随着基因组编辑技术,尤其是规律成簇间隔短回文重复序列/规律成簇间隔短回文重复序列关联蛋白(clustered regularly interspaced short palindromic repeats/CRISPR-associated proteins,CRISPR/Cas)[1-2]技术的出现和广泛应用,利用基因编辑技术对基因组进行精确修饰,并通过显微注射技术、体细胞克隆技术、胚胎移植技术获得基因编辑动物个体或群体已经越来越普遍[3]。这一方法不仅在基因功能研究、疾病模型动物制备等方面有着重要的应用意义,更在畜禽动物新品种培育方面具有重要的经济价值,市场前景十分广阔。利用基因组编辑技术进行育种,需要一系列与传统育种以及转基因育种既有联系又有区别的新理念、新策略和新方法,笔者将这一套策略和方法称之为分子编写育种(breeding by molecular writing, BMW),这一育种方法具有重要的经济价值和市场前景,利用好这一套新的育种方法不仅有助于快速培育畜禽动物新品种,而且会改变现有的育种方式,并将对世界范围内的畜禽养殖业产生重要的影响。
本文将详细论述常规动物育种技术的现状和难点以及分子编写育种方法的基本概念、研究手段、研究内容、研究现状并展望其发展前景和对育种工作的影响,为动物育种、畜禽养殖以及相关政策制定提供参考。
1 动物育种的现状与难点
培育动物新品种的主要方式是杂交育种和选择育种。以猪的育种为例,所有的现代种猪几乎都是通过不同品种的猪杂交并长期选育而成的[4]。《英国大百科全书》记载“现在欧洲的猪种,是当地猪种与中国猪杂交而成”。比如著名的约克夏猪(Yorkshire)和巴克夏猪(Berkshire)都是将广东猪种和英国当地猪种杂交,并经过长期选育而形成的。美国的切斯特白猪(Chester White)、波中猪(Poland China)则是在1817年引进华南猪杂交改良并长期选育而成。杂交育种为动物养殖业的发展做出了巨大的贡献。中国从20世纪50年代开始,开展了广泛的“土洋”猪杂交育种。通过地方品种与引进品种的杂交,把引进品种的瘦肉率高、日增重快、饲料转化率好的特点与我国地方品种的繁殖力高、肉质好、适应性强的特点结合在后代中,再通过连续不断的选择,培育出大量优质、高效的生猪新品系和配套系。而且到目前为止,杂交技术也是一项在常规的商品猪生产中广泛使用的技术,用于生产各种经济性状均较为理想的杂种商品猪,如杜长大三元杂交猪等。
但是杂交育种存在明显的不足。一是不同品种猪杂交后,后代在引入优质基因的同时,也引入了大量劣性基因,为了选育出优良的杂交后代,必须配备大量的人力并长期进行观察和测定。这种选育过程要不断持续几十年之久。如今风靡世界的长白猪、大白猪等著名品种,都是经过将近百年的选育才培育成功。漫长的选育过程导致动物新品种培育的人力成本、时间成本和经济成本居高不下。
二是常规的选育方法无法对那些难以观察和测定的复杂性状进行选育。比如抗病性状在不发病时无法观察,而在持续攻毒的条件下进行测定和选育又会对养殖场造成巨大的感染风险。另外测定成本较高的饲料转化率性状、肉质性状以及低遗传力的繁殖力等性状,用常规育种方法进行选育不仅成本高、可操作性差,而且选育进展也比较缓慢。随着环保要求以及消费水平的不断提升,规模化养殖和优良肉质品种将是未来动物养殖业的普遍需求,这就要求动物新品种的培育要向高抗病能力、优良肉质等方向发展,而这两方面恰恰是常规选育方法无法发挥其长处的地方。
三是杂交育种本身的规律就决定了,即便经过漫长的选育,劣性基因也不一定能完全从杂交后代中排除。根据基因的连锁和交换定律,染色体上相距很近的两个基因常常是连锁在一起的,作为一个单元进行遗传。所以不管如何延长选育时间,那些与优质基因连锁的劣性基因依然会存在。
2 分子编写育种的内容及研究现状
