Quantitative Trait Locus Mapping of Bruchids Resistance Based on A Novel Genetic Linkage Map in Cowpea (Vigna unguiculata)
WANG Yan,1, FAN BaoJie1, CAO ZhiMin1, ZHANG ZhiXiao1, SU QiuZhu1, WANG Shen1, WANG XueQing2, PENG XiuGuo3, MEI Li4, WU YuHua1, LIU ShaoXing1, TIAN ShengMin1, XU JunJie1, JIANG ChunZhi1, WANG WeiJuan5, LIU ChangYou,1, TIAN Jing,11Institute of Cereal and Oil Crops, Hebei Academy of Agricultural and Forestry Sciences/Hebei Laboratory of Crop Genetic and Breeding, Shijiazhuang 050035 2Hebei Academy of Agricultural and Forestry Sciences, Shijiazhuang 050031 3Shijiazhuang Botanical Garden, Shijiazhuang 050000 4Beijing Agricultural Technology Extension Centre, Beijing 100029 5Plant Protection Department of Chongli District, Zhangjiakou 076350, Hebei
Abstract 【Objective】Bruchids are the main storage pests of cowpea. The discovery of bruchids resistance genes is helpful for breeding resistant varieties and reducing the harm of bruchids to cowpea production.【Method】In this study, a RIL population of 282 lines derived from the cross between Zhongjiang No.1 (a bruchid-susceptible cultivar) and Pant-lobia-1 (a bruchid-resistant cultivar) was used for phenotype identification of bruchids resistance by artificial inoculation of Callosobruchus chinensis and Callosobruchus maculates. The two parents were used to screen polymorphic markers from 3 992 SSR markers of mung bean, adzuki bean and cowpea. Genotypes of 282 lines were analyzed using the polymorphic SSR markers. Based on the phenotype identification and genotype analysis, a genetic linkage map was constructed and the bruchids resistance gene(s) of cowpea was located using Inclusive Composite Interval Mapping (ICIM-ADD). 【Result】The results showed that Zhongjiang No. 1 and F1 seeds were 100% susceptible to Callosobruchus chinensis and Callosobruchus maculates, and the damage rates of Pant-lobia-1 were 22.5% and 42.5%, respectively. It was speculated that the resistance of Pant-lobia-1 to bruchids was recessive inheritance. 182 polymorphic markers were obtained from 3 992 SSR markers of mung bean, adzuki bean and cowpea. Using those polymorphic SSR markers, a genetic linkage map with 11 linkage groups was constructed. The map covered a total length of 1 065.23 cM with an average interval of 5.85 cM between adjacent markers. Two bruchid-resistant QTLs from linkage groups 1 and 5 were discovered, which were temporarily named as vubr1-1 and vubr5-1. QTL vuvr1-1 was located between markers XD11-44 and HAAS_VR_2274, which genetic distance was 7.6 cM, explaining 7.16% and 6.92% of the phenotypic variation in the two bruchid-resistant tests respectively. QTL vubr5-1 was located between markers XD1-14 and CP185, which genetic distance was 2.9 cM, explaining 6.96% and 6.37% of the phenotypic variation in the two bruchid-resistant tests respectively. 