0 引言
【研究意义】甘蔗(Saccharum spp.)是我国和世界首要糖料作物,并在生物能源领域发挥着愈发重要的作用。作为世界上最高产的作物之一,强大的光合能力是甘蔗的显著生理特征[1-2]。光合作用是植物生物量的来源,对作物产量和品质均有重要影响[3]。因此,高光效育种被视为作物遗传改良的重要内容和研究方向。我国甘蔗育种起步较晚,为拓宽自育品种遗传基础、提高育种效率,陆续引进了大批国外栽培品种。这些材料蕴含丰富的遗传变异,对其开展光合气体交换参数遗传变异特征研究,可为我国甘蔗高光效育种亲本选择和技术优化提供参考依据。【前人研究进展】光合效能是复杂的多因素综合性状,叶绿素荧光[4]、光合放氧[5]、气体交换[6]等生理参数均可反映叶片光合效率。其中,气体交换参数凭借实时无损、直观准确的测量技术优势,在高等植物光合生理研究中应用广泛[6]。甘蔗高光效育种实践中,气体交换参数具有重要作用。首先,气体交换参数可用于甘蔗优异种质资源筛选。罗俊等[7]发现甘蔗光合气体交换参数在不同品种间存在较大差异,其中的3个参数可用于品种光合性能的聚类筛选。徐超华等[8]分析了甘蔗近缘属野生种质资源蔗茅(Erianthus fulvus)的光合气体交换数据,发现材料间差异较大,并从中筛选出了具有较高光合效率和较高水分利用效率的群体。其次,气体交换参数还可用于分析甘蔗光合特性遗传规律。吕建林等[9]发现各项气体交换参数在甘蔗组合间和组合内均存在高度分离,从而证明在甘蔗F1代材料中进行高光效筛选是可行的,并认为净光速率、气孔导度和蒸腾效率等3个气体交换性状的遗传差异主要由非加性效应引起。罗俊等[10]在实生苗中发现甘蔗气体交换性状受组合特殊配合力影响大于亲本一般配合力。【本研究切入点】光合性能的遗传变异是高光效育种的基础。国外甘蔗优良品种的引进和利用,极大丰富了我国甘蔗杂交育种基因库,对我国自育甘蔗品种产量、糖分、抗性的提升均做出了重要贡献。但其在高光效甘蔗新品种培育方面的潜力尚未得到充分利用。要深入挖掘引进品种的高光效育种潜力准确高效的光合气体交换特性分析是十分必要的工作。目前我国对国外引进甘蔗品种的评价多集中在生产特性[11-12]、表型[13-14]和分子遗传多样性[15-16]等方面,研究已从不同角度和水平揭示了国外引进甘蔗栽培品种中蕴含的丰富遗传变异,但这些变异在光合生理、尤其是气体交换特性上的具体体现仍是有待研究的问题。【拟解决的关键问题】本研究随机选取了国家甘蔗种质资源圃内保育的50份国外引进甘蔗栽培品种,对其光合气体交换参数进行了测量和分析,旨在探明其气体交换特性差异,筛选可用于高光效育种的优异种质资源,并为更大规模的高光效资源鉴定和品种选育的技术优化提供依据。1 材料与方法
1.1 供试材料
试验材料为50份国外引进甘蔗栽培品种无性系(表1),从国家甘蔗种质资源圃中随机选取,由国家农作物种质资源共享平台提供。Table 1
表1
表1参试国外引进甘蔗栽培品种信息
Table 1Summary of exotic sugarcane cultivars
| 编号 Code | 材料名称 Clone | 育成国家 Origin | 编号 Code | 材料名称 Clone | 育成国家 Origin | |
|---|---|---|---|---|---|---|
| 1 | Q117 | 澳大利亚 Australia | 26 | Laica80-1729 | 哥斯达黎加 Costa Rica | |
| 2 | Q205 | 澳大利亚 Australia | 27 | C168-75 | 古巴 Cuba | |
| 3 | Q49 | 澳大利亚 Australia | 28 | C22-71 | 古巴 Cuba | |
| 4 | Q70 | 澳大利亚 Australia | 29 | C2858-74 | 古巴 Cuba | |
| 5 | Q71 | 澳大利亚 Australia | 30 | C323-87 | 古巴 Cuba | |
| 6 | Q80 | 澳大利亚 Australia | 31 | Mayari | 古巴 Cuba | |
| 7 | B4362 | 巴巴多斯 Barbados | 32 | MY5465 | 古巴 Cuba | |
| 8 | B45-151 | 巴巴多斯 Barbados | 33 | Mali | 马里 Mali | |
| 9 | B51-410 | 巴巴多斯 Barbados | 34 | M50-47 | 毛里求斯 Mauritius | |
| 10 | BH10-12 | 巴巴多斯 Barbados | 35 | M68-917 | 毛里求斯 Mauritius | |
