0 引言
【研究意义】黍稷(Panicum miliaceum L.)起源于中国[1],属于禾本科(Gramineae)黍属(Panicum L.),一年生草本植物,具有早熟、耐旱、耐瘠的特性[2]。全世界黍稷栽培面积约600万公顷,主产区分布在欧洲和亚洲,主产国为原苏联和中国,中国主要分布在西北、华北、东北地区,南方有零星种植[3]。黍稷能适应多种土壤,对肥力较差的砂土有较强的适应能力,耐盐碱能力也较强,常作为盐碱地和开荒沙漠的先锋作物。因其生育期短,也作为自然灾害后的补救作物[4]。中国虽是黍稷的主产国,并在种质资源方面取得了一些研究成果,但基于DNA水平分子标记的研究开发却相对较少,应用分子标记的报道则更少,尤其是随着黍稷种质资源研究的不断深入,对黍稷的国外种,野生种和野生近缘植物的搜集、研究和保护更为需要。【前人研究进展】近年来,DNA分子标记发展迅速,已作为研究遗传变异的重要手段应用于各种作物的遗传多样性研究中,如谷子[5]、玉米[6]、陆地棉[7]、野生稻[8]等。其中,SSR可直接检测DNA分子结构变异[9]、并能反映参试材料差异、灵敏度较高、稳定性较好、操作较简单[10],因而在遗传多样性分析[11]、种质鉴定、DNA指纹图谱构建[10]、基因定位和分子标记育种[12]中广泛应用。此外,SSR分子标记为共显性,具有多态性丰富、PCR结果重现性高和易于鉴别基因型的优点[13],是群体遗传标记变异的最好标记之一[14]。国内外关于黍稷种质资源遗传多样性的研究一直匮乏。M'RIBU等[15]应用RAPD技术研究4种黍稷的遗传多样性以判定其来源,根据原产地的地理区域对黍稷品种进行了划分。KARAM等[16-17]对12份来源于美国和加拿大的黍稷利用AFLP标记检测其遗传多样性。LÁGLER等[18]选用9个来自草莓属植物的ISSR标记检测了21份匈牙利黍稷,检测到15个等位基因,其中只有7个引物扩增出DNA片段。HUNT等[19]利用禾本科作物(水稻、小麦、燕麦、大麦)的46对SSR引物对118份材料进行SSR多样性分析,得出黄土高原生态型材料的遗传多样性比较丰富,有可能是黍稷的起源中心的结论。CHO等[20]通过一个SSR富集文库黍基因组DNA开发了25个多态性SSR标记并分析了50份黍稷材料的遗传多样性;连帅等[21]利用5个黍稷特异性SSR标记对5个黍稷栽培生态区的40份黍稷进行遗传多样性分析,发现SSR标记表现出了一定的地域性;LI等[22]通过核糖体DNA(rDNA)分析内部转录间隔区(ITS)和外部转录间隔区(ETS),发现黍稷在ITS具有特异性的特点,并通过对新疆墓地出土的古DNA和现代品种黍稷材料的分析,得到黍稷遗传多样性随时间流逝而部分丧失,还进一步推出新疆在黍稷驯化和传播中东西方联系的十字路口的重要地位。【本研究切入点】由于可应用的分子标记很少且多态性不高,已有研究不足以全面反映黍稷的遗传多样性水平。小宗作物课题组利用二代测序技术大规模开发了黍稷基因组SSR引物,并分析了黍稷育成品种的遗传多样性(文章已接受待出版)。【拟解决的关键问题】本研究采用前期筛选得到的162对多态性SSR标记进一步对保存在国家种质库国内不同生态区的地方栽培品种、野生种以及从国外收集引进来自不同国家的黍稷种质资源进行遗传多样性分析。以揭示黍稷种质资源群体间的遗传多样性关系与遗传多样性地理分布的特点,为黍稷种质资源的研究、保护和利用提供依据,同时也为后续进行黍稷的起源、演化、传播研究提供数据。1 材料与方法
1.1 供试材料
试验材料包括来自国家种质资源库的50份国内材料和近年新收集引进的142份国外材料(电子附表)。其中国内资源包括31份栽培种(山西2份、内蒙3份、黑龙江3份、甘肃5份、新疆3份、陕西4份、吉林3份、宁夏3份、河北2份、辽宁1份、海南1份、青海1份)和19份野生资源(山西9份、内蒙5份、黑龙江3份、甘肃1份、新疆1份)。142份国外资源分别来自蒙古(81份)、印度(28份)、美国(11份)、苏联(8份)、土耳其(4份)、波兰(3份)、德国(3份)、匈牙利(1份)、吉尔吉斯斯坦(1份)、哈萨克斯坦(1份)、韩国(1份)。根据参试材料的来源地,首先将其分成国内种和国外种两大群体;再进一步将国内栽培种和野生种质分开,分为国外种,国内栽培种和国内野生种3个组群;最后根据栽培区、地理来源将来自国内(12个省(区))和国外(11个国家)的材料分为10亚群,分别为内蒙古高原区(内蒙古)、东北区(黑龙江、吉林、辽宁、河北承德)、黄土高原区(山西、陕西、宁夏)、西北区(甘肃、新疆、青海)、南方区(海南)、蒙古(蒙古、韩国)、印度、美国、前苏联、欧洲(土耳其、波兰、德国、匈牙利、吉尔吉斯斯坦、哈萨克斯坦)。Supplement table 1
附表1
附表1参试材料
Supplement table 1Broomcorn millet varieties and landraces used in this study
| 群体编号与名称 Group number and name | 材料标号 Material number | 编号 Uniform Number | 材料名称 Material name | 材料来源 Material source |
|---|---|---|---|---|
| 1黄土高原Loess Plateau Region | 1 | 00003140 | 野生黄黍子Yeshenghuangshuzi | 晋大同市南郊区Shanxidatongnanjiaoqu |
| 黄土高原Loess Plateau Region | 2 | 00003148 | 野糜Yemi | 晋广灵Shanxiguangling |
| 黄土高原Loess Plateau Region | 3 | 00003163 | 野糜子Yemizi | 晋天镇Shanxitianzhen |
| 黄土高原Loess Plateau Region | 4 | 00003178 | 野糜Yemi | 晋阳高Shanxiyanggao |
| 黄土高原Loess Plateau Region | 5 | 00003179 | 野糜子Yemizi | 晋阳高Shanxiyanggao |
| 黄土高原Loess Plateau Region | 6 | 00003258 | 野生糜Yeshengmi | 晋平鲁Shanxipinglu |
| 黄土高原Loess Plateau Region | 7 | 00003345 | 野生糜Yeshengmi | 晋太原南郊Shanxitaiyuannanjiao |
| 黄土高原Loess Plateau Region | 8 | 00003523 | 野糜子Yemizi | 晋汾西Shanxifengxi |
| 黄土高原Loess Plateau Region | 9 | 00006762 | 野糜子Yemizi | 山西Shanxi |
| 黄土高原Loess Plateau Region | 10 | 00000936 | 跳蚤黍Tiaozaoshu | 山西Shanxi |
| 黄土高原Loess Plateau Region | 11 | 00001316 | 鸡爪红Jizhuahong | 山西Shanxi |
| 黄土高原Loess Plateau Region | 12 | 00001624 | 活剥皮糜Huobopimi | 陕西Shaanxi |
| 黄土高原Loess Plateau Region | 13 | 00001662 | 瓦灰软糜Wahuiruanmi | 陕西Shaanxi |
| 黄土高原Loess Plateau Region | 14 | 00001665 | 大红糜Dahongmi | 陕西Shaanxi |
| 黄土高原Loess Plateau Region | 15 | 00001767 | 大瓦灰Dawahui | 陕西Shaanxi |
| 黄土高原Loess Plateau Region | 16 | 00000702 | 惠农黄粘黍Huinonghuangnianshu | 宁夏Ningxia |
| 黄土高原Loess Plateau Region | 17 | 00000708 | 花软糜子Huaruanmizi | 宁夏Ningxia |
| 黄土高原Loess Plateau Region | 18 | 00002530 | 贺兰二黄Helanerhuang | 宁夏Ningxia |
| 2西北地区Northwest Region | 19 | 00006767 | 野黍子Yeshuzi | 甘肃Gansu |
| 西北地区Northwest Region | 20 | 00002674 | 酒泉小黄糜子Jiuquanxiaohuangmizi | 甘肃Gansu |
| 西北地区Northwest Region | 21 | 00002878 | 环县60天紫秆红小糜Huanxian60tianziganhongxiaomi | 甘肃Gansu |
| 西北地区Northwest Region | 22 | 00002899 | 华池红紫秆Huachihongzigan | 甘肃Gansu |
| 西北地区Northwest Region | 23 | 00007314 | 安西糜Anximi | 甘肃Gansu |
| 西北地区Northwest Region | 24 | 00007332 | 鼓鼓头糜Gugutoumi | 甘肃Gansu |
| 西北地区Northwest Region | 25 | 00003056 | 黄糜子Huangmizi | 新疆Xinjiang |
| 西北地区Northwest Region | 26 | 00003111 | 红糜Hongmi | 新疆Xinjiang |
| 西北地区Northwest Region | 27 | 00003117 | 黄糜Huangmi | 新疆Xinjiang |
| 西北地区Northwest Region | 28 | 00006766 | 野糜子Yemizi | 新疆Xinjiang |
| 西北地区Northwest Region | 29 | 00005532 | 大黄糜Dahuangmi | 青海Qinghai |
| 3东北地区Northeast region | 30 | 00000039 | 黑鹅头Heietou | 黑龙江Heilongjiang |
| 东北地区Northeast region | 31 | 00000132 | 鹌鹑尾Anchunwei | 黑龙江Heilongjiang |
| 东北地区Northeast region | 32 | 00000147 | 麦糜子Maimizi | 黑龙江Heilongjiang |