【Conclusion】A genetic linkage map containing 11 linkage groups and 182 polymorphic markers was constructed. Two QTLs linked with bruchids resistance were identified on linkage groups 1 and 5. Keywords:Vigna unguiculata;SSR;bruchid resistance;genetic mapping;quantitative trait locus
PDF (1724KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 王彦, 范保杰, 曹志敏, 张志肖, 苏秋竹, 王珅, 王学清, 彭秀国, 梅丽, 武玉华, 刘少兴, 田胜民, 徐俊杰, 蒋春志, 王伟娟, 刘长友, 田静. 基于新遗传连锁图谱的豇豆抗豆象QTL定位. 中国农业科学, 2021, 54(22): 4740-4749 doi:10.3864/j.issn.0578-1752.2021.22.003 WANG Yan, FAN BaoJie, CAO ZhiMin, ZHANG ZhiXiao, SU QiuZhu, WANG Shen, WANG XueQing, PENG XiuGuo, MEI Li, WU YuHua, LIU ShaoXing, TIAN ShengMin, XU JunJie, JIANG ChunZhi, WANG WeiJuan, LIU ChangYou, TIAN Jing. Quantitative Trait Locus Mapping of Bruchids Resistance Based on A Novel Genetic Linkage Map in Cowpea (Vigna unguiculata). Scientia Acricultura Sinica, 2021, 54(22): 4740-4749 doi:10.3864/j.issn.0578-1752.2021.22.003
开放科学(资源服务)标识码(OSID):
0 引言
【研究意义】豆象是豇豆(Vigna unguiculata (Linn.) Walp.)等食用豆类作物的主要仓储害虫,以幼虫蛀入籽粒内进行危害[1],导致豇豆品质和质量下降,发生严重时可造成整仓库的豇豆籽粒全部受损,严重影响豇豆产业的发展[2,3]。在中国危害豇豆的豆象主要是绿豆象(Callosobruchus chinensis)和四纹豆象(Callosobruchus maculates)[4,5]。其中,四纹豆象是中国进境检疫三类危险性害虫之一。目前,国内外主要通过化学防治和生物防治方法防治豆象危害[6,7,8]。化学防治方法容易导致农药残留和环境污染,而生物防治方法成本高、见效慢[9,10]。因此,利用作物本身的抗性培育抗豆象品种是防治豆象最为经济有效的方法。【前人研究进展】目前,国内外筛选到的豇豆抗豆象资源较少,1985年国际热带农业研究所(International Institute of Tropical Agriculture,IITA)筛选了8 000多份豇豆资源,仅得到3份抗性资源,分别为TVU11952、TVU11953和TVU20273抗源[11,12]。近年来,随着分子生物学技术的快速发展,国内外关于豇豆QTL定位的研究不断增多。2015年HUYNH等[13]利用1 536个SNP标记定位了2个与蚜虫抗性相关的QTL位点,分别位于第1连锁群和第7连锁群。2018年LO等[14]利用17 739个多态性标记构建了一个高密度遗传连锁图谱,并定位了9个与豇豆驯化相关的QTL位点。但是关于豇豆抗豆象性状遗传研究较少。1983年REDDEN[15]研究认为TVU2027的抗豆象特性是由1个主要隐性基因及其修饰基因起作用。2005年SINGH[16]通过研究TVU11952、TVU11953和TVU2027的抗性遗传,认为它们的抗豆象基因相同,均由2个隐性基因控制,并命名为rm1和rm2。2018年MIESHO等[17]利用41 948个SNP标记对217份豇豆核心种质进行全基因组关联分析(genome-wide association studies,GWAS),筛选到6个与抗豆象相关的候选基因,分别位于豇豆第1、2、6和8染色体。【本研究切入点】目前,国外关于豇豆抗豆象分子遗传基础研究结果不一,而国内关于豇豆抗豆象遗传研究鲜见报道。【拟解决的关键问题】本研究利用来自尼尔利亚的抗豆象豇豆品种Pant-lobia-1作为抗豆象基因来源,与中国豇豆品种中豇1号杂交构建重组自交系(recombinant inbred lines,RIL)群体,进行豇豆遗传连锁图谱构建和抗豆象QTL定位,为豇豆抗豆象分子标记辅助育种和抗豆象基因克隆提供依据。