| 11 | BF162 | 巴基斯坦 Pakistan | 36 | 95H4035 | 美国 USA | |
| 12 | RB73-9735 | 巴西 Brazil | 37 | CP44-154 | 美国 USA | |
| 13 | SP70-1143 | 巴西 Brazil | 38 | CP51-22 | 美国 USA | |
| 14 | FR93-264 | 法国 France | 39 | CP73-1547 | 美国 USA | |
| 15 | FR93-635 | 法国 France | 40 | CP86-1664 | 美国 USA | |
| 16 | FR97-31 | 法国 France | 41 | CP89-1509 | 美国 USA | |
| 17 | FR97-32 | 法国 France | 42 | CP94-1100 | 美国 USA | |
| 18 | FR97-41 | 法国 France | 43 | US67-22 | 美国 USA | |
| 19 | FR97-53 | 法国 France | 44 | Japan3 | 日本 Japan | |
| 20 | CAC113 | 菲律宾 Philippines | 45 | KN90-455 | 苏丹 Sudan | |
| 21 | CAC87 | 菲律宾 Philippines | 46 | Co330 | 印度 India | |
| 22 | Phil72-445 | 菲律宾 Philippines | 47 | Co3311-171 | 印度 India | |
| 23 | VMC71-238 | 菲律宾 Philippines | 48 | Co644 | 印度 India | |
| 24 | VMC87-95 | 菲律宾 Philippines | 49 | Co685 | 印度 India | |
| 25 | VMC96-469 | 菲律宾 Philippines | 50 | POJ2878 | 印度尼西亚 Indonesia |
新窗口打开
1.2 试验设计
试验于云南省农业科学院甘蔗研究所国家甘蔗种质资源圃内进行。试验地位于北纬23.7°,东经103.25°,海拔约1 052 m,属低纬度亚热带高原季风气候,全年以晴天为主,日温差较大,年温差较小。年平均气温20°C左右,光照充足,年日照时数2 385 h,无霜期341 d。试验地土壤为砂质红壤,有机质含量21.2 g·kg-1,pH 7.8,全氮、全磷、全钾含量分别为0.111、0.149、1.200 g·kg-1,其中有效氮、有效磷、有效钾含量分别为67、18.6、166 mg·kg-1。试验整体采用单因素完全随机设计,种植行长3 m,行距1.2 m,株距约0.5 m。于2017年3月初,选取健康完整的中上部单芽茎段下种,种植密度90 000芽/hm2。田间管理措施参照常规甘蔗高产栽培技术实行。试验期间各材料均健康生长,未见病虫害发生。
1.3 测定项目及方法
2017年7月至9月,甘蔗进入大伸长期。每一无性系选取长势具有代表性、均匀一致的植株3株,挂牌标识其+1叶进行测量,共测量5次。采用LI-6400型便携式光合仪(Li-Cor,USA)测定光合气体交换参数。于天气晴朗或少云的上午,待叶片经自然光诱导充分后进行测量,时间为9:30—11:30。以光合仪叶室固定叶片,5—10 min数值稳定后进行记录。光合仪设定参数:气体流量500 μmol·mol-1,CO2浓度400 μmol·mol-1(由小钢瓶供给),叶室温度30°C,光强1 400 μmol·m-2·s-1(使用叶室LED红蓝光源)。
测定光合气体交换参数:净光速率(photosynthesis,A),μmol·m-2·s-1;气孔导度(stomatal conductance,gs),mol·m-2·s-1;胞间CO2浓度(intercellular CO2 concentration,Ci),μmol mol-1;蒸腾速率(transpiration rate,E),mmol·m-2·s-1;叶片固有水分利用效率(intrinsic water use efficiency,WUEintr),μmol·mol-1;瞬时水分利用效率(instantanous water use efficiency,WUEinst),μmol·mol-1。其中,WUEintr=A/gs,WUEinst=A/E。
1.4 数据分析
