Effects of nitrogen enrichment on root exudative carbon inputs in Sibiraea angustata shrubbery at the eastern fringe of Qinghai-Xizang Plateau
HEWei1,2, YANGXue-Ying3, XIAOJuan4, ZHANGZi-Liang1,2, JIANGZheng1,2, YUANYuan-Shuang1,2, WANGDong1,2, LIUQing1, YINHua-Jun1,4,*, 1Key Laboratory of Mountain Ecological Restoration and Bioresource Utilization & Ecological Restoration Biodiversity Conservation Key Laboratory of Sichuan Province, Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu 610041, China;2University of Chinese Academy of Sciences, Beijing 100049, China3Lixian Forestry Bureau, Lixian, Sichuan 623100, Chinaand 4College of Environmental Science and Engineering, China West Normal University, Nanchong, Sichuan 637000, China 通讯作者:* 通信作者Author for correspondence (E-mail: yinhj@cib.ac.cn) 收稿日期:2017-04-5 接受日期:2016-10-21 网络出版日期:2017-07-28 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:基金项目 中国科学院前沿科学重点研究项目(QYZDB-SSW-SMC023)、中国科学院青年创新促进会人才项目(20132420)、国家自然科学基金(31670449)和四川省青年科技基金(2016JQ0037)
关键词:窄叶鲜卑花灌丛;氮添加;根系分泌物;细根生物量;根系分泌物通量 Abstract Aims Understanding the responses of root exudative carbon (C) to increasing nitrogen deposition is important for predicting carbon cycling in terrestrial ecosystems. However, fewer studies have investigated the dynamics of root exudation in shrubbery ecosystems compared to forests and grassland ecosystems. This objective of this study was to determine the effects of nitrogen fertilization on the rate and C flux of root exudates.Methods Three levels of nitrogen addition treatments were applied to a Sibiraea angustata shrubbery ecosystem situated at the eastern fringe of Qinghai-Xizang Plateau, including N0 (without nitrogen application), N5 (nitrogen addition rate of 5 g·m-2·a-1), and N10 (nitrogen addition rate of 10 g·m-2·a-1), respectively, in 5 m ´ 5 m plots. Root exudates were collected in June, August and October of 2015, using a modified culture-based cuvette system. Root biomass in each plot was measured with root core method.Important findings The rates of root exudates on biomass, length, and surface area basis all displayed apparent seasonal variations during the experimental period, with the magnitude ranked in the order of: August > June > October, consistent with changes in soil temperature at 5 cm depth. With increases in the nitrogen addition rate, the rate of root exudates on biomass, length, and area basis all trended lower. Compared with the control (N0), the N5 and N10 treatments significantly reduced fine root biomass in the Sibiraea angustata shrubbery, by 23.36% and 33.84%, respectively. The decreasing root exudation and fine root biomass in response to nitrogen addition significantly decreased C flux of root exudates. Our results provide additional evidences toward a robust theoretical foundation for better understanding soil C-nutrient cycling process mediated by root exudation inputs in Alpine shrubbery ecosystems under various environmental changes.
Keywords:Sibiraea angustata shrubbery;nitrogen addition;root exudation;root biomass;root exudation flux -->0 PDF (6400KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 何为, 杨雪英, 肖娟, 张子良, 蒋铮, 袁远爽, 王东, 刘庆, 尹华军. 氮素富集对青藏高原东缘窄叶鲜卑花灌丛根系分泌物碳输入的影响. 植物生态学报, 2017, 41(6): 610-621 https://doi.org/10.17521/cjpe.2016.0329 HEWei, YANGXue-Ying, XIAOJuan, ZHANGZi-Liang, JIANGZheng, YUANYuan-Shuang, WANGDong, LIUQing, YINHua-Jun. Effects of nitrogen enrichment on root exudative carbon inputs in Sibiraea angustata shrubbery at the eastern fringe of Qinghai-Xizang Plateau. Chinese Journal of Plant Ecology, 2017, 41(6): 610-621 https://doi.org/10.17521/cjpe.2016.0329 氮(N)是影响陆地生态系统植物生长的首要营养元素, 其变化深刻改变陆地生态系统的植物生长和净初级生产力, 进而影响全球碳(C)循环和其他生态过程(LeBauer & Treseder, 2008; Kopacek et al., 2013)。近年来, 由于化石燃料的燃烧、含N化肥的大量使用等人类活动的干扰, 大气中含N化合物浓度迅速增加, 并不断向陆地和水生生态系统沉降, 从而改变了生态系统的C、N循环(Galloway et al., 2008)。研究表明, 全球亚洲区域内大气N沉降已从1961年的14.4 Tg·a-1增加到2000年的67.5 Tg·a-1, 预计到2030年将达到105.3 Tg·a-1 (Zheng et al., 2002)。目前, 大气N沉降对陆地生态系统结构和功能的影响已经成为全球变化研究的重要内容, 但纵观已有研究, 主要集中在地上部分, 如植物物候、生长、生理及其生物量分配对N沉降的响应规律与机制(Fleischer et al., 2013; Talhelm et al., 2013; Xia & Wan 2013), 而对生态系统地下部分的研究相对缺乏。根系作为连接生态系统地上与地下过程的重要纽带, 由根系活动所介导的根际生态过程不仅是生态学研究中的“瓶颈”, 也是生态系统结构、功能研究中最不确定的因素, 因而严重制约着生态系统与全球变化研究的理论拓展(贺金生等, 2004)。 