| 东北地区Northeast region | 33 | 未入库 | 泰来大野糜Tailaidayemi | 黑龙江齐齐哈尔Qiqihaer, Heilongjiang |
| 东北地区Northeast region | 34 | 未入库 | 泰来小野糜Tailaixiaoyemi | 黑龙江齐齐哈尔Qiqihaer, Heilongjiang |
| 东北地区Northeast region | 35 | 未入库 | 杜尔伯特野糜Duerpoteyemi | 黑龙江齐齐哈尔Qiqihaer, Heilongjiang |
| 群体编号与名称 Group number and name | 材料标号 Material number | 编号 Uniform Number | 材料名称 Material name | 材料来源 Material source |
| 东北地区Northeast region | 36 | 00000330 | 黎糜子Limizi | 吉林Jilin |
| 东北地区Northeast region | 37 | 00000333 | 黄糜子Huangmizi | 吉林Jilin |
| 东北地区Northeast region | 38 | 00005700 | 翰章黄糜子Hanzhanghuangmizi | 吉林Jilin |
| 东北地区Northeast region | 39 | 00000440 | 大白黍Dabaishu | 辽宁Liaoning |
| 东北地区Northeast region | 40 | 00006036 | 小红黍Xiaohongshu | 河北承德Chengde, Hebei |
| 东北地区Northeast region | 41 | 00006042 | 小白黍Xiaobaishu | 河北承德Chengde, Hebei |
| 4内蒙古高原 Inner Mongolia plateau region | 42 | 00004279 | 内黍一点红Neishuyidianhong | 内蒙古Inner Mongolia |
| 内蒙古高原 Inner Mongolia plateau region | 43 | 00000525 | 巴林左疙塔黍Balinzuogatashu | 内蒙古Inner Mongolia |
| 内蒙古高原 Inner Mongolia plateau region | 44 | 00000569 | 五原黑黍子Wuyuanheishuzi | 内蒙古Inner Mongolia |
| 内蒙古高原 Inner Mongolia plateau region | 45 | 00007224 | 东胜野糜Dongshengyemi | 内蒙伊盟Inner Mongolia yimeng |
| 内蒙古高原 Inner Mongolia plateau region | 46 | 00007225 | 异交野糜Yijiaoyemi | 内蒙伊盟Inner Mongolia yimeng |
| 内蒙古高原 Inner Mongolia plateau region | 47 | 00006761 | 野糜子Yemizi | 伊盟东胜Inner Mongolia yimengdongsheng |
| 内蒙古高原 Inner Mongolia plateau region | 48 | 00006763 | 野糜子Yemizi | 伊盟东胜Inner Mongolia yimengdongsheng |
| 内蒙古高原 Inner Mongolia plateau region | 49 | 00006764 | 典型紫秆野糜Dianxingziganyemi | 伊盟东胜Inner Mongolia yimengdongsheng |
| 5南方地区Southern Region | 50 | 00006655 | 糯黍Nuoshu | 海南Hainan |
| 6蒙古Mongolia | 51 | mk-147 | Dolinskoe-86 | 蒙古Mongolia |
| 蒙古Mongolia | 52 | Mk-148 | Saratovskoe-2 | 蒙古Mongolia |
| 蒙古Mongolia | 53 | mk-153 | Uralskoe 1419 | 蒙古Mongolia |
| 蒙古Mongolia | 54 | mk-154 | Kazanskoe-506 | 蒙古Mongolia |
| 蒙古Mongolia | 55 | mk-158 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 56 | mk-163 | Mestnoe | 蒙古Mongolia |
| 蒙古Mongolia | 57 | mk-164 | -10 | 蒙古Mongolia |
| 蒙古Mongolia | 58 | mk-166 | Tuvinskoe mestnoe | 蒙古Mongolia |
| 蒙古Mongolia | 59 | mk-188 | Saratovskoe-853 | 蒙古Mongolia |
| 蒙古Mongolia | 60 | mk-168 | Uzbekstan | 蒙古Mongolia |
| 蒙古Mongolia | 61 | mk-169 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 62 | mk-172 | Beskaraganskoe-2 | 蒙古Mongolia |
| 蒙古Mongolia | 63 | mk-176 | Russia | 蒙古Mongolia |
| 蒙古Mongolia | 64 | mk-177 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 65 | mk-181 | 蒙古Mongolia | |
| 蒙古Mongolia | 66 | mk-182 | Veselo Podolyanskoe 24-273 | 蒙古Mongolia |
| 蒙古Mongolia | 67 | mk-193 | DolInskoe-12 | 蒙古Mongolia |
| 群体编号与名称 Group number and name | 材料标号 Material number | 编号 Uniform Number | 材料名称 Material name | 材料来源 Material source |
| 蒙古Mongolia | 68 | mk-199 | Veselopodolinskoe 367 | 蒙古Mongolia |
| 蒙古Mongolia | 69 | mk-207 | Omskoe-9 | 蒙古Mongolia |
| 蒙古Mongolia | 70 | mk-215 | Turgaiskoe mestnoe | 蒙古Mongolia |
| 蒙古Mongolia | 71 | mk-1046 | Veselo podolinskoe 38 | 蒙古Mongolia |
| 蒙古Mongolia | 72 | mk-1683 | Uilskoe beloe | 蒙古Mongolia |
| 蒙古Mongolia | 73 | mk-1687 | Mironovskoe-85 | 蒙古Mongolia |
| 蒙古Mongolia | 74 | mk-3835 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 75 | mk-3865 | Dolinskoe-83 | 蒙古Mongolia |
| 蒙古Mongolia | 76 | mk-3867 | Mestnoe | 蒙古Mongolia |
| 蒙古Mongolia | 77 | mk-3868 | Mestnoe | 蒙古Mongolia |
| 蒙古Mongolia | 78 | mk-3869 | Kamishenskoe 123 | 蒙古Mongolia |
| 蒙古Mongolia | 79 | mk-3870 | Kazanskoe-176 | 蒙古Mongolia |
| 蒙古Mongolia | 80 | mk-3871 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 81 | mk-3872 | Amurskoe mestnoe | 蒙古Mongolia |
| 蒙古Mongolia | 82 | mk-3873 | Beskaragaiskoe mestnoe | 蒙古Mongolia |
| 蒙古Mongolia | 83 | mk-3874 | Turgaiskoe mestnoe | 蒙古Mongolia |
| 蒙古Mongolia | 84 | mk-3877 | Ostrogojskoe-9 | 蒙古Mongolia |
| 蒙古Mongolia | 85 | mk-3880 | Kamalinskoe-591 | 蒙古Mongolia |
| 蒙古Mongolia | 86 | mk-3898 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 87 | mk-5097 | Poli ploidnoe | 蒙古Mongolia |
| 蒙古Mongolia | 88 | mk-5106 | Mestnoe | 蒙古Mongolia |
| 蒙古Mongolia | 89 | mk-5108 | Mestnoe | 蒙古Mongolia |
| 蒙古Mongolia | 90 | МК-5110 | Lobanovskoe mestnoe | 蒙古Mongolia |
| 蒙古Mongolia | 91 | mk-5111 | Mestnoe | 蒙古Mongolia |
| 蒙古Mongolia | 92 | mk-5115 | Mestnoe | 蒙古Mongolia |
| 蒙古Mongolia | 93 | mk-5118 | Mestnoe | 蒙古Mongolia |
| 蒙古Mongolia | 94 | mk-5121 | Mestnoe | 蒙古Mongolia |
| 蒙古Mongolia | 95 | mk-5126 | Mestnoe | 蒙古Mongolia |
| 蒙古Mongolia | 96 | mk-5128 | Kedabekskoe-41 | 蒙古Mongolia |
| 蒙古Mongolia | 97 | mk-5129 | Hybrid BHUC 29 | 蒙古Mongolia |
