0 引言
【研究意义】万寿菊为菊科万寿菊属植物,万寿菊是提取类胡萝卜色素的优质植物源材料[1],其中叶黄素是主要色素成分,约占万寿菊所含类胡萝卜素总量的90%[2]。番茄红素β-环化酶是类胡萝卜素合成途径中的关键酶,是调控番茄红素向叶黄素分化的限速酶,因此克隆并探索能够驱动万寿菊番茄红素β-环化酶高效转录的番茄红素β-环化酶启动子,对于提高万寿菊体内叶黄素含量的研究有重大意义。【前人研究进展】类胡萝卜素是由异戊二烯途径合成的一类自然界广泛存在的有色物质,呈现橙红色、橙色和黄色的一类萜类物质[3],约有10%的类胡萝卜素是维生素A的前体,具有抗癌、预防癌症等多种疾病的作用,是人和动物食物中不可缺少的成分[4,5,6,7]。目前有关植物类胡萝卜素生物合成途径的研究很多,参与其代谢的酶主要有八氢番茄红素合成酶(PSY)、八氢番茄红素脱氢酶(PDS)、ξ-胡萝卜素脱氢酶(ZDS)、番茄红素β-环化酶(LCYb)和番茄红素ε-环化酶(LCYe)等。其中番茄红素β-环化酶(LCYb)是类胡萝卜素合成途径的关键酶,其基因的转录水平会影响上游番茄红素和下游β-胡萝卜素、叶黄素等产物的积累。MORENO等[8]发现超量表达DcLCYb1的胡萝卜株系中能显著提高β-胡萝卜素的积累量,沉默DcLCYb1时会降低β-胡萝卜素的含量;红肉番木瓜是由于野生型番木瓜有色体中特有的番茄红素β-环化酶CYCB基因功能缺失导致番茄红素的积累产生的[9,10,11]。从番茄品种中克隆得到两种番茄红素β-环化酶基因,分别为LCYB和CYCB,研究发现红色果肉番茄与浅色果实的番茄相比,两个基因的表达量逐渐降低随着果实的成熟,但红色果肉上游番茄红素积累量显著高于浅色果实的番茄[12]。柑橘中LCYB表达量下调会导致番茄红素的积累[13,14,15],将IbLCY-β-RANi干涉载体导入到白心红薯植株体中,发现转基因植株的红薯内部变成了橘红色[16]。因此,番茄红素环化酶在参与类胡萝卜素组分调节中起着重要的作用。前人通过同源克隆的方法已经获得了万寿菊舌状花(花瓣)中类胡萝卜素生物合成途径中各种关键酶基因,叶黄素的合成需要多种酶的催化,在万寿菊中叶黄素的合成积累与LCYe、LCYb有密切关系[1, 17]。【本研究切入点】本研究利用RT-PCR的方法从万寿菊花中克隆获得一条LCYb同源基因,命名为TeLCYb,前人研究表明万寿菊中叶黄素合成可能与LCYB同源基因的调控机制直接相关[18],而关于万寿菊TeLCYb的启动子研究未见报道。【拟解决的关键问题】本研究在TeLCYb序列的基础上利用FPNI-PCR法克隆TeLCYb启动子序列,并利用在线软件预测该基因启动子区域的顺式作用调控元件,构建启动子缺失表达载体,异源转化模式植物烟草,通过GUS化学组织染色明确TeLCYb的表达部位和预测启动子顺式作用元件功能,为进一步研究TeLCYb的转录调控奠定基础。1 材料与方法
试验于2014年7月至2016年12在华中农业大学进行。1.1 材料
2014年7月中旬将万寿菊‘贵妇人’(色素品种)播种于128孔穴盘中,待幼苗长出2—3对真叶时移栽于直径为21 cm的素烧瓦盆,并摆放于基地露天广场(位于北纬30°28′36.5″,东经114°21′59.4″),常规栽培。待苗长到5—6对真叶时,采集健康的幼叶留作DNA的提取,九月下旬采集全开时期的整朵花于液氮中速冻并保存于-80℃超低温冰箱备用。转化所用烟草为导入了外源玫瑰FT的品种,表现为比正常烟草早开花的性状[19]。
1.2 试验方法
1.2.1 万寿菊番茄红素β-环化酶基因的克隆 利用Trizol试剂盒法(Invitrogen,California,The United States)提取万寿菊全开时期的花器官总RNA,利用反转录试剂盒 PrimerScript® RT Reagent Kit with gDNA Eraser(TaKaRa,大连,中国)将RNA反转录成单链cDNA,并用万寿菊的看家基因β-actin检测反转录效果。从笔者课题组测定的万寿菊转录组数据库(序列号:SRP066084)中调取TeLCYb序列,根据序列的UTR区设计TeLCYb的上下游引物TeLCYb-F/ -R(表1)。PCR反应程序为:94℃预变性4 min;94℃变性30 s,60℃退火30 s,72℃延伸2 min,36个循环;总延伸10 min。PCR产物经1%琼脂糖凝胶电泳检测后对目的片段在紫外线下进行切胶回收,将纯化后的产物与PMD-18T载体连接,再经过热激转入大肠杆菌DH5α感受态细胞中,菌落PCR检测后将阳性克隆送生工生物工程(上海)股份有限公司测序。利用DNAMAN和在线软件BoxShade(https:// embnet.vital-it.ch/software/BOX_form.html)分析万寿菊TeLCYb的生物学信息。