近年来, 随着地下生态学研究的深入, 植物根系在调控土壤功能和养分循环过程中的重要作用逐渐被人们关注和认知(Björk et al., 2007; Volder et al., 2007)。根系分泌物是植物在生长过程中通过根系分泌的一系列化合物, 主要为一些低分子(有机酸、糖类、酚类和各种氨基酸等)和高分子(蛋白质、黏液等)有机化合物(Dijkstra & Cheng, 2007), 通常被认为是源自植物最近一段时间的光合产物, 通过体内新陈代谢, 由根系持续地输入到地下(Kuzyakov & Cheng, 2001), 并最终成为根际微生态系统中植物根系-微生物-土壤之间物质、能量和信息交流的重要纽带。植物通过根系分泌物向根际输入易于土壤微生物直接利用的含C有机物, 有效地改变根际微生物数量和活性而深刻地影响土壤有机质分解和养分转化过程(Phillips et al., 2011; Yin et al., 2013a), 从而导致根系分泌物在调控土壤C-养分转化物质循环过程中发挥着与其含量不成比例的重要作用和功能(Cheng et al., 2005)。植物根系分泌物输入及其介导下的土壤生物地球化学过程已成为地下生态学研究中的关键环节(Yin et al., 2014)。 青藏高原海拔高、气温低、太阳辐射强, 是全球气候变化显现和响应的“启动器”和“放大器” (张彦成等, 2012)。窄叶鲜卑花(Sibiraea angustata)是青藏高原东缘高山灌丛中特有的、具有代表性的物种, 分布于我国青海南部、四川西部、西藏东南部和甘肃东南部, 是该地水土保持的重要植被(吴宁, 1998)。目前, 对窄叶鲜卑花灌丛的研究主要聚焦于灌丛群落学特征(吴宁, 1998)、植物生物量(曾珍英等, 2005; 雷蕾等, 2011; 金铭等, 2012)、土壤C动态(李娇等, 2014b; 高巧等, 2014)以及生态化学计量学(贺合亮等, 2015)等方面的研究, 而对该灌丛地下过程尤其是根系C源输入及其驱动的土壤生物地球化学循环过程研究甚少, 限制了我们对高寒灌丛生态系统地下生态学过程的深入认识。因此, 本研究以青藏高原东缘窄叶鲜卑花灌丛为研究对象, 采用人工施N法模拟N沉降, 在生长季节初期(6月)、中期(8月)和末期(10月)对根系分泌物进行野外原位收集, 结合细根生物量数据, 量化不同N素富集条件下窄叶鲜卑花灌丛根系分泌物C输入速率和通量的季节动态变化规律; 同时我们也将本研究根系分泌物C输入速率与前人相关研究结果进行了初步的比较分析, 以期为全球变化背景下高寒灌丛生态系统根系分泌物C输入及其生态反馈效应研究提供理论依据。
1 材料和方法
1.1 研究区域概况
试验样地位于四川省松潘县川主寺镇章腊乡境内, 地理位置为103.52° E、32.80° N, 海拔3 300 m。本区域属高原山地气候, 春秋季短促且不明显, 冷暖季分明, 年平均气温为4.8 ℃。年降水量为693.2 mm, 降水时段主要集中在5-10月, 降水量年较差大。植被覆盖率高, 主要是温带、寒温带植物。试验对象选择以窄叶鲜卑花为优势种的灌丛群落类型, 其灌木层包括窄叶鲜卑花、红花岩生忍冬(Lonicera rupicola var. syringantha)、高山绣线菊(Spiraea alpina)和金露梅(Potentilla fruticosa), 草本层包括羊茅(Festuca ovina)、条纹龙胆(Gentiana striata)、黄花野青茅(Deyeuxia flavens)、钟花报春(Primula sikkimensis)等(李娇等, 2014b)。
在不同季节, 施N对窄叶鲜卑花灌丛根系分泌物C输入速率均有显著的抑制作用(图2, p < 0.05)。具体表现为: N10处理下单位根生物量根系分泌物C输入速率在6月、8月、10月分别为13.45、18.34和11.67 μg·g-1·h-1, 与对照(N0)相比, 分别降低了51.68%、42.44%和38.44%。N10处理使灌丛单位根长根系分泌物C输入速率在6月、8月、10月分别较对照降低了47.81%、43.78%和44.52%。与对照相比, N10处理使灌丛单位根表面积根系分泌物C输入速率在6月、8月、10月分别降低了40.56%、66.71%和39.68%。 对整个生长季节而言, 施N也显著抑制了窄叶鲜卑花灌丛根系分泌物C输入速率(图2; 表2, p < 0.001), 其抑制效应随着N处理浓度增加而加剧, 具体表现为: 与对照相比, N5和N10处理使灌丛单位根生物量根系分泌物C输入速率分别降低了12.52%和44.75%; N5、N10处理下灌丛单位根长根系分泌物C输入速率分别较对照降低7.78%和45.37%; N5、N10处理的单位根表面积根系分泌物C输入速率分别较对照降低14.69%和52.59%。 显示原图|下载原图ZIP|生成PPT 图2不同施氮(N)处理下窄叶鲜卑花灌丛根系分泌物碳(C)输入速率差异(平均值±标准偏差, n = 3)。A, 单位根生物量根系分泌物C输入速率。B, 单位根长根系分泌物C输入速率。C, 单位根表面积根系分泌物C输入速率。不同小写字母表示同一采样时间各处理间在p < 0.05水平上差异显著。N0、N5和N10为3种施氮梯度(N0: 0 g·m-2·a-1; N5: 5 g·m-2·a-1; N10: 10 g·m-2·a-1)。 -->Fig. 2Differences in root exudative carbon (C) input rates in Sibiraea angustata shrubbery among different nitrogen fertilization treatments (mean ± SD, n = 3). A, Root exudative C input rate per root biomass. B, Root exudative C input rate per root length. C, Root exudative C input rate per root surface area. Different lowercase letters indicate significant differences (p < 0.05) among treatments on a given sampling date. Three nitrogen addition levels: N0 (0 g·m-2·a-1), N5 (5 g·m-2·a-1), N10 (10 g·m-2·a-1). -->
Table 2 表2 表2窄叶鲜卑花灌丛根系分泌物碳(C)输入速率对施氮(N)和采样日期的重复测量方差分析p值 Table 2Summary of repeated measures ANOVA showing the p values for responses of root exudative carbon (C) input rates of Sibiraea angustata shrubbery to nitrogen (N) fertilization and sampling date
施N N fertilization
采样日期 Sampling date
采样日期×施N Sampling date × N fertilization
单位根生物量根系分泌物C输入速率 Root exudative C input rates per root biomass
<0.001
<0.001
0.255
单位根长根系分泌物C输入速率 Root exudative C input rates per root length
<0.001
0.019
0.844
单位根表面积根系分泌物C输入速率 Root exudative C input rates per root surface area
施N显著影响了窄叶鲜卑花灌丛土壤有效N (NO3--N和NH4+-N)含量。在不同季节, 窄叶鲜卑花灌丛土壤NO3--N和NH4+-N含量均随施N水平的提高而明显上升, 但仅N10处理下土壤NO3--N和NH4+-N含量与对照间存在显著差异(表4)。 Table 4 表4 表4不同施N水平下土壤NO3--N、NH4+-N含量(平均值±标准偏差, n =3) Table 4Changes in nitrate nitrogen and ammonium nitrogen under different nitrogen fertilization treatments (mean ± SD, n = 3)
BjörkRG, MajdiH, KlemedtssonL, LewisjonssonL, MolauU (2007). Long-term warming effects on root morphology, root mass distribution, and microbial activity in two dry tundra plant communities in northern Sweden. New Phytologist, 176, 862-673. [本文引用: 1]
[2]
ChengLS, ChengW, BianZF, LeiSG (2016). Wheat root exudation and soil nitrogen transformations in farmland reclaimed from coal gangue. Journal of Ecology and Rural Environment, 32, 397-403. (in Chinese with English abstract)[ 程林森, 程伟, 卞正富, 雷少刚 (2016). 煤矸石复垦对小麦根系分泌特征和土壤氮转化的影响 . 生态与农村环境学报, 32, 397-403.]
