0 引言
【研究意义】甜叶菊(Stevia rebaudiana)是菊科、甜叶菊属多年生草本植物。甜叶菊叶片富含黄酮类和甜菊糖苷类化学成分[1,2]。黄酮类化合物具有抗肿瘤、抗氧化等多种药理活性;甜菊糖苷(steviol glycosides)则是一类新型天然甜味剂,具有甜度高、热量低、安全无毒等特点[3],逐渐成为食品和医药领域研究开发的热点[4,5]。2017年笔者在江苏省东台市富安镇甜叶菊生产基地发现甜叶菊叶片上呈现不同程度的褐斑,发病严重的植株叶片全部枯萎,严重影响了甜叶菊的产量。因此,确定甜叶菊褐斑病的致病病原菌,对该病害的防治具有重要意义。【前人研究进展】链格孢菌是自然界中广泛存在、危害严重的一种死体营养型植物病原真菌,多生于植物的枯死部分和衰弱濒死的组织,或腐生于多种有机物质上或土壤中[6]。链格孢菌可引起多种植物特别是农作物病害,能够引起包括小麦、马铃薯、玉米、烟草、番茄、苹果、梨等几十种农作物的真菌性病害,严重影响作物的产量和品质,造成巨大的经济损失[7,8]。美国伊利诺斯州的大豆曾因细极链格孢(Alternaria tenuissima)引起的大豆猝倒病,造成产量损失15%[8]。由长柄链格孢(A. longipes)和链格孢(A. alternata)引起的烟草赤星病在世界各主要烟区广泛发生,主要危害成熟期烟叶,对成熟度、采收率、外观等级和内在质量都有很大影响,是烟草的主要病害之一[9,10]。我国各地广泛发生的番茄、马铃薯等茄科蔬菜的早疫病,则是由A. solani为主的病原真菌引起的[11,12]。1982年,日本首次发现一种链格孢属真菌A. steviae能引起甜叶菊黑斑病[13],病斑呈黑色不规则形状扩展,并被褪绿区域包围;2007年,印度也首次报道了导致甜叶菊叶斑病的病原菌,鉴定结果为链格孢[14];伊朗于2015年首次分离鉴定到链格孢菌属为引起甜叶菊叶斑病的致病病原菌[15]。【本研究切入点】目前,国内关于甜叶菊病害病原物鉴定方面的报道还很少,对链格孢菌的防控并没有非常有效的方法,因此了解其侵染机理可为该真菌导致的病害防控提供指导。对2017年7月采自江苏省东台市富安镇甜叶菊生产基地的带病斑甜叶菊叶片进行病原菌分离鉴定和致病性检测,并进一步分析MeJA对甜叶菊响应链格孢菌的影响。【拟解决的关键问题】利用形态学与分子生物学手段鉴定甜叶菊褐斑病的致病病原菌,明确甜叶菊褐斑病的病原菌种类;分析MeJA对甜叶菊响应链格孢菌的影响以及JA通路是否参与该响应过程,为该病害的田间诊断、综合防控及甜叶菊的抗病育种提供理论依据。1 材料与方法
试验于2017年7月至2018年4月在江苏省农业科学院经济作物研究所完成。1.1 褐斑病标样采集及病原菌分离
2017年7月从江苏省东台市富安镇甜叶菊生产基地采集叶片带有褐斑的植株。选取叶片病健交界的组织,切成0.5 cm×0.5 cm的小块,首先用无菌水冲洗数次,用70%酒精表面消毒30 s,再用0.1%升汞处理2 min,用无菌水漂洗3次,最后用灭菌镊子夹取材料平铺于添加有链霉素(40 μg·mL-1)的PDA平板培养基上,25℃黑暗条件下培养3—5 d,从新长出的菌落边缘挑取少量菌丝接种到新的PDA平板上进一步分离纯化,记录菌落的形态学特征。1.2 病原菌的致病性测定
选择健康无病、无伤痕的甜叶菊叶片,室内进行离体叶片接种试验。每个菌株均为3次重复。用70%酒精进行表面消毒后置于铺有保湿滤纸的培养皿内。菌丝块接种法:分离纯化的菌株在PDA平板上培养7 d,用灭菌刀沿菌落边缘切取3 mm×3 mm大小的菌饼,菌丝面朝下、避开主叶脉贴于叶片背面两侧,接种部位用刺针进行人工创伤。同时接种空白的PDA培养基块作为对照。接种后封好培养皿,置于25℃条件下诱导发病,每天观察并记录发病情况。
分生孢子悬浮液接种法:将菌株在PDA平板上于黑暗条件下培养10 d,加入10 mL无菌水刮洗菌丝,所得到的悬浮液经3层纱布过滤,在显微镜下使用血球计数板来计算孢子悬浮液的浓度,最终将孢子悬浮液浓度调至1×105个/mL。将表面消毒处理后的甜叶菊叶片浸到孢子悬浮液中10 min,然后将其转移到铺有保湿滤纸的培养皿内,于25℃条件下培养,在光学显微镜下观察分生孢子的萌发以及侵染情况。
待接种后的甜叶菊叶片发病后,根据柯赫式法则,将病原菌重新进行分离纯化,观察新分离物与接种菌是否相同。
1.3 病原菌形态学观察
将分离得到的致病菌株接种到PDA培养基上,置于25℃培养箱中黑暗培养8 d后,从菌落边缘表面切取小块薄的、带菌丝的培养基,将其放置到载玻片上,覆上中心凿有1 cm2方孔的纸片,盖上盖玻片,在光学显微镜(Olympus CX41,Japan)下观察分生孢子、分生孢子梗及分生孢子链的形态并拍照,用目镜测微尺测量分生孢子的大小。结合病原菌菌落在PDA培养基上的形态及色泽特征,参考《真菌鉴定手册》[16]确定病原菌的种类。1.4 病原菌的分子生物学鉴定
1.4.1 总DNA的提取 将分离得到的菌株接种到PDA培养基上,置于25℃培养箱中培养10 d,从培养基表面刮取菌丝,加入液氮迅速研磨成粉末,参照真菌基因组DNA提取试剂盒(Solarbio,China)操作说明提取菌株DNA,并将DNA样品放于-20℃保存备用。1.4.2 ITS区扩增 采用真菌核糖体基因转录间隔区的通用引物ITS1(5′-TCCGTAGGTGAACCTGCGG -3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′)对病原菌的基因组DNA进行PCR扩增。PCR反应体系50 μL:DNA模板2 μL、上下游引物ITS1和ITS4各1 μL、10×PCR buffer(Mg2+ plus)5 μL、dNTP Mixs(10 mmol·L-1)1 μL、Taq DNA聚合酶1 μL、无菌ddH2O补齐至50 μL。PCR扩增程序:94℃预变性5 min,94℃变性30 s,56℃退火30 s,72℃延伸40 s,共33个循环,最后72℃延伸10 min,4℃保存。
1.4.3 PCR产物纯化与克隆 取5 μL 1.4.2中的PCR扩增产物加入1 μL 6×DNA loading buffer混匀,于1%琼脂糖凝胶上检测样品条带大小是否正确并特异。检测正确的PCR产物使用PCR清洁试剂盒(Axygen,USA)回收纯化,将其与pEASY-T1克隆载体(TransGen,China)连接,连接产物转入大肠杆菌Trans1-T1感受态细胞(TransGen,China)中,经PCR鉴定后,选取阳性克隆送上海英骏生物技术有限公司进行测序。
1.4.4 病原菌的ITS序列分析 将测得的ITS区序列在GenBank中进行同源性搜索,与已报道真菌菌株的ITS区序列进行同源性比较。利用MEGA 7软件的邻接法(neighbor-joining,NJ)构建系统发育树。将比较结果与病菌的形态特征、培养性状和致病性结合,对病原菌进行鉴定。
1.5 细胞学染色
等体积的乳酸、苯酚、甘油和无菌ddH2O混合,制成乳酸酚溶液;向其中加入Trypan Blue染料溶解,使其终浓度为2.5 mg·mL-1,配成Trypan Blue染液。Trypan Blue染液中加入2倍体积的无水乙醇,将样品置于染色液中100℃煮1 min,室温放置5—10 min;转移至无水乙醇﹕乳酸酚=2﹕1溶液中过夜脱色,重复直至叶片绿色完全褪去;样品于70%甘油中保存,制片并观察。
1.6 MeJA对链格孢菌生长的影响