分子编写育种是指利用锌指核酸酶(zinc-finger nucleases, ZFN)[5-6]、转录激活因子样效应因子核酸酶(transcription activator-like effector nucleases, TALEN)[7-8]]或者CRISPR/Cas等基因组编辑工具对生物基因组进行精确到分子水平的编写并定向培养新品种的技术。该技术最大的特点在于以“高精度”和“高效性”实现“高安全性”。可以对任何物种基因组的任何位置进行精确到单个碱基的编写,实现遗传序列的精确插入、替换、复制、删除等操作,大大降低非预期效应。分子编写育种的目的在于:通过基因组编辑技术、显微注射技术以及体细胞克隆技术,高效创制新的遗传标记并快速验证,对基因组进行精确的碱基序列插入、删除或者替换等操作,实现物种内或者跨物种的遗传信息的高效、快速整合,进而快速获得目标性状突出的生物个体或种群,高效培育具有目的性状的新品种,解决传统育种中耗时长、成本高、效率低的问题,实现真正意义上的“分子杂交”和“分子育种”。
分子编写的研究内容主要包括:新型育种标记的创制及验证、跨物种分子编写、基因组中碱基序列的删除以及物种内分子编写等四项(图1)。
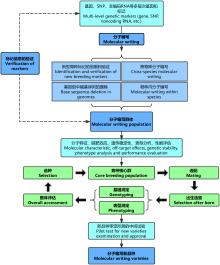 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1分子编写的研究内容及育种流程
-->Fig. 1Research contents and operational process of breeding by molecular writing
-->
2.1 新型育种标记的创制及验证
遗传标记在育种中发挥着重要作用,但是目前已发现的有效的遗传标记如主效基因、因果SNP位点等数量有限,非常不利于育种产业的发展。分子编写技术可以有效的解决这一问题。首先,分子编写技术可以高通量、大规模的创制新型的遗传标记。比如创制猪的新型遗传标记可以先构建靶向猪全基因组中启动子、内含子以及microRNA簇、lncRNA、DNA甲基化区域等重要转录调控区域的gRNA文库,并利用CRISPR蛋白对这些区域进行随机或者连续编辑,规模化的产生单碱基或大片段突变,然后进行初步的表型筛选,就能获得大量与目标性状相关的候选遗传突变和遗传标记。
另一方面,也可以综合已有的不同品种猪的基因组数据、SNP数据、全基因组关联分析结果等生物信息学数据和文献,筛选与猪抗病、生长速度、瘦肉率、产仔数、肉质等重要经济性状因果相关或紧密相关的各种遗传标记(如SNP位点、酶切位点、结构变异、拷贝数变异等)、表观遗传标记(如microRNA、lncRNA、DNA甲基化修饰等)。
以上两种方式获得的遗传标记,可以进一步通过体细胞克隆的方法进行精准验证。例如验证SNP位点与目的性状的相关性(图2),就可以先利用分子改编技术的高精准性从同一个细胞系构建出多个SNP差异细胞系,然后再利用体细胞克隆动物细胞核遗传一致性,构建SNP差异群体。这个群体理论上具有完全一致的遗传背景,只在目标SNP位点存在单碱基差异。进一步分析SNP差异群体的表型,就能验证SNP位点与目标性状的相关性,及其对目标性状的贡献度。这种方法避免了全基因组选育中依赖算法造成的统计学误差,能更准确地验证SNP位点对目标性状的贡献度,从而获得因果SNP位点。
构建的SNP差异群体除了可以精准验证SNP对目标性状的贡献外,还可深入研究SNP影响目标性状分子机制,以及挖掘与目标性状相关的其他遗传标记的好材料。比如在同一品种、同一来源的细胞中实现单核苷酸的“转换”或“颠换”培育的SNP差异群体,它们在遗传背景上基本完全一致,这时不仅可以直接验证SNP对目标性状的贡献度,还可以通过高通量测序、功能基因组学及表观遗传学深入分析其分子机制。
而在不同品种间的细胞中分别实现单核苷酸的“转换”或“颠换”培育的SNP差异群体,其SNP变异相同,但遗传背景不同,这时就可以深入挖掘、筛查与目标性状相关的其他遗传标记(图2)。