[3]
ChengWX, KuzyakovY, ZobelRW, WrightS (2005). Root effects on soil organic matter decomposition. In: Zobel RW, Wright SF eds. Roots and Soil Management: Interactions Between Roots and the Soil, Agronomy Monograph No. 48 .American Society of Agronomy, Madison, USA. 119-143. [本文引用: 1]
[4]
DijkstraFA, ChengWX (2007). Interactions between soil and tree roots accelerate long-term soil carbon decomposition. Ecology Letters, 10, 1046-1053. [本文引用: 1]
[5]
DrakeJE, DarbyBA, GiassonMA, KramerMA, PhillipsRP, FinziAC (2013). Stoichiometry constrains microbial response to root exudation-insights from a model and a field experiment in a temperate forest. Biogeosciences, 10, 821-838. [本文引用: 1]
[6]
FennME, PothMA, TerryJD, BlubaughTJ (2005). Nitrogen mineralization and nitrification in a mixed-conifer forest in southern California: Controlling factors, fluxes, and nitrogen fertilization response at a high and low nitrogen deposition site. Canadian Journal of Forest Research-revue Canadian de Recherche Forestiere, 35, 1464-1486. [本文引用: 1]
[7]
FleischerK, RebelKT, MolenMK, ErismanJW, WassenMJ, LoonEE, MontagnaniL, GoughCM, HerbstM, JanssensIA, GianelleD, DolmanAJ (2013). The contribution of nitrogen deposition to the photosynthetic capacity of forests. Global Biogeochemical Cycles, 27, 187-199. [本文引用: 1]
[8]
GallowayJN, TownsendAR, ErismanJW, BekundaMA, CaiZ, FreneyJR, MartinelliLA, SeitzingerSP, SuttonMA (2008). Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions. Science, 320, 889-892. [本文引用: 1]
[9]
GaoQ, YangXC, YinCY, LiuQ (2014). Estimation of biomass allocation and carbon density in alpine dwarf shrubs in Garzê Zangzu Autonomous Prefecture of Sichuan Province, China. Chinese Journal of Plant Ecology, 38, 355-365. (in Chinese with English abstract)[ 高巧, 阳小成, 尹春英, 刘庆 (2014). 四川省甘孜藏族自治州高寒矮灌丛生物量分配及其碳密度的估算 . 植物生态学报, 38, 355-365.] [本文引用: 1]
[10]
HeHL, YangXC, WangD, SunYY, YinCY, LiT, LiYX, ZhouGY, ZhangL, LiuQ (2015). Ecological stoichiometric characteristics of soil carbon, nitrogen and phosphorus of Sibiraea angustata shrub in eastern Qinghai-Tibetan Plateau. Chinese Journal of Applied Environmental Biology, 21, 1128-1135. (in Chinese with English abstract)[ 贺合亮, 阳小成, 王东, 孙誉育, 尹春英, 李婷, 黎云祥, 周国英, 张林, 刘庆 (2015). 青藏高原东部窄叶鲜卑花灌丛土壤C、N、P生态化学计量学特征 . 应用与环境生物学报, 21, 1128-1135.] [本文引用: 1]
[11]
HeJS, WangZQ, FangJY (2004). The below-ground ecology under global change: Problems and prospect. Chinese Science Bulletin, 49, 1226-1233. (in Chinese)[ 贺金生, 王政权, 方精云 (2004). 全球变化下的地下生态学: 问题与展望 . 科学通报, 49, 1226-1233.] [本文引用: 1]
[12]
HodgeA, GraysonSJ, OrdBG (1996). A novel method for characterization and quantification of plant root exudates. Plant and Soil, 184, 97-104. [本文引用: 1]
[13]
InagakiM, InagakiY, KamoK, TitinJ (2009). Fine-root production in response to nutrient application at three forest plantations in sabah, malaysia: Higher nitrogen and phosphorus demand by Acacia mangium. Journal of Forest Research, 14, 178-182.
[14]
JacquelineA, PetersonA, KalbitzK (2005). Short-term response on the quantity and quality of rhizo-deposited carbon from Norway spruce exposed to low and high N inputs. Journal of Plant Nutrition and Soil Science, 168, 687-693. [本文引用: 1]
[15]
JinM, LiY, WangSL, ZhangXL, LeiL (2012). Alpine shrubs biomass and its distribution characteristics in Qilian Mountains. Arid Land Geography, 35, 952-959. (in Chinese with English abstract)[ 金铭, 李毅, 王顺利, 张学龙, 雷蕾 (2012). 祁连山高山灌丛生物量及其分配特征 . 干旱区地理, 35, 952-959.] [本文引用: 1]
[16]
JonesDL, HodgeA, KuzyakovY (2004). Plant and mycorrhizal regulation of rhizodeposition. New Phytologist, 163, 459-480. [本文引用: 1]
[17]
JoslinJD, WolfeMH, HansonPJ (2001). Factors controlling the timing of root elongation intensity in a mature upland oak stand. Plant and Soil, 228, 201-212. [本文引用: 1]
[18]
KopacekJ, CosbyBJ, EvansCD, HruskaJD, MoldanF, OulehleF, SantrůckovaH, Tahovskak, WrightRF (2013). Nitrogen, organic carbon and sulphur cycling in terrestrial ecosystems: Linking nitrogen saturation to carbon limitation of soil microbial processes. Biogeochemistry, 115, 33-51. [本文引用: 1]
[19]
KuzyakovY, ChengWX (2001). Photosynthesis controls of rhizosphere respiration and organic matter decomposition. Soil Biology & Biochemistry, 33, 1915-1925. [本文引用: 1]
[20]
LeBauerDS, TresederKK (2008). Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology, 89, 371-379. [本文引用: 1]
[21]
LeiL, LiuXD, WangSL, LiY, ZhangXL (2011). Assignment rule of alpine shrubs biomass and its relationships to environmental factors in Qilian Mountains. Ecology and Environmental Sciences, 20, 1602-1607. (in Chinese with English abstract)[ 雷蕾, 刘贤德, 王顺利, 李毅, 张学龙 (2011). 祁连山高山灌丛生物量分配规律及其与环境因子的关系 . 生态环境学报, 20, 1602-1607.] [本文引用: 1]
[22]
LiDJ, MoJM, FANGYT, PengSL, GundersenP (2003). Impact of nitrogen deposition on forest plants. Acta Ecologica Sinica, 23, 1891-1900. (in Chinese with English abstract)[ 李德军, 莫江明, 方运霆, 彭少麟, PerGundersen (2003). 氮沉降对森林植物的影响 . 生态学报, 23, 1891-1900.] [本文引用: 1]
[23]
LiHY, ChenZH (2011). Growth and dissolved organic carbon exudates from roots of three wetland plants. Journal of Tropical and Subtropical Botany, 6, 536-542. (in Chinese with English abstract)[ 李海燕, 陈章和 (2011). 三种湿地植物的生长及根系溶解性有机碳分泌物研究 . 热带亚热带植物学报, 6, 536-542.]
