0 引言
【研究意义】牦牛是川西北高原地区牧民的经济来源和肉、奶等主要食品来源之一。长期以来,牧区常利用当地优势藏药资源来进行牦牛疾病防治,很少使用到抗菌药物等化学药物。然而,近年来,随着川西北高原地区旅游业的发展和人员流动的增加,加上牦牛本身传染病的肆虐,使得耐药性革兰氏阴性菌在川西北高原地区开始出现。因此, 对川西北高原地区牦牛源大肠杆菌进行耐药性变迁规律研究,对保证该地区牧民和畜禽的健康有着重要的意义。【前人研究进展】大肠杆菌感染引起动物发病率的不断上升促使了不合理使用广谱抗生素的恶性循环,导致对临床抗菌药物的敏感性降低,而生物被膜的形成使得大肠杆菌对抗菌药物的耐药性更加严重,常常引起感染的慢性发作和迁延不愈[1-3]。国内外对牦牛源大肠杆菌耐药性调查研究相对其他食品动物如猪、鸡和牛较少[4-6]。王红宁课题组分别在2006年和2008年对川西北牦牛源致病性大肠杆菌进行药敏试验,发现菌株对常用抗菌药物表现出不同程度地耐药性[7-8]。贡嘎等对西藏6个地区的牦牛源大肠杆菌进行的耐药性分析结果显示对氟喹诺酮类药物高度敏感[9]。黑占珠和谢仲强对青海果洛州牦牛源大肠杆菌耐药性调查发现对试验药物最高耐药率不超过10.0%[10]。相关研究也表明,产志贺氏菌毒素大肠杆菌是引起牦牛腹泻病的主要病原之一[11-12]。近年来,大肠杆菌耐药性变迁演变的研究在人医领域和兽医领域都有报道,结果均提示大肠杆菌耐药呈现逐年增强的趋势[13-14]。然而,对川西北高原牦牛源大肠杆菌耐药性变迁和整合子携带情况的研究目前还没有系统的报道。【本研究切入点】本研究结合抗菌药物在川西北高原地区牦牛疾病防控中的使用情况和相关文献报道结果,于2009—2016年度对川西北高原地区散养牦牛、病死和牦牛屠宰场牦牛的胃肠道内容物和脏器等进行样本采集和大肠杆菌分离鉴定,分析其耐药水平和耐药性变迁规律,同时对其整合子携带情况进行调查。【拟解决的关键问题】通过对2009—2016年度川西北高原大肠杆菌耐药性变迁和整合子携带情况调查分析,以期为川西北高原地区牦牛源大肠杆菌的流行病学和防治药物的筛选提供理论依据,确保抗菌药物在川西北地区的应用正确合理。1 材料与方法
1.1 材料
1.1.1 样本来源 大部分牦牛粪便样本于2009—2016年采自四川省甘孜藏族自治州和阿坝藏族羌族自治州各县、乡、镇牦牛养殖场和散养牦牛,胃肠道内容物和脏器样本从病死的患病牦牛或屠宰牦牛采集,部分样本采自从川西北高原运送到青白江屠宰场的牦牛(具体样品来源信息见表1)。样本采集后运送至西南民族大学药理学实验室进行细菌分离鉴定等相关试验,大肠杆菌标准株ATCC25922由西南民族大学生命科学与技术学院兽医药理实验室保存。Table 1
表1
表12009-2016年牦牛源大肠杆菌样本采集信息表
Table 1Information table of E. coli isolated from yaks between 2009 and 2016
| 样本来源 Sample origin | 年份 Year | 小计 Sub-total | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | ||
| 粪便(健康牦牛) Faces (healthy yak) | 51 | 43 | 54 | 60 | 102 | 35 | 44 | 57 | 446 |
| 粪便(患病牦牛) Faces (ill yak) | 47 | 71 | 81 | 93 | 47 | 113 | 32 | 135 | 619 |
| 胃肠道内容物(屠宰牦牛) Gastrointestinal contents (slaughtered yak) | 36 | 47 | 32 | 43 | 103 | 76 | 79 | 90 | 506 |
| 脏器样本(病死牦牛) Viscera samples (dead yak) | 0 | 46 | 26 | 61 | 19 | 73 | 77 | 35 | 337 |
