Effect of stem length to stem slender ratio of current-year twigs on the leaf display efficiency in evergreen and deciduous broadleaved trees
LIJun-Hui1, PENGGuo-Quan2, YANGDong-Mei1,* 1College of Chemistry and Life Sciences, Zhejiang Normal University, Jinhua, Zhejiang 321004, Chinaand 2College of Forestry, Northwest A & F University, Yangling, Shaanxi 712100, China; 通讯作者:* 通信作者Author for correspondence (E-mail: yangdm@zjnu.cn) 收稿日期:2017-04-5 接受日期:2016-12-12 网络出版日期:2017-07-28 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:基金项目 国家自然科学基金(31000193)和浙江省自然科学基金(LQ16C030002)
关键词:茎长度;茎纤细率;展叶效率;常绿物种;落叶物种 Abstract Aims Branches and leaves are the two main structural units of tree crown composition. Among the adaptive strategies of plants, the functional traits of branches and the relationships between branch traits and leaf traits determine the capacity of trees to access light and space. In this study, our objective is to test the hypothesis that leaf display efficiency is affected by the stem length to stem slender ratio within current-year twigs.Methods The stem length to stem slender ratios of current-year twigs were used as the proxy of stem structure traits. Leaf area ratio (total leaf area per stem mass), leaf density (leaf number per stem length) and leaf/stem mass ratio (total leaf mass per stem mass) were used as the proxies of leaf display efficiency. The relationship between stem structure traits and leaf display efficiency within current-year twigs were studied for 25 evergreen and 60 deciduous broadleaved woody species in Qingliang Mountain, Zhejiang, China. The standardized major axis estimation method was used to examine the scaling relationship between stem structural traits and leaf display efficiency within current-year twigs.Important findings The proxies of leaf display efficiency, measured by leaf area ratio, leaf density or leaf/stem mass ratio, were all significantly and negative correlated with stem length to stem slender ratio within current-year twigs in both evergreen and deciduous broadleaved woody species. This suggested that leaf display efficiency decreased with stem length to stem slender ratios within current-year twigs, which may reflect the role of mechanical safety and light within twigs. The slope of the relationship between leaf display efficiency and stem long-dimension structure traits in evergreen species was not significantly different from the one in deciduous species. In contrast, the y-intercept of the relationship between leaf density and stem long-dimension structure traits was significantly larger in evergreen species than in deciduous species, i.e. the leafing intensity of evergreen species was higher than that of deciduous species. Individual leaf area and specific leaf area were smaller in evergreen species than in deciduous species, which resulted in deciduous species have a larger leaf area per stem mass and leaf mass per stem mass at a given stem length to stem slender ratio compared to evergreen species. It may reflect the conservative adaptive strategy of high consumption and slow benefit in evergreen species. Our results demonstrated that leaf display efficiency could be affected by stem length, and would change with leaf life-span (deciduous versus evergreen).
对于植物构型性状以物种平均值进行分析。在对枝条结构与展叶效率的关系分析前, 首先将各参数的物种平均值进行对数转换(以10为底), 使之符合正态分布, 然后用关系方程y = bxa来描述, 对数转换后为log(y) = log(b) + alog(x), 其中x和y表示两个性状变量, 在双对数直角坐标系中, b表示性状关系的截距, a表示斜率, 即异速生长的指数(a = 1时为等速生长, 大于或小于1时为异速生长)(Harvey & Pagel, 1991)。异速生长方程的参数估计采用标准化主轴估计(SMA)方法(Warton et al., 2006), 由软件(S)MATR Version 2.0 (Falster et al., 2006)计算完成。回归斜率的置信区间根据(Pitman, 1939)方法计算。 在异速生长分析中, 对于常绿和落叶物种组的斜率采用Warton和Weber (2002)的方法进行异质性检验, 如果检验结果没有异质性(p > 0.05), 就给出一个共同斜率。共同斜率下的截距差异(y轴向)以及沿共同主轴方向位移的差异通过t检验或方差分析检验(Wright et al., 2001; Warton et al., 2006)进行分析。 此外, 落叶和常绿物种组的各构型性状间的比较采用独立样本的t检验。所有数据分析均在SPSS 18.0下进行。
2 结果
2.1 小枝茎结构与展叶效率的关系
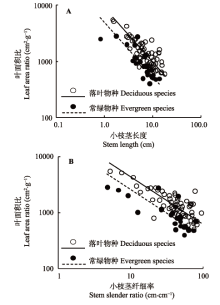
2.1.1 小枝茎结构与叶面积比(LAR)的关系 在本研究中, LAR与SL、小枝SSR在落叶和常绿物种组中均具有显著的负相关关系(图1; 表1), 其共同斜率分别为-0.961 (p = 0.773, 95%置信区间-1.097到-0.843)和-0.844 (p = 0.546, 95%置信区间-0.970到-0.736), 表明小枝上的叶面积比随小枝茎长度和茎纤细率的增加而逐渐递减。同时, 落叶物种组的叶面积比与小枝茎长度、茎纤细率相关关系共同斜率的截距显著大于常绿物种组(图1, p值均小于0.001), 表明在某一给定的茎长结构下, 落叶物种比常绿物种具有更高的叶面积比。 显示原图|下载原图ZIP|生成PPT 图1叶面积比与当年生小枝茎长度(A)、茎纤细率(B)之间的关系。 -->Fig. 1Relationship between leaf area ratio and current- year twig stem length (A) and stem slender ratio (B). -->
Table 1 表1 表1清凉峰阔叶木本植物茎结构与其他功能性状的标准化主轴估计回归关系 Table 1Summary of standardized major axis estimation regression parameters for the scaling relationships between twig stem structure and other functional trains in plant twigs of broadleaved woody species, Qingliang Mountain.