1.6.1 MeJA培养基的配制 取223.6 μL的MeJA溶于10 mL无水乙醇中,配成100 mmol·L-1的浓缩液,分别加入到PDA培养基中配成0、40、100、200、500 μmol·L-1含不同浓度MeJA的混合培养基。1.6.2 菌丝生长检测 链格孢菌株ST3在PDA培养基上培养5 d后,用打孔器在菌落边缘均匀打下菌丝块,将菌丝块分别放在上述含不同浓度MeJA的PDA平板中央,每个处理重复6次。将其置于25℃培养箱中黑暗培养,每2 d观察并测量菌落直径。
1.7 离体叶片饲喂及培养
选择生长状态良好、长势一致的甜叶菊植株,剪取离体叶片放入铺有一层滤纸的培养皿中,滤纸加灭菌水浸湿,用浸泡100 μmol·L-1 MeJA溶液的无菌棉包裹叶柄切口,进行离体饲喂,对照用浸泡无菌水的棉球处理。室温下静置10 h后取下棉球,重新用无菌水浸湿的棉球包裹叶柄处(方法参照SUN等[17])。接种方法参照1.2的菌丝块接种法。接种后的叶片置于培养皿中于温度28/23℃、光周期16/8 h的光照培养箱中培养,分别于接种后3、4、5 d测量病斑并拍照记录。1.8 RNA提取及qRT-PCR
样品RNA的提取采用E.Z.N.A.? Plant RNA Kit(OMEGA,美国)试剂盒,按照说明书进行RNA的提取操作。基因组DNA去除及RNA反转录采用PrimeScriptTM RT Reagent Kit(TaKaRa,Japan)试剂盒并按说明书操作。qRT-PCR采用SYBR? Premix Ex TaqTM(Tli RNaseH Plus)(TaKaRa,Japan)试剂盒,PCR反应在QuantStudio 5 Real-Time PCR System(Applied Biosystems,American)仪器上进行。PCR反应结束后扩增Ct值经QuantStudioTM Design & Analysis Software 1.3.1软件初步处理后导出至Excel中,利用2-ΔΔCt方法计算目的基因的相对表达量。数据采用Dunnett’ s test进行显著性分析。所用甜叶菊内参基因26 S rRNA及JA通路各目的基因引物序列如表1所示。
Table 1
表1
表1本研究中所用引物核酸序列
Table 1List of qPCR primer sequences used in this study
| 引物名称 Primer name | 核酸序列 Primer sequence (5′-3′) | 基因登录号 Accession number |
|---|---|---|
| SrJAR1-F SrJAR1-R SrLOX3-F SrLOX3-R SrJAZ1-F SrJAZ1-R SrJAZ4-F SrJAZ4-R 26 S rRNA-F 26 S rRNA-R | ACCGATGTCAAGCTCGGTGA GGGTCGCGTTATGGAACCCA TGCTTTGGCTCGAGAAGCAC TCAGCTGGCAATGCTTCATGGT GCCGTGCAGAAGCAGCATGA CGTCTGGTTTCGATGGTTTCCG GCACCACAACCAAGGACCCA GAGACGGGTGAGCAGGGTTG CAAAACGACTCTCGGCAACG CACACACGAGACACTCCGAT | BG523392.1 BG524650.1 BG525787.1 BG526796.1 AB457301.1 |
新窗口打开
2 结果
2.1 病原菌的分离纯化及形态学特征
从甜叶菊守田3号发病叶片中分离出7种疑似致病真菌,依次编号为ST1—ST7。所有菌株在PDA培养基上呈近圆形等径辐射生长,边缘光滑略呈波浪状;气生菌丝较为发达,初期为白色,后期逐渐变为不同程度的灰黑色,基质呈现不同类型的同心轮纹(图1);在PDA平板上培养8 d的菌落产生大量的分生孢子,显微镜下观察,分生孢子梗直立或略弯,分隔,偶分枝,淡褐色,大小为(24.0—77.5)×(2.3—5.0)μm;分生孢子单生或成链,形态多样,多为近球形、倒棒状或倒梨形,大小为(20.5—45.5)×(6.5—16.0)μm(图2)。其中ST2、ST3、ST5、ST6具有短分枝的孢子链,作合轴式延伸,分生孢子表面光滑或具微刺,具3—8个横膈膜和1—4个纵、斜隔膜;ST1、ST4、ST7则形成超过10个孢子的分生孢子长链,少分支,处于链基部的孢子,表面偶生明显的疣突,分生孢子具4—7个横膈膜,1—4个纵或斜隔膜,常有1—4个主横膈膜,主隔膜处有明显的隘缩。根据这些特征,参照魏景超的《真菌鉴定手册》[16]及张天宇的《中国真菌志》[6]初步判定此类菌为链格孢属霉菌,其中ST2、ST3、ST5、ST6可能是Alternaria alternata,ST1、ST4、ST7则推测是Alternaria tenuissima。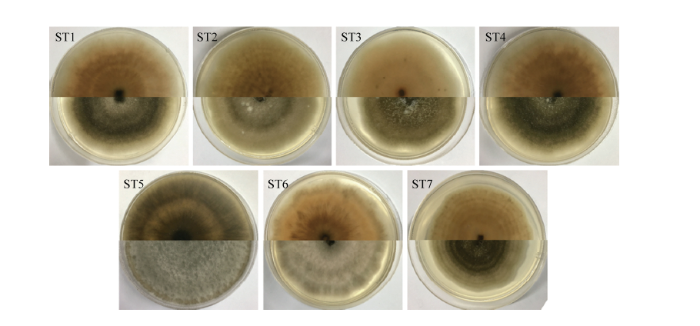 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1分离菌株菌落形态特征
ST1—ST7在PDA平板上25℃条件下培养10 d后的正面和反面菌落形态
-->Fig. 1Morphological characteristics of isolated strains
Colony morphology of ST1-ST7 which cultured on potato dextrose agar at 25℃ for 10 days, from above and below
-->
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2分离病原真菌分生孢子形态的显微观察
-->Fig. 2Conidial morphology of 7 isolated strains
-->
2.2 病原菌的致病性检测
采用离体叶片接种的方法,对叶片发病情况进行观察统计,结果发现7个菌株对甜叶菊叶片致病程度存在一定的差异,ST2、ST3和ST7 3个菌株侵染叶片后病斑扩展速度快,在接种后72 h病斑直径达到3—5 mm,发病位置呈黑褐色(图3-B、3-C、3-G),而ST1、ST4、ST5和ST6在接种后的第7天病斑仍局限于叶片的创伤位点(图3-A、3-D、3-E、3-F)。因此,引起甜叶菊叶片褐斑病的ST2、ST3和ST7菌株的致病力较强,而ST1、ST4、ST5和ST6对甜叶菊的致病力则相对较弱。从接种发病的叶片病斑上刮取少量的病组织镜检,可观察到与接种菌株一致的菌丝和分生孢子。对发病组织进行病原菌的再分离,同样获得与接种病原真菌菌落形态一致的培养物,完成柯赫氏法则(Koch postulates)致病性检测。因此,接种所用的菌株是引起甜叶菊褐斑病的病原菌。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3甜叶菊叶片分别接种7株分离病原真菌后的表型
A—G:甜叶菊叶片分别接种ST1-ST7菌株的表型;H:接种无菌空白PDA培养基的对照
-->Fig. 3Symptoms on S. rebaudiana leaves after inoculated with isolated fungi strains ST1-ST7, respectively
The phenotype of S. rebaudiana leaves inoculated with ST1-ST7 strains, respectively;The control leaf inoculated with PDA medium
-->