[24]
LiJ, JiangXM, YinHJ, YinCY, WeiYH, LiuQ (2014a). Root exudates and soil microbes in three Picea asperata plantations with different stand ages. Chinese Journal of Applied Ecology, 25, 325-332. (in Chinese with English abstract)[ 李娇, 蒋先敏, 尹华军, 尹春英, 魏宇航, 刘庆 (2014a). 不同林龄云杉人工林的根系分泌物与土壤微生物 . 应用生态学报, 25, 325-332.]
[25]
LiJ, YinCY, ZhouXB, WeiYH, GaoQ, LiuQ (2014b). Effects of nitrogen addition on soil respiration of Sibiraea angustata shrub in the eastern margin of Qinghai-Tibetan Plateau. Acta Ecologica Sinica, 34, 5558-5569. (in Chinese with English abstract)[ 李娇, 尹春英, 周晓波, 魏宇航, 高巧, 刘庆 (2014b). 施氮对青藏高原东缘窄叶鲜卑花灌丛土壤呼吸的影响 . 生态学报, 34, 5558-5569.]
[26]
LiX, DuanZQ (2013). Progress on the research methods for root exudates. Genomics and Applied Biology, 32, 540-547. (in Chinese with English abstract)[ 李讯, 段增强 (2013). 植物根系分泌物的研究方法 . 基因组学与应用生物学, 32, 540-547.] [本文引用: 1]
[27]
LiuZX, ZhuTH, ZhangJ (2005). Research advances in root exudates and rhizosphere microorganisms of forest trees. World Forestry Research, 18(6), 25-31. (in Chinese with English abstract)[ 刘子雄, 朱天辉, 张建 (2005). 林木根系分泌物与根际微生物研究进展 . 世界林业研究, 18(6), 25-31.] [本文引用: 1]
[28]
LuCQ, TianHQ (2007). Spatial and temporal patterns of nitrogen deposition in China synthesis of observational data. Journal of Geophysical Research, 112(D22), 1-10.
[29]
LuoYQ, ZhaoXY, LiMX (2012). Ecological effect of plant root exudates and related affecting factors: A review. Chinese Journal of Applied Ecology, 23, 3496-3504. (in Chinese with English abstract)[ 罗永清, 赵学勇, 李美霞 (2012). 植物根系分泌物生态效应及其影响因素研究综述 . 应用生态学报, 23, 3496-3504.] [本文引用: 1]
[30]
MagillAH, AberJD, CurrieWS, NadelhofferKJ, MartinME, McDowellWH, MelilloJM, SteudlerP (2004). Ecosystems response to 15 years of chronic nitrogen additions at the Harvard Forest LTER, Massachusetts, USA. Forest Ecology and Management, 196, 7-28. [本文引用: 1]
[31]
MajdiH (2001). Changes in fine root production and longevity in relation to water and nutrient availability in a Norwaye spruce stand in northern Sweden. Tree Physiology, 21, 1057-1061.
[32]
MeierIC, AvisPG, PhillipsRP (2013). Fungal communities influence root exudation rates in pine seedlings. FEMS Microbiology Ecology, 83, 585-595.
[33]
NadelhofferKJ (2000). The potential effects of nitrogen deposition on fine root production in forest ecosystems. New Phytologist, 147, 131-139. [本文引用: 1]
[34]
PatersonE, GebbingT, AbelC, SimA, TelferG (2007). Rhizodeposition shapes rhizosphere microbial community structure in organic soil. New Phytologist, 173, 600-610. [本文引用: 1]
[35]
PersoneniE, NguyenC, MarchaP, PagesL (2007). Experimental evaluation of an efflux-influx model of C exudation by individual apical root segments. Journal of Experimental Botany, 58, 2091-2099.
[36]
PhillipsRP, BernhardtES, SchlesingerWH (2009). Elevated CO2 increases root exudation from loblolly pine ( Pinus taeda) seedlings as an N-mediated response. Tree Physiology, 29, 1513-1523.
[37]
PhillipsRP, FaheyTJ (2007). Fertilization effects on fine root biomass, rhizosphere microbes and respiratory fluxes in hard wood forest soils. New Phytologist, 176, 655-664. [本文引用: 1]
[38]
PhillipsRP, FinziAC, BernhardtES (2011). Enhanced root exudation induces microbial feedbacks to N cycling in a pine forest under long-term CO2 fumigation. Ecology Letters, 14, 187-194. [本文引用: 3]
[39]
QiaoMF (2015). Effects of Night Warming and Nitrogen Fertilization on Root Exudates of the Seedlings of Two Subalpine Coniferous Species in Western Sichuan. PhD dissertation, Sichuan University, Chengdu. (in Chinese with English abstract)[ 乔明锋 (2015). 增温条件下川西亚高山针叶林云杉和冷杉根系分泌物及其生态学效应研究 . 博士学位论文, 四川大学, 成都.] [本文引用: 1]
[40]
TalhelmAF, BurtonAJ, PregitzerKS, CampioneMA (2013). Chronic nitrogen deposition reduces the abundance of dominant forest understory and groundcover species. Forest Ecology and Management, 293, 39-48. [本文引用: 1]
[41]
TuSX, WuJ (2010). A review on research methods of root exudates. Ecology and Environmental Sciences, 19, 2493-2500. (in Chinese with English abstract)[ 涂书新, 吴佳 (2010). 植物根系分泌物研究方法评述 . 生态环境学报, 19, 2493-2500.]