指标 (y 轴-x 轴) Index (y axis-x axis)
功能组 Functional group
样本量 Sample size
决定系数 Coefficient of determination
斜率(95%置信区间) Slope (95% confidence interval)
截距 Intercept
叶数量-小枝茎长度 Leaf number-stem length
落叶 Deciduous
60
0.192
0.510 (0.404, 0.645)
0.238
常绿 Evergreen
25
0.279
0.463 (0.323, 0.662)
0.415
叶面积比-小枝茎纤细率 Leaf area ratio-stem slender ratio
落叶 Deciduous
60
0.613
-0.867 (-1.021, -0.737)
4.525
常绿 Evergreen
25
0.620
-0.791 (-1.028, -0.608)
4.175
叶密度-小枝茎纤细率 Density of leaf number-stem slender ratio
落叶 Deciduous
60
0.617
-0.804 (-0.945, -0.684)
1.115
常绿 Evergreen
25
0.781
-0.717 (-0.876, -0.587)
1.106
叶茎质量比-小枝茎纤细率 Leaf/stem mass ratio-stem slender ratio
落叶 Deciduous
60
0.629
-0.869 (-1.019, -0.741)
2.231
常绿 Evergreen
25
0.823
-0.613 (-0.734, -0.512)
1.859
叶面积比-小枝茎长度 Leaf area ratio-stem length
落叶 Deciduous
60
0.628
-0.973 (-1.141, -0.829)
3.955
常绿 Evergreen
25
0.683
-0.939 (-1.195, -0.738)
3.679
叶密度-小枝茎长度 Density of leaf number-stem length
落叶 Deciduous
60
0.741
-0.902 (-1.030, -0.789)
0.587
常绿 Evergreen
25
0.787
-0.852 (-1.038, -0.699)
0.657
叶茎质量比-小枝茎长度 Leaf/stem mass ratio-stem length
落叶 Deciduous
60
0.614
-0.974 (-1.146, -0.828)
1.660
常绿 Evergreen
25
0.852
-0.729 (-0.859, -0.618)
1.474
总叶面积-茎干质量 Total leaf area-stem dry mass
落叶 Deciduous
60
0.437
0.815 (0.670, 0.991)
0.504
常绿 Evergreen
25
0.439
0.722 (0.526, 0.992)
0.502
All scaling relationships were significant (p < 0.05).回归关系均达到显著水平(p < 0.05)。 新窗口打开 2.1.2 小枝茎结构与叶密度的关系 无论在落叶和常绿物种组内, 小枝茎长度均与其上的叶数量呈显著的正相关关系(图2; 表1), 其共同斜率为0.495 (p = 0.653, 95%置信区间0.407到0.602), 显著小于1.0 (p < 0.000β1), 即存在显著的小于1的异速生长关系, 表明叶数量的增加速度没有小枝茎长度的增加速度快, 长的枝条相对于短枝具有低的出叶强度。此外, 落叶物种的小枝茎长度与叶数量的相关关系共同斜率的截距显著低于常绿物种组(图2, p < 0.001), 表明在某一给定的茎长度下, 常绿物种比落叶物种具有更多的叶数量。 显示原图|下载原图ZIP|生成PPT 图2小枝上的叶数量与当年生小枝茎长度之间的关系。 -->Fig. 2Relationship between leaf number per twig and current-year twig stem length. -->
小枝茎纤细率和茎长度在落叶和常绿物种组中均与叶密度呈显著的负相关关系(图3; 表1), 其共同斜率分别为-0.766 (p = 0.377, 95%置信区间-0.873到-0.677)和-0.885 (p = 0.622, 95%置信区间-0.988到-0.793), 表明随着小枝茎纤细率和长度的增加, 叶密度逐渐减小。此外, 落叶物种的小枝结构性状与叶密度相关关系共同斜率的截距显著低于常绿物种组(图3, p值均小于0.001), 表明在某一给定的小枝茎纤细率和茎长度下, 常绿物种比落叶物种具有更高的叶密度。 显示原图|下载原图ZIP|生成PPT 图3叶密度与小枝茎纤细率(A)和小枝茎长度(B)的关系。 -->Fig. 3Relationship between the density of leaf number and stem slender ratio (A) and stem length (B). -->
2.1.3 小枝茎结构与叶茎生物量比的关系 小枝的茎纤细率和茎长度在落叶和常绿物种组中均与叶茎生物量比呈显著的负相关关系(图4; 表1), 且在两组关系中, 同时表现出落叶物种关系斜率显著低于常绿物种(p值均小于0.02), 表明在小枝茎纤细率和长度增加相同的情况下, 落叶物种的小枝叶生物量分配比例比常绿物种下降得更快。 显示原图|下载原图ZIP|生成PPT 图4叶茎质量比与小枝茎纤细率(A)和小枝茎长度(B)的关系。 -->Fig. 4Relationship between the leaf/stem mass ratio and stem slender ratio (A) and stem length (B). -->
2.2 常绿和落叶物种展叶效率及各功能性状的比较
在本研究区域, 常绿物种和落叶物种在展叶效率和部分茎、叶功能性状上表现出显著的差异(表2)。落叶物种比常绿物种具有更高的单叶面积和当年生小枝上的总叶面积(p ≤ 0.001); 具有更低的叶密度和叶面积比(p < 0.05)。然而, 两个物种组在茎长度、直径、叶干质量、茎干质量及叶茎质量比和茎纤细率上无显著差异(p > 0.05)。 Table 2 表2 表2清凉峰不同生活型物种的茎、叶功能特征(平均值±标准误差) Table 2Stem and leaf functional traits of different life forms in Qingliang Moutain (mean ± SE)
性状 Trait
常绿 Evergreen
落叶 Deciduous
显著水平 Significant value, p
茎纤细率 Stem slender ratio (cm·cm-1)
41.80 ± 3.98
46.87 ± 2.80
0.318
茎直径 Stem diameter (cm)
0.18 ± 0.01
0.18 ± 0.01
0.887
茎长度 Stem length (cm)
6.77 ± 0.62
8.01 ± 0.50
0.158
茎干质量 Stem mass (mg)
112.03 ± 11.72
132.50 ± 13.22
0.352
总叶干质量 Total leaf mass (mg)
853.32 ± 74.69
840.87 ± 62.55
0.909
叶密度指数 Leaf density index (No.·cm-1)
1.20 ± 0.20
0.76 ± 0.06
0.007
叶面积比 Leaf area ratio (cm2·g-1)
1β081.71 ± 146.01
1β573.41 ± 130.10
0.030
单叶面积 Individual leaf area (cm2)
16.21 ± 1.97
34.66 ± 2.34
<0.001
总叶面积 Total leaf area (cm2)
95.19 ± 10.34
163.68 ± 11.69
0.001
比叶面积 Specific leaf area (cm2·g-1)
114.31 ± 6.89
200.76 ± 4.42
<0.001
叶茎质量比 Leaf/stem mass ratio (mg·mg-1)
9.14 ± 0.99
7.96 ± 0.66
0.335
P values are the results of student’s t-test in different traits between evergreen and deciduous species.p值为t检验结果。 新窗口打开
BellAD(1984). Dynamic morphology: A contribution to plant population ecology. In: Dirzo R ed. Perspectives on Plant Population Ecology . Sinaner, Sunderland, USA. [本文引用: 1]
[2]
BicknellSH (1982). Development of canopy stratification during early succession in northern hardwoods. Forest Eco- logy & Management, 4, 41-51. [本文引用: 1]
[3]
BoojhR, RamakrishnanPS (1982). Growth strategy of trees related to successional status. I. Architecture and extension growth. Forest Ecology & Management, 4, 359-374. [本文引用: 1]
[4]
BorchertR, SladeNA (1981). Bifurcation ratios and the adaptive geometry of trees. Botanical Gazette, 142, 394-401. [本文引用: 1]
[5]
CaesarJC, MacdonaldAD (1984). Shoot development in Betula papyrifera. IV. Comparisons between growth characteristics and expression of vegetative long and short shoots . Canadian Journal of Botany, 62, 446-453. [本文引用: 1]
[6]
Cavender-BaresJ, CortesP, RambalS, JoffreR, MilesB, RocheteauA (2005). Summer and winter sensitivity of leaves and xylem to minimum freezing temperatures: A comparison of co-occurring Mediterranean oaks that differ in leaf lifespan. New Phytologist, 168, 597-612. [本文引用: 2]
[7]