| 蒙古Mongolia | 98 | I-3256 | saratovskoe 2 | 蒙古Mongolia |
| 蒙古Mongolia | 99 | I-3887 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 100 | I-5020 | Kormovoe-45 | 蒙古Mongolia |
| 蒙古Mongolia | 101 | I-7979 | Unnown | 蒙古Mongolia |
| 蒙古Mongolia | 102 | I-7984 | Bershevskoe-18 | 蒙古Mongolia |
| 蒙古Mongolia | 103 | I-7988 | Saratovskoe-3 | 蒙古Mongolia |
| 群体编号与名称 Group number and name | 材料标号 Material number | 编号 Uniform Number | 材料名称 Material name | 材料来源 Material source |
| 蒙古Mongolia | 104 | I-7993 | Omskoe-5 | 蒙古Mongolia |
| 蒙古Mongolia | 105 | I-8003 | Abakanskoe korm | 蒙古Mongolia |
| 蒙古Mongolia | 106 | I-9131 | Mutant 84-10909 | 蒙古Mongolia |
| 蒙古Mongolia | 107 | I-9134 | Uralskoe109 | 蒙古Mongolia |
| 蒙古Mongolia | 108 | I-9143 | Bezinnukskoe 10 | 蒙古Mongolia |
| 蒙古Mongolia | 109 | I-10068 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 110 | I-15425 | Gorlinko | 蒙古Mongolia |
| 蒙古Mongolia | 111 | I-15426 | Lipekts | 蒙古Mongolia |
| 蒙古Mongolia | 112 | I-18898 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 113 | I-7983 | Uilskoe mestnoe | 蒙古Mongolia |
| 蒙古Mongolia | 114 | Mk-1687 | 蒙古Mongolia | |
| 蒙古Mongolia | 115 | mk-1130 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 116 | mk-185 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 117 | mk-1043 | Sibirskoe | 蒙古Mongolia |
| 蒙古Mongolia | 118 | mk-1040 | Kamalinskoe-40 | 蒙古Mongolia |
| 蒙古Mongolia | 119 | I-23287 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 120 | I-23288 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 121 | I-23289 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 122 | I-23290 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 123 | I-23291 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 124 | I-23292 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 125 | I-23293 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 126 | I-23294 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 127 | I-23295 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 128 | I-23296 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 129 | I-3543 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 130 | I-5646 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 131 | I-18897 | Local | 蒙古Mongolia |
| 蒙古Mongolia | 132 | PI649380 | Mooju 6 | 韩国Korea |
| 7印度India | 133 | 00007372 | 790032 | 印度India |
| 印度India | 134 | 00007373 | 790033 | 印度India |
| 印度India | 135 | 00007374 | 790034 | 印度India |
| 印度India | 136 | 00007375 | 790035 | 印度India |
| 印度India | 137 | 00007376 | 790036 | 印度India |
| 印度India | 138 | 00007377 | 790037 | 印度India |
| 印度India | 139 | 00007378 | 790038 | 印度India |
| 群体编号与名称 Group number and name | 材料标号 Material number | 编号 Uniform Number | 材料名称 Material name | 材料来源 Material source |
| 印度India | 140 | 00007379 | 790039 | 印度India |
| 印度India | 141 | 00007380 | 790040 | 印度India |
| 印度India | 142 | 00007381 | 790041 | 印度India |
| 印度India | 143 | 00007382 | 790042 | 印度India |
| 印度India | 144 | 00007383 | 790043 | 印度India |
| 印度India | 145 | 00007384 | 790044 | 印度India |
| 印度India | 146 | 00007386 | 790046 | 印度India |
| 印度India | 147 | 00007388 | 790048 | 印度India |
| 印度India | 148 | 00007389 | 790049 | 印度India |
| 印度India | 149 | 00007390 | 790050 | 印度India |
| 印度India | 150 | 00007391 | 790051 | 印度India |
| 印度India | 151 | 00007392 | 790052 | 印度India |
| 印度India | 152 | 00007393 | 790053 | 印度India |
| 印度India | 153 | 00007394 | 790054 | 印度India |
| 印度India | 154 | 00007395 | 790055 | 印度India |
| 印度India | 155 | 00007396 | 790056 | 印度India |
| 印度India | 156 | 00007397 | 790057 | 印度India |
| 印度India | 157 | 00007398 | 790058 | 印度India |
| 印度India | 158 | 00007399 | 790059 | 印度India |
| 印度India | 159 | 00007400 | 790060 | 印度India |
| 印度India | 160 | 00007401 | 790061 | 印度India |
| 8美国U.S.A | 161 | 00007664 | 外引黍1号Waiyinshu 1 hao | 美国U.S.A |
| 美国U.S.A | 162 | 00007665 | 外引黍2号Waiyinshu 2 hao | 美国U.S.A |
| 美国U.S.A | 163 | 00007666 | 外引黍3号Waiyinshu 3 hao | 美国U.S.A |
| 美国U.S.A | 164 | 00007667 | 外引黍4号Waiyinshu 4 hao | 美国U.S.A |
| 美国U.S.A | 165 | 00007668 | 外引黍5号Waiyinshu 5 hao | 美国U.S.A |
| 美国U.S.A | 166 | 00007669 | 外引黍6号Waiyinshu 6 hao | 美国U.S.A |
| 美国U.S.A | 167 | 00007670 | 外引黍7号Waiyinshu 7 hao | 美国U.S.A |
| 美国U.S.A | 168 | 00007671 | 外引黍8号Waiyinshu 8 hao | 美国U.S.A |
| 美国U.S.A | 169 | PI578073 | EARLYBIRD | 美国,内布拉斯加州U.S.A Nebraska |
| 美国U.S.A | 170 | PI578074 | HUNTSMAN | 美国,内布拉斯加州U.S.A Nebraska |
| 美国U.S.A | 171 | PI583347 | SUNRISE | 美国,内布拉斯加州U.S.A Nebraska |
| 9前苏联Former Soviet Union | 172 | PI260053 | VESELOPOLIANSKOYI367 | 前苏联Former Soviet Union |