| 合计 Total | 1908 | ||||||||
新窗口打开
1.1.2 培养基和生化试剂 麦康凯琼脂培养基、伊红美兰培养基、蛋白胨大豆肉汤和MH肉汤等购自北京陆桥技术有限公司;肠杆菌科生化鉴定编码管(15e)购自杭州天和微生物试剂有限公司;分子生物学试剂购自宝生物工程(大连)有限公司;整合子整合酶基因扩增引物由生工生物工程(上海)股份有限公司合成。
1.1.3 抗菌药物 27个抗菌药物原料大多数购自中国兽药监察所或试剂公司,部分由兽药企业提供,主要包括以下几类:氨基糖苷类8个(庆大霉素、链霉素、妥布霉素、阿米卡星、大观霉素、新霉素、卡那霉素和安普霉素)、氟喹诺酮类6个(萘啶酸、沙拉沙星、左旋氧氟沙星、环丙沙星、恩诺沙星和达氟沙星)、β-内酰胺类4个(阿莫西林、头孢曲松、头孢喹肟和头孢噻呋)、磺胺类3个(磺胺嘧啶钠、磺胺间甲氧嘧啶和磺胺二甲嘧啶)、酰胺醇类2个(氯霉素和氟苯尼考)、四环素类2个(多西环素和土霉素)、利福霉素类1个(利福平)和喹啉类1个(乙酰甲喹)等。试验药物根据其理化性质配成5 120 μg·mL-1或640 μg·mL-1备用,临用前稀释至试验用浓度。
1.2 方法
1.2.1 不同样本来源大肠杆菌的分离和鉴定 参照大肠杆菌常规分离方法对不同年份采集的牦牛粪便、胃肠道内容物和脏器等样本进行处理,采用麦康凯培养基和伊红美兰培养基交替培养方法反复纯化,纯化菌株利用15e肠杆菌科生化鉴定编码管对之进行生化鉴定。1.2.2 药物敏感性试验 参照NCCLS(美国临床实验室标准化委员会)推荐的微量肉汤稀释法操作程序对1 908株大肠杆菌进行药物敏感性试验,每个药物重复3次,同时设立阴性对照和阳性对照[15]。药物敏感性试验结果利用WHONet5.6软件包进行处理和分析。
1.2.3 2009-2016年川西北牦牛源大肠杆菌耐药性变迁分析 根据1.2.2的结果,分别对2009—2016年不同年度采集的大肠杆菌进行耐药水平统计和处理,分析不同年份大肠杆菌耐药性变迁规律,以比较其耐药谱型的差异和分析耐药性变迁与耐药谱型的相关性。
1.2.4 2009-2016年川西北牦牛源大肠杆菌整合子携带情况调查分析 参考文献报道引物扩增Ⅰ类整合子和Ⅱ类整合子整合酶基因[16-17],对不同年份采集大肠杆菌的Ⅰ类整合子和Ⅱ类整合子携带情况进行调查,分析其与耐药性变迁的相关性。
2 结果
2.1 不同样本来源大肠杆菌分离情况
从不同年份健康和患病牦牛的粪便、病死和宰杀牦牛胃肠道内容物和脏器样本中共分离到大肠杆菌1 908株(表1,按样本来源统计结果为:健康牦牛粪便样本来源446株,患病牦牛粪便样本来源619株,屠宰场牦牛胃肠道内容物样本来源506株,病死牦牛脏器样本来源337株)。2.2 2009-2016年川西北牦牛源大肠杆菌耐药性变迁分析
1908株牦牛源大肠杆菌耐药性分析结果如表2所示,从中可以得出以下结论:(1)2009—2016各年度分离的牦牛源大肠杆菌对27个药物的耐药水平表现出逐年增高的趋势;除了2014—2016年度分离菌株对土霉素的耐药水平和2015—2016年度分离菌株对3个磺胺类药物的耐药水平超过60.00%外,其他年度分离菌株对其余试验药物的耐药水平均低于60.00%。(2)氨基糖苷类的敏感性分析结果显示:各年度分离菌株对妥布霉素和阿米卡星较敏感,其耐药水平比其余6个氨基糖苷类药物都低;分离菌株对链霉素和新霉素的耐药率较高。(3)氟喹诺酮类的敏感性分析结果显示:各年度分离菌株对萘啶酸的耐药率最高,其中2015年度分离菌株的耐药率达到为59.48%;各年度分离菌株对其余5个氟喹诺酮类药物耐药率都低于40.00%,且各年度的耐药率差别不明显。(4)β-内酰胺类的敏感性分析结果显示:各年度分离菌株对阿莫西林的耐药率最高,其中2013和2014年的耐药率分别高达54.98%和58.92%,对其余3个β-内酰胺类的敏感性差异不显著,除2014和2015年度头孢曲松的耐药率高于40.00%外,其余各年度的耐药率均在40.00%以下。