (P50(leaf)) either increased or decreased with the nutrient treatment depending on the species, but the average P50(leaf) did not change with nutrient addition. The P50(leaf) decreased linearly with increasing LMA and LL across species and treatments. These trade-offs have an important functional significance because more expensive (higher LMA) and less vulnerable leaves (lower P50(leaf)) are retained for a longer period of time. Osmotic potentials at TLP and at full turgor became more negative with decreasing P50(leaf) regardless of nutrient treatment. The K(leaf) on a mass basis was negatively correlated with LMA and LL, indicating that there is a carbon cost associated with increased water transport that is compensated by a longer LL. The vulnerability to cavitation of stems and leaves were similar, particularly in fertilized plants. Leaves in the species studied may not function as safety valves at low water potentials to protect the hydraulic pathway from water stress-induced cavitation. The lack of rainfall seasonality in the subtropical forest studied probably does not act as a selective pressure to enhance hydraulic segmentation between leaves and stems.
WartonDI, WrightIJ, FalsterDS, WestobyM (2006). Bivariate line-fitting methods for allometry. Biological Reviews, 81, 259-291.https://doi.org/10.1017/S1464793106007007URLPMID:16573844 [本文引用: 2]摘要 ABSTRACT Fitting a line to a bivariate dataset can be a deceptively complex problem, and there has been much debate on this issue in the literature. In this review, we describe for the practitioner the essential features of line-fitting methods for estimating the relationship between two variables: what methods are commonly used, which method should be used when, and how to make inferences from these lines to answer common research questions. A particularly important point for line-fitting in allometry is that usually, two sources of error are present (which we call measurement and equation error), and these have quite different implications for choice of line-fitting method. As a consequence, the approach in this review and the methods presented have subtle but important differences from previous reviews in the biology literature. Linear regression, major axis and standardised major axis are alternative methods that can be appropriate when there is no measurement error. When there is measurement error, this often needs to be estimated and used to adjust the variance terms in formulae for line-fitting. We also review line-fitting methods for phylogenetic analyses. Methods of inference are described for the line-fitting techniques discussed in this paper. The types of inference considered here are testing if the slope or elevation equals a given value, constructing confidence intervals for the slope or elevation, comparing several slopes or elevations, and testing for shift along the axis amongst several groups. In some cases several methods have been proposed in the literature. These are discussed and compared. In other cases there is little or no previous guidance available in the literature. Simulations were conducted to check whether the methods of inference proposed have the intended coverage probability or Type I error. We identified the methods of inference that perform well and recommend the techniques that should be adopted in future work.