1.2.2 番茄红素β-环化酶基因启动子序列克隆和其生物信息学信息分析 以幼叶为材料,采用改良的CTAB法[20]提取万寿菊总DNA。根据上述TeLCYb序列测序结果,利用FPNI-PCR法克隆TeLCYb启动子,设计3个嵌套式的特异引物SPb1-1-R、SPb1-2-R、SPb1-3-R(表1),3轮反应程序具体参照WANG等[21]方法,在第三轮中发现一条特异条带,经切胶回收、热激连接T载体,阳性菌落检测,送样测序。采用同样的FPNI-PCR法,以第一次启动子测序结果为模板设计3个嵌套式的特异引物SPb2-1-R、SPb2-2-R、SPb2-3-R(表1),最后根据两次启动子序列拼接结果分别在距离ATG上游1 945 bp和164 bp处设计扩增全长启动子上下游引物,命名为pTeLCYb-F(-1945)和pTeLCYb-F(-164)(表1),PCR扩增程序体系及后续测序具体步骤参照1.2.1。
Table 1
表1
表1引物序列
Table 1Corresponding primer sequences
| 引物名称 Primer name | 序列 Sequences (5′ to 3′) | 退火温度 Tm | 产物 Products |
|---|---|---|---|
| TeLCYb-F | CACCATTTGAGGCATCCACA | 60.0℃ | cDNA |
| TeLCYb-R | CACTGCTAACCCTGAAGGACCA | 60.0℃ | cDNA |
| SPb1-1-R | CACTGCTAACCCTGAAGGACCA | 61.9℃ | 启动子 Promoter |
| SPb1-2-R | GGTATCCATGAAACCCAGATGTAAT | 58.7℃ | 启动子 Promoter |
| SPb1-3-R | GAGAGATTGGATAATGGGAAGTGGT | 60.3℃ | 启动子 Promoter |
| Spb2-1-R | ATGTGGATGCCTCAAATGGGG | 60.0℃ | 启动子 Promoter |
| Spb2-2-R | TTACGGTGACATTCAATCGCAT | 56.3℃ | 启动子 Promoter |
| Spb2-3-R | CCAGTGAGTTCAATGTTTTTCGTGT | 58.7℃ | 启动子 promoter |
| pTeLCYb-F(-1945) | CAGAGCGTATTGCACTTTTTCTAGT | 58.0℃ | 启动子 Promoter |
| pTeLCYb-F(-1117) | AGAAGCGATTGAATGTCACCGTAA | 56.0℃ | 启动子 Promoter |
| pTeLCYb-R(-164) | GCGAAGTGGGTTTGTGAAGAA | 58.0℃ | 启动子 Promoter |
| V097-F | GATGCCTGGCAGTTCCCTACTC | 58.0℃ | 载体引物 Primer of vector |
| V152-R | AGTTCACCTTGATGCCGTTCTT | 58.0℃ | 载体引物 Primer of vector |
新窗口打开
利用PlantCare(http://bioinformatics.psb.ugent. be/webtools/plantcare/html/)在线数据库分析万寿菊TeLCYb启动子调控元件。启动子序列生物信息学分析发现在ATG上游1 140 bp和164 bp区间有大量的光响应元件,而在ATG上游1 969 bp和1 140 bp区间主要是激素响应元件和耐热响应元件,为寻找具有增强启动子功能的的作用元件,对启动子序列进行缺失克隆,因此在ATG上游1 117 bp处设计引物pTeLCYb-F(-1117)(表1)。
1.2.3 植物表达载体的构建和农杆菌的转化 利用Gateway技术分别构建不同长度启动子的表达载体,分别命名为:pTeLCYb(-1969)∷GUS和pTeLCYb(-1140)∷GUS(数字为距离ATG上游的碱基长度)。用EcoRⅠ和SalⅠ分别双酶切含有目的片段克隆载体PMD-18T和中间载体V097,分别回收酶切产物后进行连接,将连接液用热激法导入到DH5α大肠杆菌中,检测阳性菌落提取质粒,再通过Getway技术将目的片段重组到含有GUS报告基因的V152表达载体上,LR反应采用5.0 μL的反应体系:入门克隆载体V097 1.0 μL(75 ng),表达载体V152 1.0 μL(75 ng),LR clonase enzyme mix 1.0 μL,TE溶液补充到5.0 μL。25℃反应12 h后,加入2.0 μL Proteinase K(蛋白酶K)solution,37℃温浴10 min,终止LR,反应液转化大肠杆菌DH5α感受态细胞中,挑菌落检测阳性,提取阳性菌落质粒,分别命名为V152-PTeLCYb(-1969)和V152-PTeLCYb(-1140)。各取1.0 μL阳性质粒经电转分别导入GV3101农杆菌感受态中,将转化细胞均匀涂布含100 mg∙L-1壮观霉素固体LB培养基上28℃暗培养2 d,以特异引物pTeLCYb-F(-1945)、pTeLCYb-F(-1117)和载体引物V152-R做阳性菌落PCR检测,挑取阳性菌落于相同浓度壮观霉素LB培养基内28℃震荡暗培养2 d,随后采用叶盘转化法转化早花烟草,具体转化方法参考HORSCH等[22]。