(5)四环素类的敏感性分析结果显示:各年度分离菌株对多西环素和土霉素的敏感性差别不明显,对多西环素的耐药水平比土霉素低,试验结果也表明从2013年开始,川西北牦牛源大肠杆菌对四环素类药物的耐药率已高达40.00%以上。(6)酰胺醇类的敏感性分析结果显示:各年度分离菌株对氯霉素和氟苯尼考的敏感性差别不大。尽管氯霉素在兽医临床禁止使用,但2014和2016年度分离菌株对氯霉素的耐药水平均超过50.00%,应引起足够的重视。(7)磺胺类的敏感性分析结果显示:各年度分离菌株对磺胺嘧啶钠的耐药水平比磺胺间甲氧嘧啶和磺胺二甲嘧啶低,磺胺间甲氧嘧啶和磺胺二甲嘧啶的耐药水平差别不大。值得注意的是,从2015年度开始,分离菌株对3个磺胺类药物的耐药率均超过60.00%。(8)利福霉素类和喹啉类的敏感性分析结果显示:各年度分离菌株对两个药物的敏感性较高,各年度的耐药率均在30.00%以下,对乙酰甲喹的敏感性高于利福平。Table 2
表2
表22009-2016年牦牛源大肠杆菌耐药菌株数(株)和耐药率(%)汇总表
Table 2Summary sheet of numbers (strain) and drug resistance rates (%) of E. coli isolated from yaks between 2009 and 2016
| 药物 Drugs | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 菌株数(株) Number (strain) | 耐药率 Drug resistance rate (%) | 菌株数(株) Number (strain) | 耐药率 Drug resistance rate (%) | 菌株数(株) Number (strain) | 耐药率 Drug resistance rate (%) | 菌株数(株) Number (strain) | 耐药率 Drug resistance rate (%) | 菌株数(株) Number (strain) | 耐药率 Drug resistance rate (%) | 菌株数(株) Number (strain) | 耐药率 Drug resistance rate (%) | 菌株数(株) Number (strain) | 耐药率 Drug resistance rate (%) | 菌株数(株) Number (strain) | 耐药率 Drug resistance rate (%) | |
| 庆大霉素Gentamycin | 14 | 10.45 | 35 | 16.91 | 44 | 22.8 | 66 | 25.68 | 79 | 29.15 | 94 | 31.65 | 71 | 30.6 | 105 | 33.12 |
| 卡那霉素Kanamycin | 18 | 13.43 | 21 | 10.14 | 31 | 16.06 | 45 | 17.51 | 71 | 26.20 | 87 | 29.29 | 84 | 36.21 | 132 | 41.64 |
| 链霉素Streptomycin | 37 | 27.61 | 53 | 25.60 | 48 | 24.87 | 71 | 27.63 | 91 | 33.58 | 138 | 46.46 | 90 | 38.79 | 163 | 51.42 |
| 大观霉素Spectinomycin | 28 | 20.90 | 34 | 16.43 | 46 | 23.83 | 67 | 26.07 | 77 | 28.41 | 106 | 35.69 | 103 | 44.4 | 118 | 37.22 |