WestobyM, ReichPB, WrightIJ (2013). Understanding ecological variation across species: Area-based vs. mass- based expression of leaf traits. New Phytologist, 199, 322-323.https://doi.org/10.1111/nph.12345URLPMID:23692294 [本文引用: 1]摘要 No abstract is available for this article.
[44]
WuT, GengYF, ChaiY, HaoJB, YuanCM (2014). Response of leaf anatomical structure and photosynthesis characteristics of Parthenocissus himalayana to three habitat types. Ecology and Environmental Sciences, 23, 1586-1592.(in Chinese with English abstract) [吴涛, 耿云芬, 柴勇, 郝佳波, 袁春明 (2014). 三叶爬山虎叶片解剖结构和光合生理特性对3种生境的响应. 生态环境学报,23, 1586-1592.]https://doi.org/10.3969/j.issn.1674-5906.2014.10.004URL [本文引用: 1]摘要 藤本植物生活环境的时空变化较为剧烈,为适应异质性生境常表现出较大的可塑性,其形态解剖结构及光合生理特征被认为能很好地体现对异质生境的适应。为了明确藤本植物叶片结构和光合作用对不同生境光强的响应策略,以木质藤本三叶爬山虎(Parthenocissus himalayana)为对象,采用光合仪测定和解剖显微观察的方法研究了哀牢山亚热带湿性常绿阔叶林的林外(全光照)、林缘(遮荫)和林内(荫生)3种自然生境中三叶爬山虎的叶片解剖结构和光合生理特征的变化,以期阐述三叶爬山虎对不同光环境的生态适应能力及策略,为森林生态系统的管理和物种多样性的保护及群落优化配置提供理论依据。结果表明:从林内到林外随着生境光强增加,叶片厚度(157.77~299.17μm)、上表皮厚度(21.30~28.40μm)、栅栏组织厚度(30.83~124.65μm)、栅栏组织细胞面积(430.95~652.97μm2)显著增大(P<0.01),栅栏组织细胞长度(29.23~49.54μm)和周长(86.58~155.17μm)、下表皮厚度(16.14~19.01μm)、气孔长度(24.13~27.10μm)和气孔密度(86.20~129.41个·mm-2)呈显著上升趋势(P<0.05)。栅栏组织细胞宽度(19.67~22.81μm)在3种生境中无显著差异。叶片解剖结构性状的平均可塑性值为0.37,其中最大值是栅栏组织细胞长度(0.67),最小值是气孔长度(0.11)。光饱和点(201.27~1299.17μmol·m-2·s-1)、光补偿点(3.86~29.88μmol·m-2·s-1)、饱和光强最大光合速率(2.20~12.03μmol·m-2·s-1)、暗呼吸速率(0.17~2.19μmol·m-2·s-1)、CO2补偿点(83.01~237.26μmol·m-2·s-1)、饱和CO2最大净光合速率(2.07~25.49μmol·m-2·s-1)、光呼吸速率(0.36~7.57μmol·m-2·s-1)、初始羧化效率(0.006~0.035μmol·μmol-1)随着生境光强的增高呈上升趋势,而表观量子效率(0.067~0.031μmol·μmol-1)、CO2饱和点(2062.56~1385.31μmol·m-2·s-1)呈下降趋势。与光合生理参数相关性显著的解剖性状主要是叶片厚度、上表皮厚度及其细胞周长/面积、栅栏组织细胞长度、栅栏组织厚度及维管束占叶脉面积的比例等。研究说明,三叶爬山虎在林外生境中具有明显的阳生叶特征,而在林内生境中具有明显的阴生叶特征,表现出对异质生境很强的适应性,从叶片解剖结构和光合生理特性方面解释了其在林外、林缘和林内均有分布的原因。
[45]
ZhangHX, LiS, ZhangSX, XiongXY, CaiJ (2013). Relationships between xylem structure and embolism vulnerability in four Populus clones. Scientia Silvae Sinicae, 49(5), 54-61.(in Chinese with English abstract) [张海昕, 李珊, 张硕新, 熊晓燕, 蔡靖 (2013). 4个杨树无性系木质部导管结构与栓塞脆弱性的关系. 林业科学,49(5), 54-61.]https://doi.org/10.11707/j.1001-7488.20130508URLMagsci [本文引用: 5]摘要 以杨属84K、I-101及其杂交子代02-8-21、02-9-22为研究对象,采用Cochard cavitron离心机法测定木质部栓塞脆弱性(<EM>P</EM><SUB>50</SUB>),利用染色法、硅胶注射法等测定每个无性系木质部导管的直径、长度、导管壁上纹孔膜面积,探究木质部导管结构与栓塞脆弱性的关系,以期建立杨树无性系的木质部结构指标体系。结果表明: 4个无性系枝条的栓塞脆弱性为02-8-21>I-101>02-9-22>84K,子代02-9-22比02-8-21抗栓塞。4个无性系的导管直径为02-8-21>02-9-22>84K>I-101,导管长度为02-9-22>02-8-21> I-101>84K,纹孔膜面积为02-8-21>02-9-22>I-101>84K。回归分析表明随着杨树无性系枝条导管直径和每个导管上纹孔面积的增大,其栓塞脆弱性也随之增大,显示出较强的正相关(<EM>R</EM><SUP>2</SUP>>0.7),导管长度与栓塞脆弱性显示出较弱的相关性(<EM>R</EM><SUP>2</SUP>=0.019)。导管直径大小对栓塞脆弱性起决定性作用。
[46]
ZhangMJ, LiuMS, XuC, ChiT, HongC (2012). Spatial pattern responses of Achnatherum splendens to environmental stress in different density levels. Acta Ecologica Sinica, 32, 595-604.(in Chinese with English abstract) [张明娟, 刘茂松, 徐驰, 池婷, 洪超 (2012). 不同密度条件下芨芨草空间格局对环境胁迫的响应. 生态学报,32, 595-604.]https://doi.org/10.5846/stxb201012171806URL [本文引用: 1]摘要 基于小尺度上植物间相互作用与空间格局的高度相关性,选用可避免环境异质性影响的K2点格局函数,研究了沙枣-芨芨草群落中63个芨芨草样方(3密度条件×3生境条件×7重复)中芨芨草种群在小尺度下(0—0.5 m)的空间格局,探讨了在3种密度条件(高、中、低)及3种生境条件下(冠盖区、过渡区、空旷区),芨芨草种群空间格局对土壤理化性质胁迫(盐分、有机质及容重)程度的响应。研究结果表明,就土壤盐分,养分含量及土壤容重而言,土壤理化性质胁迫程度沿冠盖区、过渡区和空旷区增加的趋势。相应地,芨芨草种群呈聚集分布的样方比例在沿冠盖区(6/21)较低,而在过渡区(11/21)和空旷区(11/21)较高。然而,在不同密度条件下,芨芨草种群空间格局对土壤性质胁迫的响应不同。在低密度条件下,芨芨草种群在冠盖区多数为聚集分布(4/7),在过渡区和空旷区全部为聚集分布;中密度条件下,芨芨草聚集分布样方比例沿冠盖区-过渡区-空旷区梯度增加(分别为2/7,3/7,4/7),但低于同等胁迫条件下低密度样方中聚集分布数量;高密度条件下,在各胁迫条件下,除了一个过渡区高密度样方,其余芨芨草种群均为随机分布。总体上,随着环境胁迫增强,在中低密度下,芨芨草种群趋向于聚集分布;但在高密度下,芨芨草种群均以随机分布为主。此外,芨芨草种群空间格局随密度变化趋势比随土壤理化性质胁迫梯度变化趋势更加明显,可能表明相对于土壤理化性质胁迫程度,芨芨草种群密度对其空间格局影响更大。因此,在考虑芨芨草种群空间格局对环境胁迫的响应时,应当考虑种群密度因素。