2.3 病原菌的rDNA-ITS序列分析
以菌株ST1—ST7的基因组DNA为模板,利用真菌rDNA-ITS区通用引物ITS1和ITS4,扩增得到571、569、570、570、570、571和570 bp大小的ITS序列(图4),并进行测序分析,将测序结果提交至NCBI网站进行BLAST比对(图5)。结果表明,ST2、ST3、ST5、ST6与KY075667.1、KY814634.1等8个菌株(Alternaria alternata、Alternaria sp.)的相似性达到99%—100%,ST1、ST4、ST7与HQ402558.1(Alternaria tenuissima)的相似性达到99%,结合病原菌的形态学观察结果,进一步确认引起甜叶菊褐斑病的病原菌为链格孢和细极链格孢。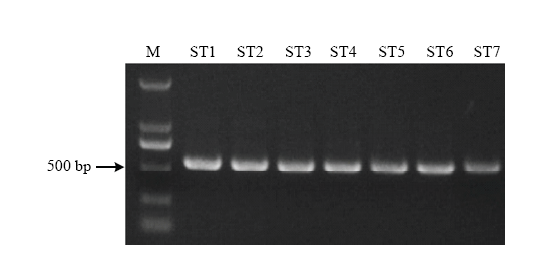 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图47株链格孢菌的rDNA-ITS PCR产物电泳图
-->Fig. 4PCR results of 7 Alternaria rDNA-ITS
-->
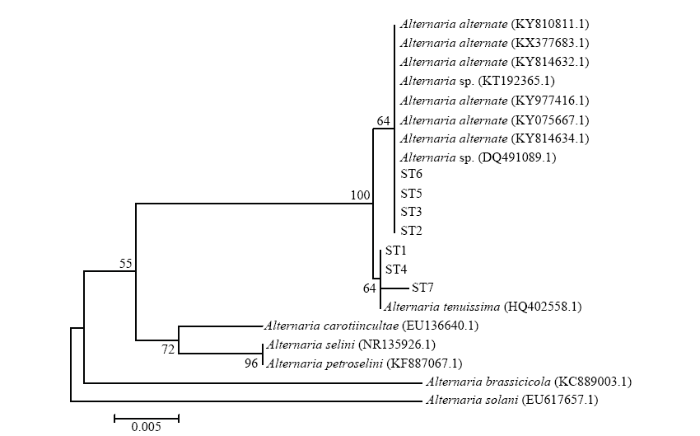 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5甜叶菊褐斑病7个致病菌与链格孢菌属相关种rDNA-ITS序列系统发育树
-->Fig. 5Phylogenetic tree of 7 strains from S. rebaudiana and related species of Alternaria based on sequences of rDNA-ITS region
-->
2.4 细极链格孢分生孢子在叶片上的萌发及侵入
甜叶菊品种守田3号叶片接种细极链格孢ST7(以下试验将该菌株作为研究对象)分生孢子悬浮液(1×105个/mL),利用光学显微镜观察发现,接种6 h后,分生孢子已经开始萌发,分生孢子可以从孢子的头部、侧面、尾部多个位置萌发(图6-A、6-B、6-C),菌丝沿叶片表面蔓延生长。接种24 h后,对菌丝侵入表皮细胞的方式进行观察,发现一部分菌丝通过叶片的气孔侵入表皮细胞(图6-D、6-E、6-F),一部分则从表皮细胞间隙直接侵入叶片表皮细胞内(图6-G、6-H)。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6细极链格孢分生孢子在甜叶菊叶片上的萌发与侵入
甜叶菊品种守田3号接种细极链格孢ST7 6 h后,分生孢子从孢子的头部(A)、侧面(A、B)、尾部萌发(C);接种24 h后,菌丝从叶片气孔(D、E、F)、表皮细胞间隙(G、H)侵入叶片表皮细胞内
-->Fig. 6Germination and invasion of A. tenuissima conidia on S. rebaudiana leaves
The A. tenuissima conidia of ST7 germinated from the head (A), lateral (A, B) and caudal (C) of the spores on the S. rebaudiana leaves of cultivar Shoutian 3 at 6 h post inoculation; mycelium invaded into leaf epidermal cells from the leaf stomata (D, E, F) and epidermal cell gap (G, H) at 24 h post inoculation
-->
2.5 JA通路参与甜叶菊对链格孢菌的响应
2.5.1 MeJA对链格孢菌菌丝生长的影响 选择细极链格孢菌株ST7在含有不同浓度MeJA的PDA培养基上培养,菌丝生长面积统计结果如图7所示。MeJA浓度低于200 μmol·L-1时,菌落生长面积与对照相比无明显差异;当MeJA浓度增至200 μmol·L-1时,链格孢菌菌丝的生长受到明显抑制,且随着MeJA浓度增至500 μmol·L-1,对其生长的抑制效果更显著。MeJA在较低浓度范围内对细极链格孢菌ST7没有直接的抑制作用,因此,选择浓度为100 μmol·L-1的MeJA用于后续的试验处理。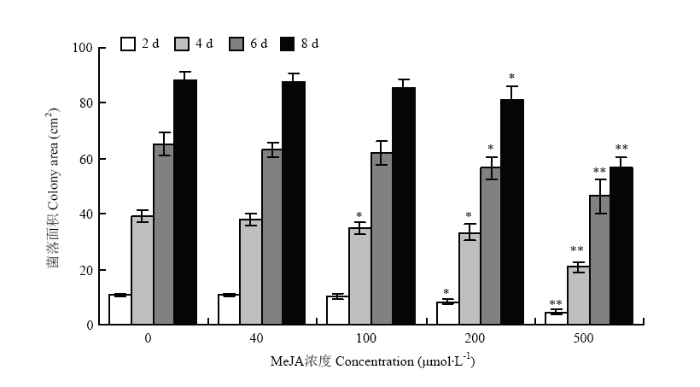 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图7MeJA对细极链格孢ST7菌丝生长的影响
-->Fig. 7Effect of MeJA on mycelial growth of A. tenuissima ST7 on PDA medium
-->
2.5.2 外源施加MeJA增强甜叶菊对细极链格孢的抗性 对外源施加100 μmol·L-1 MeJA的甜叶菊离体叶片接种细极链格孢ST7菌丝块,同时对叶片接种无菌PDA培养基作为空白对照。接种后的病斑表型如图8-A所示,相比于对照H2O处理的叶片,MeJA处理后甜叶菊叶片的病斑明显减小。病斑统计结果(图8-B)显示,接种后3、4、5 d,对照叶片的病斑面积分别是MeJA处理后叶片病斑面积的1.41、1.50和1.58倍,结果表明外源施加MeJA后,甜叶菊叶片对细极链格孢ST7的抗性明显增强。
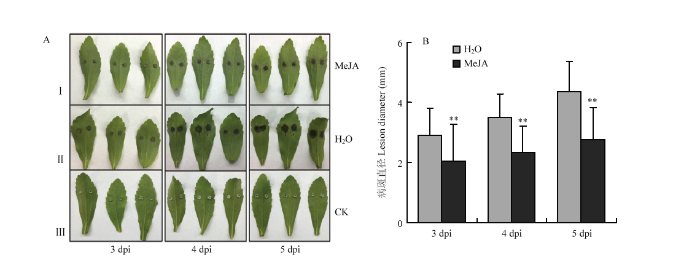 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图8MeJA影响甜叶菊叶片对细极链格孢ST7的抗性
I:甜叶菊叶片饲喂100 μmol·L-1 MeJA 10 h后接种细极链格孢ST7的表型;叶柄处饲喂ddH2O后接种细极链格孢ST7的表型;叶片饲喂ddH2O并接种无菌PDA培养基的空白对照;拍照及测量病斑时间点均为接种后3、4、5 d
-->Fig. 8Exogenous application of MeJA enhances the resistance of S. rebaudiana leaves to A. tenuissima
The phenotype of S. rebaudiana leaves which inoculated with ST7 after feeding 100 μmol·L-1 MeJA for 10 h;II: The phenotype of S. rebaudiana leaves which inoculated with A. tenuissima ST7 after petiole feeding ddH2O;III: The control which leaves inoculated with PDA medium。 The time points for taking pictures and measuring the disease spots are 3, 4, 5 days after inoculation
-->
2.5.3 JA通路相关基因的表达分析 LOX是JA合成通路基因,在甜叶菊叶片接种细极链格孢菌株ST7分生孢子悬浮液12 h后,LOX3明显上调表达;JAR则是JA通路中的信号传导因子,在接种后48 h后,JAR1的表达量显著增加;JAZ是茉莉酸途径的抑制因子,接种链格孢菌后,JAZ1和JAZ4呈现不同程度的下调表达(图9)。表明JA通路相关基因参与甜叶菊对细极链格孢ST7的响应。