综上所述,利用分子编写技术不仅能规模化的精准创制遗传突变,便于发现新型遗传标记,更可以精准验证SNP位点对目标性状的贡献度,改变目前全基因组选育中完全依赖算法造成的统计学误差,从而获得一批具有高度育种价值的因果SNP位点,形成自主知识产权及核心技术。另外在进行SNP验证时,还可以进一步解析SNP位点影响目标性状的分子机制,挖掘和筛查与目标性状相关的其他遗传标记。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2通过分子编写技术进行SNP位点的规模化验证
-->Fig. 2SNPs validation using breeding by molecular writing
-->
2.2 跨物种分子编写育种
跨物种分子编写育种,即通过ZFN、TALEN或者CRISPR/Cas系统以及显微注射技术、体细胞核移植技术将一个物种中的基因精确插入或替换到另外一个物种基因组的特定位置中,或者在一个物种的基因组特定位置上,模仿另一个物种基因组中特定位置的碱基序列信息。常见的是将外源基因插入或替换到某一物种基因组上的特定位点中,如友好基因座中。该方法不仅能打破物种间的生殖隔离,引入外物种的优良基因,而且能在大大降低非预期效应的同时,充分发挥外源基因的育种价值。现有的跨物种分子编写育种研究可以分为以下三类:外物种基因的精确插入,外物种基因的精确替换以及外物种突变序列的模仿。2.2.1 外物种基因的精确插入 外物种基因的精确插入目前已有报道。其主要方法是将具有特殊功能的外源基因的完整表达框,精确的定点整合到受体动物基因组中的特定位置中。使其整合位置以及表达模式完全可控,避免传统转基因技术中的非预期效应的产生。目前常选择动物基因组中的“友好基因座(safe harbor)”进行定点整合。友好基因座是指动物基因组中转录活跃且不影响动物生长、发育和生殖的基因位点,这些位点非常适合外源基因的表达,对外源基因是友好的,因此称为友好基因座。在开发和利用友好基因座进行外物种基因的精确插入方面,我国的科研工作者走在了世界前列。中国科学院广州生物医药与健康研究院鉴定出猪的Rosa26友好基因座,并在该位点实现外源基因的定点插入和替换[9]。
中国农业科学院北京畜牧兽医研究所鉴定出猪的一个新型友好基因座H11位点,并利用CRISPR/Cas技术高效、准确的在该位点定点插入长达9.4 kb的外源基因片段[10]。同时,该团队还进一步制备了两种在该友好位点分别插入三个基因的II型糖尿病模型猪:转GIPRdn-hIAPP-11β-HSD1巴马猪和转GIPRdn-hIAPP-PNPLA3 I148M巴马猪,该项研究首次在国际上创制胰岛β细胞和肝脏特异性表达多基因小型猪2型糖尿病模型,并率先在国际上利用基因组编辑技术创制定点敲入大片段基因工程猪(数据暂未发表)。
西北农林科技大学利用CRISPR/Cas9n切口酶系统将结核抗性基因NRAMP1定点整合到奶牛基因组的友好基因座F-A位点中,不仅建立了一套高效、低脱靶率的将外物种基因精确插入奶牛基因组的技术体系[11],而且有望利用此成果培育出抗牛结核病的新型奶牛品种,市场潜力巨大。
2.2.2 外物种基因的精确替换 在外物种基因的精确替换方面,美国密苏里大学通过CRISPR/Cas技术将猪CD163基因的第7外显子精确替换为人的同源序列[12]。猪CD163蛋白是巨噬细胞表面的一种蛋白,在猪繁殖与呼吸综合征病毒(porcine reproductive and respiratory syndrome virus,PRRSV)感染过程中能介导PRRSV病毒进入巨噬细胞,而人的同源蛋白却不介导PRRSV的感染,从而使得PRRSV病毒无法感染人类。该研究精确替换了CD163基因的第7外显子,不仅能深入探索PRRSV病毒感染机制,更有望在保留猪CD163蛋白基本功能的前提下,删除CD163对PRRSV病毒的介导作用,从而培育出既能抵抗PRRSV病毒,又不会因CD163敲除而可能携带某些缺陷的新型抗病猪。