| 前苏联Former Soviet Union | 173 | PI531410 | KAMUSINSZKOE67 | 前苏联Former Soviet Union |
| 前苏联Former Soviet Union | 174 | PI531411 | KOMSOMOLSKOE996 | 前苏联Former Soviet Union |
| 前苏联Former Soviet Union | 175 | PI531412 | KAZANSKOE176 | 前苏联Former Soviet Union |
| 群体编号与名称 Group number and name | 材料标号 Material number | 编号 Uniform Number | 材料名称 Material name | 材料来源 Material source |
| 前苏联Former Soviet Union | 176 | PI531414 | KRASNOKUTSKOE48 | 前苏联Former Soviet Union |
| 前苏联Former Soviet Union | 177 | PI531415 | KUSOTA | 前苏联Former Soviet Union |
| 前苏联Former Soviet Union | 178 | 00007366 | 78 | 前苏联Former Soviet Union |
| 前苏联Former Soviet Union | 179 | 00007367 | 支多奥斯支Zhiduoaosizhi | 前苏联Former Soviet Union |
| 10欧洲Europe | 180 | PI649373 | TU-86-42-03 | 土耳其Turkey |
| 欧洲Europe | 181 | PI649374 | Garvis | 土耳其Turkey |
| 欧洲Europe | 182 | PI654403 | TU-85-074-03 | 土耳其,比特利斯Turkey Bitlis |
| 欧洲Europe | 183 | PI654404 | TU-85-087-01 | 土耳其,比特利斯Turkey Bitlis |
| 欧洲Europe | 184 | 00007369 | 色查尔托Sechaertuo | 波兰Poland |
| 欧洲Europe | 185 | 00007370 | 黑糜Heimi | 波兰Poland |
| 欧洲Europe | 186 | 00007371 | 灰糜Huimi | 波兰Poland |
| 欧洲Europe | 187 | PI531408 | HARKOVSKOE65 | 德国Germany |
| 欧洲Europe | 188 | PI531413 | KORNBERGER MITTELFRUHE | 德国Germany |
| 欧洲Europe | 189 | PI649371 | Index Seminum #568 | 德国,萨克森Germany Saxony |
| 欧洲Europe | 190 | PI346936 | TOKOQULSK | 吉尔吉斯斯坦Kyrgyzstan |
| 欧洲Europe | 191 | PI346938 | URAL1419 | 哈萨克斯坦Kazakhstan |
| 欧洲Europe | 192 | PI531416 | MALCALTOR"A" | 匈牙利Hungary |
新窗口打开
Supplement table 2
附表2
附表2所用SSR引物的序列及退火温度
Supplement table 2Sequence of SSR primers and annealing temperature
| 位点 Locus | 正向引物 Forward primer (5'-3') | 退火温度 Tm (℃) | 反向引物 Reverse primer (5'-3') | 退火温度 Tm (℃) |
|---|---|---|---|---|
| BM5 | CCCTTCTTCCCTACTTTGCC | 60.07 | GTGTGCGTGCATGGGTGT | 62.82 |
| BM484 | GGACAGGATGAGGAGGATCA | 60.01 | CGGTTACCATCGCCTTCTTA | 60.09 |
| BM552 | GCAGCAATAGATTTGCCTCC | 59.81 | TCTAGCCTACCCGAACTTGC | 59.48 |
| BM637 | CACACTTGTGCTGTTGGGTC | 60.2 | TCTAGCCTACCCCAACTTGC | 59.34 |
| BM787 | TCCGTGACATGATGCAGACT | 60.28 | GCACTATTTCTGCAGGAGCC | 59.98 |
| BM994 | AACAGAACCGCACATCAGTG | 59.75 | TCAGGTGAGCGATTCTTCCT | 59.95 |
| BM1303 | TTGACGCGTACAACAAGAGC | 60.06 | ACCGAAGAGAAGGTTGCTGA | 59.99 |
| BM1332 | CGGTTCAGTATTCAGGGCAT | 59.96 | CACTACACGTACCCGGTCCT | 59.91 |
| BM1419 | CGGTTCAGTATTCAGGGCAT | 59.96 | CACAAGGATCAGGCCAAAGT | 60.11 |
| BM1420 | CGCGCACACAAAAACATAGT | 59.8 | ACCGGTCGAGGATGTGTAAG | 59.99 |
| BM1477 | GCGAAAGATGAACTGCCAAT | 60.22 | GCTGCGACGGATACTGATCT | 60.39 |
| BM1533 | CTCTTGTCGTCTTGGTCCGT | 60.3 | CGTGCGTGTGTCGAGAGAG | 61.87 |
| BM1701 | CAGCTGATCATTTGAAAAGTATGG | 60.02 | TAGTTGTCGCGATCATGAGG | 59.82 |
| BM1710 | TATTCGAGCCCCATTTCTTG | 60.03 | GCGTTATCCGGATGATGAAG | 60.44 |
| BM1745 | CAGCTGATCATTTGAAAAGTATGG | 60.02 | TAGTTGTCGCGATCATGAGG | 59.82 |
| LMX265 | GGCTTTGCTAGGGTTTCTCC | 60.21 | GGTGTGAAGTTGCCCAGATT | 59.97 |
| LMX503 | GTGGTACAGCTGCTCGTTCA | 60.06 | AGGAGGAACCAGGAAGCAAT | 60.07 |
| LMX510 | GTGGTACAGCTGCTCGTTCA | 60.06 | CAAGACAGGGAAGGAACCAA | 60.08 |
| 位点 Locus | 正向引物 Forward primer (5'-3') | 退火温度 Tm (℃) | 反向引物 Reverse primer (5'-3') | 退火温度 Tm (℃) |
| LMX515 | CGTTTTCTCGCTACACACGA | 60.05 | TGGACAACGGAAAACGTACA | 60 |
| LMX630 | ATGAATCACCCGATCCACAT | 60.02 | ACGCCAACATCAGCATATCA | 60.1 |
| LMX632 | GCTGTCGGTCAGTCCTGTTT | 60.31 | ACGCCAACATCAGCATATCA | 60.1 |
| LMX645 | GCTAGGTGTCGTTTTCTCGC | 60.02 | GGGGCGTCTAAGCTTGATATT | 59.61 |
| LMX653 | CGATGAACGAAAATTCACCC | 60.31 | GTTCATTCGTCCAAATGCCT | 59.94 |
| LMX684 | GGCCTCCTGTGGTCTTCTCT | 60.79 | GACCCTCTCCTCCTACCACC | 59.93 |
| LMX691 | ACTCATGGTTACGGCAACTG | 58.65 | GCGCGAGAGAGAGAGAGAGA | 60.27 |
| LMX746 | CGTCCACCTTGGTGCTTATT | 59.99 | GCTGATTTTCTAACGGCTGC | 59.99 |
| LMX780 | CTGCATTCTCTGTTCACCCA | 59.83 | ATCCTTTCACTCGAGGGGTT | 59.93 |
| LMX786 | CCTGGACACACACACACACA | 60.09 | TCTTGTCACTGTCGGCGTAG | 60.05 |
| LMX794 | GTGGTGAGAAGAAGCCTTGC | 60 | GATGGGTCGATCAGCAGAAT | 60.04 |
| LMX836 | GCGCAGTAATATATTTCAGTAATTCA | 58.07 | GCATCATCGTCAAGACCTCA | 59.79 |
| LMX845 | GCGCAGTAATATATTTCAGTAATTCA | 58.07 | GCATCATCGTCAAGACCTCA | 59.79 |
| LMX850 | GCGCAGTAATATATTTCAGTAATTCA | 58.07 | GCATCATCGTCAAGACCTCA | 59.79 |
| LMX1036 | CAGAGCAGTGCGGTATTGTG | 60.47 | TCGTTTGTTGTTCGGTTGTC | 59.59 |
| LMX1065 | TCTGGACATGCTTTCACCAG | 59.83 | CCTACCTCGTAACACTGCGG | 60.7 |
| LMX1067 | ATCGACGACTAGGCCCTGTA | 59.72 | TGCGGAGTGTCTTTGTTCTG | 60.02 |
| LMX1071 | ATCGACGACTAGGCCCTGTA | 59.72 | TGTATGGAAAGCTCTGGCCT | 59.84 |
| LMX1072 | TCTGGACATGCTTTCACCAG | 59.83 | TACCTGTAACACTGCGGCTG | 59.93 |
| LMX1097 | TGCCAAGGACTAGCTGAAAGA | 60.14 | TTCCGGTTACAGACGGTAGG | 59.99 |