使用SPSS 19.0软件进行方差分析、相关分析、主成分分析、判别分析。广义遗传力计算参照杜荣骞[17]方法。使用Origin 2017 Pro软件绘制拟合曲线。使用HemI软件[18]绘制热图,并进行聚类分析。原始数据标准化为Z分数进行主成分分析、判别分析、聚类热图绘制。2 结果
2.1 光合气体交换参数遗传差异分析
50份国外引进甘蔗栽培品种光合气体交换参数基因型间差异如表2所示。各材料A平均为27.3 μmol·m-2·s-1,最高的无性系VMC87-95(36.2 μmol·m-2·s-1)与最低的CAC113(15.9 μmol·m-2·s-1)间存在2.27倍差异;gs均值为0.229 mol·m-2·s-1,最高为MY5465(0.367 mol·m-2·s-1),最低为CAC113(0.117 mol·m-2·s-1),二者间差异达3.14倍;Ci均值为144 μmol·mol-1,最高的MY5465(190 μmol·mol-1)与最低的FR93- 264(103 μmol·mol-1)间差异为1.85倍;E均值为3.98 mmol·m-2·s-1,最高为Co330(5.42 mmol·m-2·s-1),最低为CAC113(2.63 mmol·m-2·s-1),差异达2.06倍;WUEintr均值为126 μmol·mol-1,最高的FR93-264(157 μmol·mol-1)与最低的MY5465(95 μmol·mol-1)间存在1.65倍差异;WUEinst均值为6.86 μmol·mol-1,最高为C2858-74(7.66 μmol·mol-1),与最低的CAC113(5.65 μmol·mol-1)间差异为1.36倍。倍数差异和变异系数均显示gs为变异最剧烈的参数,其次为A,再次为E,其后为Ci和WUEintr,WUEinst变异程度最小。方差分析结果显示,所有光合气体交换参数在参试基因型间差异均达到极显著水平(P<0.001)。总体而言,各参数广义遗传力较高,其中最高为A,达74.8%;其后依次为gs、Ci、E、WUEintr,均达70%以上;WUEinst最低,亦达58.8%。
Table 2
表2
表2国外引进甘蔗栽培品种间气体交换参数遗传差异
Table 2Genetic variation in gas exchange parameters among exotic sugarcane cultivars
| 编号 Code | 材料名称 Clone | A (μmol·m-2·s-1) | gs (mol·m-2·s-1) | Ci (μmol·mol-1) | E (mmol·m-2·s-1) | WUEintr (μmol·mol-1) | WUEinst (μmol·mmol-1) |
|---|---|---|---|---|---|---|---|
| 1 | Q117 | 25.9 | 0.218 | 158 | 3.69 | 118 | 6.92 |
| 2 | Q205 | 22.4 | 0.160 | 121 | 3.37 | 145 | 6.63 |
| 3 | Q49 | 31.7 | 0.266 | 146 | 4.30 | 120 | 7.32 |
| 4 | Q70 | 25.7 | 0.230 | 165 | 4.32 | 113 | 5.93 |
| 5 | Q71 | 19.8 | 0.163 | 160 | 3.24 | 123 | 6.25 |
| 6 | Q80 | 20.7 | 0.150 | 125 | 3.31 | 143 | 6.22 |
| 7 | B4362 | 28.9 | 0.212 | 123 | 4.23 | 136 | 6.89 |
| 8 | B45-151 | 34.5 | 0.299 | 134 | 5.06 | 124 | 7.00 |
| 9 | B51-410 | 29.2 | 0.216 | 125 | 4.16 | 135 | 7.11 |
| 10 | BH10-12 | 28.8 | 0.223 | 124 | 4.02 | 137 | 7.30 |
| 11 | BF162 | 24.6 | 0.188 | 132 | 3.84 | 135 | 6.51 |
| 12 | RB73-9735 | 24.8 | 0.243 | 161 | 3.81 | 117 | 6.69 |
| 13 | SP70-1143 | 23.7 | 0.159 | 109 | 3.50 | 151 | 6.86 |
| 14 | FR93-264 | 22.0 | 0.142 | 103 | 3.01 | 157 | 7.36 |
| 15 | FR93-635 | 28.8 | 0.256 | 154 | 4.18 | 118 | 6.89 |