[42]
UselmanSM, QuallsRG, ThomasRB (1999). A test of a potential short cut in the nitrogen cycle: The role of exudation of symbiotically fixed nitrogen from the roots of a N-fixing tree and the effects of increased atmospheric CO2 and temperature. Plant and Soil, 210, 21-32. [本文引用: 1]
[43]
VolderA, GiffordRM, EvansJR (2007). Effects of elevated atmospheric CO2, cutting frequency, and differential day/night atmospheric warming on root growth and turnover of phalaris swards. Global Change Biology, 13, 1040-1052. [本文引用: 1]
[44]
WellsCE, GlennDM, EissenstatD (2005). Soil insects alter fine root demography in peach ( Prunus persica). Plant, Cell & Environment, 25, 431-439. [本文引用: 1]
[45]
WhiteDC, StairJO, RingelbergDB (1996). Quantitative comparisons of in situ microbial biodiversity by signature biomarker analysis. Journal of Industrial Microbiology, 17, 185-196. [本文引用: 1]
[46]
WuN (1998). The community types and biomass of Sibiraea angustata scrub and their relationship with environmental factors in northwestern Sichuan. Acta Botanica Sinica, 40, 860-870. (in Chinese with English abstract)[ 吴宁 (1998). 川西北窄叶鲜卑花灌丛的类型和生物量及其与环境因子的关系 . 植物学报, 40, 860-870.] [本文引用: 2]
[47]
XiaJ, WanS (2013). Independent effects of warming and nitrogen addition on plant phenology in the Inner Mongolian steppe. Annals of Botany, 111, 1207-1217. [本文引用: 1]
[48]
XiaoJ (2013). Effects of Night Warming and Nitrogen Fertilization on Root Exudates of the Seedlings of Two Subalpine Coniferous Species in Western Sichuan. PhD dissertation, Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu. (in Chinese with English abstract)[ 肖娟 (2013). 夜间增温和施N对两种川西亚高山针叶树幼苗根系分泌物的影响研究 . 博士学位论文, 中国科学院成都生物研究所, 成都.] [本文引用: 1]
[49]
XiongDC, HuangJX, ChenGS, XieJS, YangYS (2015). A preliminary report on the fine root exudate of Castanopsis carlesii and Cunninghamia lanceolata in subtropical zone. Journal of Subtropical Resources and Environment, 10(1), 83-86. (in Chinese)[ 熊德成, 黄锦学, 陈光水, 谢锦升, 杨玉盛 (2015). 中亚热带米槠和杉木细根分泌物研究初报 . 亚热带资源与环境学报, 10(1), 83-86.] [本文引用: 1]
[50]
YangJH (2015). Influence of Nitrogen Deposition and Rainfall Manipulation on Concentration of Forest Tree Root Exudates. Master degree dissertation, Institute of Applied Ecology, Chinese Academy of Sciences, Shenyang. (in Chinese with English abstract)[ 杨建华 (2015). N沉降与降水变化对森林根系分泌物的影响 . 硕士学位论文, 中国科学院沈阳应用生态研究所, 沈阳.]
[51]
YinHJ, LiY, XiaoJ, XuZF, ChengXY, LiuQ (2013a). Enhanced root exudation stimulates soil nitrogen transformations in a subalpine coniferous forest under experimental warming. Global Change Biology, 19, 2158-2167. [本文引用: 1]
[52]
YinHJ, WheelerE, PhillipsRP (2014). Root-induced changes in nutrient cycling in forests depend on exudation rates. Soil Biology & Biochemistry, 78, 213-221. [本文引用: 3]
[53]
YinHJ, XiaoJ, LiYF, ChenZ, ChengXY, ZhaoCZ, LiuQ (2013b). Warming effects on root morphological and physiological traits the potential consequences on soil C dynamics as altered root exudation. Agricultural and Forest Meteorology, 180, 287-296.
[54]
ZengZY, LiuQJ, ZhangJP, ZengHQ, CaiZ (2005). A study on the pertinence of measure factors and organic biomass of shrub. Acta Agriculturae Universitatis Jiangxiensis, 27, 694-699. (in Chinese with English abstract)[ 曾珍英, 刘琪璟, 张建萍, 曾慧卿, 蔡哲 (2005). 灌木各测树因子相关性以及器官生物量相关性的研究 . 江西农业大学学报, 27, 694-699.] [本文引用: 1]
[55]
ZhangYC, HouSG, PangHX (2012). Preliminary study on spatiotemporal pattern of climate change over Tibet Plateau during past millennium. Marine Geology & Quaternary Geology, 32(3), 135-146. (in Chinese with English abstract)[ 张彦成, 侯书贵, 庞洪喜 (2012). 青藏高原地区近千年气候变化的时空特征 . 海洋地质与第四纪地质, 32(3), 135-146.] [本文引用: 1]
[56]
ZhangZL, QiaoMF, LiDD, YinHJ, LiuQ (2016). Do warming-induced changes in quantity and stoichiometry of root exudation promote soil N transformations via stimulation of soil nitrifiers, denitrifiers and ammonifiers? European Journal of Soil Biology, 74, 60-68.
[57]
ZhengXH, FuCB, XuXK, YanXD, HuangY, HanSH, HuF, ChenGX (2002). The Asian nitrogen cycle case study. AMBIO, 31, 79-87. [本文引用: 1]
[58]
ZongN, ShiPL, JiangJ, XiongDP, MengFS, SongMH, ZhangXZ, ShenZX (2013). Interactive effects of short-term nitrogen enrichment and simulated grazing on ecosystem respiration in an alpine meadow on the Tibetan Plateau. Acta Ecologica Sinica, 33, 6191-6201. (in Chinese with English abstract)[ 宗宁, 石培礼, 蒋婧, 熊定鹏, 孟丰收, 宋明华, 张宪洲, 沈振西 (2013). 短期N素添加和模拟放牧对青藏高原高寒草甸生态系统呼吸的影响 . 生态学报, 33, 6191-6201.] [本文引用: 1]
Long-term warming effects on root morphology, root mass distribution, and microbial activity in two dry tundra plant communities in northern Sweden. 1 2007
... 近年来, 随着地下生态学研究的深入, 植物根系在调控土壤功能和养分循环过程中的重要作用逐渐被人们关注和认知(Björk et al., 2007; Volder et al., 2007).根系分泌物是植物在生长过程中通过根系分泌的一系列化合物, 主要为一些低分子(有机酸、糖类、酚类和各种氨基酸等)和高分子(蛋白质、黏液等)有机化合物(Dijkstra & Cheng, 2007), 通常被认为是源自植物最近一段时间的光合产物, 通过体内新陈代谢, 由根系持续地输入到地下(Kuzyakov & Cheng, 2001), 并最终成为根际微生态系统中植物根系-微生物-土壤之间物质、能量和信息交流的重要纽带.植物通过根系分泌物向根际输入易于土壤微生物直接利用的含C有机物, 有效地改变根际微生物数量和活性而深刻地影响土壤有机质分解和养分转化过程(Phillips et al., 2011; Yin et al., 2013a), 从而导致根系分泌物在调控土壤C-养分转化物质循环过程中发挥着与其含量不成比例的重要作用和功能(Cheng et al., 2005).植物根系分泌物输入及其介导下的土壤生物地球化学过程已成为地下生态学研究中的关键环节(Yin et al., 2014). ...