1.2.4 阳性苗的检测 采用SDS磁珠法[23]分别提取转化不同长度启动子的抗性烟草叶片的DNA,用特异引物pTeLCYb-F(-1945)、pTeLCYb-F(-1117)和载体引物V152-R对所提取的DNA做PCR检测。PCR扩增反应体系:PCR-Mix 12.5 μL,上、下游引物各1.0 μL,DNA模板1.0 μL,加去离子水补充到25 μL(Aidlab,北京,中国)。反应程序:95℃预变性4 min;95℃变性30 s,58℃退火 30 s,72℃延伸2 min,36个循环;72℃总延伸10 min,4℃保温10 min。PCR产物经1% 琼脂糖凝胶电泳检测,统计阳性植株数。
1.2.5 转基因烟草GUS化学组织染色 采集PCR检测阳性植株幼苗和花器官进行GUS化学组织染色,以未转化烟草作为阴性对照。GUS化学组织染色具体步骤参考JEFFERSON等[24]。
2 结果
2.1 万寿菊番茄红素β-环化酶基因获得和序列分析
根据已有的基因转录组数据设计扩增TeLCYb引物,测序结果发现万寿菊TeLCYb cDNA全长为1 865 bp,最大开放阅读框(Open Reading Frame,ORF)为1 527 bp,编码508个氨基酸残基。利用DNAMAN和BoxShade软件同源性比对分析发现该核苷酸序列推导的氨基酸序列与同科的西洋蒲公英Taraxacum officinale(BAE79544.1)、菊花Chrysanthemum morifolium(BAE79544.1)、刺苞菜蓟Cynara cardunculus var. Scolymus(KVI03465.1)的同源性分别为87.20%、85.10%、81.56%。对其氨基酸结构分析发现,TeLCYb氨基酸序列含有典型LCYb氨基酸结构如:β-lcyb保守域、LCY特异元件、β-lcyb CAD等(图1)。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1TeLCYb的氨基酸序列比对
-->Fig. 1Alignments of amino acid sequences of TeLCYb
-->
为探讨万寿菊TeLCYb与菊科中其他植物之间的关系,并进一步探讨与其他物种之间的系统进化关系,利用MEGA6.0构建含有13个LCYb氨基酸序列的进化树,采用邻近法1 000重复的自展检验完成。结果表明万寿菊TeLCYb与菊科植物单独聚为一个进化分枝,万寿菊TeLCYb与菊花Chrysanthemum morifolium和甘菊Chrysanthemum boreale的亲缘关系较近,与刺苞菜蓟Cynara cardunculus var. Scolymus的亲缘关系较远(图2)。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2基于MEGA6.0构建的TeLCYb系统进化树^括号内为该物种的LCYb氨基酸序列在GenBank中的登录号,分支上面的数字采用1 000重复的自展检验生成Numbers in parentheses represent the
-->Fig. 2Phylogenetic tree based on TeLCYb amino acid sequence using MEGA6.0^ sequences' accession number in GenBank. The number at each branch point is the percentage supported by 1 000 bootstrap
-->
2.2 万寿菊TeLCYb启动子克隆和顺式元件预测