[47]
ZhangQM, ZhangC, LiuMS, YuW, XuC, WangHJ (2007). The influences of arboraceous layer on spatial patterns and morphological characteristics of herbaceous layer in an arid plant community. Acta Ecologica Sinica, 27, 1265-1271.https://doi.org/10.1016/S1872-2032(07)60027-4URL [本文引用: 1]摘要 Our study objective is to examine the effects of arboraceous layer on the spatial pattern and morphological features of herbaceous layer in Elacagnus angustifolia-Achnatherum splendens community in Ningxia Autonomous Region.In a 50m脳50m plot,we measured crown diameter,DBH(for E.angustifolia),height,and location of every individual by species.A 100 cm soil profile was divided into five strata at 0-10,10-30,30-60cm,and 60-100 cm in depth and sampled to measure the soil water contents,salinity,and physical and chemical properties.Wavelet analysis and buffer analysis were applied to examine the spatial relationship between the arboraceous and herbaceous layers.Additionally,three 24 m lines marked with A,B and C in the canopy projective region,and two 32 m lines marked with D and E in the open region were randomly placed for measuring individuals that touch the lines.The mean diameter,crown,height and density of A.splendens were recorded.We found that the E.angustifolia had a mean height of 7.9 m,a mean canopy diameter of 6.5 m,and the coverage up to 38.5 %,while the dominant herbaceous species,A.splendens, contributed 97.8 % of the total abundance.Soil moisture at 0-10 cm depth was only 4.67 %,but increased with depth with little variation in the 30-100 cm layers.The change in electrical conductivity was inconsistent with soil moisture and reached maximum value of 11.1脳103 S/cm in the 60-100 cm layer.This suggests that the deeper soil had lower water potential and implied that arbors roots received water mainly from deep soil,causing an anti-gradient soil salinity and water potential,while herbaceous plants tended to use water in shallow soil.Wavelet transform showed that the intrinsic scale was 6.2 m for the canopy projective regions and was 8-12 m in the open area,which corresponded well to the canopy size in the canopy projective regions and 1.5 height of E.angustifolia in the open area.Buffer analysis further suggested a more complex relationship.In the arbor canopy vertical projection area,we found no obvious differences in diameter,crown size,and height of A.splendens,but from zone 6#,the average crown diameter and the height of A.splendens increased gradually along the transect.From zone 10# to 13#,the crown diameter decreased and rapidly reverted in zone 14# and 15#.These results suggested that the control of arboraceous layer on herbaceous layer on spatial pattern and morphological characteristics were influenced not only by canopy shading,but also by other causes such as root distribution pattern as the morphological characters not changed monotonically with distance.