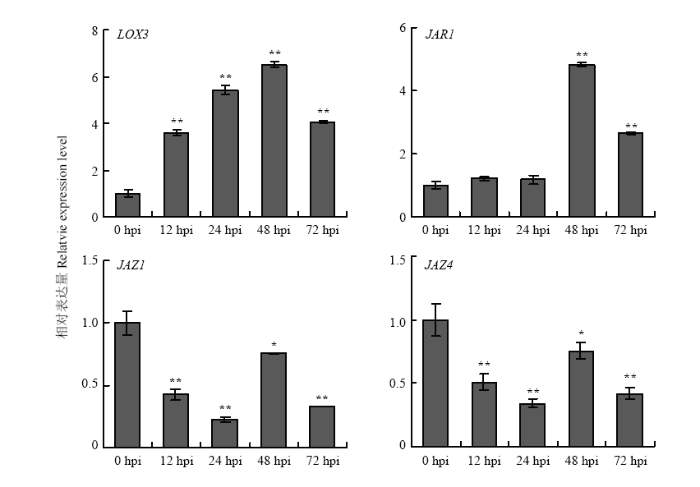 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图9JA通路相关基因响应细极链格孢侵染的表达分析
-->Fig. 9Expression pattern of JA pathway related genes in response to A. tenuissima infection
-->
3 讨论
甜叶菊提取物甜菊糖苷作为甜味食品添加剂在南美已使用了数个世纪[18],最近在欧盟和美国也被批准使用[19]。甜菊糖苷同时具有一定的药用价值,例如降低血压、增强胰岛β细胞功能治疗2型糖尿病、预防动脉粥样硬化、预防某些癌症以及清除人体自由基等功能[19,20,21,22]。我国的甜叶菊自1976年南京中山植物园从日本引种试验成功后,80年代初全国各地开始种植[23]。自2006年起,中国成为世界甜菊糖产品最大的出口国[4]。目前关于甜叶菊病害已有相关报道。2002年,朱东顺等[24]在山东省高密地区发现甜叶菊生产中发生的主要病害有立枯病、花叶病毒病和叶斑病等,对甜叶菊的干叶质量和产量均造成不同程度的影响。近年来甘肃河西走廊地区甜叶菊育苗期的病害呈逐年加重的趋势,猝倒病、立枯病和叶斑病成为育苗期最容易发生的3大病害,给农业生产带来较大损失[25]。2017年,笔者在江苏省东台市富安镇甜叶菊生产基地调查发现甜叶菊叶片上呈现不同程度的褐斑,发病严重的植株叶片全部枯萎。经病原菌的分离、培养、纯化,共分离得到7株病原真菌,均为链格孢属真菌,致病性鉴定结果发现有3株致病力较强,为细极链格孢和链格孢。JA是植物细胞间和细胞内的重要信号分子,通过与转录因子间的相互作用来调控防御蛋白的表达以及次生物质的合成,参与植物对病原菌的应答反应和信号传递[26,27]。本试验表明,100 μmol·L-1的MeJA对链格孢菌无直接抑制作用,而用该浓度MeJA处理后的甜叶菊叶片对链格孢菌的抗性明显增强,具有显著的诱抗作用,说明MeJA在增强甜叶菊抗链格孢菌方面起着重要作用。脂氧合酶(lipoxygenase)基因LOX是JA合成通路中的关键基因,当植物受到伤诱导时,LOX被激活,诱导JA及MeJA的合成与积累,而生成的JA又可进一步激活LOX,促进JA的积累[28],在甜叶菊叶片受链格孢菌侵染后,LOX3的表达水平明显上调。JAZ蛋白作为E3泛素连接蛋白降解复合体SCF的靶蛋白,是JA信号转导途径的关键组分,其能与茉莉酸信号途径的转录因子(如MYC2、MYC3)相结合从而抑制这些因子的转录活性,JAZ蛋白是茉莉酸信号途径重要的负调控因子[29],本研究用链格孢菌侵染甜叶菊后,叶片中JAZ1、JAZ4的表达受到抑制,与接种前相比,整体呈下调表达的趋势。JAR1是一种茉莉酸-异亮氨酸(JA-Ile)合成酶,被认为是拟南芥JA通路中必需的信号转导因子[30],本研究中甜叶菊JAR1响应链格孢菌的侵染,接种48 h后表达量显著上调,表明在甜叶菊受链格孢菌侵染时,JA信号通路基因的表达发生相应的改变,提高植物体内JA水平,对病原菌的侵染作出响应。
4 结论
根据真菌形态鉴定、致病性鉴定以及病原菌rDNA-ITS序列同源性分析结果,确定甜叶菊褐斑病的致病菌为链格孢及细极链格孢。显微观察发现细极链格孢菌丝可以从叶片的气孔以及表皮细胞间隙侵入表皮细胞。外源施加MeJA能够有效抑制细极链格孢菌丝的生长,MeJA处理后的甜叶菊叶片对细极链格孢的抗性明显增强,具有显著的诱抗作用,在甜叶菊褐斑病的防治中具有很好的应用前景。The authors have declared that no competing interests exist.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | . . |
| [2] | . |
| [3] | . Stevia rebaudiana (Bertoni) is a herbaceous perennial plant (2n1722) of genus Stevia Cav., which consists of approximately 230 species of herbaceous, shrub and sub-shrub plants. Leaves of stevia produce diterpene glycosides (stevioside and rebaudiosides), non-nutritive, non-toxic, high-potency sweeteners and may substitute sucrose as well as other synthetic sweetners, being 300 times sweeter than sucrose. In addition to its sweetening property, it has medicinal values and uses. Stevia is self-incompatible plant and the pollination behaviour is entomophilous. Rebaudioside-A is of particular interest among the glycosides produced in the leaves of stevia because of the most desirable flavour profile, while, stevioside is responsible for aftertaste bitterness. Development of new varieties of S. rebaudiana with a higher content of rebaudioside-A and a reduced content of stevioside is the primary aim of plant breeders concerned with the improvement and utilization of this source of natural sweeteners. The proportions of rebaudioside-A and -C are controlled by a single additive gene known to be co-segregating suggesting synthesis by the same enzyme. Stevioside and rebaudioside-A are negatively correlated, while rebaudioside-A and -C are positively correlated. Conventional plant breeding approaches such as selection and intercrossing among various desirable genotypes is the best method for improving quality traits in a highly cross-pollinated crop like stevia. Various plant types with larger amounts