Cavender-BaresJ, HolbrookNM (2001). Hydraulic properties and freezing-induced cavitation in sympatric evergreen and deciduous oaks with contrasting habitats. Plant, Cell & Environment, 24, 1243-1256. [本文引用: 2]
[8]
ChenB, SongYC, DaLJ (2002). Woody plant architecture and its research in plant ecology. Chinese Journal of Ecology, 21, 52-56. (in Chinese with English abstract)[ 陈波, 宋永昌, 达良俊 (2002). 木本植物的构型及其在植物生态学研究的进展 . 生态学杂志, 21, 52-56.] [本文引用: 1]
[9]
ChengLY, CaoT, ZhangHW, YuJ (2016). Studies on flora of bryophytes in Qingliangfeng National Reserve, Zhejiang Province. Acta Botanica Boreali-Occidentalia Sinica, 23, 398-403. (in Chinese with English abstract)[ 程丽媛, 曹同, 张宏伟, 于晶 (2016). 浙江省清凉峰自然保护区苔藓植物区系成分研究 . 西北植物学报, 36, 398-403.] [本文引用: 2]
[10]
ColeyPD (1988). Effects of plant growth rate and leaf lifetime on the amount and type of anti-herbivore defense. Oecologia, 74, 531-536. [本文引用: 1]
[11]
CornelissenJHC (1999). A triangular relationship between leaf size and seed size among woody species: Allometry, ontogeny, ecology and taxonomy. Oecologia, 118, 248-255. [本文引用: 1]
[12]
CornerEJH (1949). The durian theory or the origin of the modern tree. Annals of Botany, 13, 367-414. [本文引用: 1]
[13]
DayME, GreenwoodMS, DiazsalaC (2002). Age- and size- related trends in woody plant shoot development: Regulatory pathways and evidence for genetic control. Tree Physiology, 22, 507-513. [本文引用: 1]
[14]
DeanTJ, LongJN (1986). Validity of constant-stress and elastic-instability principles of stem formation in Pinus contorta and Trifolium pratense. Annals of Botany, 58, 833-840. [本文引用: 2]
[15]
DiemerM (1998). Life span and dynamics of leaves of herbaceous perennials in high-elevation environments: “News from the elephant’s leg”. Functional Ecology, 12, 413-425. [本文引用: 1]
[16]
DongL, JiaGX, SuXH (2003). Change of the leaf tissue structure of evergreen broad-leaf plants during overwintering. Acta Horticulture Sinica, 30, 59-64. (in Chinese with English abstract)[ 董丽, 贾桂霞, 苏雪痕 (2003). 常绿阔叶植物越冬期间叶片组织结构的适应性变化 . 园艺学报, 30, 59-64.] [本文引用: 1]
HornHS (1971). The adaptive geometry of trees. Botanical Gazette, 142, 394-401. [本文引用: 1]
[24]
KikuzawaK (1991). A cost-benefit analysis of leaf habit and leaf longevity of trees and their geographical pattern. The American Naturalist, 138, 1250-1263. [本文引用: 1]
[25]
KikuzawaK (1995). Leaf phenology as an optimal strategy for carbon gain in plants. Canadian Journal Botany, 73, 158-163. [本文引用: 1]
[26]
KingD (1981). Tree dimensions: Maximizing the rate of height growth in dense stands. Oecologia, 51, 351-356. [本文引用: 1]
[27]
KingD, LoucksOL (1978). The theory of tree bole and branch form. Radiation and Environmental Biophysics, 15, 141-165. [本文引用: 1]
[28]
KingDA (1986). Tree form, height growth, and susceptibility to wind damage in Acer saccharum. Ecology, 67, 980-990. [本文引用: 1]
[29]
KleimanD, AarssenLW (2007). The leaf size/number trade-off in trees. Journal of Ecology, 95, 376-382. [本文引用: 2]
[30]
KoikeF (1989). Foliage-crown development and interaction in Quercus gilva and Q. acuta. Journal of Ecology, 77, 92-111. [本文引用: 3]
[31]
LiG, YangD, SunS (2008). Allometric relationships between lamina area, lamina mass and petiole mass of 93 temperate woody species vary with leaf habit, leaf form and altitude. Functional Ecology, 22, 557-564. [本文引用: 1]
[32]
LiYX, LiuYC (1996). The neighbourhood modular interference effects of seedlings of Gordonia acuminata. Journal of Southwest China Normal University (Natural Science), 21, 271-275. (in Chinese with English abstract)[ 黎云祥, 刘玉成 (1996). 四川大头茶苗木构件水平的邻体干扰效应 . 西南师范大学学报自然科学版, 21, 271-275.] [本文引用: 3]
[33]
LiuYF (2015). The Study on Anatomical Structures Character and Its Environmental Adaptation of Leaves from Broad- Leaf Woody Plants in Gongga Mountain. Master degree dissertation, Southwest University, Chongqing. (in Chinese with English abstract)[ 刘艳芳 (2015). 贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究 . 硕士学位论文, 西南大学, 重庆.] [本文引用: 1]
[34]
LongJN, SmithFW, ScottDRM (1981). The role of Douglas- fir stem sapwood and heartwood in the mechanical and physiological support of crowns and development of stem form. Canadian Journal of Forest Research, 11, 459-464. [本文引用: 1]
[35]
MaruyamaK (1983). Shoot characteristics as a function of the bud length on Japanese beech trees. Journal of the Japanese Forestry Society, 65, 43-51. [本文引用: 1]
[36]
McmahonT (1973). Size and shape in biology. Science, 179, 1201-1204. [本文引用: 1]
[37]
NiklasKJ, KerchnerV (1984). Mechanical and photosynthetic constraints on the evolution of plant shape. Paleobiology, 10, 79-101. [本文引用: 1]
[38]
NiklasKJ (1988). The role of phyllotactic pattern as a “developmental constraint” on the interception of light by leaf surfaces. Evolution, 42, 1-16. [本文引用: 1]
[39]
PanSA, PengGQ, YangDM (2015). Biomass allocation strategies within a leaf: Implication for leaf size optimization. Chinese Journal of Plant Ecology, 39, 971-979. (in Chinese with English abstract)[ 潘少安, 彭国全, 杨冬梅 (2015). 从叶内生物量分配策略的角度理解叶大小的优化 . 植物生态学报, 39, 971-979.] [本文引用: 1]
[40]