煤矸石复垦对小麦根系分泌特征和土壤氮转化的影响 2016
Root effects on soil organic matter decomposition. In: Zobel RW, Wright SF eds. Roots and Soil Management: Interactions Between Roots and the Soil, Agronomy Monograph No. 48 1
... 近年来, 随着地下生态学研究的深入, 植物根系在调控土壤功能和养分循环过程中的重要作用逐渐被人们关注和认知(Björk et al., 2007; Volder et al., 2007).根系分泌物是植物在生长过程中通过根系分泌的一系列化合物, 主要为一些低分子(有机酸、糖类、酚类和各种氨基酸等)和高分子(蛋白质、黏液等)有机化合物(Dijkstra & Cheng, 2007), 通常被认为是源自植物最近一段时间的光合产物, 通过体内新陈代谢, 由根系持续地输入到地下(Kuzyakov & Cheng, 2001), 并最终成为根际微生态系统中植物根系-微生物-土壤之间物质、能量和信息交流的重要纽带.植物通过根系分泌物向根际输入易于土壤微生物直接利用的含C有机物, 有效地改变根际微生物数量和活性而深刻地影响土壤有机质分解和养分转化过程(Phillips et al., 2011; Yin et al., 2013a), 从而导致根系分泌物在调控土壤C-养分转化物质循环过程中发挥着与其含量不成比例的重要作用和功能(Cheng et al., 2005).植物根系分泌物输入及其介导下的土壤生物地球化学过程已成为地下生态学研究中的关键环节(Yin et al., 2014). ...
Interactions between soil and tree roots accelerate long-term soil carbon decomposition. 1 2007
... 近年来, 随着地下生态学研究的深入, 植物根系在调控土壤功能和养分循环过程中的重要作用逐渐被人们关注和认知(Björk et al., 2007; Volder et al., 2007).根系分泌物是植物在生长过程中通过根系分泌的一系列化合物, 主要为一些低分子(有机酸、糖类、酚类和各种氨基酸等)和高分子(蛋白质、黏液等)有机化合物(Dijkstra & Cheng, 2007), 通常被认为是源自植物最近一段时间的光合产物, 通过体内新陈代谢, 由根系持续地输入到地下(Kuzyakov & Cheng, 2001), 并最终成为根际微生态系统中植物根系-微生物-土壤之间物质、能量和信息交流的重要纽带.植物通过根系分泌物向根际输入易于土壤微生物直接利用的含C有机物, 有效地改变根际微生物数量和活性而深刻地影响土壤有机质分解和养分转化过程(Phillips et al., 2011; Yin et al., 2013a), 从而导致根系分泌物在调控土壤C-养分转化物质循环过程中发挥着与其含量不成比例的重要作用和功能(Cheng et al., 2005).植物根系分泌物输入及其介导下的土壤生物地球化学过程已成为地下生态学研究中的关键环节(Yin et al., 2014). ...
Stoichiometry constrains microbial response to root exudation-insights from a model and a field experiment in a temperate forest. 1 2013
... N素富集对窄叶鲜卑花灌丛根系分泌物C输入速率的抑制作用可通过如下机制来解释: (1)导致窄叶鲜卑花灌丛根系分泌物C输入速率降低的一个重要原因可能与N富集下植物根系生长抑制有关.研究表明, 植物根系分泌物输入与根系生长及生理活性密切相关(刘子雄等, 2005).一般地, N沉降在短期内通常会促进植物地上部分生物量的积累, 抑制植物地下根系的生长(Fenn et al., 2005; Phillips & Fahey, 2007), 从而导致细根生物量和地下C分配降低(Magill et al., 2004).本研究中N素富集下窄叶鲜卑花灌丛细根生物量降低这一结果(图4)也进一步佐证了上述推测.(2) N富集所导致的窄叶鲜卑花灌丛根系对养分的微生物需求变化可能也是植物根系分泌物C输入速率降低的重要原因.根系分泌物是驱动根际微生物群落组成和活性的重要因子, 在调控土壤生物地球化学过程中具有重要的生态学作用(Drake et al., 2013).本研究中, 窄叶鲜卑花灌丛受气候条件的限制, 土壤有机质分解缓慢, 养分匮乏, 属于典型N素受限的高寒生态系统类型(宗宁等, 2013).在非施肥条件下, 由于受土壤N有效性的限制, 土壤微生物将根系分泌物输入的C作为能源, 以刺激和促进土壤有机质分解和N素转化, 从而满足植物和微生物生长所需要的养分(Jones et al., 2004; Phillips et al., 2011).相反, 持续而高强度的N素富集在一定程度上增加了窄叶鲜卑花灌丛土壤有效N水平, 使得植物能更轻易地获取所需的养分, 从而缓和了植物根系对养分的微生物驱动需求.在此情况下, 植物采取了低N收益-低C投入的养分获取策略(Phillips et al., 2011; Yin et al., 2014), 即植物在面对N素富集时通过自我生理调节降低了根系分泌物C输入.本研究中N素富集下窄叶鲜卑花土壤有效N含量(表4)与微生物PLFAs总量(图5)降低也进一步佐证了我们的推测. ...