| 16 | FR97-31 | 23.2 | 0.175 | 132 | 3.70 | 137 | 6.52 |
| 17 | FR97-32 | 30.8 | 0.262 | 144 | 4.42 | 121 | 6.91 |
| 续表2 Continued table 2 | |||||||
| 编号 Code | 材料名称 Clone | A (μmol·m-2·s-1) | gs (mol·m-2·s-1) | Ci (μmol·mol-1) | E (mmol·m-2·s-1) | WUEintr (μmol·mol-1) | WUEinst (μmol·mmol-1) |
| 18 | FR97-41 | 29.4 | 0.237 | 141 | 4.03 | 126 | 7.21 |
| 19 | FR97-53 | 34.8 | 0.321 | 157 | 5.04 | 110 | 6.85 |
| 20 | CAC113 | 15.9 | 0.117 | 149 | 2.63 | 133 | 5.72 |
| 21 | CAC87 | 18.7 | 0.148 | 162 | 2.89 | 122 | 6.20 |
| 22 | Phil72-445 | 23.7 | 0.185 | 133 | 3.33 | 136 | 7.17 |
| 23 | VMC71-238 | 25.8 | 0.197 | 138 | 3.59 | 131 | 7.26 |
| 24 | VMC87-95 | 36.2 | 0.343 | 159 | 4.9 | 108 | 7.33 |
| 25 | VMC96-469 | 29.4 | 0.246 | 149 | 3.99 | 120 | 7.31 |
| 26 | Laica80-1729 | 29.7 | 0.282 | 162 | 4.45 | 112 | 6.61 |
| 27 | C168-75 | 29.4 | 0.285 | 173 | 4.82 | 105 | 6.21 |
| 28 | C22-71 | 34.1 | 0.363 | 168 | 4.6 | 104 | 7.31 |
| 29 | C2858-74 | 31.0 | 0.263 | 151 | 4.05 | 118 | 7.59 |
| 30 | C323-87 | 30.1 | 0.232 | 126 | 4.13 | 134 | 7.39 |
| 31 | Mayari | 26.2 | 0.182 | 113 | 3.61 | 146 | 7.34 |
| 32 | MY5465 | 28.8 | 0.367 | 190 | 4.54 | 95 | 6.31 |
| 33 | Mali | 26.1 | 0.184 | 118 | 3.67 | 143 | 7.06 |
| 34 | M50-47 | 26.9 | 0.207 | 134 | 4.08 | 131 | 6.65 |
| 35 | M68-917 | 26.2 | 0.228 | 160 | 3.87 | 116 | 6.72 |
| 36 | 95H4035 | 18.4 | 0.127 | 120 | 2.68 | 149 | 7.01 |
| 37 | CP44-154 | 33.4 | 0.271 | 137 | 4.83 | 123 | 6.95 |
| 38 | CP51-22 | 33.2 | 0.326 | 170 | 4.65 | 103 | 7.07 |
| 39 | CP73-1547 | 31.6 | 0.309 | 162 | 4.52 | 110 | 6.91 |
| 40 | CP86-1664 | 25.8 | 0.232 | 167 | 3.87 | 113 | 6.59 |
| 41 | CP89-1509 | 18.3 | 0.151 | 162 | 2.92 | 123 | 6.33 |
| 42 | CP94-1100 | 30.1 | 0.269 | 159 | 4.74 | 112 | 6.27 |
| 43 | US67-22 | 31.4 | 0.249 | 130 | 4.58 | 129 | 6.96 |
| 44 | Japan3 | 23.9 | 0.166 | 119 | 3.31 | 145 | 7.31 |
| 45 | KN90-455 | 30.5 | 0.274 | 157 | 4.52 | 114 | 6.60 |
| 46 | Co330 | 34.2 | 0.32 | 159 | 5.42 | 108 | 6.29 |