PickupM, WestobyM, BasdenA (2005). Dry mass costs of deploying leaf area in relation to leaf size. Functional Ecology, 19, 88-97. [本文引用: 1]
[41]
PitmanE (1939). A note on normal correlation. Biometrika, 31, 9-12. [本文引用: 4]
[42]
ReichP, WaltersM, EllsworthD (1992). Leaf life-span in relation to leaf, plant, and stand characteristics among diverse ecosystems. Ecological Monographs, 62, 365-392. [本文引用: 1]
[43]
ReichPB, BowmanWD (1999). Generality of leaf trait relationships: A test across six biomes. Ecology, 80, 1955-1969. [本文引用: 1]
[44]
ReichPB, WaltersMB, EllsworthDS, VoseJM, VolinJC, GreshamC, BowmanWD (1998). Relationships of leaf dark respiration to leaf nitrogen, specific leaf area and leaf life-span: A test across biomes and functional groups. Oecologia, 114, 471-482. [本文引用: 1]
[45]
RyserP, UrbasP (2000). Ecological significance of leaf life span among Central European grass species. Oikos, 91, 41-50. [本文引用: 1]
[46]
SchulzeE, KelliherFM, KornerC, LloydJ, LeuningR (1994). Relationships among maximum stomatal conductance, ecosystem surface conductance, carbon assimilation rate, and plant nitrogen nutrition: A global ecology scaling exercise. Ecology, Evolution, and Systematics, 25, 629-662. [本文引用: 1]
[47]
SunS, JinD, ShiP (2006). The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient: An invariant allometric scaling relationship. Annals of Botany, 97, 97-107. [本文引用: 2]
[48]
TakenakaA (1997). Structural variation in current-year shoots of broad-leaved evergreen tree saplings under forest canopies in warm temperate Japan. Tree Physiology, 17, 205-210. [本文引用: 1]
[49]
ThomasSC, WinnerWE (2002). Photosynthetic differences between saplings and adult trees: An integration of field results by meta-analysis. Tree Physiology, 22, 117-127.
[50]
WartonDI, WeberNC (2002). Common slope tests for bivariate errors-in-variables models. Biometrical Journal, 44, 161-174. [本文引用: 2]
WengDM, ZhangL, ChenXD, ShenGC, ZhangHW, ZhangFG, YuMJ (2009). Species diversity of Fagus hayatae community in Qingliangfeng National Nature Reserve. Journal of Zhejiang Forestry Science and Technology, 29(4), 1-6.[翁东明, 张磊, 陈晓栋, 沈国春, 张宏伟, 张方钢, 于明坚 (2009). 清凉峰自然保护区台湾水青冈群落物种多样性研究 . 浙江林业科技, 29(4), 1-6.] [本文引用: 4]
[53]
WestobyM, FalsterDS, MolesAT, VeskPA, WrightIJ (2002). Plant ecological strategies: Some leading dimensions of variation between species. Annual Review of Ecology and Systematics, 33, 125-159. [本文引用: 1]
[54]
WrightIJ, AckerlyDD, BongersF, HarmsKE, Ibarra- ManriquezG, Martinez-RamosM, MazerSJ, Muller- LandauHC, PazH, PitmanNCA, PoorterL, SilmanMR, VriesendorpCF, WebbCO, WestobyM, WrightSJ (2007). Relationships among ecologically important dimensions of plant trait variation in seven neotropical forests. Annals of Botany, 99, 1003-1015. [本文引用: 1]
[55]
WrightIJ, ReichPB, WestobyM (2001) Strategy shifts in leaf physiology, structure and nutrient content between species of high- and low-rainfall and high- and low-nutrient habitats. Functional Ecology, 15, 423-434. [本文引用: 3]
XiangSA, LiuYL, FangF, WuN, SunSC (2009). Stem architectural effect on leaf size, leaf number, and leaf mass fraction in plant twigs of woody species. International Journal of Plant Sciences, 170, 999-1008. [本文引用: 2]
[58]
YagiT (2000). Morphology and biomass allocation of current-year shoots of ten tall tree species in cool temperate Japan. Journal of Plant Research, 113, 171-183. [本文引用: 1]
[59]
YagiT (2004). Within-tree variations in shoot differentiation patterns of 10 tall tree species in a Japanese cool-temperate forest. Canadian Journal of Botany, 82, 228-243. [本文引用: 9]
[60]
YagiT, KikuzawaK (1999). Patterns in size-related variations in current-year shoot structure in eight deciduous tree species. Journal of Plant Research, 112, 343-352. [本文引用: 1]
[61]
YangDM, LiGY, SunSC (2008). The generality of leaf size versus number trade-off in temperate woody species. Annals of Botany, 102, 623-629. [本文引用: 2]
[62]
YangDM, LiGY, SunSC (2009). The effects of leaf size, leaf habit, and leaf form on leaf/stem relationships in plant twigs of temperate woody species. Journal of Vegetation Science, 20, 359-366. [本文引用: 2]
[63]
YangDM, ZhanF, ZhangHW (2012). Trade-off between leaf size and number in current-year twigs of deciduous broad-leaved woody species at different altidudes on Qingliang Mountain, southeastern China. Chinese Journal of Plant Ecology, 36, 281-291. (in Chinese with English abstract)[ 杨冬梅, 占峰, 张宏伟 (2012). 清凉峰不同海拔木本植物小枝内叶大小-数量权衡关系 . 植物生态学报, 36, 281-291.] [本文引用: 3]
[64]
ZhangL, LuoTX (2004). Advances in ecological studies on leaf lifespan and associated leaf traits. Acta Phytoecologica Sinica, 28, 844-852. (in Chinese with English abstract)[ 张林, 罗天祥 (2004). 植物叶寿命及其相关叶性状的生态学研究进展 . 植物生态学报, 28, 844-852.] [本文引用: 1]
[65]
ZhangYC, DuXJ, ZhangQY, GaoXM, SuZX (2005). Functions of branches of the clonal tree Symplocos laurina. Acta Phytoecologica Sinica, 29, 799-806. (in Chinese with English abstract)[ 张运春, 杜晓军, 张桥英, 高贤明, 苏智先 (2005). 克隆乔木黄牛奶树枝条的功能特征 . 植物生态学报, 29, 799-806.]