2.2.3 外物种突变序列的模仿 除了外物种基因的精确插入和替换外,还可以通过精细的模仿外物种碱基突变培育动物新品种。这一方面我国的研究也处于世界前列。肌肉生成抑制素(myostatin, MSTN),也称为生长分化因子8(growth and differentiation factor 8, GDF8)是一种肌肉生长的负性调控因子。世界著名的比利时蓝牛,正是由于MSTN基因第三外显子上11个碱基的缺失,形成了提前终止密码子,造成了其“双肌臀”表型[13]。比利时蓝牛肌肉量特别丰富,屠宰率最高达71%,比其他品种牛多提供肌肉18%—20%。中国农业科学院北京畜牧兽医研究所使用ZFN技术和TALEN技术在大白猪体内模拟比利时蓝牛MSTN基因的自然突变,获得“仿比利时蓝牛MSTN基因突变”的大白猪。这种“仿比利时蓝牛MSTN基因突变猪”经过精确的基因编辑,所携带的MSTN基因突变类型与比利时蓝牛的自然突变完全一致,从而在提高肌肉量、降低脂肪沉积、提高生长速度的同时进一步降低了MSTN基因突变可能引起的不良效应,具有重要的应用价值和良好的商业前景。目前,该基因编辑新品种猪已经顺利完成中间试验(数据暂未发表)。
在抗病育种方面,英国罗斯林研究所研究团队发现:非洲疣猪通常会携带非洲猪瘟病毒,却不发病,而家猪在感染非洲猪瘟病毒后,在很短的时间内就会发病倒下。基因组测序分析发现,与感染非洲猪瘟病毒相关的一个关键基因RELA在家猪和非洲疣猪中存在3个不同的SNP位点,这可能是家猪易感猪瘟病毒的一个重要遗传差异。非洲疣猪与家猪同属猪科,但是属于不同的亚科。家猪属于猪亚科,非洲疣猪属于疣猪亚科。该团队利用ZFN技术,成功地将家猪RELA基因的3个SNP更改为非洲疣猪的SNP,有希望获得天然抵抗非洲猪瘟病毒的基因编辑猪[14]。
2.3 基因组中碱基序列的删除
分子编写育种的第二方面是:基因组中碱基序列的删除。除了打破生殖隔离,跨物种的引入基因或碱基突变外,分子编写育种还包括在同一物种内对基因组进行精细操作,精确删除基因组上的某些无用甚至有害的碱基序列。比如在进化中整合到基因组上的内源性逆转录病毒、导致应激综合征的基因、疾病易感基因以及某些病毒或细菌的受体基因等。将这些基因或序列精确的剔除掉,就能定向培育出更为优质,更符合人类养殖、食用、医药研发等多种不同需求的生物新品种。2.3.1 内源性冗余序列的删除 猪的内源性逆转录病毒是在长期进化过程中整合到猪基因组中的,对猪本身已经没有毒性,但是在进行异种器官移植时却有可能对人体安全造成威胁。通过分子编写,美国哈佛大学的杨璐菡团队成功删除了猪基因组中几乎全部的逆转录病毒序列,并通过体细胞克隆技术获得了世界上首批适合进行异种器官移植的无内源性逆转录病毒的小型猪[15-16],该研究对培育异种器官移植用小型猪新品种具有重大意义。同样,通过分子编写技术,删除其他物种基因组中的内源性病毒序列,也能够培育出更安全,适用于医学研究和应用的生物新品种。
2.3.2 负作用基因和因子的删除 除删除内源性逆转录病毒外,通过分子编写技术删除基因组中某些负调控基因和因子,同样可实现相应性状的快速改良。如猪、牛、羊基因组中的MSTN基因对肌肉生长具有抑制作用,删除该基因可提高动物的产肉量。早在2012年,中国农业科学院北京畜牧兽医研究所就通过ZFN技术获得了MSTN基因敲除梅山猪,其瘦肉率显著提高。至今已繁育到第五代,并且已获准开展转基因生物安全评价环境释放试验。2014年,该单位又利用TALEN技术精确模拟比利时蓝牛的MSTN基因突变,获得了“仿比利时蓝牛MSTN基因突变”的大白猪初代群。 2015年,中国科学家与韩国科学家合作利用TALEN技术获得具有显著“双肌臀”表型的MSTN基因编辑猪[17]。之后,吉林大学[18]和湖北省农业科学院畜牧兽医研究所[19]又分别利用CRISPR/Cas9技术成功制备了MSTN基因编辑猪。除MSTN基因敲除新品种猪外,利用CRISPR/Cas9技术还可获得MSTN基因敲除绵羊、山羊、兔等高产肉率新品种动物。