| 47 | Co3311-171 | 22.9 | 0.167 | 125 | 3.14 | 142 | 7.37 |
| 48 | Co644 | 23.3 | 0.186 | 149 | 3.48 | 126 | 6.76 |
| 49 | Co685 | 28.2 | 0.211 | 126 | 3.64 | 136 | 7.59 |
| 50 | POJ2878 | 30.9 | 0.262 | 145 | 4.13 | 121 | 7.36 |
| 均值 Mean | 27.3 | 0.229 | 144 | 3.98 | 126 | 6.86 | |
| 倍数差异 Fold variation | 2.27 | 3.14 | 1.85 | 2.06 | 1.65 | 1.36 | |
| 变异系数 CV (%) | 24.9 | 39.9 | 19.9 | 23.8 | 16.3 | 10.8 | |
| F值 | 19.640*** | 17.338*** | 6.692*** | 17.727*** | 7.329*** | 5.234*** | |
| 广义遗传力 Hb (%) | 74.8 | 70.9 | 70.1 | 72.2 | 72.4 | 58.8 | |
新窗口打开
2.2 光合气体交换参数间相关性
虽然WUEinst为A与E的比值,但其与E的相关系数仅为0.008,相关性不显著。除此以外,其他光合气体交换参数间相关性均达到极显著水平(表3)。除WUEinst外,gs与其他气体交换参数间的相关性均较强。其中,与A、Ci、E呈正相关,与WUEintr呈负相关。这些相关关系均为非线性,使用指数函数和幂函数衍生的超越方程可较好描述其函数关系(图1-a—图1-d)。E与A间同样存在较强的非线性正相关关系,可以幂函数方程y= 167.36x0.12 - 170.16(R2=0.85)拟合(图1-e)。WUEintr与A、gs、Ci、E均呈负相关,其中,与Ci的相关性极强,使用线性方程y= -0.68x + 222.92进行拟合,R2可达0.90(图1-f)。虽然同为水分利用效率,WUEinst与WUEintr相关系数较低,相关性较弱。
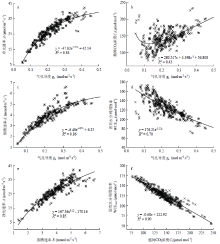 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1气体交换参数间相关关系
-->Fig. 1Relationships between gas exchange parameters
-->
2.3 光合气体交换参数主成分分析
表4表明,以特征根≥1且积累贡献率≥85%为判别条件,共可提取两项主成分因子。PC1特征根达3.459,对总体方差的贡献率高达57.651%;PC2特征根为1.907,所含方差占总方差的31.777%。两项主成分因子共解释了89.427%的总体变异,表明主成分因子提取是有效的,包含了本试验国外引进甘蔗栽培品种光合气体交换数据的主要信息。Table 3
表3
表3光合气体交换参数间相关系数
Table 3Correlation coefficients among photosynthetic gas exchange parameters
| A | gs | Ci | E | WUEintr | WUEinst | |
|---|---|---|---|---|---|---|
| A | 1 | |||||
| gs | 0.865** | 1 | ||||
| Ci | 0.223** | 0.599** | 1 | |||
| E | 0.893** | 0.859** | 0.380** | 1 | ||
| WUEintr | -0.524** | -0.806** | -0.947** | -0.635** | 1 | |
| WUEinst | 0.437** | 0.180** | -0.362** | 0.008 ns | 0.193** | 1 |
新窗口打开
Table 4
表4
表4光合气体交换参数主成分因子得分及载荷
Table 4Score and load of principal component factors in gas exchange parameters
| 参数 Parameter | 得分 Score | 载荷 Load | ||
|---|---|---|---|---|
| PC1 | PC2 | PC1 | PC2 | |
| A | 0.324 | 0.221 | 0.968 | 0.195 |