*、**和***分别表示该标记在0.05、0.01和0.001显著水平上表现偏分离 Fig. 4A genetic linkage map of cowpea
*, **, and ***: Indicated markers exhibiting significant deviation from the expected segregation ratio at the 0.05, 0.01 and 0.001 levels, respectively
JIN WL, TAN RJ, WANG JZ, ZHANG ZY, LIU CA, PU SJ, ZHAOB. Preliminary analysis on spatial distribution patterns of Callosobruchus chinensis eggs in adzuki bean fields Plant Protection, 2004, 30(6): 34-36. (in Chinese) [本文引用: 1]
LIU CY, TIANJ, FAN BJ, CAO ZM, SU QZ, ZHANG ZX, WANG SH. Advances in breeding research on bruchid-resistant cultivars of three main Vigna food legumes Scientia Agricultura Sinica, 2010, 43(12): 2410-2417. (in Chinese) [本文引用: 1]
GUJAR GT, YADAV TD. Feeding of Callosobruchus maculatuts (Fab.) and Callosobruchus chinensis (Linn.) in green gram Indian Journal of Entomology, 1978, 40(2): 108-112. [本文引用: 1]
BABARINDE SA, PITANO, AJALA MO, OLATUNDE GO. Insectifugal and insecticidal potentials of two tropical botanical essential oils against cowpea seed bruchid Environmental Science and Pollution Research, 2017, 24(24): 19785-19794. DOI:10.1007/s11356-017-9589-xURL [本文引用: 1]
BABARINDE SA, PITANO, OLATUNDE GO, AJALA MO. Chemical composition of the essential oil of nigeria grown hoslundia opposita Vahl (Lamiaceae) dried leaves and its bioactivity against cowpea seed bruchid Chemistry & Biodiversity, 2017, 14(6): e1600418. [本文引用: 1]
SANGW, SPEAKMONM, ZHOUL, WANGY, LEIC, PILLAI SD, ZHU-SALZMANK. Detrimental effects of electron beam irradiation on the cowpea bruchid Callosobruchus maculatus Pest Management Science, 2016, 72(4): 787-795. DOI:10.1002/ps.2016.72.issue-4URL [本文引用: 1]
AMEVOINK, SANONA, APOSSABAM, GLITHO IA. Biological control of bruchids infesting cowpea by the introduction of dinarmus basalis (Rondani) (Hymenoptera: Pteromalidae) adults into farmers' stores in West Africa Journal of Stored Products Research, 2007, 43(3): 240-247. DOI:10.1016/j.jspr.2006.06.004URL [本文引用: 1]
AMUSA OD, OGUNKANMI LA, ADETUNBI JA, AKINYOSOYE ST, OGUNDIPE OT. Assessment of bruchid (Callosobruchus maculatus) tolerance of some elite cowpea (Vigna unguiculata) varieties Journal of Agriculture & Sustainability, 2014, 6(2): 164-178. [本文引用: 1]
HUYNH BL, EHLERS JD, NDEVEA, WANAMAKERS, LUCAS MR, CLOSE TJ, ROBERTS PA. Genetic mapping and legume synteny of aphid resistance in African cowpea (Vigna unguiculata L. Walp.) grown in California Molecular Breeding, 2015, 35(1): 36. DOI:10.1007/s11032-015-0254-0URL [本文引用: 2]