牛奶和羊奶中的β-乳球蛋白是人乳中不含有的蛋白,是奶制品中的过敏原之一。通过分子编写技术将牛或羊基因组中的β-乳球蛋白基因敲除[20],从而可降低牛羊乳中过敏原的含量,培育出不含β-乳球蛋白的牛羊新品种,缓解部分人群牛乳过敏症状。2011年,中国农业大学利用ZFN技术成功敲除牛基因组中的β-乳球蛋白基因,获得β-乳球蛋白敲除的奶牛,为培育低过敏原奶牛打下了良好的基础。
2.3.3 与疾病易感相关基因和因子的删除 抗病能力是动物育种中具有重要经济价值的一种表型。但是传统育种手段在选育抗病品种时缺乏合适的指示性状,而攻毒后再测定则成本高昂,并会对生产有不利影响。分子标记辅助育种存在分子标记较少,各个抗病分子标记间的关系复杂,抗病性状与部分生产性状存在拮抗等多种问题。因此抗病动物品种的培育工作一直进展缓慢。而分子编写育种则可以通过直接删除疾病易感基因或者病毒受体基因,快速培育具有抗病性状的动物新品种,将大大加快抗病动物新品种的培育速度。
动物传染性海绵状脑病(transmissible spongiform encephalopathy, TSE)是一种可以感染人类和牛、羊等动物的致死性中枢神经系统疾病。这种疾病不仅对牛、羊等动物养殖业造成巨大损失,还直接威胁着人类健康。通过动物分子编写技术,敲除动物中引起TSE疾病的PRNP基因,就能获得抗TSE的动物。如敲除PRNP基因的抗疯牛病牛[21],和抗羊瘙痒病绵羊[22]、山羊[23-24]等动物新品种。这类抗TSE动物新品种的进一步推广和应用,将会大大提升畜牧业生产和人类健康的安全保障。
猪繁殖与呼吸综合征又称为猪蓝耳病,是严重威胁猪养殖业的头号烈性传染病。该病由一种小RNA病毒引起,变异速度快,防控难度大。从2014年开始,美国密苏里大学研究团队利用基因组编辑技术先后敲除了与PRRSV病毒感染相关的CD169和CD163基因,获得了CD169和CD163基因敲除猪。攻毒实验发现CD163双等位基因敲除猪对PRRSV具有完全抗性[25-28],就是说CD163基因敲除猪能够完全抵抗猪蓝耳病。国际著名猪育种公司Genus plc(www.genusplc.com)已经与密苏里大学合作,开展抗蓝耳病新品种猪的培育工作,该公司在2015年宣布计划用5年时间将抗蓝耳病新品种猪提供给猪肉生产商。截止到2017年12月,该公司已相继拿到CD163基因、分子剪刀等知识产权许可,并培育出祖代抗蓝耳病猪群体,整体培育工作进展顺利(图3)。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3Genus pIc公司抗蓝耳病猪开发时间表(数据源自Genus pIc公司网站)
-->Fig. 3The R&D timeline of Genus pIc’s PRRSv resistance programme(Cited form Genus pIc website)
-->
2.4 物种内分子编写育种
分子编写育种的第三方面是:物种内分子编写育种。常规的杂交育种手段无法精确的将“好基因”和“坏基因”分开,因此往往在导入优质基因的同时也导入了大量的劣性基因。尤其是那些互相连锁的基因,常规的杂交育种手段是无法分开的。物种内分子编写育种就能解决这一问题。分子编写育种可以在同一物种内对动物基因组进行精细操作,模仿同一物种中其他品种上的碱基突变或SNP位点,将多个品种的优质遗传突变信息,如基因突变以及启动子、增强子、内含子等多种非编码区域上的SNP位点、突变序列等模仿和替换到一个品种上,实现真正意义上的“分子杂交”。比如在猪的育种中,分子编写育种可以精确的将地方猪的肉质优良、抗逆性强的基因替换到商用品种中,或者将商用品种生长速度快、饲料利用率高、瘦肉率高的基因替换到地方品种中。甚至可以精确的实现品种间、个体间碱基突变或SNP位点的模仿,直接获得分子杂种优势,替代有性杂交,实现真正的猪个体和群体水平的“分子杂交”育种。