| gs | 0.274 | -0.01 | 0.956 | -0.209 |
| Ci | 0.041 | -0.425 | 0.439 | -0.839 |
| E | 0.264 | 0.024 | 0.896 | -0.138 |
| WUEintr | -0.141 | 0.305 | -0.700 | 0.679 |
| WUEinst | 0.201 | 0.493 | 0.352 | 0.801 |
| 特征根 Eigenvalue | — | — | 3.459 | 1.907 |
| 方差贡献率 Proportion of variation | — | — | 57.651 | 31.777 |
| 累计贡献率 Cumulative proportion | — | — | 57.651 | 89.427 |
新窗口打开
主成分因子PC1的载荷主要来自A、gs、E,且均为正向(表4)。A值为叶片CO2同化速率的直接体现,gs、E为叶片固碳性能的重要组成部分和表征因子,与A具有极显著的正相关关系(表3,图1-a—图1-e)。可见,PC1因子可称为“碳同化性能”。主成分因子PC2主要由WUEinst和WUEintr提供正向载荷,其负向载荷主要由Ci承担(表4)。Ci与WUEintr间存在极显著的线性负相关关系(表2,图1-f),为WUEintr的负向指征。因此,PC2因子可称为“水分利用效率”。
根据各基因型的主成分因子得分分析因子间相关性,发现PC1与PC2间无相关关系,彼此独立(R2 = 0.02,P = 0.85)(图2)。这为同时具有优异碳同化性能和水分利用效率的优异材料筛选提供了可能。
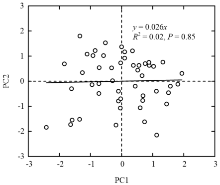 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2主成分因子间关系
-->Fig. 2Relationship between two principal component factors
-->
2.4 聚类与判别分析
R型聚类结果显示,6项气体交换参数中,A、gs、Ci、E等固碳相关参数聚成一组,WUEintr、WUEinst等两项水分利用效率参数聚成一组(图3)。Q型聚类中,50个基因型总体划分为两个群。其中一个群包含I、II 2个组,另一个群由III、IV、V 3个组组成。热图中,两个群特征差异明显,其中一个群固碳相关指标水平较弱,水分利用效率较高,另一个群则相反(图3)。总体而言,聚类结果与主成分因子分析结果较为一致。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3国外引进甘蔗栽培品种气体交换参数聚类热图
-->Fig. 3Hierarchical clustering heatmap based on gas exchange parameters of exotic sugarcane cultivars
-->
以上述聚类结果作为先验概率,使用步进法对Q型聚类结果进行贝叶斯判别分析。经Wilk’s Lambda检验,gs、E、WUEintr、WUEinst等4项参数作为引入变量进入判别模型,其在各组分类判别函数中的系数如表5所示。上述模型判别结果显示分组正判率为100%,说明图3所示聚类结果是可靠的。
各基因型的聚类分组的气体交换特征如表6所示。I组包含7个基因型,A、gs、E、WUEinst均为5组中最低,Ci和WUEintr为中等水平,表明该组材料固碳性能和水分利用高效率均较差,属气体交换性能最差的类型;II组包含11个基因型,其中6项气体交换性状均属中等水平,表明该组材料为固碳性能和水分利用效率较为均衡的类型;III组包含6个基因型,A和E均较低,WUEintr和WUEinst为各组最高,可视为水分利用效率的极端材料;IV组包含11个基因型,具有中等水平的A和较高的gs、E,但其Ci过高,WUEintr和WUEinst均较属最低水平,为气体交换性能较差的类型;V组包含16个基因型,其A、gs和E均为最高水平,Ci和WUEinst较高,WUEintr较低,综合认为该组材料为固碳性能的极端类型,同时具一定水平的水分利用效率。
Table 5
表5
表5各组判别函数系数
Table 5Coefficients in discriminant function of each group
| 引入变量 Introduced variate | 组别 Group | ||||
|---|---|---|---|---|---|