| 妥布霉素Tobramycin | 9 | 6.72 | 23 | 11.11 | 27 | 13.99 | 41 | 15.95 | 46 | 16.97 | 74 | 24.92 | 65 | 28.02 | 77 | 24.29 |
| 安普霉素Apramycin | 19 | 14.18 | 40 | 19.32 | 49 | 25.39 | 66 | 25.68 | 73 | 26.94 | 112 | 37.71 | 87 | 37.5 | 90 | 28.39 |
| 新霉素Neomycin | 40 | 29.85 | 54 | 26.09 | 61 | 31.61 | 74 | 28.79 | 104 | 38.38 | 146 | 49.16 | 102 | 43.97 | 147 | 46.37 |
| 阿米卡星Amikacin | 19 | 14.18 | 37 | 17.87 | 41 | 21.24 | 61 | 23.74 | 71 | 26.20 | 84 | 28.28 | 61 | 26.29 | 79 | 24.92 |
| 萘啶酸Nalidixic acid | 48 | 35.82 | 68 | 32.85 | 75 | 38.86 | 113 | 43.97 | 147 | 54.24 | 171 | 57.58 | 138 | 59.48 | 179 | 56.47 |
| 恩诺沙星Enrofloxacin | 18 | 13.43 | 41 | 19.81 | 44 | 22.80 | 57 | 22.18 | 76 | 28.04 | 111 | 37.37 | 89 | 38.36 | 112 | 35.33 |
| 沙拉沙星Sarafloxacin | 11 | 8.21 | 30 | 14.49 | 43 | 22.28 | 55 | 21.40 | 74 | 27.31 | 95 | 31.99 | 70 | 30.17 | 103 | 32.49 |
| 达氟沙星Danofloxacin | 17 | 12.69 | 33 | 15.94 | 38 | 19.69 | 47 | 18.29 | 81 | 29.89 | 101 | 34.01 | 71 | 30.6 | 100 | 31.55 |
| 左旋氧氟沙星Levofloxacin | 20 | 14.93 | 48 | 23.19 | 51 | 26.42 | 77 | 29.96 | 94 | 34.69 | 125 | 42.09 | 80 | 34.48 | 143 | 45.11 |
| 环丙沙星Ciprofloxacin | 21 | 15.67 | 43 | 20.77 | 50 | 25.91 | 83 | 32.30 | 91 | 33.58 | 130 | 43.77 | 82 | 35.34 | 109 | 34.38 |
| 头孢噻肟Cefquinome | 22 | 16.42 | 37 | 17.87 | 44 | 22.80 | 71 | 27.63 | 77 | 28.41 | 92 | 30.98 | 59 | 25.43 | 86 | 27.13 |
| 头孢噻呋Ceftiofur | 22 | 16.42 | 44 | 21.26 | 56 | 29.02 | 82 | 31.91 | 93 | 34.32 | 108 | 36.36 | 73 | 31.47 | 93 | 29.34 |
| 阿莫西林Amoxicillin | 52 | 38.81 | 79 | 38.16 | 77 | 39.90 | 105 | 40.86 | 149 | 54.98 | 175 | 58.92 | 117 | 50.43 | 167 | 52.68 |
| 头孢曲松Ceftriaxone | 32 | 23.88 | 56 | 27.05 | 69 | 35.75 | 84 | 32.68 | 104 | 38.38 | 134 | 45.12 | 93 | 40.09 | 94 | 29.65 |