通过对“瘦肉型”和“肥肉型”猪的QTL分析发现,“瘦肉型”猪(如大白猪、皮特蓝猪)与“肥肉型”猪(如野猪、梅山猪)相比,其类胰岛素生长因子2(Insulin-like growth factor 2, IGF2)的第三内含子的第3072位碱基存在G到A的SNP位点,该SNP位点对猪的生长速度、瘦肉率、背膘厚、背最长肌面积以及心脏重量等性状有着非常显著的影响,可以使猪的肌肉量增加15%—30%,背膘厚减少10%—20%[29]。该SNP位点应该是在“瘦肉型”猪的培育过程中自然突变后经过漫长的人工选育固定下来的。如果通过物种内分子编写育种,将该SNP位点直接精确的引入到我国地方品种猪中,就能快速的提高我国地方品种猪的产肉性能,对我国地方品种猪的改良意义重大。
在欧洲和美国的奶牛养殖业中,常常出于生产安全以及对工作人员的保护将奶牛的角去掉。但是去角的过程不仅增加了养殖成本,对奶牛来说也非常痛苦,与动物福利和动物保护的宗旨不符[30]。目前已选育出某些肉用品种的牛有无角性状,如安格斯肉牛(Angus cattle) [31],其无角性状主要由1号染色体上的POLLED基因决定[32]。如果通过杂交育种的方法培育无角奶牛,不仅需要非常长的选育时间,而且杂交后代的产奶性能一定会受到肉牛某些基因的影响,产奶性能会降低。TAN等利用TALEN技术成功将肉牛的无角基因POLLED定点整合到有角奶牛的基因组中,由此可培育出既保持高生产性能,又无角的奶牛新品种,免除了奶牛切角的痛苦[33]。
禽流感病毒(avian influenza virus,AIV)极大的威胁着禽类养殖业和人类健康。培育能抵抗禽流感的禽类动物,一直以来都是养殖和育种从业人员的梦想。2011年,英国罗斯林研究所通过转入干扰RNA来抑制禽流感病毒的增殖[34],获得了较好的抗病效果。未来则可以借助分子编写技术,将其他品种中对禽流感病毒有抵抗力的基因[35]直接模仿和替换到生产性能较高的品种上,进而快速获得对禽流感病毒具有一定抵抗力的新品种动物。
2015年,一种能快速生长的转基因三文鱼——水优三文鱼(aquAdvantage salmon)通过美国食品药品监督管理局(food and drug administration, FDA)的审批,获准上市销售[36-37]。2017年,首批水优三文鱼已经正式上市销售[38]。这种三文鱼本身为大西洋三文鱼(atlantic salmon),转入了另外一种三文鱼——大鳞三文鱼(chinook salmon)的一个生长激素调节基因,以及一个来自美洲大绵鳚(ocean pout)的启动子,用来调节生长激素的合成,从而使得它们能够全年生长,因此生长速度比普通三文鱼快一倍,节省约75%的饲料。该品种采用的是传统的转基因技术,因此经历了较为漫长的审批过程。如果采用品种内分子编写技术,将其他品种三文鱼的优质基因或启动子序列替换到大西洋三文鱼中,相信将更加容易获得消费者的认可。
3 分子编写育种流程
分子编写育种的目的是改良品种并将优良的遗传基因传递至商品代。因此除了上述的对动物基因组的分子操作外,分子编写育种还需要紧密结合目前广泛使用的常规育种方法,持续对分子编写群体进行选配、测定、评估,根据评估结果选择优良个体建立核心群(图1)。然后依次建立纯种扩繁群商品生产群,将优良基因传递给商品群。在核心群的建立和维持过程中,测定和评估是关键环节。不仅要进行性能测定、体型评估等常规测定,还需要进行基因测定,获取其遗传标记信息,以便通过全基因组选择和测序分型进行更准确的选种。也就是说,通过分子编写技术获得的、遗传性状得到“质变”的群体,还需要结合全基因组选择、标记辅助选择等多种育种方法进一步不断选育、不断迭代、不断积累“量变”,才能保持遗传性状的优良。4 分子编写育种的前景