of specific glycoside have already been patented, such as RSIT 94-1306, RSIT 94-75, RSIT 95-166-1 through selection and intercrossing. Composites and synthetics can be used to capture part of the available heterosis because of the high degree of natural out-crossing and the absence of an efficient system of pollination control. Synthetics and composites like ‘‘AC Black Bird’’ and ‘‘PTA-444’’ have already been developed. Polyploidy results in better adaptability of individuals and increased organ and cell sizes. Tetraploids have larger leaf size, thickness and have potential use in increasing biomass and yield in comparison with diploid strains. Characters of interest with low variability in the population may be improved through mutation breeding. Use of biotechnological approaches, such as tissue culture for the mass propagation of elite genotypes, anther culture for development of pure homozygous doubled haploid and molecular marker technology for identification of marker loci linked to rebaudioside-A trait, can create new opportunities for plant breeders. Understanding the mechanism and pathway of biosynthesis of steviol glycosides can help to improve the glycoside profile by up-regulation and down-regulation of genes. |
| [4] | . 介绍了全球甜叶菊的发展历史、甜菊糖苷等产品的提取方法、作为糖源的机理、安全性、各国或地区准许使用与合法地位、甜叶菊产量、市场额度以及甜叶菊产业发展的未来展望. . 介绍了全球甜叶菊的发展历史、甜菊糖苷等产品的提取方法、作为糖源的机理、安全性、各国或地区准许使用与合法地位、甜叶菊产量、市场额度以及甜叶菊产业发展的未来展望. |
| [5] | . 在介绍甜菊糖优点的基础上,总结了甜菊糖的主要功能,包括作为糖尿病人的甜味剂和辅助治疗剂、改善胃肠功能、降低血液中胆固醇及甘油三酯的含量、降低血压、抗过敏、预防龋齿等口腔疾病、减肥及预防心血管疾病功能、用于肥料及饲料等,以为甜菊糖的应用提供参考。 . 在介绍甜菊糖优点的基础上,总结了甜菊糖的主要功能,包括作为糖尿病人的甜味剂和辅助治疗剂、改善胃肠功能、降低血液中胆固醇及甘油三酯的含量、降低血压、抗过敏、预防龋齿等口腔疾病、减肥及预防心血管疾病功能、用于肥料及饲料等,以为甜菊糖的应用提供参考。 |
| [6] | |
| [7] | . |
| [8] | [D]. [D]. |
| [9] | . |
| [10] | . [Objective] The paper was to study the effect of tobacco blown spot on the yield and output value of tobacco leaf.[Method]The upper,middle and lower leaves in tobacco plant were selected during the harvest period of tobacco to carry out loss rate estimation of yield and output value of tobacco leaf caused by different harmful levels of brown spot.Regression correlation analysis was also conducted.[Result]The disease levels of brown spot had extremely significant negative correlation with single-leaf weight of tobacco leaf,and it had extremely significant positive correlation with the loss rate of single-leaf weight.The increase speed of loss rate of single-leaf weight of middle and upper leaves was obviously faster than that of lower leaves.The loss rates of single-leaf weight of upper,middle and lower leaves were 10.07%-26.90%,3.43%-28.88% and 3.18%-28.95%,respectively.The higher the disease level of blown spot was,the lower the yield and output value of tobacco leaf was,and the corresponding loss rate was also higher.Correlation analysis showed that the disease level of blown spot had extremely significant negative correlation with the yield and output value of tobacco leaf,and it had extremely significant positive correlation with the loss rate of yield and output value.The negative impact of blown spot on the output value of tobacco leaf was far greater than that on the yield.The highest loss rate of the yield of tobacco leaf was 28.56%,while the highest loss rate of output value reached 89.67%.[Conclusion] The study provided theoretical basis for accurately holding the critical period for the control of blown spot,thus reducing the damage on tobacco leaf and improving the output value of tobacco leaf. |