利用染色体步移技术FPNI-PCR方法对TeLCYb启动子序列进行两次克隆,第一次PCR扩增产物经测序验证为一条1 343 bp的长度的启动子序列,采用同样的方法在第一次序列的基础上继续向上游扩增获得587 bp长度序列,经两次克隆拼接得到了ATG上游2 036 bp的序列。由于ATG上游区域包含基因5′非翻译区,因此本研究克隆全长启动子引物的下游引物设计在ATG上游164 bp处的位置,上游引物设计在ATG上游1 945 bp处的位置,最终获得一条长1 806 bp含有核心元件的启动子片段,命名为pTeLCYb(-1969)。利用PlantCare在线分析软件对TeLCYb启动子序列进行生物信息学分析,结果(表2)显示,该启动子除了典型的TATA-box和CAAT-box核心启动子元件外,还包含多种光应答元件、激素响应元件,值得注意的是该序列中有13个参与光应答及光调控的元件和5个参与激素响应的元件AuxRR-core、ARE、ERE等。另外,还有一些除了光响应元件和激素响应元件之外的其他环境响应元件的存在,如抗旱和高温响应元件CCAAT-box、MBS、HSE,厌氧必需的顺式元件ARE等。除了上述顺式作用元件外,还发现4个胚乳表达所需要的元件(Skn-1_motif)等。
通过以上对启动子顺式作用元件分析发现,光响应元件主要分布在ATG上游1 140 bp和164 bp区间,而在ATG上游1 969 bp和1 140 bp区间主要是激素响应元件(CGTCA-motif、ERE)和逆境胁迫响应元件(HSE),因此为了探究光响应元件以及激素响应元件(CGTCA-motif、ERE)和逆境胁迫响应元件(HSE)的特殊功能,设计上游引物TeLCYb(-1117),通过PCR扩增获得一条长度为977 bp的启动子片段,命名为pTeLCYb(-1140)(图3)。
Table 2
表2
表2启动子顺式作用元件分析
Table 2Analysis of promoter elements
| 元件 Elements | 序列及位置 Sequences and position | 数量 Number | 功能 Function |
|---|---|---|---|
| A-box | CCGTCC: -287 | 1 | 顺式调控元件 Cis-acting regulatory element |
| AuxRR-core | GGTCCAT: -370 | 1 | 参与生长素响应的顺式作用调节元件 Cis-acting regulatory element involved in auxin responsiveness |
| CCAAT-box | CAACGG: -1103 | 1 | MYBHv1结合位点 MYBHv1 binding site |
| MBS | TAACTG: -530 | 1 | 与抗性诱导相关的MTB结合位点 MYB binding site involved in drought-inducibility |
| CCGTCC-box | CCGTCC: -287 | 1 | 与分生组织特异性活化相关的顺式作用调节元件 Cis-acting regulatory element related to meristem specific activation |
| CGTCA-motif | CGTCA: -1450 | 1 | 参与茉莉酸甲酯响应的顺式调控作用元件 Cis-acting regulatory element involved in the MeJA- responsiveness |
| TGACG-motif | TGACG: -609, -1593 | 2 | 参与茉莉酸甲酯响应的顺式调控作用元件 Cis-acting regulatory element involved in the MeJA-responsiveness |
| ERE | ATTTCAAA:-1242 | 1 | 乙烯响应元件 Ethylene-responsive element |
| Skn-1_motif | GTCAT: -285, -372, -421, -921 | 4 | 胚乳表达所需的顺式调控元件 Cis-acting regulatory element required for endosperm expression |
| ARE | TGGTTT: -25, -30, -1625 | 3 | 厌氧诱导所必需的顺式作用调节元件 Cis-acting regulatory element essential for the anaerobic induction |
| HSE | AAAAAATTTC: -1766 | 1 | 热激响应顺式调控元件 Cis-acting element involved in heat stress responsiveness |
| GA-motif | AAAGATGA: -966 | 1 | 部分光响应元件 Part of a light responsive element |
| chs-CMA1a | TTACTTAA: -1937 | 1 | 部分光响应元件 Part of a light responsive element |