| 土霉素Oxytetracycline | 20 | 14.93 | 47 | 22.71 | 53 | 27.46 | 79 | 30.74 | 143 | 52.77 | 184 | 61.95 | 143 | 61.64 | 207 | 65.3 |
| 多西环素Doxycycline | 20 | 14.93 | 38 | 18.36 | 49 | 25.39 | 70 | 37.24 | 115 | 42.44 | 167 | 56.23 | 97 | 41.81 | 101 | 31.86 |
| 氟苯尼考Florfenicol | 29 | 21.64 | 50 | 24.15 | 47 | 24.35 | 76 | 29.57 | 107 | 39.48 | 143 | 48.15 | 84 | 36.21 | 124 | 39.12 |
| 氯霉素Chloramphenicol | 51 | 38.06 | 36 | 17.39 | 51 | 26.42 | 87 | 33.85 | 147 | 54.24 | 164 | 55.22 | 104 | 44.83 | 173 | 54.57 |
| 磺胺间甲嘧啶* Sulfamonomethoxine* | 60 | 44.78 | 89 | 43.00 | 77 | 39.90 | 106 | 41.25 | 113 | 41.70 | 126 | 42.42 | 141 | 60.78 | 192 | 60.57 |
| 磺胺二甲嘧啶* Benzenesulfonamide* | 59 | 44.03 | 82 | 39.62 | 83 | 43.01 | 117 | 45.53 | 135 | 49.82 | 153 | 51.52 | 153 | 65.95 | 207 | 65.3 |
| 磺胺嘧啶钠*Sulfadizine sodium* | 16 | 11.94 | 33 | 15.94 | 44 | 22.80 | 61 | 23.74 | 83 | 30.63 | 98 | 33.00 | 147 | 63.36 | 197 | 62.15 |
| 乙酰甲喹Mequindox | 8 | 5.97 | 31 | 14.98 | 36 | 18.65 | 45 | 17.51 | 60 | 22.14 | 64 | 21.55 | 57 | 24.57 | 91 | 28.71 |
| 利福平Rifampicin | 20 | 14.93 | 50 | 24.15 | 57 | 29.53 | 68 | 26.46 | 84 | 31.00 | 91 | 30.64 | 61 | 26.29 | 92 | 29.02 |
新窗口打开
2.3 2009-2016年川西北牦牛源大肠杆菌整合子携带情况调查分析
采用常规PCR技术对Ⅰ类整合子和Ⅱ类整合子的整合酶基因进行扩增,分析不同年份分离菌株的整合子携带情况。以菌株Y20130606为例,其扩增结果如图1所示,其余菌株的Ⅰ类整合子和Ⅱ类整合子的整合酶扩增片段大小均与Y20130606一致。 2009—2016年川西北牦牛源大肠杆菌整合子携带情况调查分析结果见表3:1 908株细菌中,1 120株大肠杆菌不携带任何种类的整合子,占58.70%(1120/1908),其余788株携带至少一种类型整合子,占41.30%(788/1908)。从携带整合子类型统计分析结果来看,携带Ⅰ类整合子的菌株共有582株,占30.50%(582/1908),携带Ⅱ类整合子的菌株共有346株,占18.13%(346/1908),同时携带Ⅰ类整合子和Ⅱ类整合子的菌株共有140株,占7.34%(140/1908)。不同年份统计分析结果显示:分离菌株中Ⅰ类整合子携带率最高的年份是2016年,Ⅰ类整合子携带率为40.06%(127/317);Ⅱ类整合子携带率最高的年份是2011年,Ⅱ类整合子携带率为26.94%(52/193);2014年分离菌株中,同时携带Ⅰ类整合子和Ⅱ类整合子的菌株有32株,携带率为10.77%(32/297),高于其他年份。Table 3