综上所述,分子编写育种一方面可以在物种间实现优质基因或序列甚至是SNP的流动、替换,打破物种间的生殖隔离,充分发挥优质基因的育种价值。另一方面可以在物种内真正实现个体和群体水平的分子杂交育种,快速获得具有分子杂种优势的生物新品种。跨物种分子编写育种与转基因技术相比,精确性大大提高,因此可以大大降低外源基因随机整合以及表达量不可控导致的非预期效应,更加精准,更加高效,更加安全。而物种内的动物分子编写育种,在真正实现分子杂交的同时,完全不含其他物种的DNA序列,在本质上与传统育种方法获得的个体完全没有差异,可以说等同于传统育种方法获得的遗传变异个体。尽管分子编写育种具有诸多优势,但目前尚有两方面的问题需要进一步解决。一是基因编辑技术的准确性及特异性。分子编写技术所依赖的基因组操作工具——ZFN、TALEN和CRISPR/Cas等技术发展迅速,其精确性和特异性都在不断提高。如通过Scr7抑制小鼠细胞中的非同源末端连接修复(non-homologous end joining, NHEJ)可将精确修复的效率提高数倍甚至数十倍[39-40],除抑制NHEJ修复外,还可以通过直接增强同源重组修复(homology directed repair, HDR)以提高精确修复的比率,主要方法有调控细胞周期[39, 41]、引入外源性DNA[42]、对sgRNA进行化学修饰[43]或利用小分子化合物激活HDR[44]等。除了以上几种增强精确修复的方法,还可以利用胞嘧啶核苷脱氨酶[45]或碱基修饰酶[46]与Cas9蛋白直接融合,进而实现对单个胞嘧啶到胸腺嘧啶的直接转换。
除不断提高CRISPR/Cas技术的精确修复能力外,CRISPR/Cas技术的特异性也已经得到极大的改善。主要优化方法可分为对Cas9蛋白的改造和针对sgRNA的改进。在Cas9蛋白的改造方面,其主要策略包括将Cas9蛋白的核酸内切酶失活,只保留DNA识别能力,同时与其他核酸内切酶融合形成复合体,如FokⅠ-dCas9复合体[47-48],进而提高对靶序列切割的特异性。其次是对Cas9蛋白进行点突变,筛选与DNA结合特异性更高的突变体,如增强型eSpCas9[49]、高保真型Cas9-HF1[50]等。针对sgRNA的改造是改变sgRNA的长度,研究表明在sgRNA的5'端的减少2—3个碱基[51]或在识别序列前加2个鸟嘌呤[52],能够在不影响切割活性的情况下,降低脱靶率大约5 000倍。
第二方面是政府监管措施及公众接受度。由于基因编辑技术发展的速度太快,远远超过了人们对其了解和熟悉的速度,不仅科学界对基因编辑作物是否属于转基因生物(genetically modified organism, GMO)尚未有明确的定论,而且全球多数国家的监管标准也尚未明确。2016年《Nature genetics》发表评述文章[53],提出了5项基因编辑作物的管理框架,该框架对基因编辑作物的管理提出了系统性的建议,在最大限度减少基因编辑作物传播风险的同时,建议“以和常规育种作物相同的制度来管理基因组编辑作物”从而简化基因编辑作物的管理,以加快基因编辑作物的商业化应用。目前,美国农业部对于通过基因编辑工具生产的、不含有外源基因的食品免于监管。如2016年美国农业部就宣布免除对CRISPR/Cas9技术编辑的抗褐变蘑菇的监管[5]。而转基因动物在美国是作为药物受FDA监管,且FDA声明基因组编辑动物依然要受其监管。而澳大利亚、新西兰和阿根廷等国家对不含有外源基因的产品免于监管,对于有少数DNA碱基插入的产品实行个案分析原则,对于有大片段插入的食品则进行严格监管。
2018年中国科学院遗传与发育生物学研究所高彩霞研究员发表评述文章[55],认为政府监管措施和公众接受度是影响基因编辑技术农业应用的最大因素,全球监管环境的不确定性限制了将该技术进行应用的信心。同时她认为目前基因编辑技术所取得的成就只是冰山一角,这种新型育种技术将为农业带来可持续发展的未来。因此,我们有理由相信,随着基因编辑工具的不断更新,分子编写育种也将不断的发展与成熟,其安全性和优越性将进一步被消费者认识和熟知。未来的分子编写品种和产品将会越来越多,动物育种将迎来全新的发展机遇,为人类提供更加优质、健康的产品。
The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}