| I | II | III | IV | V | |
| gs | 18.509 | 3.333 | -1.56 | -13.622 | -0.469 |
| E | -19.132 | -2.655 | -0.31 | 12.471 | 1.854 |
| WUEintr | 2.967 | 2.039 | 4.168 | -3.821 | -1.745 |
| WUEinst | -10.427 | -0.815 | 1.485 | 3.901 | 2.009 |
| 常量 Constant | -8.331 | -2.113 | -6.654 | -4.358 | -2.819 |
新窗口打开
II组和V组具有进一步筛选的价值,且所含材料较多,可各自划分出II-a、II-b、V-a、V-b等4个亚组(图3,表6)。其中,II-a亚组包含B4362、B51-410、US67-22、BH10-12、C323-87、Co685等6个基因型,总体表现为A高,Ci低,gs和E适中,WUEintr和WUEinst为4个亚组中最高,同时具有优异的固碳性能和水分利用效率,为本研究中气体交换特征较为理想的一组材料。
Table 6
表6
表6聚类分组气体交换特征
Table 6Gas exchange characteristics of each group
| 组别 Group | 材料份数 Clone Number | A (μmol·m-2·s-1) | gs (mol·m-2·s-1) | Ci (μmol·mol-1) | E (mmol·m-2·s-1) | WUEintr (μmol·mol-1) | WUEinst (μmol·mmol-1) |
|---|---|---|---|---|---|---|---|
| I | 7 | 19.9a | 0.150a | 141bc | 3.12a | 134.3b | 6.32a |
| II | 12 | 27.0c | 0.207b | 131b | 3.90b | 133.6b | 7.05b |
| III | 6 | 23.3b | 0.161a | 116a | 3.24a | 147.0c | 7.24b |
| IV | 11 | 28.3c | 0.268c | 165d | 4.37c | 111.3a | 6.47a |
| V | 14 | 32.1d | 0.287c | 151c | 4.48c | 116.2a | 7.10b |
| 亚组Subgroup | |||||||
| II-a | 6 | 29.4b | 0.224b | 126a | 4.13b | 135c | 7.21b |
| II-b | 6 | 24.6a | 0.190a | 136b | 3.67a | 133c | 6.81a |
| V-a | 9 | 31.1b | 0.262c | 144c | 4.33b | 121b | 7.17b |
| V-b | 5 | 34.0c | 0.332d | 163d | 4.74c | 107a | 7.09ab |
新窗口打开
3 讨论
3.1 国外引进甘蔗栽培品种光合气体交换参数遗传变异
光合作用为作物生长和产量形成提供必要的物质基础。作为主要光合器官,叶片的固碳作用为高等植物积累了近90%的生物量[2-3]。叶片光合能力的遗传改良,已成为作物高光效育种的热点方向之一[3]。本文对50份国外引进甘蔗栽培品种的叶片气体交换参数进行了分析,发现各项参数中存在巨大变异,且广义遗传力较高(表2)。这表明遗传因素,而非环境饰变,是国外引进甘蔗品种叶片气体交换参数变异产生的主要原因。前人利用国外引进栽培品种开展了甘蔗叶片光合效率研究[7,10],也显示了气体交换参数在不同材料间存在显著差异,但未解释其产生的原因。JACKSON[19]和LI等[20]此前的工作表明遗传差异对甘蔗及近缘属植物叶片光合水平差异的形成具有决定性作用,但所用无性系群体由多个种属材料混合而成,遗传背景复杂,天然具有较高的异质性。本研究在遗传背景相对狭窄的国外引进栽培品种无性系群体中,揭示了由其内在遗传差异导致的气体交换参数的巨大变异。原种和近缘属资源的杂交利用可拓宽甘蔗种质基础,但作为高代材料,现有国外引进甘蔗栽培品种亦可为我国甘蔗高光效育种提供较为丰富的遗传资源,种质创新与品种选育实践中应予关注。
3.2 优异基因型筛选及其气体交换特征