[48]
ZhangSB, ZhangJL, CaoKF (2016). Effects of seasonal drought on water status, leaf spectral traits and fluorescence parameters in Tarenna depauperata Hutchins, a Chinese savanna evergreen species. Plant Science Journal, 34, 117-126.(in Chinese with English abstract) [张树斌, 张教林, 曹坤芳 (2016). 季节性干旱对白皮乌口树水分状况、叶片光谱特征和荧光参数的影响. 植物科学学报,34, 117-126.] [本文引用: 1]
[49]
ZhangYF, WangW, LiangCZ, WangLX, PeiH, WangCY, WangWF (2012). Suitable habitat for the Achnatherum splendens community in typical steppe region of Inner Mongolia. Acta Ecologica Sinica, 32, 1193-1201.(in English with Chinese abstract) [张翼飞, 王炜, 梁存柱, 王立新, 裴浩, 王成燕, 王伟峰 (2012). 内蒙古典型草原区芨芨草群落适生生境. 生态学报,32, 1193-1201.]https://doi.org/10.5846/stxb201012301871URL [本文引用: 1]摘要 芨芨草群落是干旱与半干旱地区重要的植被资源,内蒙古典型草原区常在地形稍低的部位镶嵌有芨芨草群落,且芨芨草群落常与低洼湿草甸群落相连.为了探究芨芨草群落的适生生境,在锡林郭勒盟典型草原区选取了4个代表性调查区,从芨芨草群落及其相邻群落分布的地形特征、调查区的地下水埋深、1 m深土层的土壤电导率、土壤pH值和土壤质地等指标上对芨芨草适生生境进行探究,结果表明,芨芨草群落在内蒙古典型草原区常分布在河漫滩和丘间蝶形洼地地区,呈现出由高程梯度导致的与地带性针茅群落、湿草甸群落带状分布的特点,导致这种成带分布的原因与地下水埋深及地表径流有关,芨芨草群落生境的地下水埋深一般在1-3 m且有地表径流的补充;芨芨草群落耐盐碱性强,其生境也与反映土壤盐分含量的土壤电导率和土壤pH值有关;但芨芨草群落生境的形成不是由土壤机械组成所致.
[50]
ZhangYJ, CaoKF, SackL, LiN, WeiXM, GoldsteinG (2015). Extending the generality of leaf economic design principles in the cycads, an ancient lineage. New Phytologist, 206, 817-829.https://doi.org/10.1111/nph.13274URLPMID:25622799 [本文引用: 1]摘要 Abstract Cycads are the most ancient lineage of living seed plants, but the design of their leaves has received little study. We tested whether cycad leaves are governed by the same fundamental design principles previously established for ferns, conifers and angiosperms, and characterized the uniqueness of this relict lineage in foliar trait relationships. Leaf structure, photosynthesis, hydraulics and nutrient composition were studied in 33 cycad species from nine genera and three families growing in two botanical gardens. Cycads varied greatly in leaf structure and physiology. Similarly to other lineages, light-saturated photosynthetic rate per mass (A m ) was related negatively to leaf mass per area and positively to foliar concentrations of chlorophyll, nitrogen (N), phosphorus and iron, but unlike angiosperms, leaf photosynthetic rate was not associated with leaf hydraulic conductance. Cycads had lower photosynthetic N use efficiency and higher photosynthetic performance relative to hydraulic capacity compared with other lineages. These findings extend the relationships shown for foliar traits in angiosperms to the cycads. This functional convergence supports the modern synthetic understanding of leaf design, with common constraints operating across lineages, even as they highlight exceptional aspects of the biology of this key relict lineage. 漏 2015 The Authors. New Phytologist 漏 2015 New Phytologist Trust.