Shoot development in Betula papyrifera. IV. Comparisons between growth characteristics and expression of vegetative long and short shoots 1 1984
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978; Niklas & Kerchner, 1984).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971; Yagi & Kikuzawa, 1999), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989; Yagi & Kikuzawa, 1999).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999).尽管短枝上的总叶面积比长枝小(Gregory, 1980; Yagi & Kikuzawa, 1999), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984; Yagi & Kikuzawa, 1999).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983; Koike, 1989; Takenaka, 1997; Yagi & Kikuzawa, 1999)和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al., 2002; Pickup et al., 2005; Wright et al., 2007; Li et al., 2008; Xiang et al., 2009; Yang et al., 2009; 杨冬梅等, 2012), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M)主要由枝条长度(L)和枝条所承受的力(F)共同决定, 即M = L × F.对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F)又是叶面积(A)和风压(W)的乘积, 即F = A × W (Dean & Long, 1986).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
Summer and winter sensitivity of leaves and xylem to minimum freezing temperatures: A comparison of co-occurring Mediterranean oaks that differ in leaf lifespan. 2 2005
... 同时, 对于生活在同一群落里的常绿和落叶物种而言, 由于展叶物候和生物学习性的不同, 它们的功能特征通常也会有很大的差异.例如, 相比于落叶物种, 常绿物种通常具有长的叶寿命、低的比叶面积(SLA)、低的光合速率和高的养分含量(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; Li et al., 2008); 另外, 由于常绿物种为了确保在冬季低温等不利环境下叶片不凋落, 需要水分运输系统保持畅通, 确保水分运输安全, 其木质部需要比落叶物种具有更强的抵抗冰冻引起栓塞的能力(Cavender-Bares & Holbrook, 2001; Cavender-Bares et al., 2005), 致使其投入在支撑结构上的成本增加, 表现出在相同的茎投入时, 常绿物种通常比落叶物种具有更小的总叶面积和单叶面积(Sun et al., 2006; Yang et al., 2009).这种不同的成本-收益关系将有可能导致常绿和落叶物种采取不同的枝条伸展策略以改变小枝上的叶排布方式, 使两类物种都能够捕获足够的光能以保持相对稳定的碳收益.因此, 两类物种可能会具有不同的茎结构与展叶效率之间的变化关系. ...
... 常绿与落叶是植物通过长期进化适应形成的两种不同的生活史对策, 它们在叶寿命和叶片结构(如比叶面积)(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; Li et al., 2008)、茎木质部抗栓塞能力(Cavender-Bares & Holbrook, 2001; Cavender- Bares et al., 2005)等许多方面都显著不同.在本研究中, 虽然当年生小枝茎结构性状与表征展叶效率的性状之间的负相关关系斜率在常绿和落叶物种间无显著差异, 但截距差异显著.在3个表征展叶效率的性状中, 尽管常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 在某一给定的茎长或茎纤细率下, 常绿物种比落叶物种具有更多的叶数量, 即具有较高的出叶强度, 与以前相关研究发现的叶大小与叶数量之间是相互权衡的结果一致(Corner, 1949; Kleiman & Aarssen, 2007; Yang et al., 2008).但由于常绿物种的单叶面积16.21 cm2显著小于落叶物种的单叶面积34.66 cm2 (p < 0.001, 表2), 导致其叶面积比(单位茎质量的叶面积)和叶茎生物量比(单位茎质量的叶质量)与小枝茎结构性状之间的关系截距反而显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低.对数据结果进一步分析表明, 常绿物种更低展叶效率的主要原因是其比叶面积显著小于落叶物种 (p < 0.001, 表2), 因为我们的结果显示, 在不考虑小枝茎长结构影响的情况下, 茎叶生物量比在常绿和落叶物种中没有显著差异(p = 0.335, 表2), 表明总生物量在叶和茎之间的分配比例在常绿和落叶物种中是没有显著差异的, 可是落叶物种的单叶面积(p < 0.001, 表2)和总叶面积(p = 0.001, 表2)都要显著大于常绿物种, 并且常绿物种茎干质量与总叶面积相关关系共同斜率下的截距显著小于落叶物种(p ˂ 0.05, 表1), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积.许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; 潘少安等, 2015).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
Hydraulic properties and freezing-induced cavitation in sympatric evergreen and deciduous oaks with contrasting habitats. 2 2001
... 同时, 对于生活在同一群落里的常绿和落叶物种而言, 由于展叶物候和生物学习性的不同, 它们的功能特征通常也会有很大的差异.例如, 相比于落叶物种, 常绿物种通常具有长的叶寿命、低的比叶面积(SLA)、低的光合速率和高的养分含量(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; Li et al., 2008); 另外, 由于常绿物种为了确保在冬季低温等不利环境下叶片不凋落, 需要水分运输系统保持畅通, 确保水分运输安全, 其木质部需要比落叶物种具有更强的抵抗冰冻引起栓塞的能力(Cavender-Bares & Holbrook, 2001; Cavender-Bares et al., 2005), 致使其投入在支撑结构上的成本增加, 表现出在相同的茎投入时, 常绿物种通常比落叶物种具有更小的总叶面积和单叶面积(Sun et al., 2006; Yang et al., 2009).这种不同的成本-收益关系将有可能导致常绿和落叶物种采取不同的枝条伸展策略以改变小枝上的叶排布方式, 使两类物种都能够捕获足够的光能以保持相对稳定的碳收益.因此, 两类物种可能会具有不同的茎结构与展叶效率之间的变化关系. ...