| [11] | . 对甘肃省马铃薯贮藏期真菌性病害的病原菌进行了分离鉴定,共鉴定到8种病原真菌,引起6种贮藏期真菌病害,分别为:致病疫霉(Phytophthora infestans)引起马铃薯晚疫病,茄链格孢(Alternaria solani)引起马铃薯早疫病,茄丝核菌(Rizoctonia solani)引起马铃薯黑痣病,具柄细基束梗孢(Cephalotri- chum stemonitis)引起马铃薯褐腐病,Verticillium tricopsis引起马铃薯萎蔫病,Fusarium solani和F.sambucinum引起马铃薯干腐病.马铃薯为V.tricopsis的新寄主,具柄细基束梗孢(C.stemonitis)引起马铃薯褐腐病、F.sambucinum引起马铃薯干腐病在我国属首次报道. . 对甘肃省马铃薯贮藏期真菌性病害的病原菌进行了分离鉴定,共鉴定到8种病原真菌,引起6种贮藏期真菌病害,分别为:致病疫霉(Phytophthora infestans)引起马铃薯晚疫病,茄链格孢(Alternaria solani)引起马铃薯早疫病,茄丝核菌(Rizoctonia solani)引起马铃薯黑痣病,具柄细基束梗孢(Cephalotri- chum stemonitis)引起马铃薯褐腐病,Verticillium tricopsis引起马铃薯萎蔫病,Fusarium solani和F.sambucinum引起马铃薯干腐病.马铃薯为V.tricopsis的新寄主,具柄细基束梗孢(C.stemonitis)引起马铃薯褐腐病、F.sambucinum引起马铃薯干腐病在我国属首次报道. |
| [12] | . . |
| [13] | . |
| [14] | . |
| [15] | . |
| [16] | |
| [17] | . Abstract Alternaria alternata (tobacco pathotype) is a necrotrophic fungus causing severe losses in Nicotiana species by infection of mature leaves. Similar to what has been observed in cultivated tobacco, N. tabacum, young leaves of wild tobacco, N. attenuata, were more resistant to A. alternata than mature leaves, and this was correlated with stronger blue fluorescence induced after infection. However, the nature of the fluorescence-emitting compound, its role in defence, and its regulation were not clear. Silencing feruloyl-CoA 6'-hydroxylase 1 (F6'H1), the gene encoding the key enzyme for scopoletin biosynthesis, by virus-induced gene silencing (VIGS) revealed that the blue fluorescence was mainly emitted by scopoletin and its -glycoside form, scopolin. Further analysis showed that scopoletin exhibited strong antifungal activity against A. alternata in vitro and in vivo. Importantly, jasmonic acid (JA) levels were highly elicited in young leaves but much less in mature leaves after infection; and fungus-elicited scopoletin was absent in JA-deficient plants, but was largely restored with methyl jasmonate treatments. Consistent with this, plants strongly impaired in JA biosynthesis and perception were highly susceptible to A. alternata in the same way scopoletin/scopolin-depleted VIGS F6'H1 plants. Furthermore, silencing MYC2, a master regulator of most JA responses, reduced A. alternata-induced NaF6'H1 transcripts and scopoletin. Thus, it is concluded that JA signalling is activated in N. attenuata leaves after infection, which subsequently regulates scopoletin biosynthesis for the defence against A. alternata partly through MYC2, and higher levels of scopoletin accumulated in young leaves account for their strong resistance. The Author 2014. Published by Oxford University Press on behalf of the Society for Experimental Biology. |
| [18] | . |