表3
表32009-2016年牦牛源大肠杆菌整合子携带情况表
Table 3Integron-carrying of E. coli isolated from yaks between 2009 and 2016
| 整合子 Integrons | 年份 Year | 小计 Sub-total | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | ||
| 不携带整合子 No integrons | 103 | 141 | 93 | 147 | 157 | 172 | 151 | 156 | 1120 |
| Ⅰ类整合子 Class 1 integron | 21 | 48 | 60 | 80 | 95 | 86 | 65 | 127 | 582 |
| Ⅱ类整合子 Class 2 integron | 14 | 31 | 52 | 51 | 43 | 71 | 30 | 54 | 346 |
| Ⅰ类整合子和Ⅱ类整合子 Class 1 and class 2 integrons | 4 | 13 | 12 | 21 | 24 | 32 | 14 | 20 | 140 |
| 合计 Total | 1908 | ||||||||
新窗口打开
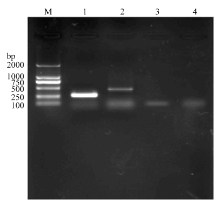 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1Y20130606整合子PCR扩增
M:DNA 标准 DL2000;1.Ⅱ类整合子整合酶基因;2. Ⅰ类整合子整合酶基因;3. Ⅱ类整合子整合酶基因阴性对照;4. Ⅰ类整合子整合酶基因阴性对照
-->Fig. 1PCR amplification of Y20130606 integrons
M: DNA marker DL2000; 1. Integrase gene of class 2 integron; 2. Integrase gene of class 1 integron; 3. Negative control of integrase gene of class 2 integron; 4. Negative control of integrase gene of class 1 integron
-->
3 讨论
3.1 川西北牦牛源大肠杆菌耐药性变迁分析
本研究对川西北牦牛源大肠杆菌进行分离鉴定和药物敏感性试验,样品涉及健康和患病牦牛的粪便、病死和宰杀牦牛的胃肠道内容物和脏器样本,从中共分离到1 908株。尽管本研究的药物敏感性结果表明大多数试验药物的耐药性变迁表现出逐年上升的趋势,但仍存在的一定的不足,分析本试验耐药性变迁的影响因素主要有以下两点:(1)试验菌株数量:试验中对2009—2016年度的大肠杆菌进行耐药性变迁演变分析,不同年份的菌株数量不同(其中2009年134株,2010年207株,2011年193株,2012年257株,2013年271株,2014年297株,2015年232株,2016年317株),样本数量的差异会造成耐药率有一定的偏差。尽管各年度的菌株数量均在100株以上,但笔者认为从统计分析的角度来讲,试验菌株数量越大其统计学意义越有说服力,各年份之间菌株数量的相差越小越能保证各年度菌株耐药性变迁的差异越小。在各年度大肠杆菌的分离鉴定过程中,虽然采样样本差别不大,但各年度的大肠杆菌分离率也不同,这可能与样本自身特点有一定关系,比如粪便样本几乎都能保证分离到大肠杆菌,而其他样本未必一定能分离到大肠杆菌。(2)菌株来源及其致病性:本研究中各年度分离菌株对相关药物的耐药水平均远低于相关报道[4],其原因是样本来源较广,而文献报道的样本仅采自患病动物,且动物的种类也不同。