[51]
ZhangYQ, LiangCZ, WangW, WangLX, PengJT, YangJC, JiaCZ (2010). Soil salinity and distribution. Chinese Journal of Ecology, 29, 2438-2443.(in Chinese with English abstract) [张雅琼, 梁存柱, 王炜, 王立新, 彭江涛, 闫建成, 贾成朕 (2010). 芨芨草群落土壤盐分特征. 生态学杂志,29, 2438-2443.]URL [本文引用: 1]
[52]
ZhaoYT, XuMS, ZhangZH, ZhouLL, ZhangQQ, ArshadA, SongYJ, YanER (2016). Hydraulic architecture of evergreen broad-leaved woody plants at different successional stages in Tiantong National Forest Park, Zhejiang Province, China. Chinese Journal of Plant Ecology, 40, 116-126.(in Chinese with English abstract) [赵延涛, 许洺山, 张志浩, 周刘丽, 张晴晴, AliARSHAD, 宋彦君, 阎恩荣 (2016). 浙江天童常绿阔叶林不同演替阶段木本植物的水力结构特征. 植物生态学报,40, 116-126.]https://doi.org/10.17521/cjpe.2015.0258URL [本文引用: 1]摘要 水力结构是植物应对环境形成的与水分运输相关的形态策略.探索不同演替阶段和群落不同高度层植物的水力结 构特征,有助于理解植物的水分运输和利用策略.该研究以浙江天童常绿阔叶林演替前中后期群落的上层木(占据林冠层的树种)和下层木(灌木层物种)为对象, 测定了演替共有种(至少存在于两个演替阶段的物种)和更替种(仅存在于某一演替阶段的物种)的枝边材比导率、叶比导率和胡伯尔值,以及边材疏导面积、末端 枝总叶面积和枝条水势,分析植物水力结构在群落上层木和下层木间以及在演替阶段间的差异,及其与枝叶性状的相关关系.结果显示:(1)上层木植物边材比导 率和叶比导率显著高于下层木植物(p<0.05);(2)上层木和下层木的边材比导率与叶比导率在演替阶段间均无显著差异(p>0.05);上层木的胡伯 尔值在演替阶段间无显著差异,下层木的胡伯尔值随演替显著下降(p<0.05);(3)上层木共有种仅边材比导率随演替进行显著降低(p<0.05),更 替种的3个水力结构参数在演替阶段间无显著差异;下层木共有种水力结构参数在演替阶段间无明显差异,更替种仅胡伯尔值随演替减小(p<0.05);(4) 植物边材比导率与枝疏导面积和末端枝所支撑的总叶面积显著正相关(p<0.01),胡伯尔值与枝条水势及末端枝总叶面积显著负相关(p<0.01).以上 结果表明:天童常绿阔叶林演替各阶段上层木比下层木具有更大的输水能力和效率;随着演替进行,上层木与下层木的共有种和更替种边材比导率的相反变化表明上 层木水力结构的变化可能由微生境变化引起,而下层木水力结构特征的变化可能由物种更替造成.
[53]
ZouCM, WangYQ, LiuY, ZhangXH, TangS (2015). Responses of photosynthesis and growth to weak light regime in four legume species. Chinese Journal of Plant Ecology, 39, 909-916.(in Chinese with English abstract) [邹长明, 王允青, 刘英, 张晓红, 唐杉 (2015). 四种豆科作物的光合生理和生长发育对弱光的响应. 植物生态学报,39, 909-916.]https://doi.org/10.17521/cjpe.2015.0087URL [本文引用: 1]摘要 为了确定豆科作物作为林果行间套种作物的适宜性,于2014年通过田间试验和盆栽试验对4种豆科作物的耐阴能力及其光合生理进行了研究。测定全光和弱光处理(全光的48%)下豆科作物在初花期的叶片光合特征参数、光合色素含量与核酮糖-1,5-二磷酸羧化酶(RuBPCase)活性,监测其生长发育对弱光的响应。结果表明:弱光使供试作物的最大净光合速率、光饱和点、光补偿点、表观量子效率和暗呼吸速率不同程度地向耐阴植物变化;弱光还使供试作物的净光合速率、气孔导度、蒸腾速率、瞬时水分利用效率和RuBPCase活性不同程度地下降,而胞间CO2浓度显著上升;遮阴后,乌豇豆(Vigna cylindrica)和绿豆(Vigna radiata)的叶绿素(Chl) a和Chl b含量显著增加, Chl a/b值显著降低,大猪屎豆(Crotalaria assamica)和望江南(Senna occidentalis)的光合色素含量也有不同程度的变化;弱光使供试作物茎变细,侧枝数减少,生物量和干物质积累效率降低,根冠比降低,根瘤量减少,叶片变小变薄,叶片数减少,叶面积指数降低;弱光胁迫下,大猪屎豆不开花,望江南只开花不结实,而绿豆和乌豇豆开花数减少,花期缩短,种子产量显著降低。根据供试作物的光合特性和生长发育对弱光的响应,得出它们的耐阴能力排序是:乌豇豆>绿豆>望江南>大猪屎豆,其中乌豇豆和绿豆适宜与林果套种,而望江南和大猪屎豆不适宜套种。