... 常绿与落叶是植物通过长期进化适应形成的两种不同的生活史对策, 它们在叶寿命和叶片结构(如比叶面积)(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; Li et al., 2008)、茎木质部抗栓塞能力(Cavender-Bares & Holbrook, 2001; Cavender- Bares et al., 2005)等许多方面都显著不同.在本研究中, 虽然当年生小枝茎结构性状与表征展叶效率的性状之间的负相关关系斜率在常绿和落叶物种间无显著差异, 但截距差异显著.在3个表征展叶效率的性状中, 尽管常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 在某一给定的茎长或茎纤细率下, 常绿物种比落叶物种具有更多的叶数量, 即具有较高的出叶强度, 与以前相关研究发现的叶大小与叶数量之间是相互权衡的结果一致(Corner, 1949; Kleiman & Aarssen, 2007; Yang et al., 2008).但由于常绿物种的单叶面积16.21 cm2显著小于落叶物种的单叶面积34.66 cm2 (p < 0.001, 表2), 导致其叶面积比(单位茎质量的叶面积)和叶茎生物量比(单位茎质量的叶质量)与小枝茎结构性状之间的关系截距反而显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低.对数据结果进一步分析表明, 常绿物种更低展叶效率的主要原因是其比叶面积显著小于落叶物种 (p < 0.001, 表2), 因为我们的结果显示, 在不考虑小枝茎长结构影响的情况下, 茎叶生物量比在常绿和落叶物种中没有显著差异(p = 0.335, 表2), 表明总生物量在叶和茎之间的分配比例在常绿和落叶物种中是没有显著差异的, 可是落叶物种的单叶面积(p < 0.001, 表2)和总叶面积(p = 0.001, 表2)都要显著大于常绿物种, 并且常绿物种茎干质量与总叶面积相关关系共同斜率下的截距显著小于落叶物种(p ˂ 0.05, 表1), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积.许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; 潘少安等, 2015).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
Tree form, height growth, and susceptibility to wind damage in 1 1986
... 常绿与落叶是植物通过长期进化适应形成的两种不同的生活史对策, 它们在叶寿命和叶片结构(如比叶面积)(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; Li et al., 2008)、茎木质部抗栓塞能力(Cavender-Bares & Holbrook, 2001; Cavender- Bares et al., 2005)等许多方面都显著不同.在本研究中, 虽然当年生小枝茎结构性状与表征展叶效率的性状之间的负相关关系斜率在常绿和落叶物种间无显著差异, 但截距差异显著.在3个表征展叶效率的性状中, 尽管常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 在某一给定的茎长或茎纤细率下, 常绿物种比落叶物种具有更多的叶数量, 即具有较高的出叶强度, 与以前相关研究发现的叶大小与叶数量之间是相互权衡的结果一致(Corner, 1949; Kleiman & Aarssen, 2007; Yang et al., 2008).但由于常绿物种的单叶面积16.21 cm2显著小于落叶物种的单叶面积34.66 cm2 (p < 0.001, 表2), 导致其叶面积比(单位茎质量的叶面积)和叶茎生物量比(单位茎质量的叶质量)与小枝茎结构性状之间的关系截距反而显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低.对数据结果进一步分析表明, 常绿物种更低展叶效率的主要原因是其比叶面积显著小于落叶物种 (p < 0.001, 表2), 因为我们的结果显示, 在不考虑小枝茎长结构影响的情况下, 茎叶生物量比在常绿和落叶物种中没有显著差异(p = 0.335, 表2), 表明总生物量在叶和茎之间的分配比例在常绿和落叶物种中是没有显著差异的, 可是落叶物种的单叶面积(p < 0.001, 表2)和总叶面积(p = 0.001, 表2)都要显著大于常绿物种, 并且常绿物种茎干质量与总叶面积相关关系共同斜率下的截距显著小于落叶物种(p ˂ 0.05, 表1), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积.许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; 潘少安等, 2015).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
The leaf size/number trade-off in trees. 2 2007
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978; Niklas & Kerchner, 1984).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971; Yagi & Kikuzawa, 1999), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989; Yagi & Kikuzawa, 1999).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999).尽管短枝上的总叶面积比长枝小(Gregory, 1980; Yagi & Kikuzawa, 1999), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984; Yagi & Kikuzawa, 1999).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983; Koike, 1989; Takenaka, 1997; Yagi & Kikuzawa, 1999)和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al., 2002; Pickup et al., 2005; Wright et al., 2007; Li et al., 2008; Xiang et al., 2009; Yang et al., 2009; 杨冬梅等, 2012), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M)主要由枝条长度(L)和枝条所承受的力(F)共同决定, 即M = L × F.对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F)又是叶面积(A)和风压(W)的乘积, 即F = A × W (Dean & Long, 1986).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... ; Koike, 1989; Takenaka, 1997; Yagi & Kikuzawa, 1999)和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al., 2002; Pickup et al., 2005; Wright et al., 2007; Li et al., 2008; Xiang et al., 2009; Yang et al., 2009; 杨冬梅等, 2012), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M)主要由枝条长度(L)和枝条所承受的力(F)共同决定, 即M = L × F.对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F)又是叶面积(A)和风压(W)的乘积, 即F = A × W (Dean & Long, 1986).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
Foliage-crown development and interaction in 3 1989
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978; Niklas & Kerchner, 1984).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971; Yagi & Kikuzawa, 1999), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989; Yagi & Kikuzawa, 1999).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999).尽管短枝上的总叶面积比长枝小(Gregory, 1980; Yagi & Kikuzawa, 1999), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984; Yagi & Kikuzawa, 1999).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983; Koike, 1989; Takenaka, 1997; Yagi & Kikuzawa, 1999)和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al., 2002; Pickup et al., 2005; Wright et al., 2007; Li et al., 2008; Xiang et al., 2009; Yang et al., 2009; 杨冬梅等, 2012), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M)主要由枝条长度(L)和枝条所承受的力(F)共同决定, 即M = L × F.对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F)又是叶面积(A)和风压(W)的乘积, 即F = A × W (Dean & Long, 1986).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... 同时, 对于生活在同一群落里的常绿和落叶物种而言, 由于展叶物候和生物学习性的不同, 它们的功能特征通常也会有很大的差异.例如, 相比于落叶物种, 常绿物种通常具有长的叶寿命、低的比叶面积(SLA)、低的光合速率和高的养分含量(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; Li et al., 2008); 另外, 由于常绿物种为了确保在冬季低温等不利环境下叶片不凋落, 需要水分运输系统保持畅通, 确保水分运输安全, 其木质部需要比落叶物种具有更强的抵抗冰冻引起栓塞的能力(Cavender-Bares & Holbrook, 2001; Cavender-Bares et al., 2005), 致使其投入在支撑结构上的成本增加, 表现出在相同的茎投入时, 常绿物种通常比落叶物种具有更小的总叶面积和单叶面积(Sun et al., 2006; Yang et al., 2009).这种不同的成本-收益关系将有可能导致常绿和落叶物种采取不同的枝条伸展策略以改变小枝上的叶排布方式, 使两类物种都能够捕获足够的光能以保持相对稳定的碳收益.因此, 两类物种可能会具有不同的茎结构与展叶效率之间的变化关系. ...