| GAG-motif | GGAGATG: -1063, -1718 | 2 | 部分光响应元件 Part of a light responsive element |
| GATA-motif | GATAGGG: -990 | 1 | 部分光响应元件 Part of a light responsive element |
| I-box | GATAGGG: -989 | 1 | 部分光响应元件 Part of a light responsive element |
| GT1-motif | GGTTAAT: -631 | 1 | 光响应元件 Light responsive element |
| Box I | TTTCAAA: -493, -1242 | 2 | 部分光响应元件 Light responsive element |
| Box 4 | ATTAAT: -429, -437 | 2 | 光响应中一个保守基因组件的部分 Part of a conserved DNA module involved in light responsiveness |
| ACE | AAAACGTTTA: -466 | 1 | 参与光应答的顺式元件 Cis-acting element involved in light responsiveness |
| G-Box | CACGTT: -1011 | 1 | 参与光应答的顺式调控元件 Cis-acting regulatory element involved in light responsiveness |
| AAGAA-motif | GAAAGAA: -1929 | 1 | 未知 Unknown |
新窗口打开
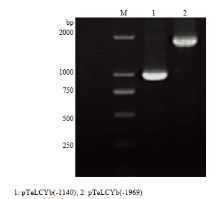 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3 TeLCYb启动子的克隆
-->Fig. 3Amplication of TeLCYb gene promoter
-->
2.3 转化烟草阳性苗的检测
PCR扩增检测转化苗叶片DNA,结果显示21棵转pTeLCYb(-1140)∷GUS的烟草植株中有16棵克隆出目的片段(图4-A);18棵转 pTeLCYb(-1969)∷GUS的烟草植株中有15棵克隆出目的片段,其他3棵为假阳性苗(图4-B)。2.4 GUS植物组织染色
采集阳性苗和未转化烟草的幼苗和整朵花进行GUS组织化学染色,结果显示阴性对照组无任何蓝色斑点出现;转化pTeLCYb(-1969)∷GUS的阳性植株在茎、花瓣、花药、柱头部位检测到GUS的表达,而其根、叶片和花萼中均无蓝色斑点;转化pTeLCYb(-1140)∷GUS的阳性植株的根、茎、叶、花萼、花瓣、花药以及柱头均有蓝色斑点出现,其中在花组织中,花药和柱头中GUS酶活性较花萼中强。不同长度启动子GUS组织化学染色结果比较,转化全长启动子的阳性苗的组织蓝色斑点颜色均浅于转pTeLCYb(-1140)∷GUS组织染色结果,且在绿色组织中转化pTeLCYb(-1969)∷GUS的阳性植株GUS不表达或表达水平很弱(图5),说明克隆的TeLCYb启动子能够成功的驱动下游GUS报告基因的表达,但不同长度的启动子对下游基因的作用强度不同。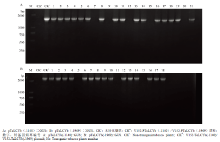 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4转基因烟草PCR检测
-->Fig. 4The identification of transgenic tobacco plants by PCR
-->
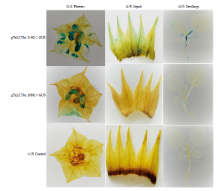 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5GUS活性检测
-->Fig. 5Detection of transient expression of GUS gene
-->
3 讨论