优异资源的鉴定与筛选是开展甘蔗高光效品种选育的基础与前提。从气体交换角度来看,光合作用即为一个“以散失自身水分为代价吸收外界CO2”的过程[21]。前人的报道[7-8,22]和本文主成分分析结果(表4)也显示,CO2固定和水分散失是叶片光合气体交换的两大主题。因此,高光效优异材料的筛选,不应一味追求光合速率,而应注重碳同化性能与水分利用效率的平衡。借助聚类分析和判别分析,笔者筛选和鉴定出了可供甘蔗高光效育种优先选择的6份材料,分别为B4362、B51-410、US67-22、BH10-12、C323-87,以及Co685。这一组材料的气体交换特征在于固碳性能优异,同时水分利用效率极佳(表6),在较高水平上取得了二者间平衡。由于碳同化性能与水分利用效率可各自独立在基因型间变化(图2),二者间的平衡是可以达成的。但更多情况下,植物固碳能力和水分利用效率间的矛盾难以调和[23]。本文和其他大量研究[8,19-20]结果表明,光合速率与固有水分利用效率间存在中等以上强度(r>0.5)的显著负相关关系(表2),即为该矛盾的具体体现。本研究中部分材料取得了二者的平衡,应为人工选择压力间接作用的结果。参试材料虽为不同国家的不同育种机构育成,但均由常规选择方法选育,无一为专门的高光效育种计划的产物。甘蔗的常规选育是基于表型的,而部分表型性状,尤其是叶部表型,与其光合能力紧密相关。如叶片厚度可决定单位面积叶片的Rubisco酶含量,从而影响光合速率[1-2];而叶型则与耐旱性有关,叶片长宽比与水分利用效率具有一定正相关关系[2]。相关表型性状的综合考量,可间接完成作物叶片固碳能力和水分利用效率的微妙调和与平衡[24-25]。以上6个优异基因型固碳能力与水分利用效率在较高水平达成平衡,即为表型选择压力间接塑造甘蔗叶片气体交换生理特征的典型体现。
3.3 甘蔗大规模高光效种质资源筛选方案优化
气体交换数据是目前评价叶片光合能力的主流标准[6],但其测量结果易受环境影响。云层遮盖、日光角度、植株自身和邻株的遮光、空气流动等难以避免的扰动,都将通过气孔行为对各气体交换参数数值产生影响[3,22]。广义遗传力可反映环境方差在总体方差中的比例[17],因此可作为气体交换数据质量的衡量指标[20]。为取得更高的广义遗传力,往往需要提高测量强度[17]。这使得气体交换测量成为一项劳动密集工作,对大规模的高光效材料筛选犹是如此。根据全生长期测量结果,LI等[20]预计甘蔗每个基因型约需至少20次测量才能令Hb达到70%。但本研究仅以每个基因型15次(3棵植株×5次测量/株)的测量强度便获得了这一水平的广义遗传力(表2),这可能与选择在大伸长期进行集中测量有关。这一时期,同一品种的各植株长势趋于一致,而不同品种的生长状态差异愈发凸显[1-2],测量结果能够充分体现遗传效应带来的差异,压缩环境方差所占比例。我们结合严格控制仪器测量参数以避免误差项过大,最终以较低的测量强度取得了较好的测量结果。
仅针对气孔导度开展筛选,亦被证明是有效的替代方案。这一方案的优势在于气孔导度直接或间接测量设备价格均远低于光合仪,且测量更为快捷便利。研究机构可负担多台,并以更快的单叶测量速度提高测量通量,从而满足大规模筛选的需求。气孔行为对叶片光合水平的关键影响,是该方案合理性的基础。以本文为例,气孔导度在所有气体交换参数中遗传变异最为强烈(表2),对其他参数具有显著调节作用(图1),同时还是“碳同化性能”主成分因子的主要载荷性状(表4)和聚类结果的判别性状(表5)。在水稻(Oryza sativa)[26]、小麦(Triticum aestivum)[27]、马铃薯(Solanum tuberosum)[28]等作物的生理选育中,针对气孔导度的筛选已得到实际运用。甘蔗中,BASNAYAKE[29]和LUO[30]已明确提出气孔导度可作为耐旱品种选育指标。鉴于适宜的气孔导度亦为本文6份优异材料气体交换的显著特征,笔者认为这一替代方案对甘蔗高光效种质筛选同样具有应用潜力。
4 结论
通过50份国外引进甘蔗栽培品种的叶片气体交换测量和分析发现,净光速率、气孔导度、胞间CO2浓度、蒸腾速率、固有水分利用效率和瞬时水分利用效率等6项参数存在极显著差异,材料间遗传差异是其产生的主要原因。各基因型间,碳同化性能与水分利用效率可以兼顾,B4362、B51-410、US67-22、BH10-12、C323-87、Co685等6份材料碳同化性能和水分利用效率俱佳,可供高光效育种利用。本文研究结果还为甘蔗高光效育种大规模气体交换测量提出了优化方案。致谢:感谢澳大利亚联邦科学与工业研究组织(Commonwealth Scientific and Industrial Research Organization,CSIRO)高级研究员Phillip Jackson博士对本文的帮助。
The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}