... 常绿与落叶是植物通过长期进化适应形成的两种不同的生活史对策, 它们在叶寿命和叶片结构(如比叶面积)(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; Li et al., 2008)、茎木质部抗栓塞能力(Cavender-Bares & Holbrook, 2001; Cavender- Bares et al., 2005)等许多方面都显著不同.在本研究中, 虽然当年生小枝茎结构性状与表征展叶效率的性状之间的负相关关系斜率在常绿和落叶物种间无显著差异, 但截距差异显著.在3个表征展叶效率的性状中, 尽管常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 在某一给定的茎长或茎纤细率下, 常绿物种比落叶物种具有更多的叶数量, 即具有较高的出叶强度, 与以前相关研究发现的叶大小与叶数量之间是相互权衡的结果一致(Corner, 1949; Kleiman & Aarssen, 2007; Yang et al., 2008).但由于常绿物种的单叶面积16.21 cm2显著小于落叶物种的单叶面积34.66 cm2 (p < 0.001, 表2), 导致其叶面积比(单位茎质量的叶面积)和叶茎生物量比(单位茎质量的叶质量)与小枝茎结构性状之间的关系截距反而显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低.对数据结果进一步分析表明, 常绿物种更低展叶效率的主要原因是其比叶面积显著小于落叶物种 (p < 0.001, 表2), 因为我们的结果显示, 在不考虑小枝茎长结构影响的情况下, 茎叶生物量比在常绿和落叶物种中没有显著差异(p = 0.335, 表2), 表明总生物量在叶和茎之间的分配比例在常绿和落叶物种中是没有显著差异的, 可是落叶物种的单叶面积(p < 0.001, 表2)和总叶面积(p = 0.001, 表2)都要显著大于常绿物种, 并且常绿物种茎干质量与总叶面积相关关系共同斜率下的截距显著小于落叶物种(p ˂ 0.05, 表1), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积.许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; 潘少安等, 2015).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
Allometric relationships between lamina area, lamina mass and petiole mass of 93 temperate woody species vary with leaf habit, leaf form and altitude. 1 2008
Relationships among maximum stomatal conductance, ecosystem surface conductance, carbon assimilation rate, and plant nitrogen nutrition: A global ecology scaling exercise. 1 1994
... 同时, 对于生活在同一群落里的常绿和落叶物种而言, 由于展叶物候和生物学习性的不同, 它们的功能特征通常也会有很大的差异.例如, 相比于落叶物种, 常绿物种通常具有长的叶寿命、低的比叶面积(SLA)、低的光合速率和高的养分含量(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; Li et al., 2008); 另外, 由于常绿物种为了确保在冬季低温等不利环境下叶片不凋落, 需要水分运输系统保持畅通, 确保水分运输安全, 其木质部需要比落叶物种具有更强的抵抗冰冻引起栓塞的能力(Cavender-Bares & Holbrook, 2001; Cavender-Bares et al., 2005), 致使其投入在支撑结构上的成本增加, 表现出在相同的茎投入时, 常绿物种通常比落叶物种具有更小的总叶面积和单叶面积(Sun et al., 2006; Yang et al., 2009).这种不同的成本-收益关系将有可能导致常绿和落叶物种采取不同的枝条伸展策略以改变小枝上的叶排布方式, 使两类物种都能够捕获足够的光能以保持相对稳定的碳收益.因此, 两类物种可能会具有不同的茎结构与展叶效率之间的变化关系. ...
The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient: An invariant allometric scaling relationship. 2 2006
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978; Niklas & Kerchner, 1984).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971; Yagi & Kikuzawa, 1999), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989; Yagi & Kikuzawa, 1999).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999).尽管短枝上的总叶面积比长枝小(Gregory, 1980; Yagi & Kikuzawa, 1999), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984; Yagi & Kikuzawa, 1999).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983; Koike, 1989; Takenaka, 1997; Yagi & Kikuzawa, 1999)和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al., 2002; Pickup et al., 2005; Wright et al., 2007; Li et al., 2008; Xiang et al., 2009; Yang et al., 2009; 杨冬梅等, 2012), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M)主要由枝条长度(L)和枝条所承受的力(F)共同决定, 即M = L × F.对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F)又是叶面积(A)和风压(W)的乘积, 即F = A × W (Dean & Long, 1986).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978; Niklas & Kerchner, 1984).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971; Yagi & Kikuzawa, 1999), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989; Yagi & Kikuzawa, 1999).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999).尽管短枝上的总叶面积比长枝小(Gregory, 1980; Yagi & Kikuzawa, 1999), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984; Yagi & Kikuzawa, 1999).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983; Koike, 1989; Takenaka, 1997; Yagi & Kikuzawa, 1999)和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al., 2002; Pickup et al., 2005; Wright et al., 2007; Li et al., 2008; Xiang et al., 2009; Yang et al., 2009; 杨冬梅等, 2012), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M)主要由枝条长度(L)和枝条所承受的力(F)共同决定, 即M = L × F.对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F)又是叶面积(A)和风压(W)的乘积, 即F = A × W (Dean & Long, 1986).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... 同时, 对于生活在同一群落里的常绿和落叶物种而言, 由于展叶物候和生物学习性的不同, 它们的功能特征通常也会有很大的差异.例如, 相比于落叶物种, 常绿物种通常具有长的叶寿命、低的比叶面积(SLA)、低的光合速率和高的养分含量(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; Li et al., 2008); 另外, 由于常绿物种为了确保在冬季低温等不利环境下叶片不凋落, 需要水分运输系统保持畅通, 确保水分运输安全, 其木质部需要比落叶物种具有更强的抵抗冰冻引起栓塞的能力(Cavender-Bares & Holbrook, 2001; Cavender-Bares et al., 2005), 致使其投入在支撑结构上的成本增加, 表现出在相同的茎投入时, 常绿物种通常比落叶物种具有更小的总叶面积和单叶面积(Sun et al., 2006; Yang et al., 2009).这种不同的成本-收益关系将有可能导致常绿和落叶物种采取不同的枝条伸展策略以改变小枝上的叶排布方式, 使两类物种都能够捕获足够的光能以保持相对稳定的碳收益.因此, 两类物种可能会具有不同的茎结构与展叶效率之间的变化关系. ...