| LMX1220 | AGTTACTTTTCCACACCCGC | 59.1 | TTTATTGGGAGTTCGGATGG | 59.76 |
| LMX1251 | GCAGTTTTGTTTCGATCGGT | 60.12 | TGGTGTCAAGACATGTGGATG | 60.43 |
| LMX1380 | GCCTCCTGTCTTGTAGCGTC | 60.02 | AGGGTAGGCTGAGAGCCTGT | 60.41 |
| LMX1387 | TTTCAGGGACTGGACTGGAC | 60.09 | GTAGGGGGTAGCTGAGAGCC | 60.23 |
| LMX1400 | AGCAACGGAGGTGAGAGAGA | 60.14 | TCGACACACACGACACACAC | 60.27 |
| LMX1477 | GCTACTACGCACACATTGCAC | 59.43 | CTCTCATCCACTCAGCCACA | 59.98 |
| LMX1553 | ATTTGACCTGTGACCTCGCT | 59.73 | AGGGCTCTCGAGGAGTGTTT | 60.39 |
| LMX1672 | ATTTGACCTGTGACCTCGCT | 59.73 | CCTTTCTGTTTCTGCAAGCC | 59.99 |
| LMX1749 | CTCAACACCACACCTTCCCT | 60 | CTTTTGATCCATGGGCGTAG | 60.46 |
| LMX1760 | TGTGGGAGAGAAGTGGGC | 59.75 | CAAGGAAGGAATAAACCGCA | 60.07 |
| LMX1761 | GAGATGGTGCGGATTCTGAG | 60.77 | TCATTTCCACTGTCACTGCC | 59.68 |
| LMX1959 | GTGGTGAGAAGAAGCCTTGC | 60 | GATGGGTCGATCAGCAGAAT | 60.04 |
| LMX2019 | CCTCTCCTTACACGGGGATT | 60.32 | TTGATTATGCTTTGGAGGGG | 59.89 |
| LMX2068 | TGAGATTGGCATCAAGCAAG | 59.95 | TTTCTGGTCAGTTCGGTCAG | 58.85 |
| LMX2074 | GCCACACTAAATAAGCTTTGTGTC | 59.29 | TGGTCGTCACTGATTACGGA | 60.11 |
| LMX2175 | GTGACCGACGACTACCGTTT | 60.04 | CACCCCTTTCCTCTCGCTAT | 60.59 |
| LMX2187 | CGATTGCACACACGGATAAC | 60 | GTCGTTGTGCTTGCACTTGT | 59.95 |
| LMX2281 | TTCTCCGTCAGCTCACATTG | 59.98 | TCCATTGTTCATTTAGTAGAAACCT | 57.49 |
| LMX2305 | CGCACTAGCCCTTGTCTTTC | 60.02 | CGCCCTACGAACAAATCACT | 60.13 |
| LMX2410 | CTCAACACCACACCTTCCCT | 60 | CTTTTGATCCATGGGCGTAG | 60.46 |
| LMX2551 | TATTCGAGCCCCATTTCTTG | 60.03 | AGCGTATCCGGATGATGAAG | 60.06 |
| LMX2734 | CACACAGATATTTGGCACCG | 59.99 | TGAGGATCCGAAAAGATTGG | 60.01 |
| LMX2782 | GCCGGAGTATAGATCCGACA | 60.06 | GTCAGGCCGTGAACGTTATT | 60 |
| LMX2901 | ATGCACGCACGAACACATA | 59.72 | TCTTGATCATCACCAGCACC | 59.64 |
新窗口打开
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT附图1凝胶电泳图
-->Supplement Fig. 1Polyacrylamide gel electrophoresis (SDS-PAGE) spectra
-->
1.2 DNA提取与检测
每个参试材料随机选取20个单株中的幼嫩叶片组织60 mg混合后,用新型植物基因组DNA快速提取试剂盒(北京鼎国昌盛生物有限公司)提取基因组DNA,采用1%的琼脂糖凝胶电泳检测其质量,之后再用NanoDropND-1000(NanoDrop,Wilmington,DE,USA)测定浓度后加入100 mL双纯水将其稀释成DNA母液于-80℃低温下保存备用。1.3 SSR扩增及电泳
采用162对小宗作物课题组通过高通量测序技术开发的黍稷基因组SSR引物,选用6份不同地理来源、性状差异显著的黍稷种质资源(国外3份,国内3份)为DNA模板进行引物的PCR扩增和多态性筛选,最终筛选出63对条带清晰,多态性稳定的SSR引物用于后续的遗传多样性分析。SSR扩增反应在MY-CYCLERPCR仪上进行。20 μL反应体系中含1.2 μL基因组DNA、0.6 μL引物、0.4 μL Taq酶(2.5 U·μL-1)、2 μL 10×buffer、0.8 μL dNTP(10 mmol·L-1)和ddH2O 15.0 μL。PCR程序为94℃ 5 min;94℃ 30 s,55℃ 30 s,72℃ 1 min,30个循环;72℃ 5 min。然后于16℃保存。PCR扩增产物在8%聚丙烯酰胺凝胶上进行电泳分离,之后用硝酸银染色显影。
1.4 数据统计分析
分别记录同一SSR引物扩增电泳条带(等位变异)在各参试材料中的有无。若有,根据分子量从大到小的顺序,记录为A、B、C……;若无,记为..。不同群体间和群体内的某一位点的观测等位变异数(observed number of alleles,Na)、有效等位变异数(effective number of alleles,Ne)、观测杂合度(observed heterozygosity,Ho)、期望杂合度(expected heterozygosity,He)及Shannon-Weaver指数等遗传参数的统计计算和不同群体间的Nei78遗传距离在Popgen1.32软件包[23]中完成;在PowerMarkerV3.25软件包和MEGA6软件包[24]中完成参试材料间多态性信息含量指数(polymorphism information content index,PIC)和Nei83遗传距离频率计算及群体间遗传距离频率的聚类图绘制。2 结果
2.1 SSR多态性引物的筛选
利用随机选择的6份材料对162对SSR引物进行PCR退火温度和引物扩增多态性筛选,有77对SSR引物能扩增出清晰、稳定、可辨识的谱带,有效扩增比率为48.1%,其中63对SSR引物具有多态性,多态性引物比率为77.8%。最终选用63对具有稳定多态性的引物进行全部参试材料的遗传多样性分析。2.2 SSR位点多样性分析
192份参试材料在63对SSR引物上共检测出161个等位变异(表1),变幅为2—4个,平均每对SSR引物扩增出2.56个等位变异,其中有效等位变异数1.74,有效等位变异所占比重为67.97%。其中,引物BM552、BM1701、LMX1251等位变异最多,为4个,其有效等位变异数分别为2.0741、1.7599和1.1824。多态性信息量(PIC)的变幅介于0.0877—0.8020,平均值为0.4855,PIC指数最大的标记是BM637,最小的标记是LMX786。观测杂合度(Ho)在0(LMX1071、1387)—0.9405(LMX1332)变化,平均为0.2940。期望杂合度(He)的平均值为0.3887,最大是0.6664(BM637),最小为0.028(LMX684)。Nei基因多样性指数的变化范围是0.0279(LMX684)—0.6645(BM637)。Shannon-Weaver指数值最大的位点是BM637,最小的为LMX684,Shannon-Weaver指数变化范围为0.0743—1.0953,平均0.6275。结果表明,获得的SSR位点的遗传多样性比较丰富,且不同指标能从不同角度揭示黍稷SSR引物在遗传多样性方面存在的巨大差异,其中,效等位变异数、Shannon-Weaver指数与Nei基因多样性指数呈现正相关,能够反映群体遗传多样性。因此,有效等位变异数、Nei基因多样性指数和Shannon-Weaver指数这三个参数对于遗传多样性分析意义重大。Table 1
表1
表163对SSR引物测定的遗传参数
Table 1Genetic parameters from the 63 polymorphic SSR markers used in this study
| 位点 Locus | 观测等位基因数 Na | 有效等位基因数 Ne | 多样性指数 I | 观察杂合度 Ho | 期望杂合度 He | Nei’s期望杂合度 Nei | 多态性信息 含量指数PIC |
|---|---|---|---|---|---|---|---|
| BM5 | 2 | 1.2047 | 0.3111 | 0.8296 | 0.1704 | 0.1699 | 0.3010 |
| BM484 | 2 | 1.714 | 0.6072 | 0.5819 | 0.4181 | 0.4166 | 0.6099 |
| BM552 | 4 | 2.0741 | 0.8807 | 0.4806 | 0.5194 | 0.5179 | 0.7208 |
| BM637 | 3 | 2.9804 | 1.0953 | 0.3336 | 0.6664 | 0.6645 | 0.8020 |
| BM787 | 2 | 1.6835 | 0.596 | 0.5926 | 0.4074 | 0.406 | 0.6083 |
| BM994 | 2 | 1.1308 | 0.2315 | 0.8839 | 0.1161 | 0.1157 | 0.4295 |
| BM1303 | 2 | 1.9956 | 0.692 | 0.4998 | 0.5002 | 0.4989 | 0.2577 |
| BM1332 | 3 | 2.6195 | 1.0177 | 0.3801 | 0.6199 | 0.6182 | 0.5027 |
| BM1419 | 2 | 1.9476 | 0.6796 | 0.5122 | 0.4878 | 0.4865 | 0.2883 |