[54]
ZwienieckiMA, BrodribbTJ, HolbrookNM (2007). Hydraulic design of leaves: Insights from rehydration kinetics. Plant, Cell & Environment, 30, 910-921.URL
Venation networks and the origin of the leaf economics spectrum. 1 2011
Water supply and demand remain balanced during leaf acclimation of Nothofagus cunninghamii trees. 1 2011
... 在干旱区湿地的乔-草群落中, 树冠的遮蔽作用使得草本植物所处的树冠下层常形成较好的微生境(Milios et al., 2007; 张明娟等, 2012), 如遮阴可以减少强烈的太阳辐射、促进土壤有机质积累和降低环境温度变幅等, 对植株构建最佳的生理和功能状态发挥着重要的调控作用(Zhang et al., 2007).光照是调控植物生长和分布的主要生态因子, 长期处于光照异质性分布的环境中, 植物种群间可获得性资源的数量受到抑制(党晶晶等, 2014), 进而影响植物生长发育和干物质积累效率(张明娟等, 2012; 邹长明等, 2015), 为了保证充足的光能截获和水力运输能力以维持稳定的光合作用和实现光合产物投资-收益最大化(Brodribb et al., 2010; 覃凤飞等, 2012), 植物会主动调整、转换和补偿叶脉性状的功能及状态(Navas et al., 2010), 将生物量优先分配给最易获取能力的性状是植物提高资源利用效率和风险规避能力的主要生活史对策(Heberling & Fridley, 2012; Funk & Cornwell, 2013; Westoby et al., 2013), 进而促使植物权衡在构建和维持维管束组织间的资源投入(Brodribb & Jordan, 2011), 引起叶形、叶表面特征的变化以及叶脉密度和叶脉直径的生物量权衡分配, 保证了叶片光合作用中水分运输的有效性与安全性的协调统一(Cai et al., 2010b; 吴涛等, 2014), 有利于维持植物蒸腾失水与土壤-植物-大气连续体(SPAC)系统有效供水之间的动态平衡.因此, 研究不同群落组合生境下植物叶脉密度和叶脉直径的权衡关系, 有利于理解叶脉网络建成中碳投入的经济权衡机制, 可为建立植物茎干、枝、叶水力传导系统的机理性模型奠定良好的理论基础. ...
The impact of vessel size on vulnerability curves: Data and models for within-species variability in saplings of aspen, Populus tremuloides Michx. 1 2010
A computational algorithm addressing how vessel length might depend on vessel diameter. 1 2010
... 在干旱区湿地的乔-草群落中, 树冠的遮蔽作用使得草本植物所处的树冠下层常形成较好的微生境(Milios et al., 2007; 张明娟等, 2012), 如遮阴可以减少强烈的太阳辐射、促进土壤有机质积累和降低环境温度变幅等, 对植株构建最佳的生理和功能状态发挥着重要的调控作用(Zhang et al., 2007).光照是调控植物生长和分布的主要生态因子, 长期处于光照异质性分布的环境中, 植物种群间可获得性资源的数量受到抑制(党晶晶等, 2014), 进而影响植物生长发育和干物质积累效率(张明娟等, 2012; 邹长明等, 2015), 为了保证充足的光能截获和水力运输能力以维持稳定的光合作用和实现光合产物投资-收益最大化(Brodribb et al., 2010; 覃凤飞等, 2012), 植物会主动调整、转换和补偿叶脉性状的功能及状态(Navas et al., 2010), 将生物量优先分配给最易获取能力的性状是植物提高资源利用效率和风险规避能力的主要生活史对策(Heberling & Fridley, 2012; Funk & Cornwell, 2013; Westoby et al., 2013), 进而促使植物权衡在构建和维持维管束组织间的资源投入(Brodribb & Jordan, 2011), 引起叶形、叶表面特征的变化以及叶脉密度和叶脉直径的生物量权衡分配, 保证了叶片光合作用中水分运输的有效性与安全性的协调统一(Cai et al., 2010b; 吴涛等, 2014), 有利于维持植物蒸腾失水与土壤-植物-大气连续体(SPAC)系统有效供水之间的动态平衡.因此, 研究不同群落组合生境下植物叶脉密度和叶脉直径的权衡关系, 有利于理解叶脉网络建成中碳投入的经济权衡机制, 可为建立植物茎干、枝、叶水力传导系统的机理性模型奠定良好的理论基础. ...