| [19] | . Foodstuff is usually contaminated by more than one mycotoxin, however toxicological data are lacking as regards the effects in combinations compared to their individual effect. This study investigated the in vitro effects of enniatins (ENs) A, A 1 , B and B 1 , alone and in combinations, on Caco-2 cells viability by MTT assay after 24h of exposure. Cells were treated with concentrations ranging from 0.9 to 15.0μM, individually and in combination of two, three and four mycotoxins. Dose–response curves were generated for each mycotoxin and the isobologram method was used to determine the interactive effects of tested mixtures. Tested ENs produced significant cytotoxic effects both individually and in combination in a dose-dependent manner. IC 50 values obtained for all individually tested mycotoxins ranged from 1.3 to >15μM. In ENs combination tests, synergistic effect in Caco-2 viability are observed for EN B+EN A 1 , EN B 1 +EN A 1 and EN A+EN A 1 +EN B (CI=0.33–0.52). All other combinations showed additive effect at medium and high affected fraction with exception of lower fraction affected and the EN B+EN B 1 mixture that produced antagonistic effect (CI=1.76–10.36). The use of combination index-isobole method could help to better understand the potential interaction between co-occurring mycotoxins and may contribute to their risk assessment. |
| [20] | . |
| [21] | . A series of sodium 2-sulfoethyl alkanoates RCOO(CH2)(2)SO3Na was prepared from sodium isethionate and fatty acids containing 9, 11, 13, and 15 carbon atoms. The Krafft point, the critical micelle concentration (CMC), the surface tension, the emulsifying power, the wetting time, the resistance to acid hydrolysis, and the calcium-ion stability of this series of surfactants were examined, and the results were compared with those of the corresponding sodium alkyl beta-sulfopropionates ROCO(CH2)(2)SO3Na, which have another structure of ester-linkage. The CMC values of sodium 2-sulfoethyl alkanoates almost coincide with those value of the sodium alkyl beta-sulfopropionates with the same number of carbon atoms. However, the Krafft point and surface tension at the CMC of sodium 2-sulfoethyl alkanoates were lower than those of the sodium alkyl beta-sulfopropionates with the same number of carbon atoms. Sodium 2-sulfoethyl alkanoates are more stable than sodium alkyl beta-sulfopropionates in their resistance to hydrolysis and to the calcium-ion. |
| [22] | . |
| [23] | . . |
| [24] | . 山东省高密地区甜叶菊生产中发生的主要病害有立枯病、花叶病毒病和叶斑病等。本文分别介绍了各病害的症状,侵染发病条件及防治措施,对指导甜叶菊生产,做到早预防病害,早实施防治措施,把病害损失减少到最低限度,以保证甜叶菊优质高产。 . 山东省高密地区甜叶菊生产中发生的主要病害有立枯病、花叶病毒病和叶斑病等。本文分别介绍了各病害的症状,侵染发病条件及防治措施,对指导甜叶菊生产,做到早预防病害,早实施防治措施,把病害损失减少到最低限度,以保证甜叶菊优质高产。 |
| [25] | . 介绍了河西走廊甜叶菊拱棚育苗最严重的猝倒、立枯和叶斑三大病害症状及发生原因。并提出了具体的综合性防治策略。 . 介绍了河西走廊甜叶菊拱棚育苗最严重的猝倒、立枯和叶斑三大病害症状及发生原因。并提出了具体的综合性防治策略。 |
| [26] | . Unexpected roles in defense and signaling processes for oxygenated forms of fatty acids were recently uncovered, broadening the horizons of lipid biochemistry. |
| [27] | . Induced plant resistance depends on the production of specialized metabolites that repel attack by biotic aggressors and is often associated with reduced growth of vegetative tissues. Despite progress in understanding the signal transduction networks that control growth-defense tradeoffs, much remains to be learned about how growth rate is coordinated with changes in metabolism during growth-to-defense transitions. Here, we highlight recent advances in jasmonate research to suggest