| BM1420 | 2 | 1.6082 | 0.5658 | 0.6206 | 0.3794 | 0.3782 | 0.6070 |
| BM1477 | 2 | 1.1263 | 0.2259 | 0.8875 | 0.1125 | 0.1121 | 0.3704 |
| BM1533 | 2 | 1.5355 | 0.5332 | 0.6503 | 0.3497 | 0.3487 | 0.4541 |
| BM1701 | 4 | 1.7599 | 0.7401 | 0.5671 | 0.4329 | 0.4318 | 0.5850 |
| BM1710 | 2 | 1.9266 | 0.674 | 0.5176 | 0.4824 | 0.481 | 0.4546 |
| BM1745 | 3 | 1.3492 | 0.4934 | 0.7403 | 0.2597 | 0.2588 | 0.5292 |
| LMX265 | 3 | 2.4656 | 0.9909 | 0.404 | 0.596 | 0.5944 | 0.6176 |
| LMX503 | 3 | 2.6693 | 1.029 | 0.3722 | 0.6278 | 0.6254 | 0.7674 |
| LMX510 | 3 | 2.2019 | 0.8561 | 0.4522 | 0.5478 | 0.5458 | 0.7049 |
| LMX515 | 3 | 2.1395 | 0.9121 | 0.4659 | 0.5341 | 0.5326 | 0.7107 |
| LMX621 | 3 | 1.55 | 0.6605 | 0.6442 | 0.3558 | 0.3548 | 0.5183 |
| LMX630 | 3 | 1.4333 | 0.5811 | 0.6968 | 0.3032 | 0.3023 | 0.4524 |
| LMX632 | 3 | 2.0421 | 0.8632 | 0.4883 | 0.5117 | 0.5103 | 0.6435 |
| LMX645 | 3 | 1.3235 | 0.4827 | 0.7549 | 0.2451 | 0.2444 | 0.3876 |
| LMX653 | 2 | 1.962 | 0.6834 | 0.5066 | 0.4934 | 0.4903 | 0.5434 |
| LMX684 | 2 | 1.0287 | 0.0743 | 0.972 | 0.028 | 0.0279 | 0.3956 |
| LMX691 | 2 | 1.8556 | 0.6537 | 0.5377 | 0.4623 | 0.4611 | 0.3918 |
| LMX746 | 3 | 1.4144 | 0.5681 | 0.7062 | 0.2938 | 0.293 | 0.4958 |
| LMX780 | 2 | 1.4928 | 0.5121 | 0.6689 | 0.3311 | 0.3301 | 0.5148 |
| LMX786 | 2 | 1.0816 | 0.1656 | 0.9244 | 0.0756 | 0.0755 | 0.0877 |
| 位点 Locus | 观测等位基因数 Na | 有效等位基因数 Ne | 多样性指数 I | 观察杂合度 Ho | 期望杂合度 He | Nei’s期望杂合度 Nei | 多态性信息 含量指数PIC |
| LMX794 | 2 | 1.6364 | 0.5775 | 0.6098 | 0.3902 | 0.3889 | 0.5653 |
| LMX836 | 3 | 1.6664 | 0.6582 | 0.599 | 0.401 | 0.3999 | 0.4399 |
| LMX845 | 2 | 1.5117 | 0.5216 | 0.6606 | 0.3394 | 0.3385 | 0.3773 |
| LMX850 | 3 | 1.6674 | 0.7255 | 0.5986 | 0.4014 | 0.4003 | 0.5692 |
| LMX1036 | 3 | 1.9091 | 0.7448 | 0.5226 | 0.4774 | 0.4762 | 0.4403 |
| LMX1065 | 3 | 2.9717 | 1.0939 | 0.3348 | 0.6652 | 0.6635 | 0.6204 |
| LMX1067 | 3 | 1.4663 | 0.5989 | 0.6811 | 0.3189 | 0.318 | 0.5084 |
| LMX1071 | 2 | 1.9912 | 0.6909 | 0.4938 | 0.5062 | 0.4978 | 0.2583 |
| LMX1072 | 3 | 2.1828 | 0.8493 | 0.4566 | 0.5434 | 0.5419 | 0.5715 |
| LMX1097 | 3 | 1.8858 | 0.7218 | 0.5291 | 0.4709 | 0.4697 | 0.3983 |
| LMX1220 | 3 | 2.0277 | 0.7836 | 0.4918 | 0.5082 | 0.5068 | 0.4107 |
| LMX1251 | 4 | 1.1824 | 0.362 | 0.8453 | 0.1547 | 0.1542 | 0.2757 |
| LMX1380 | 3 | 1.9786 | 0.8197 | 0.504 | 0.496 | 0.4946 | 0.5825 |
| LMX1387 | 2 | 1.489 | 0.5101 | 0.6706 | 0.3294 | 0.3284 | 0.4180 |
| LMX1400 | 3 | 1.6448 | 0.6682 | 0.6069 | 0.3931 | 0.392 | 0.3898 |
| LMX1477 | 2 | 1.0977 | 0.1887 | 0.9107 | 0.0893 | 0.089 | 0.1955 |
| LMX1553 | 2 | 1.6234 | 0.5722 | 0.615 | 0.385 | 0.384 | 0.4608 |
| LMX1672 | 2 | 1.869 | 0.6577 | 0.5337 | 0.4663 | 0.465 | 0.6325 |
| LMX1749 | 2 | 1.0794 | 0.1623 | 0.9262 | 0.0738 | 0.0736 | 0.1866 |
| LMX1760 | 2 | 1.1403 | 0.2428 | 0.8766 | 0.1234 | 0.1231 | 0.3026 |
| LMX1761 | 3 | 1.9608 | 0.7829 | 0.5086 | 0.4914 | 0.49 | 0.5336 |
| LMX1959 | 2 | 1.9878 | 0.6901 | 0.5014 | 0.4986 | 0.4969 | 0.6416 |
| LMX2019 | 3 | 1.4169 | 0.568 | 0.7049 | 0.2951 | 0.2942 | 0.4741 |
| LMX2068 | 3 | 2.2602 | 0.9269 | 0.4408 | 0.5592 | 0.5576 | 0.6762 |
| LMX2074 | 2 | 1.4474 | 0.4878 | 0.6896 | 0.3104 | 0.3091 | 0.5541 |
| LMX2175 | 2 | 1.5433 | 0.5369 | 0.647 | 0.353 | 0.3521 | 0.4743 |
| LMX2187 | 2 | 1.441 | 0.4841 | 0.6931 | 0.3069 | 0.306 | 0.4074 |
| LMX2281 | 3 | 1.5565 | 0.6561 | 0.6414 | 0.3586 | 0.3575 | 0.5101 |
| LMX2305 | 2 | 1.6125 | 0.5677 | 0.6189 | 0.3811 | 0.3799 | 0.5726 |
| LMX2410 | 3 | 1.647 | 0.6718 | 0.6061 | 0.3939 | 0.3928 | 0.5227 |
| LMX2551 | 2 | 1.9997 | 0.6931 | 0.4985 | 0.5015 | 0.4999 | 0.4641 |
| LMX2734 | 3 | 2.5493 | 1.0127 | 0.3906 | 0.6094 | 0.6077 | 0.7006 |
| LMX2782 | 3 | 1.214 | 0.3552 | 0.8232 | 0.1768 | 0.1763 | 0.2126 |
| LMX2901 | 2 | 1.6303 | 0.575 | 0.6122 | 0.3878 | 0.3866 | 0.4897 |
| 平均值Mean | 2.5556 | 1.7397 | 0.6275 | 0.6113 | 0.3887 | 0.3874 | 0.4855 |
| 标准差SD | 0.5896 | 0.4517 | 0.2337 | 0.1562 | 0.1562 | 0.1557 |
新窗口打开
2.3 不同来源黍稷资源群体间的遗传变异分析
由表2可知,各参试群体的有效等位变异变化范围较窄,最小的是南方群体,为1.2407±0.4315;最大的是内蒙古高原群体,为1.8846±0.4892;总体看来,有效等位变异,国内5个群体大于国外各群体。国内群体间的有效等位变异依次为内蒙古高原>东北地区>黄土高原>西北地区>南方地区,变化范围为(1.2407±0.4315)—(1.8846±0.4892)。国外群体间有效等位变异依次是前苏联>欧洲>蒙古>印度>美国。变化范围为(1.5522±0.4658)—(1.7409±0.4662)。分析有效等位变异结果可知,国内外群体间的有效等位变异差异不大,但国内群体间的遗传丰富度总体大于国外群体。Table 2
表2
表210个群体的遗传多样性分析
Table 2Estimates of genetic diversity within 10 populations of the main cultivation areas of broomcorn millet
| 群体 Population | 观测等位基因数Na | 有效等位基因数 Ne | 多样性指数 I | 观测杂合度 Ho | 期望杂合度 He | Nei’s期望 杂合度Nei | 多态性信息 含量指数PIC |
|---|---|---|---|---|---|---|---|