Summer and winter sensitivity of leaves and xylem to minimum freezing temperatures: A comparison of co-occurring Mediterranean oaks that differ in leaf lifespan. 1 2005
... 在干旱区湿地的乔-草群落中, 树冠的遮蔽作用使得草本植物所处的树冠下层常形成较好的微生境(Milios et al., 2007; 张明娟等, 2012), 如遮阴可以减少强烈的太阳辐射、促进土壤有机质积累和降低环境温度变幅等, 对植株构建最佳的生理和功能状态发挥着重要的调控作用(Zhang et al., 2007).光照是调控植物生长和分布的主要生态因子, 长期处于光照异质性分布的环境中, 植物种群间可获得性资源的数量受到抑制(党晶晶等, 2014), 进而影响植物生长发育和干物质积累效率(张明娟等, 2012; 邹长明等, 2015), 为了保证充足的光能截获和水力运输能力以维持稳定的光合作用和实现光合产物投资-收益最大化(Brodribb et al., 2010; 覃凤飞等, 2012), 植物会主动调整、转换和补偿叶脉性状的功能及状态(Navas et al., 2010), 将生物量优先分配给最易获取能力的性状是植物提高资源利用效率和风险规避能力的主要生活史对策(Heberling & Fridley, 2012; Funk & Cornwell, 2013; Westoby et al., 2013), 进而促使植物权衡在构建和维持维管束组织间的资源投入(Brodribb & Jordan, 2011), 引起叶形、叶表面特征的变化以及叶脉密度和叶脉直径的生物量权衡分配, 保证了叶片光合作用中水分运输的有效性与安全性的协调统一(Cai et al., 2010b; 吴涛等, 2014), 有利于维持植物蒸腾失水与土壤-植物-大气连续体(SPAC)系统有效供水之间的动态平衡.因此, 研究不同群落组合生境下植物叶脉密度和叶脉直径的权衡关系, 有利于理解叶脉网络建成中碳投入的经济权衡机制, 可为建立植物茎干、枝、叶水力传导系统的机理性模型奠定良好的理论基础. ...
An analysis of winter temperature and dew point under the canopy of a common Sonoran Desert nurse and the implications for positive plant interactions. 1 2007
Effects of drought stress and nutrient availability on dry matter allocation, phenolic glycosides, and rapid induced resistance of poplar to two Lymantriid defoliators. 4 2005
... 芨芨草(Achnatherum splendens)是我国西北地区多年生密丛性禾本科草本植物, 叶片较完整的机械组织和输导系统使得芨芨草具有较强的耐旱、耐盐碱和适应性强的生态可塑性, 是一种优良的饲用、水土保持和沙化盐碱地改良植物(张翼飞等, 2012; 韩玲等, 2016).近年来, 学术界对叶脉网络功能性状与光合作用(Brodribb et al., 2010)和叶片导水能力(Zwieniecki et al., 2007; Sellin et al., 2008)等的耦合关系、叶脉密度与叶大小的权衡关系(Sack et al., 2012; 韩玲等, 2016), 以及不同环境中的叶脉网络构建的碳投入(Reich & Cornelissen, 2014)等已有深入研究, 说明叶脉性状间的关联性可能受研究尺度或环境因子影响, 对芨芨草叶脉密度与叶大小的权衡关系对密度制约的响应(韩玲等, 2016), 以及芨芨草空间格局和对环境胁迫的响应(张明娟等, 2012)等已有研究, 但芨芨草叶脉性状间的生长规律研究不深入, 尤其湿地乔-草群落中芨芨草叶脉直径和叶脉密度的生长关系与叶脉构型构建机理尚不清晰.鉴于此, 我们研究了张掖洪泛平原湿地乔-草群落中芨芨草种群的叶脉直径-叶脉密度的关系及变化规律, 旨在理解湿地乔-草功能群的相互作用及对异质生境中的耐受性与适应性机制. ...