... 常绿与落叶是植物通过长期进化适应形成的两种不同的生活史对策, 它们在叶寿命和叶片结构(如比叶面积)(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; Li et al., 2008)、茎木质部抗栓塞能力(Cavender-Bares & Holbrook, 2001; Cavender- Bares et al., 2005)等许多方面都显著不同.在本研究中, 虽然当年生小枝茎结构性状与表征展叶效率的性状之间的负相关关系斜率在常绿和落叶物种间无显著差异, 但截距差异显著.在3个表征展叶效率的性状中, 尽管常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 在某一给定的茎长或茎纤细率下, 常绿物种比落叶物种具有更多的叶数量, 即具有较高的出叶强度, 与以前相关研究发现的叶大小与叶数量之间是相互权衡的结果一致(Corner, 1949; Kleiman & Aarssen, 2007; Yang et al., 2008).但由于常绿物种的单叶面积16.21 cm2显著小于落叶物种的单叶面积34.66 cm2 (p < 0.001, 表2), 导致其叶面积比(单位茎质量的叶面积)和叶茎生物量比(单位茎质量的叶质量)与小枝茎结构性状之间的关系截距反而显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低.对数据结果进一步分析表明, 常绿物种更低展叶效率的主要原因是其比叶面积显著小于落叶物种 (p < 0.001, 表2), 因为我们的结果显示, 在不考虑小枝茎长结构影响的情况下, 茎叶生物量比在常绿和落叶物种中没有显著差异(p = 0.335, 表2), 表明总生物量在叶和茎之间的分配比例在常绿和落叶物种中是没有显著差异的, 可是落叶物种的单叶面积(p < 0.001, 表2)和总叶面积(p = 0.001, 表2)都要显著大于常绿物种, 并且常绿物种茎干质量与总叶面积相关关系共同斜率下的截距显著小于落叶物种(p ˂ 0.05, 表1), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积.许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; 潘少安等, 2015).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
... ; Westoby et al., 2002; Wright et al., 2004; 潘少安等, 2015).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
Plant ecological strategies: Some leading dimensions of variation between species. 1 2002
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978; Niklas & Kerchner, 1984).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971; Yagi & Kikuzawa, 1999), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989; Yagi & Kikuzawa, 1999).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999).尽管短枝上的总叶面积比长枝小(Gregory, 1980; Yagi & Kikuzawa, 1999), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984; Yagi & Kikuzawa, 1999).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983; Koike, 1989; Takenaka, 1997; Yagi & Kikuzawa, 1999)和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al., 2002; Pickup et al., 2005; Wright et al., 2007; Li et al., 2008; Xiang et al., 2009; Yang et al., 2009; 杨冬梅等, 2012), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M)主要由枝条长度(L)和枝条所承受的力(F)共同决定, 即M = L × F.对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F)又是叶面积(A)和风压(W)的乘积, 即F = A × W (Dean & Long, 1986).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
Relationships among ecologically important dimensions of plant trait variation in seven neotropical forests. 1 2007
... 在异速生长分析中, 对于常绿和落叶物种组的斜率采用Warton和Weber (2002)的方法进行异质性检验, 如果检验结果没有异质性(p > 0.05), 就给出一个共同斜率.共同斜率下的截距差异(y轴向)以及沿共同主轴方向位移的差异通过t检验或方差分析检验(Wright et al., 2001; Warton et al., 2006)进行分析. ...
Strategy shifts in leaf physiology, structure and nutrient content between species of high- and low-rainfall and high- and low-nutrient habitats. 3 2001
... 同时, 对于生活在同一群落里的常绿和落叶物种而言, 由于展叶物候和生物学习性的不同, 它们的功能特征通常也会有很大的差异.例如, 相比于落叶物种, 常绿物种通常具有长的叶寿命、低的比叶面积(SLA)、低的光合速率和高的养分含量(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; Li et al., 2008); 另外, 由于常绿物种为了确保在冬季低温等不利环境下叶片不凋落, 需要水分运输系统保持畅通, 确保水分运输安全, 其木质部需要比落叶物种具有更强的抵抗冰冻引起栓塞的能力(Cavender-Bares & Holbrook, 2001; Cavender-Bares et al., 2005), 致使其投入在支撑结构上的成本增加, 表现出在相同的茎投入时, 常绿物种通常比落叶物种具有更小的总叶面积和单叶面积(Sun et al., 2006; Yang et al., 2009).这种不同的成本-收益关系将有可能导致常绿和落叶物种采取不同的枝条伸展策略以改变小枝上的叶排布方式, 使两类物种都能够捕获足够的光能以保持相对稳定的碳收益.因此, 两类物种可能会具有不同的茎结构与展叶效率之间的变化关系. ...
... 常绿与落叶是植物通过长期进化适应形成的两种不同的生活史对策, 它们在叶寿命和叶片结构(如比叶面积)(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; Li et al., 2008)、茎木质部抗栓塞能力(Cavender-Bares & Holbrook, 2001; Cavender- Bares et al., 2005)等许多方面都显著不同.在本研究中, 虽然当年生小枝茎结构性状与表征展叶效率的性状之间的负相关关系斜率在常绿和落叶物种间无显著差异, 但截距差异显著.在3个表征展叶效率的性状中, 尽管常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 在某一给定的茎长或茎纤细率下, 常绿物种比落叶物种具有更多的叶数量, 即具有较高的出叶强度, 与以前相关研究发现的叶大小与叶数量之间是相互权衡的结果一致(Corner, 1949; Kleiman & Aarssen, 2007; Yang et al., 2008).但由于常绿物种的单叶面积16.21 cm2显著小于落叶物种的单叶面积34.66 cm2 (p < 0.001, 表2), 导致其叶面积比(单位茎质量的叶面积)和叶茎生物量比(单位茎质量的叶质量)与小枝茎结构性状之间的关系截距反而显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低.对数据结果进一步分析表明, 常绿物种更低展叶效率的主要原因是其比叶面积显著小于落叶物种 (p < 0.001, 表2), 因为我们的结果显示, 在不考虑小枝茎长结构影响的情况下, 茎叶生物量比在常绿和落叶物种中没有显著差异(p = 0.335, 表2), 表明总生物量在叶和茎之间的分配比例在常绿和落叶物种中是没有显著差异的, 可是落叶物种的单叶面积(p < 0.001, 表2)和总叶面积(p = 0.001, 表2)都要显著大于常绿物种, 并且常绿物种茎干质量与总叶面积相关关系共同斜率下的截距显著小于落叶物种(p ˂ 0.05, 表1), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积.许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al., 1992; Westoby et al., 2002; Wright et al., 2004; 潘少安等, 2015).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT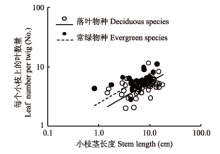 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}