LOS, Muñoz-AmatriaínM, BOUKARO, HERNITERI, CLOSE TJ. Identification of QTL controlling domestication-related traits in cowpea (Vigna unguiculata L. Walp) Scientific Reports, 2018, 8(1): 6261. DOI:10.1038/s41598-018-24349-4URL [本文引用: 1]
REDDENR. The inheritance of seed resistance to Callosobruchus maculatus F. in cowpea (Vigna unguiculata L. Walp.): II. Analyses of percentage emergence and emergence periods of bruchids in F4 seed generation of two reciprocal crosses Australian Journal of Agricultural Research, 1983, 34(6): 697-705. DOI:10.1071/AR9830697URL [本文引用: 2]
MIESHOB, HAILAYM, MSISKAU, BRUNOA, KYAMANYWAS. Identification of candidate genes associated with resistance to bruchid (Callosobruchus maculatus) in cowpea Plant Breeding, 2019, 138(5): 1-9. DOI:10.1111/pbr.2019.138.issue-1URL [本文引用: 3]
AFANADOR LK, HALEY SD, KELLY JD. Adoption of a mini-prep DNA extraction method for RAPD marker analysis in common bean Bean Improvement Cooperative, 1993, 36: 10-11. [本文引用: 1]
LIUC, FANB, CAOZ, SUQ, WANGY, ZHANGZ, WUJ, TIANJ. A deep sequencing analysis of transcriptomes and the development of EST-SSR markers in mungbean (Vigna radiata) Journal of Genetics, 2016, 95(3): 1-9. [本文引用: 2]
CHENX, LAUDEMAN TW, RUSHTON PJ, SPRAGGINS TA, TIMKO MP. CGKB: An annotation knowledge base for cowpea (Vigna unguiculata L.) methylation filtered genomic genespace sequences BMC Bioinformatics, 2007, 8(1): 129. DOI:10.1186/1471-2105-8-129URL [本文引用: 1]
MENGL, LIH, ZHANGL, WANGJ. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations The Crop Journal, 2015, 3(3): 269-283. DOI:10.1016/j.cj.2015.01.001URL [本文引用: 1]
LIH, RIBAUT JM, LIZ, WANGJ. Inclusive composite interval mapping (ICIM) for digenic epistasis of quantitative traits in biparental populations Theoretical & Applied Genetics, 2008, 116(2): 243-260. [本文引用: 1]
IWATAA, GREENLAND CM, JACKSON SA. Cytogenetics of legumes in the phaseoloid clade Plant Genome, 2013, 6(3): 1-8. [本文引用: 1]
WANG LX, CHENG XZ, WANG SH, LIU CY, LIANGH. Transferability of SSR from adzuki bean to mung bean Acta Agronomica Sinica, 2009, 35(5): 816-820. (in Chinese) [本文引用: 1]
ZHONGM, CHENG XZ, WANG LX, WANG SH, WANG XB. Transferability of mungbean genomic-SSR markers in other Vigna species Acta Agronomica Sinica, 2012, 38(2): 223-230. (in Chinese) DOI:10.3724/SP.J.1006.2012.00223URL [本文引用: 3]
FATOKUN CA, MENANCIO-HAUTEAD I, DANESHD, YOUNGN D. Evidence for orthologous seed weight genes in cowpea and mung bean based on RFLP mapping Genetics, 1992, 132(3): 841-846. DOI:10.1093/genetics/132.3.841URL [本文引用: 1]