The ecological significance of long-distance water transport: Short-term regulation, long- term acclimation and the hydraulic costs of stature across plant life forms. 1 2003
... 在干旱区湿地的乔-草群落中, 树冠的遮蔽作用使得草本植物所处的树冠下层常形成较好的微生境(Milios et al., 2007; 张明娟等, 2012), 如遮阴可以减少强烈的太阳辐射、促进土壤有机质积累和降低环境温度变幅等, 对植株构建最佳的生理和功能状态发挥着重要的调控作用(Zhang et al., 2007).光照是调控植物生长和分布的主要生态因子, 长期处于光照异质性分布的环境中, 植物种群间可获得性资源的数量受到抑制(党晶晶等, 2014), 进而影响植物生长发育和干物质积累效率(张明娟等, 2012; 邹长明等, 2015), 为了保证充足的光能截获和水力运输能力以维持稳定的光合作用和实现光合产物投资-收益最大化(Brodribb et al., 2010; 覃凤飞等, 2012), 植物会主动调整、转换和补偿叶脉性状的功能及状态(Navas et al., 2010), 将生物量优先分配给最易获取能力的性状是植物提高资源利用效率和风险规避能力的主要生活史对策(Heberling & Fridley, 2012; Funk & Cornwell, 2013; Westoby et al., 2013), 进而促使植物权衡在构建和维持维管束组织间的资源投入(Brodribb & Jordan, 2011), 引起叶形、叶表面特征的变化以及叶脉密度和叶脉直径的生物量权衡分配, 保证了叶片光合作用中水分运输的有效性与安全性的协调统一(Cai et al., 2010b; 吴涛等, 2014), 有利于维持植物蒸腾失水与土壤-植物-大气连续体(SPAC)系统有效供水之间的动态平衡.因此, 研究不同群落组合生境下植物叶脉密度和叶脉直径的权衡关系, 有利于理解叶脉网络建成中碳投入的经济权衡机制, 可为建立植物茎干、枝、叶水力传导系统的机理性模型奠定良好的理论基础. ...
Structure and regeneration patterns of the Juniperus excelsa Bieb. stands in the central part of the Nestos valley in the northeast of Greece, in the context of anthropogenic disturbances and nurse plant facilitation. 1 2007
Developmentally based scaling of leaf venation architecture explains global ecological patterns. 1 2012
... 芨芨草(Achnatherum splendens)是我国西北地区多年生密丛性禾本科草本植物, 叶片较完整的机械组织和输导系统使得芨芨草具有较强的耐旱、耐盐碱和适应性强的生态可塑性, 是一种优良的饲用、水土保持和沙化盐碱地改良植物(张翼飞等, 2012; 韩玲等, 2016).近年来, 学术界对叶脉网络功能性状与光合作用(Brodribb et al., 2010)和叶片导水能力(Zwieniecki et al., 2007; Sellin et al., 2008)等的耦合关系、叶脉密度与叶大小的权衡关系(Sack et al., 2012; 韩玲等, 2016), 以及不同环境中的叶脉网络构建的碳投入(Reich & Cornelissen, 2014)等已有深入研究, 说明叶脉性状间的关联性可能受研究尺度或环境因子影响, 对芨芨草叶脉密度与叶大小的权衡关系对密度制约的响应(韩玲等, 2016), 以及芨芨草空间格局和对环境胁迫的响应(张明娟等, 2012)等已有研究, 但芨芨草叶脉性状间的生长规律研究不深入, 尤其湿地乔-草群落中芨芨草叶脉直径和叶脉密度的生长关系与叶脉构型构建机理尚不清晰.鉴于此, 我们研究了张掖洪泛平原湿地乔-草群落中芨芨草种群的叶脉直径-叶脉密度的关系及变化规律, 旨在理解湿地乔-草功能群的相互作用及对异质生境中的耐受性与适应性机制. ...
Effects of light intensity and duration on leaf hydraulic conductance and distribution of resistance in shoots of silver birch (Betula pendula). 2 2008
 , 冯威, 徐婷, 郑慧玲, 段贝贝
, 冯威, 徐婷, 郑慧玲, 段贝贝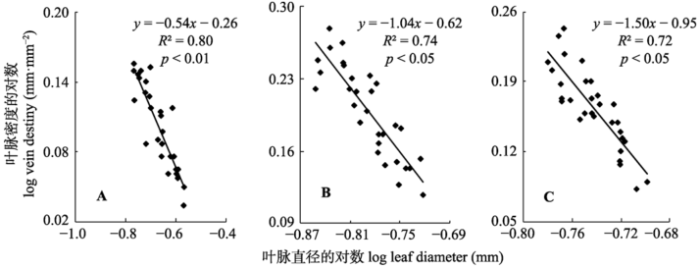 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}