| 黄土高原Loess Plateau | 2.3175±0.6678 | 1.7597±0.5053 | 0.6065±0.3112 | 0.3430±0.3174 | 0.3917±0.2035 | 0.3784±0.1972 | 0.4329 |
| 西北地区Northwest | 2.2381±0.6651 | 1.6759±0.5085 | 0.5544±0.3068 | 0.3279±0.3324 | 0.3650±0.2093 | 0.3461±0.1995 | 0.3873 |
| 东北地区Northeast | 2.3810±0.5214 | 1.8489±0.4554 | 0.6621±0.2469 | 0.3144±0.3391 | 0.4431±0.1690 | 0.4218±0.1605 | 0.4393 |
| 内蒙古高原 Inner Mongolia plateau region | 2.3016±0.6126 | 1.8846±0.4892 | 0.6694±0.2799 | 0.3966±0.3250 | 0.4622±0.1862 | 0.428±0.1717 | 0.4673 |
| 南方地区Southern | 1.2407±0.4315 | 1.2407±0.4315 | 0.1669±0.2991 | 0.2407±0.4315 | 0.2407±0.4315 | 0.1204±0.2158 | 0.2344 |
| 蒙古Mongolia | 2.3492±0.6263 | 1.6437±0.4608 | 0.5449±0.2790 | 0.2696±0.2802 | 0.346±0.1847 | 0.3436±0.1834 | 0.4510 |
| 印度India | 2.3065±0.6164 | 1.5874±0.4757 | 0.5169±0.2822 | 0.2372±0.2962 | 0.3254±0.1914 | 0.3175±0.1865 | 0.3684 |
| 美国U.S.A | 1.9677±0.6521 | 1.5522±0.4658 | 0.4584±0.3102 | 0.3101±0.3712 | 0.3114±0.2203 | 0.2957±0.2094 | 0.3025 |
| 前苏联Former Soviet Union | 2.1311±0.5620 | 1.7409±0.4662 | 0.5916±0.2722 | 0.3539±0.3347 | 0.4118±0.1863 | 0.3825±0.1735 | 0.3769 |
| 欧洲Europe | 2.2097±0.6308 | 1.6848±0.4714 | 0.5673±0.2783 | 0.3232±0.3494 | 0.3758±0.1882 | 0.3594±0.1800 | 0.3922 |
新窗口打开
国内群体的Shannon-Weaver指数最小值为0.1669±0.2991;最大值为0.6694±0.2799,均值为0.5319± 0.2888,分布也较国外更为宽泛。国内群体Shannon- Weaver指数为内蒙古高原>东北地区>黄土高原>西北地区>南方地区,其中内蒙古高原群体与东北地区群体的Shannon-Weaver指数接近,只差0.007。国外群体Shannon-Weaver指数范围为(0.4584±0.3102)—(0.5916±0.2722),均值为0.5358±0.2849。Shannon-Weaver指数排序依次为前苏联>欧洲>蒙古>印度>美国。由Shannon- Weaver指数分析结果可知,国内群体间的Shannon- Weaver指数即遗传均匀度差异不大,但国内群体总体高于国外。
从Nei’s基因杂合度分析,观察杂合度(Ho)最小的是印度群体,为0.2372±0.2962,最大的是内蒙古高原群体,为0.3966±0.3250;前苏联群体次之,为0.3539±0.3347。期望杂合度(He)最小的是美国群体,为0.3114±0.2203;最大的是内蒙古高原群体,为0.4622±0.1862。总体来看,Ho值国内群体比国外群体略小,而He值国内群体却比国外群体稍大。
对于有效等位变异数和Shannon-Weaver指数,不同群体的变化趋势是一致的。如国内内蒙古高原、东北群体都远大于国外群体。而对于观察杂合度和期望杂合度来说,变化趋势却并不完全一致,但总的结果是国内群体的Nei基因多样性指数是高于国外群体。因此,等位变异数、有效等位变异数以及有效等位变异所占比重等指标,可从不同侧面揭示各群体材料的特点。Nei基因多样性指数、Shannon-Weaver指数则综合反映了各群体的遗传多样性。
据此,综合各遗传参数可知,国内黍稷种质遗传多样性总体高于国外种质。国外群体中,前苏联、欧洲的遗传多样性较高。而在国内群体中,内蒙古高原、东北地区的遗传多样性较高,除南方地区外基本均高于国外群体。南方地区因不是黍稷栽培的主载区,取材少,类型单一,多样性低。
就国内资源和国外资源两大群体而言(表3),国内资源的有效等位基因数(1.8145±0.4519)、Shannon-Weaver指数(0.6657±0.2413)和Nei基因多样性指数(0.412±0.1574)均大于国外资源(1.6862±0.4527、0.5897±0.2469、0.3652±0.1655)。因此,认为此次分析的国内黍稷资源的遗传多样性大于国外种质资源。而从国外种、国内栽培种和国内野生种3个大群体来看(表4),野生种质资源有效等位基因数(1.9285±0.5101)、Shannon-Weaver指数(0.6948±0.2852)、Nei基因多样性指数(0.4373± 0.1773)远大于国外种和国内栽培种,而由于缺少了野生种的国内栽培种也同时因材料过少,有效等位基因数(1.6727±0.4612)、Shannon-Weaver指数(0.5528± 0.2877)、Nei基因多样性指数(0.352±0.1925)略低于国外种(1.6862±0.4527、0.5897±0.2469、0.3652± 0.1655),由此可知,国内野生资源在黍稷遗传多样性中的重要地位。
Table 3
表3
表3国内外群体遗传多样性分析
Table 3Estimates of genetic diversity of populations of broomcorn millet from home and abroad
| 群体 Population | 观测等位基因数 Na | 有效等位基因数 Ne | 多样性指数 I | 观测杂合度 Ho | 期望杂合度 He | Nei’s期望杂合度Nei | 多态性信息含量指数PIC |
|---|---|---|---|---|---|---|---|
| 国内群体Domestic | 2.5238±0.5345 | 1.8145±0.4519 | 0.6657±0.2413 | 0.3396±0.3045 | 0.4168±0.1591 | 0.412±0.1574 | 0.4794 |
| 国外群体Abroad | 2.5079±0.5922 | 1.6862±0.4527 | 0.5897±0.2469 | 0.2770±0.2767 | 0.367±0.1664 | 0.3652±0.1655 | 0.4666 |
新窗口打开
Table 4
表4
表4野生种与国内外群体遗传多样性分析
Table 4Estimates of genetic diversity of wild species and populations of broomcorn millet from home and abroad
| 群体 Population | 观测等位基因数Na | 有效等位基因数Ne | 多样性指数 I | 观测杂合度 Ho | 期望杂合度 He | Nei’s期望杂合度Nei | 多态性信息 含量指数PIC |
|---|---|---|---|---|---|---|---|
| 国内栽培群体Domestic | 2.254±0.6468 | 1.6727±0.4612 | 0.5528±0.2877 | 0.2956±0.3439 | 0.3588±0.1962 | 0.352±0.1925 | 0.3919 |
| 国外群体Abroad | 2.5079±0.5922 | 1.6862±0.4527 | 0.5897±0.2469 | 0.277±0.2767 | 0.367±0.1664 | 0.3652±0.1655 | 0.4666 |
| 野生种质Wild germplasm | 2.4194±0.6153 | 1.9285±0.5101 | 0.6948±0.2852 | 0.4124±0.3185 | 0.4507±0.1827 | 0.4373±0.1773 | 0.4702 |
新窗口打开
2.4 遗传相似性分析
采用POPGENE1.32计算不同群体黍稷种质间的遗传相似度(表5),结果显示遗传距离的变化范围为0.0207—0.2948,平均值为0.1030,遗传一致度的变化范围0.7447—0.9796,平均值0.9046。其中,南方群体与国内外群体间遗传距离均达0.2以上,内蒙古高原群体和印度、美国遗传距离也达0.1水平,而遗传距离最小的是黄土高原和西北群体,只有0.0207,遗传距离越大,说明亲缘关系越远,遗传相似性越低,而各群体间遗传距离变化区间只有0.27左右,遗传一致度接近于1,表明各群体间遗传距离大不,亲缘关系都很近,遗传相似性高。Table 5
表5
表510个群体遗传距离与遗传一致度
Table 5Nei’s unbiased measures of genetic identity and genetic distance
| 群体 Population | 黄土高原 Loess Plateau | 西北地区 Northwest | 东北地区 Northeast | 内蒙古高原 Inner Mongolia plateau region | 南方地区 Southern | 蒙古 Mongolia | 印度 India | 美国 U.S.A | 前苏联 Former Soviet Union | 欧洲 Europe |
|---|---|---|---|---|---|---|---|---|---|---|
| 黄土高原Loess Plateau | — | 0.9796 | 0.9667 | 0.9746 | 0.7940 | 0.9521 | 0.9108 | 0.9317 | 0.9121 | 0.9357 |
| 西北地区Northwest | 0.0207 | — | 0.9546 | 0.9562 | 0.8068 | 0.9350 | 0.9073 | 0.9286 | 0.9049 | 0.9074 |