Nitrogen mineralization and nitrification in a mixed-conifer forest in southern California: Controlling factors, fluxes, and nitrogen fertilization response at a high and low nitrogen deposition site. 1 2005
... N素富集对窄叶鲜卑花灌丛根系分泌物C输入速率的抑制作用可通过如下机制来解释: (1)导致窄叶鲜卑花灌丛根系分泌物C输入速率降低的一个重要原因可能与N富集下植物根系生长抑制有关.研究表明, 植物根系分泌物输入与根系生长及生理活性密切相关(刘子雄等, 2005).一般地, N沉降在短期内通常会促进植物地上部分生物量的积累, 抑制植物地下根系的生长(Fenn et al., 2005; Phillips & Fahey, 2007), 从而导致细根生物量和地下C分配降低(Magill et al., 2004).本研究中N素富集下窄叶鲜卑花灌丛细根生物量降低这一结果(图4)也进一步佐证了上述推测.(2) N富集所导致的窄叶鲜卑花灌丛根系对养分的微生物需求变化可能也是植物根系分泌物C输入速率降低的重要原因.根系分泌物是驱动根际微生物群落组成和活性的重要因子, 在调控土壤生物地球化学过程中具有重要的生态学作用(Drake et al., 2013).本研究中, 窄叶鲜卑花灌丛受气候条件的限制, 土壤有机质分解缓慢, 养分匮乏, 属于典型N素受限的高寒生态系统类型(宗宁等, 2013).在非施肥条件下, 由于受土壤N有效性的限制, 土壤微生物将根系分泌物输入的C作为能源, 以刺激和促进土壤有机质分解和N素转化, 从而满足植物和微生物生长所需要的养分(Jones et al., 2004; Phillips et al., 2011).相反, 持续而高强度的N素富集在一定程度上增加了窄叶鲜卑花灌丛土壤有效N水平, 使得植物能更轻易地获取所需的养分, 从而缓和了植物根系对养分的微生物驱动需求.在此情况下, 植物采取了低N收益-低C投入的养分获取策略(Phillips et al., 2011; Yin et al., 2014), 即植物在面对N素富集时通过自我生理调节降低了根系分泌物C输入.本研究中N素富集下窄叶鲜卑花土壤有效N含量(表4)与微生物PLFAs总量(图5)降低也进一步佐证了我们的推测. ...
The contribution of nitrogen deposition to the photosynthetic capacity of forests. 1 2013
... 氮(N)是影响陆地生态系统植物生长的首要营养元素, 其变化深刻改变陆地生态系统的植物生长和净初级生产力, 进而影响全球碳(C)循环和其他生态过程(LeBauer & Treseder, 2008; Kopacek et al., 2013).近年来, 由于化石燃料的燃烧、含N化肥的大量使用等人类活动的干扰, 大气中含N化合物浓度迅速增加, 并不断向陆地和水生生态系统沉降, 从而改变了生态系统的C、N循环(Galloway et al., 2008).研究表明, 全球亚洲区域内大气N沉降已从1961年的14.4 Tg·a-1增加到2000年的67.5 Tg·a-1, 预计到2030年将达到105.3 Tg·a-1 (Zheng et al., 2002).目前, 大气N沉降对陆地生态系统结构和功能的影响已经成为全球变化研究的重要内容, 但纵观已有研究, 主要集中在地上部分, 如植物物候、生长、生理及其生物量分配对N沉降的响应规律与机制(Fleischer et al., 2013; Talhelm et al., 2013; Xia & Wan 2013), 而对生态系统地下部分的研究相对缺乏.根系作为连接生态系统地上与地下过程的重要纽带, 由根系活动所介导的根际生态过程不仅是生态学研究中的“瓶颈”, 也是生态系统结构、功能研究中最不确定的因素, 因而严重制约着生态系统与全球变化研究的理论拓展(贺金生等, 2004). ...
Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions. 1 2008
... 氮(N)是影响陆地生态系统植物生长的首要营养元素, 其变化深刻改变陆地生态系统的植物生长和净初级生产力, 进而影响全球碳(C)循环和其他生态过程(LeBauer & Treseder, 2008; Kopacek et al., 2013).近年来, 由于化石燃料的燃烧、含N化肥的大量使用等人类活动的干扰, 大气中含N化合物浓度迅速增加, 并不断向陆地和水生生态系统沉降, 从而改变了生态系统的C、N循环(Galloway et al., 2008).研究表明, 全球亚洲区域内大气N沉降已从1961年的14.4 Tg·a-1增加到2000年的67.5 Tg·a-1, 预计到2030年将达到105.3 Tg·a-1 (Zheng et al., 2002).目前, 大气N沉降对陆地生态系统结构和功能的影响已经成为全球变化研究的重要内容, 但纵观已有研究, 主要集中在地上部分, 如植物物候、生长、生理及其生物量分配对N沉降的响应规律与机制(Fleischer et al., 2013; Talhelm et al., 2013; Xia & Wan 2013), 而对生态系统地下部分的研究相对缺乏.根系作为连接生态系统地上与地下过程的重要纽带, 由根系活动所介导的根际生态过程不仅是生态学研究中的“瓶颈”, 也是生态系统结构、功能研究中最不确定的因素, 因而严重制约着生态系统与全球变化研究的理论拓展(贺金生等, 2004). ...
Chronic nitrogen deposition reduces the abundance of dominant forest understory and groundcover species. 1 2013
... 氮(N)是影响陆地生态系统植物生长的首要营养元素, 其变化深刻改变陆地生态系统的植物生长和净初级生产力, 进而影响全球碳(C)循环和其他生态过程(LeBauer & Treseder, 2008; Kopacek et al., 2013).近年来, 由于化石燃料的燃烧、含N化肥的大量使用等人类活动的干扰, 大气中含N化合物浓度迅速增加, 并不断向陆地和水生生态系统沉降, 从而改变了生态系统的C、N循环(Galloway et al., 2008).研究表明, 全球亚洲区域内大气N沉降已从1961年的14.4 Tg·a-1增加到2000年的67.5 Tg·a-1, 预计到2030年将达到105.3 Tg·a-1 (Zheng et al., 2002).目前, 大气N沉降对陆地生态系统结构和功能的影响已经成为全球变化研究的重要内容, 但纵观已有研究, 主要集中在地上部分, 如植物物候、生长、生理及其生物量分配对N沉降的响应规律与机制(Fleischer et al., 2013; Talhelm et al., 2013; Xia & Wan 2013), 而对生态系统地下部分的研究相对缺乏.根系作为连接生态系统地上与地下过程的重要纽带, 由根系活动所介导的根际生态过程不仅是生态学研究中的“瓶颈”, 也是生态系统结构、功能研究中最不确定的因素, 因而严重制约着生态系统与全球变化研究的理论拓展(贺金生等, 2004). ...