各年度分离菌株对庆大霉素、新霉素、土霉素和恩诺沙星的耐药水平均比王红宁[7-8]报道的结果要高,而对阿莫西林和链霉素等耐药率要低。与贡嘎等报道结果相比,除了卡那霉素的耐药水平相差较小外,各年度分离菌株对阿莫西林、四环素、多西环素、链霉素、庆大霉素、阿米卡星和新霉素的耐药水平均低于报道结果[9]。此外,牦牛用药习惯不同、样本的采集数量和动物的健康状况对菌株的耐药水平也有一定的影响,这可能也是本研究的耐药率高于芮亚培等的报道结果的原因[18-19],而且耐药性和毒力基因在不同菌株之间的传播,也可能导致在抗生素压力下,致病性菌株的耐药性和毒力发生改变[4,20]。3.2 川西北牦牛源大肠杆菌耐药性变迁与整合子携带情况相关性分析
对不同年份采集的大肠杆菌耐药水平进行比较分析,分离菌株对27个试验药物的耐药性变迁表现出逐年增加的趋势(表3)。不同年份的分离菌株整合子携带率虽未表现出线性递增的变化趋势,但从一定程度也反映出耐药性的逐年增加导致整合子的携带率增加。对试验菌株进行整合子携带情况分析的结果表明,788株大肠杆菌携带至少一种类型整合子,占41.30%(788/1908)。其中Ⅰ类整合子的检出率和同时携带Ⅰ类整合子和Ⅱ类整合子的检出率均比相关报道的要低[17,21-23]。而本研究中1 908株大肠杆菌中Ⅱ类整合子的检出率为18.13%,高于相关文献报道[24-28]。可以推测分离的牦牛源大肠杆菌Ⅱ类整合子携带率高可能与大肠杆菌生物被膜的形成有关,其整合子携带情况与生物被膜形成之间是否具有一定相关性还有待于进一步研究。3.3 川西北牦牛源大肠杆菌多重耐药性分析及其应对策略
在试验中笔者试图对不同年份分离的实验菌株进行优势耐药谱型分析,但结果显示耐药谱型较为分散,分离菌株对27个试验药物均表现出一定的耐药性,且出现多重耐药菌株。多重耐药菌株的出现,与整合子的携带有一定的关系,整合子携带的基因盒元件能够在整合酶的作用下将不同的耐药基因整合,使得基因盒的大小和耐药基因的种类有一定的差异。此外,整合子作为细菌的可移动元件,能够使耐药基因在不同菌株个体之间进行传播,尤其是抗生素压力下大肠杆菌生物被膜的形成导致耐药性的广泛传递和感染的迁延不愈,给临床治疗带来一定的困难,必须引起高度的重视[20]。川西北牦牛源大肠杆菌耐药性变迁规律,能够为后续研究中其多重耐药性与生物被膜表型和毒力谱型之间的相关性提供参考。革兰氏阴性菌尤其是大肠杆菌、铜绿假单孢杆菌和鲍曼不动杆菌的多重耐药性问题已引起全球的关注[29-31]。针对细菌耐药性广泛传播的现状,采用植物药物提取物来应对耐药性引起传染病肆虐传播和迁延不愈的棘手问题提供了一个的新的思路和方法[32-34]。此外,基于抗菌药物作用机制的化学合成方法来寻求新构型化合物也是解决细菌耐药性问题的有效途径之一[35]。这些研究内容提示,在后续研究工作中,可以结合川西北独特的藏兽药资源优势,从中寻求对牦牛常见病和多发病起到防治作用的有效成分或药物组合,不仅能够为藏兽药的现代药理学研究及其在藏区兽医临床的广泛应用开辟新的空间,而且对藏兽医文化的传承和保护也有一定的促进作用。
4 结论
川西北高原地区分离的牦牛源大肠杆菌表现出一定的演变规律,分离菌株对试验药物的耐药水平呈现逐年增加的趋势。此外,整合子携带情况分析结果也预示着耐药性的增加导致多重耐药性菌株的出现,耐药性变迁和整合子携带存在一定的相关性。因此后续研究应重点探讨牦牛源多重耐药大肠杆菌耐药谱型与整合子携带相关性,从分子水平更加深入分析整合子-基因盒系统在川西北牦牛源细菌耐药性传播过程中的作用机理。同时,结合川西北藏区广袤的藏兽药资源优势,从中筛选针对牦牛常见病和多发病有效的主要化学成分或药物组合,不仅为牦牛的健康养殖保驾护航提供一条崭新的防治途径和方法,而且能够为藏兽药的推广和发扬光大提供更为广阔的发展空间。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}