how a major branch of plant immunity is dynamically regulated to calibrate growth-defense balance with shifts in carbon availability. We review evidence that diminished growth, as an integral facet of induced resistance, may optimize the temporal and spatial expression of defense compounds without compromising other critical roles of central metabolism. New insights into the evolution of jasmonate signaling further suggest that opposing selective pressures associated with too much or too little defense may have shaped the emergence of a modular jasmonate pathway that integrates primary and specialized metabolism through the control of repressor-transcription factor complexes. A better understanding of the mechanistic basis of growth-defense balance has important implications for boosting plant productivity, including insights into how these tradeoffs may be uncoupled for agricultural improvement. |
| [28] | . Exposure of soybean (Glycine max) seedlings to low levels of atmospheric methyl jasmonate induced the expression and accumulation of one or more lipoxygenase(s) in the primary leaves, hypocotyls, epicotyls, and cotyledons. In the primary leaf, the major site of lipoxygenase accumulation in response to methyl jasmonate was in the vacuoles of paraveinal mesophyll cells. In the other organs, however, most of the methyl jasmonate-responsive lipoxygenase(s) were associated with both the epidermal and cortical cells and were present in both vacuoles and plastids. In plastids, the methyl jasmonate-responsive lipoxygenase was sequestered into protein inclusion bodies; no lipoxygenase was evident in either the thylakoids or the stroma. Both spectrophotometric measurement of conjugated diene formation and thin layer chromatography of lipoxygenase product formation indicated that methyl jasmonate caused an increase in the amount of lipoxygenase activity. Electron microscopy of the methyl jasmonate-responsive lipoxygenase protein in the vacuoles showed that it was arranged into a stellate, paracrystalline structure in various cell types other than the paraveinal mesophyll cells. The paracrystals appeared to be composed of tubular elements of between 5 and 8 nm in diameter, were of variable length, and were observed in most cell types of the seedling organs. |
| [29] | . Jasmonic acid (JA) and its derivates, collectively known as jasmonates (JAs), are essential signalling molecules that coordinate the plant response to biotic and abiotic challenges, in addition to several developmental processes. The COI1 F-box and additional SCF modulators have long been known to have a crucial role in the JA-signalling pathway. Downstream JA-dependent transcriptional re-programming is regulated by a cascade of transcription factors and MYC2 plays a major role. Recently, JAZ family proteins have been identified as COI1 targets and repressors of MYC2, defining the 'missing link' in JA signalling. JA090009Ile has been proposed to be the active form of the hormone, and COI1 is an essential component of the receptor complex. These recent discoveries have defined the core JA-signalling pathway as the module COI1/JAZs/MYC2. |
| [30] | . |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}