番茄红素β-环化酶是类胡萝卜素生物合成途径中的关键酶,是催化番茄红素向下游合成路径的重要分支点酶[25,26]。目前已在多种园艺作物中克隆出β-环化酶基因并对其功能进行了验证,如榴莲、红薯、柑橘[16,27-28]。本研究利用RT-PCR的方法获得了一条LCYb同源序列,采用生物信息学分析发现,该基因推导的氨基酸序列具有LCYb蛋白质特有的保守功能位点和特征多肽序列,同源序列比对和进化树分析均表明其与菊科植物同源性最高,进化关系最近,说明菊科植物在进化上是相对保守的。目前虽然在许多植物中LCYb已经被成功分离并进行了相应的功能验证,对于调控这些关键酶基因的启动子研究多是集中在经济作物[29,30,31],而对花卉植物LCYB启动子的研究除龙胆[32]外却鲜有报道。植物启动子在调控基因表达过程中起关键作用,本研究克隆得到了一条长度为1 806 bp的TeLCYb启动子序列,发现该启动子含有多种生物学顺式作用元件,推测TeLCYb的转录水平可能受到多种因素的调控。pTeLCYb(-1969)序列中含有多个光应答和参与光调控的元件,如:Box-I、I-box、Box-4、ACE、G-box等。YANG等[33]研究发现光响应元件Box-I、G-boxes、GAG motif、Box-4和chs-CMA1a等通过调节日长和其他刺激途径调控花发育过程中胡萝卜素的合成,另外早期的其他研究报道也发现胡萝卜色素的合成受光的调控[34,35],暗示万寿菊中TeLCYb的表达同样受光信号的调控。相比西瓜、柑橘、杧果LCYb启动子序列分析结果[30-31,36],万寿菊具有较多的且独有的光响应元件,如:chs-CMA1a、BOX-4、BOX-I,推测光信号调控在调节TeLCYb表达中具有主导作用。除了光应答元件,该启动子序列还包含多个参与激素响应的顺式作用元件,其中WISUTIAMONKUL等[28]研究发现内源乙烯调控榴莲LCYb的表达水平,推测TeLCYb的转录水平与激素的调节有关。除以上顺式作用元件外,该序列中还包含MYBHv1结合位点元件、耐热响应元件等非生物胁迫响应元件,暗示TeLCYb可能提高植物对非生物胁迫的耐受能力[16]。另外该序列还包含一些生理调控作用元件,如:ARE、Skn-1_motif,进一步说明TeLCYb的表达还可能受到生理因素的调控。
对转化pTeLCYb(-1969)启动子的转基因烟草阳性苗进行GUS化学组织染色,结果说明该启动子具有启动下游基因表达的功能,但在不同组织中启动子的作用强度不同,也进一步说明该启动子具有组织的特异性,同时也印证了TeLCYb在植物体内的广泛表达,这与前人[1,17]对万寿菊TeLCYb研究结果较为一致,不同之处是万寿菊花瓣中TeLCYb表达量最高,此差异可能是不同物种调控路径不同所导致,猜测叶黄素积累量高的植物组织LCYb表达量高。
由转化不同长度启动子的转基因烟草GUS植物组织染色结果的差异,推测pTeLCYb(-1140)启动子中的光响应元件可能具有增强下游基因在绿色组织中表达的作用,而pTeLCYb(-1969)启动子中的激素响应元件(CGTCA-motif、ERE)以及逆境胁迫响应元件(HSE)可能具有抑制作用;另外在花的组织染色中同样有较明显的差异,该区段启动子对下游GUS表达水平的正向调控能力明显大于pTeLCYb(-1969)启动子的调控作用,推测光响应元件可能具有增强子的功能。相关研究发现光响应元件Box-I、G-boxes、GAG motif、Box-4等通过调节日长和其他刺激途径调控花发育过程中胡萝卜素的合成[33];另外,早前的其他研究报道中也发现胡萝卜色素的合成受光的调控[34,35],这些报道更加证明了光响应元件具有正调控TeLCYb的表达。激素和逆境响应元件相关报道发现茉莉酸能够促进番茄红素在果实中的积累[37],乙烯响应元件ERE在橘柑有色体特有的CYCB启动子中能够调节果实的成熟[27],而果实的成熟和番茄红素的积累有关[34,35],猜测这些激素响应元件在万寿菊中通过抑制或减弱启动子的功能来降低TeLCYb的表达量,从而提高植株体内番茄红素的积累。关于HSE热响应元件在类胡萝卜素合成途径中的作用目前尚不清楚,根据本研究的结果推测其同样可能具有抑制或减弱启动子驱动下游基因表达的功能。因此针对本试验不同启动子作用下的GUS化学组织染色差异,推测光响应元件在调控下游基因的表达上具有主导作用且扮演着增强子的角色,而部分激素和逆境响应元件可能具有部分抑制子的功能。
4 结论
本研究利用RT-PCR的方法从万寿菊花中获得一条LCYb同源基因,生物信息学分析发现该基因推导的氨基酸序列具有典型的LCYb蛋白质特有的保守功能位点和特征多肽序列,同源序列比对和进化树分析均表明其和菊科植物同源性最高,进化关系最近。TeLCYb启动子顺式作用元件预测分析发现该启动子含有多种光响应元件、激素响应元件及逆境响应元件等。构建该启动子缺失表达载体,异源转化模式植物烟草,GUS化学组织染色结果显示不同长度的启动子均能驱动下游基因的表达,但GUS酶活性强度和表达部位存在差异,推测在ATG上游1 140 bp和164 bp区间的光响应元件可能具有增强子的功能,而全长启动子特有的激素响应元件和热响应元件可能具有抑制或降低启动子功能的作用。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}