| 东北地区Northeast | 0.0339 | 0.0465 | — | 0.9498 | 0.8135 | 0.9625 | 0.9287 | 0.9386 | 0.9319 | 0.9443 |
| 内蒙古高原 Inner Mongolia plateau region | 0.0257 | 0.0448 | 0.0515 | — | 0.7769 | 0.9100 | 0.8868 | 0.8951 | 0.9151 | 0.9296 |
| 南方地区Southern | 0.2307 | 0.2147 | 0.2064 | 0.2524 | — | 0.7657 | 0.7857 | 0.7734 | 0.7447 | 0.7649 |
| 蒙古Mongolia | 0.0491 | 0.0672 | 0.0382 | 0.0943 | 0.2669 | — | 0.9384 | 0.9364 | 0.9301 | 0.9577 |
| 印度India | 0.0934 | 0.0973 | 0.0740 | 0.1201 | 0.2412 | 0.0636 | — | 0.9465 | 0.9174 | 0.9444 |
| 美国U.S.A | 0.0708 | 0.0740 | 0.0634 | 0.1108 | 0.2569 | 0.0657 | 0.055 | — | 0.9432 | 0.9445 |
| 前苏联Former Soviet Union | 0.092 | 0.0999 | 0.0706 | 0.0887 | 0.2948 | 0.0725 | 0.0862 | 0.0585 | — | 0.9751 |
| 欧洲Europe | 0.0665 | 0.0972 | 0.0573 | 0.073 | 0.2681 | 0.0432 | 0.0573 | 0.0571 | 0.0252 | — |
新窗口打开
2.5 参试群体的UPGMA聚类分析
利用10个参试群体绘制的基于UPGMA法的树状聚类图,从遗传距离频率0.09处可将所有参试群体聚类为3大组群,分别命名为组群Ⅰ、组群Ⅱ和组群Ⅲ(图1)。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1基于SSR标记数据的种质资源群体间聚类分析图
-->Fig. 1Dendrogram generated by UPGMA cluster analysis of 10 populations of the main cultivation areas of broomcorn millet accessions based on data from 63 SSR markers
-->
组群I由来自于国内黄土高原、东北、西北、内蒙古高原生态区和蒙古的材料组成。聚类分析表明,遗传关系与地理分布有关,整个组群I地理分布上紧密连接,由国内黍稷主栽区与接壤的蒙古构成。
组群Ⅱ全部由国外群体组成。前苏联与欧洲群体遗传关系较近,美国和印度群体遗传关系较近,前苏联和欧洲群体地理分布相连,而美国和印度材料遗传关系近,可能因为引种地相似一致和长年驯化的结果。总体上,组群Ⅰ和组群Ⅱ有可能是长期地理隔离引起的生殖隔离造成的。
组群Ⅲ由南方群体构成,南方群体自成一支与国内外群体遗传关系均较远,从地理分布上看,它离黍稷主栽区较远,长期的自然条件、人为影响或许是造成与大部分群体差异较大的原因。但因南方群体的取材较少,地区、品种单一,具体原因有待进一步研究。
从192个参试个体绘制的基于UPGMA法的树状聚类图(图2)来看,所有材料的遗传距离频率在0.06—0.37变化,其中来自东北黑龙江齐齐哈尔的泰来小野糜(34号)在截距0.37处被独立分为一支,来自甘肃的野黍子(19号)在截距0.34处被分为独立个体,表明这两个材料与其他材料遗传差异较大。其余材料在截距0.31处被分为若干个小群体,个体与个体间差异不大,没有分成大的聚类,整体分布与地理分布有一定关联,也存在区域间和国内外间的互相渗透。
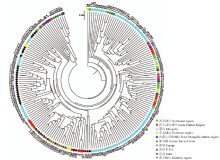 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2基于SSR标记数据的192份材料聚类图
-->Fig. 2Dendrogram generated by UPGMA cluster analysis of 192 broomcorn millet accessions based on data from 63 SSR markers
-->
综上,基于遗传距离频率的UPGMA法构建的树状聚类图与遗传距离、遗传一致度的分析结果,相互印证、吻合一致。
3 讨论
3.1 黍稷遗传多样性
黍稷虽栽培历史久远,但作为小宗作物,种植面积狭小,食用人群较少,所关注程度一直较低,所以研究水平也一直落后于水稻、小麦、玉米主要作物。尽管国家种质库中保存着8 600多份黍稷种质资源,但开展的研究主要集中于形态学水平的遗传多样性评价和抗旱耐盐资源的初步鉴定等方面,而对分子水平的遗传多样性则所知甚少。如胡兴雨等[24]对来自国家种质资源库中的8 016份黍稷种质资源进行了株高、千粒重、生育期等11个农艺性状进行主成分分析和聚类分析;WANG等[25]选择《中国黍稷(糜)品种资源目录》中山西省1 192份黍稷种质,分析了黍稷种质资源穗型与主要农艺性状的关系;LIU等[26]研究了不同耐性黍稷在混合盐胁迫下的生理应答机制并分析了195份黍稷核心种质的耐盐性[27],为后续开展耐盐基因挖掘奠定了基础;王纶等[28]选用山西省有代表性的500份黍稷种质资源采用反复干旱法进行了抗旱性鉴定评价;刘晓欢等[29]研究了黍稷品种间农艺性状的形态解剖差异;张盼盼等[30]对黍稷苗期PEG胁迫下抗旱指标进行了鉴选研究。而在分子水平,黍稷中已有AFLP[16-17]、RAPD[15]和ISSR[18]等标记应用于遗传多样性研究。KARAM等[16-17]检测了黍稷遗传多样性用AFLP银染标记;LÁGLER等[18]选用9个来自草莓属植物的ISSR标记检测了21份匈牙利黍稷;2008年,HUNT等[19]对来源于中国不同生态区的118份黍稷材料选取了水稻、小麦、大麦和燕麦的46个SSR标记进行了遗传变异分析;2010年,CHO等[20]通过构建黍稷基因组DNA的富SSR文库,以此开发了25个多态性微卫星标记。上述研究所用标记,特别是SSR标记的数量较少,很难全面反映黍稷种质资源的多样性,因此从黍稷自身基因组开发SSR标记对促进其遗传多样性的研究具有十分重要的意义。随着测序技术的成熟发展及其价格降低,应用二代测序技术与磁珠富集法相结合大规模的开发了谷子[31]、燕麦[32]、蚕豆[33]和山黧豆[34]等作物的基因组SSR标记,大大促进了相关小宗作物分子水平的研究进展。本研究对来自国内外不同生态区的的192份种质(栽培种和野生种)利用前期开发的黍稷基因组SSR引物进行遗传多样性分析,共检测出161个等位变异,其中每对引物有效等位变异数1.74,有效等位变异所占比重为67.97%。多态性信息量PIC介于0.0877—0.8020,Nei基因多样性指数的变化范围是0.0279—0.6645,Shannon-Weaver指数变化范围为0.0743—1.0953,其数值结果均高于先前研究,基因组SSR标记的成功应用,对黍稷种质资源的遗传多样性有了更全面多样的认识,并有助于黍稷遗传资源的进一步挖掘利用。同时本研究建立的SSR标记在分子水平也为国内黍稷资源研究打下了坚实基础。3.2 黍稷起源、演化与传播
黍稷是人类最早驯化栽培的谷物之一,中国是目前已知的黍稷栽培历史最悠久的国家,研究中国黍稷和国外黍稷的遗传多样性水平,对于探讨黍稷的起源、演化与传播也具有重要的意义。本研究通过对来自黍稷主栽区192份黍稷品种进行SSR遗传多样性的系统分析,揭示了各群体间遗传多样性差异。其中,内蒙古高原、东北、黄土高原群体在有效等位变异、Shannon-Weaver指数和Nei基因多样性指数都高于其他群体,结合其他各参数而言,国内黍稷资源的遗传多样性也大于国外种质资源,同时还发现不同群体黍稷种质间的遗传距离变化范围为0.0207—0.2948,遗传一致度变化范围为0.7447—0.9796,更进一步显示出10个群体来源的黍稷资源群体间的遗传距离不大,亲缘关系都很近,遗传相似性高。通过分析各栽培区的遗传多样性可以为黍稷的起源和演化研究提供依据和思路。在先前研究中,中国、中亚和中欧都被提出为黍和其野生祖先驯化的可能起源中心,但具体位置尚有待于进一步研究确认[35]。而中国黍稷品种资源群体间的显著差异,作为全世界大面积栽培黍稷的国家,栽培环境、自然气候条件千差万别,同时还可能隐含着自然和人工优胜选择、栽培资源与野生资源的互相基因渗透,这也是世界的其他地方不能比拟的。从研究中可知,中国黍稷种质的遗传多样性高于国外品种,拥有更多样多元的遗传多样性,此结果与胡兴雨等[24]的研究结果一致。其中,内蒙古高原群体和东北群体品种的遗传多样性要高于其他地区,该区域有可能是黍稷起源地,并以此向周边扩展,此结果与LI等[22]推得新疆是黍稷驯化和传播的十字路口,是由中国北方通过新疆向中亚、西亚进而进入欧洲的结果一致。同时,蒙古与国内归为一组,俄罗斯与欧洲归为一组的研究互相佐证,表明群体聚类与驯化、传播及地理分布有一定关联。
3.3 特殊群体与特殊个体
黑龙江齐齐哈尔的泰来小野糜(34号)和甘肃的野黍子(19号)在聚类分析中非常独特,远远分离于其他黍稷材料之外。这有可能是由于野生种质没有经过人为驯化不受人为环境影响又或是与其他野生近缘种杂交影响所致。而对于南方种群虽仅有一份材料但在群体聚类中却自成一个独立的群体,与国内外其他各群体遗传距离均达0.2以上,亲缘关系较远,可能与自然环境,所在的气候区有关,真实原因还需利用植物学、细胞生物学和分子生物学等手段对其他性状进行深入研究。同时通过将国内栽培种和野生种分别分析,进一步发现了国内野生种质的丰富遗传多样性与其在黍稷种质资源中的重要地位,此次野生种质只有19份,但其有效等位基因数、香农信息值数、Nei基因多样性指数均远高于其他群体,充分显示其遗传多样性的丰富。所以,野生种质和南方零星种质资源有必要在今后的种质保存、资源管理和基础研究工作中重点关注。
4 结论
中国黍稷种质资源具有较丰富的遗传多样性,普遍高于国外材料,而国内资源又以内蒙古高原、东北地区材料的遗传多样性最高,进一步验证了“中国是黍稷起源中心”的论点。同时应进一步加强野生材料、南方种质的收集和国外资源的引进,丰富现有资源的遗传多样性,并促进黍稷遗传改良、演化传播等研究。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}