植物根系分泌物研究方法评述 2010
A test of a potential short cut in the nitrogen cycle: The role of exudation of symbiotically fixed nitrogen from the roots of a N-fixing tree and the effects of increased atmospheric CO2 and temperature. 1 1999
... 前期研究表明, 植物根系分泌物输入受多种生物和非生物因素影响, 主要包括土壤理化特性、植物种类和发育阶段、矿质营养、光温条件以及根际微生物组成和数量等(Hodge et al., 1996; Uselman et al., 1999).通过比较分析发现, 不同植物间根系分泌物C输入速率呈现出较明显的差异(表1), 这说明植物种类、自身遗传特性对根系分泌物输入有着决定性的影响(罗永清等, 2012).总体而言, 自然状况下窄叶鲜卑花灌丛根系分泌物C输入速率介于草本植物和乔木之间.这可能与灌丛植物自身的光合速率、根系形态、生理特征、生产力以及地下C分配格局有关.另外, 同一树种在幼苗阶段根系分泌物C输入速率低于成株, 这可能与植物所处生长阶段的不同生理状态以及环境条件有关, 但具体机理尚不清楚, 有待进一步深入研究分析. ...
Effects of elevated atmospheric CO2, cutting frequency, and differential day/night atmospheric warming on root growth and turnover of phalaris swards. 1 2007
... 近年来, 随着地下生态学研究的深入, 植物根系在调控土壤功能和养分循环过程中的重要作用逐渐被人们关注和认知(Björk et al., 2007; Volder et al., 2007).根系分泌物是植物在生长过程中通过根系分泌的一系列化合物, 主要为一些低分子(有机酸、糖类、酚类和各种氨基酸等)和高分子(蛋白质、黏液等)有机化合物(Dijkstra & Cheng, 2007), 通常被认为是源自植物最近一段时间的光合产物, 通过体内新陈代谢, 由根系持续地输入到地下(Kuzyakov & Cheng, 2001), 并最终成为根际微生态系统中植物根系-微生物-土壤之间物质、能量和信息交流的重要纽带.植物通过根系分泌物向根际输入易于土壤微生物直接利用的含C有机物, 有效地改变根际微生物数量和活性而深刻地影响土壤有机质分解和养分转化过程(Phillips et al., 2011; Yin et al., 2013a), 从而导致根系分泌物在调控土壤C-养分转化物质循环过程中发挥着与其含量不成比例的重要作用和功能(Cheng et al., 2005).植物根系分泌物输入及其介导下的土壤生物地球化学过程已成为地下生态学研究中的关键环节(Yin et al., 2014). ...
Soil insects alter fine root demography in peach ( 1 2005
Do warming-induced changes in quantity and stoichiometry of root exudation promote soil N transformations via stimulation of soil nitrifiers, denitrifiers and ammonifiers? 2016
The Asian nitrogen cycle case study. 1 2002
... 氮(N)是影响陆地生态系统植物生长的首要营养元素, 其变化深刻改变陆地生态系统的植物生长和净初级生产力, 进而影响全球碳(C)循环和其他生态过程(LeBauer & Treseder, 2008; Kopacek et al., 2013).近年来, 由于化石燃料的燃烧、含N化肥的大量使用等人类活动的干扰, 大气中含N化合物浓度迅速增加, 并不断向陆地和水生生态系统沉降, 从而改变了生态系统的C、N循环(Galloway et al., 2008).研究表明, 全球亚洲区域内大气N沉降已从1961年的14.4 Tg·a-1增加到2000年的67.5 Tg·a-1, 预计到2030年将达到105.3 Tg·a-1 (Zheng et al., 2002).目前, 大气N沉降对陆地生态系统结构和功能的影响已经成为全球变化研究的重要内容, 但纵观已有研究, 主要集中在地上部分, 如植物物候、生长、生理及其生物量分配对N沉降的响应规律与机制(Fleischer et al., 2013; Talhelm et al., 2013; Xia & Wan 2013), 而对生态系统地下部分的研究相对缺乏.根系作为连接生态系统地上与地下过程的重要纽带, 由根系活动所介导的根际生态过程不仅是生态学研究中的“瓶颈”, 也是生态系统结构、功能研究中最不确定的因素, 因而严重制约着生态系统与全球变化研究的理论拓展(贺金生等, 2004). ...
短期N素添加和模拟放牧对青藏高原高寒草甸生态系统呼吸的影响 1 2013
... N素富集对窄叶鲜卑花灌丛根系分泌物C输入速率的抑制作用可通过如下机制来解释: (1)导致窄叶鲜卑花灌丛根系分泌物C输入速率降低的一个重要原因可能与N富集下植物根系生长抑制有关.研究表明, 植物根系分泌物输入与根系生长及生理活性密切相关(刘子雄等, 2005).一般地, N沉降在短期内通常会促进植物地上部分生物量的积累, 抑制植物地下根系的生长(Fenn et al., 2005; Phillips & Fahey, 2007), 从而导致细根生物量和地下C分配降低(Magill et al., 2004).本研究中N素富集下窄叶鲜卑花灌丛细根生物量降低这一结果(图4)也进一步佐证了上述推测.(2) N富集所导致的窄叶鲜卑花灌丛根系对养分的微生物需求变化可能也是植物根系分泌物C输入速率降低的重要原因.根系分泌物是驱动根际微生物群落组成和活性的重要因子, 在调控土壤生物地球化学过程中具有重要的生态学作用(Drake et al., 2013).本研究中, 窄叶鲜卑花灌丛受气候条件的限制, 土壤有机质分解缓慢, 养分匮乏, 属于典型N素受限的高寒生态系统类型(宗宁等, 2013).在非施肥条件下, 由于受土壤N有效性的限制, 土壤微生物将根系分泌物输入的C作为能源, 以刺激和促进土壤有机质分解和N素转化, 从而满足植物和微生物生长所需要的养分(Jones et al., 2004; Phillips et al., 2011).相反, 持续而高强度的N素富集在一定程度上增加了窄叶鲜卑花灌丛土壤有效N水平, 使得植物能更轻易地获取所需的养分, 从而缓和了植物根系对养分的微生物驱动需求.在此情况下, 植物采取了低N收益-低C投入的养分获取策略(Phillips et al., 2011; Yin et al., 2014), 即植物在面对N素富集时通过自我生理调节降低了根系分泌物C输入.本研究中N素富集下窄叶鲜卑花土壤有效N含量(表4)与微生物PLFAs总量(图5)降低也进一步佐证了我们的推测. ...

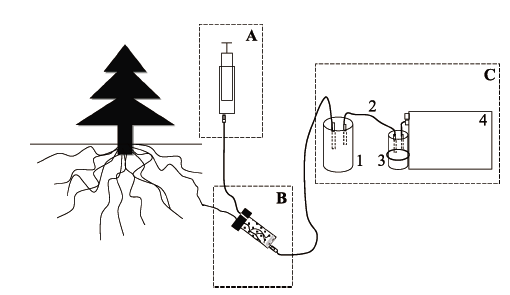 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}