MUÑOZ-AMATRIAÍNM, MIREBRAHIMH, XUP, WANAMAKER SI, LUO MC, ALHAKAMIH, ALPERTM, ATOKPLEI, BATIENO BJ, BOUKARO. Genome resources for climate-resilient cowpea, an essential crop for food security The Plant Journal, 2017, 89(5): 1042-1054. DOI:10.1111/tpj.2017.89.issue-5URL [本文引用: 2]
OHLSON EW, THIO GI, MAHAMADOUS, PACOS, TIMKO MP. Quantitative trait loci analysis of brown blotch resistance in cowpea variety KN1 Molecular Breeding, 2018, 38(9): 110. DOI:10.1007/s11032-018-0867-1URL [本文引用: 2]
NDEVE AD, SANTOSJ, MATTHEWS WC, HUYNH BL, GUO YN, LOS, MUOZ-AMATRIA N M, ROBERTSP A. A novel root-knot nematode resistance QTL on chromosome Vu01 in cowpea Genes Genomes Genetics, 2018, 9: 1199-1209. [本文引用: 2]
KITAMURAK, ISHIMOTOM, SAWAM. Inheritance of resistance to infestation with adzuki bean weevil in Vigna sublobata and vuccessful incorporation to V. radiata Japanese Journal of Breeding, 2008, 38(4): 459-464. DOI:10.1270/jsbbs1951.38.459URL [本文引用: 1]
CHEN HM, KU HM, SCHAFLEITNERR, BAINS TS, KUO CG, LIU CA, NAIR RM. The major quantitative trait locus for mungbean yellow mosaic Indian virus resistance is tightly linked in repulsion phase to the major bruchid resistance locus in a cross between mungbean [Vigna radiata (L.) Wilczek] and its wild relative Vigna radiata ss Euphytica, 2013, 192(2): 205-216. DOI:10.1007/s10681-012-0831-9URL [本文引用: 1]
WU CS. Development of SSR markers and construction of a genetic linkage map in mungbean (Vigna radiate L.) [D]. Lanzhou: Gansu Agricultural University, 2014. (in Chinese) [本文引用: 1]
SUNL, CHENG XZ, WANG SH, WANG LX, LIU CY, MEIL, XUN. Heredity analysis and gene mapping of bruchid resistance of a mungbean cultivar V2709 Scientia Agricultura Sinica, 2008, 41(5): 1291-1296. (in Chinese) [本文引用: 1]
LIU CY, SU QZ, FAN BJ, CAO ZM, ZHANG ZX, WUJ, CHENG XZ, TIANJ. Genetic mapping of bruchid resistance gene in mungbean V1128 Acta Agronomica Sinica, 2018, 44(12): 1875-1881. (in Chinese) DOI:10.3724/SP.J.1006.2018.01875URL [本文引用: 1]
 ,1, 范保杰1, 曹志敏1, 张志肖1, 苏秋竹1, 王珅1, 王学清2, 彭秀国3, 梅丽4, 武玉华1, 刘少兴1, 田胜民1, 徐俊杰1, 蒋春志1, 王伟娟5, 刘长友
,1, 范保杰1, 曹志敏1, 张志肖1, 苏秋竹1, 王珅1, 王学清2, 彭秀国3, 梅丽4, 武玉华1, 刘少兴1, 田胜民1, 徐俊杰1, 蒋春志1, 王伟娟5, 刘长友

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT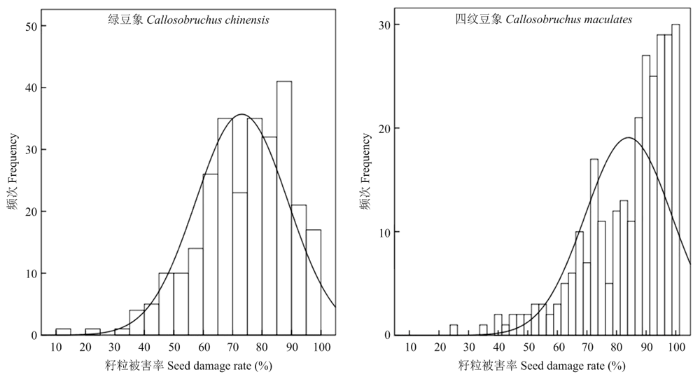 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT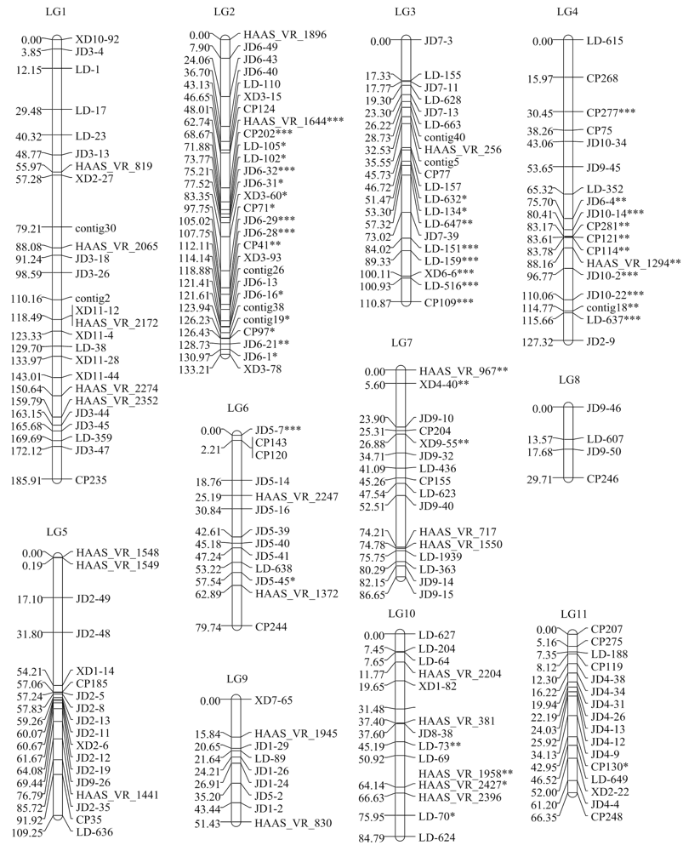 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT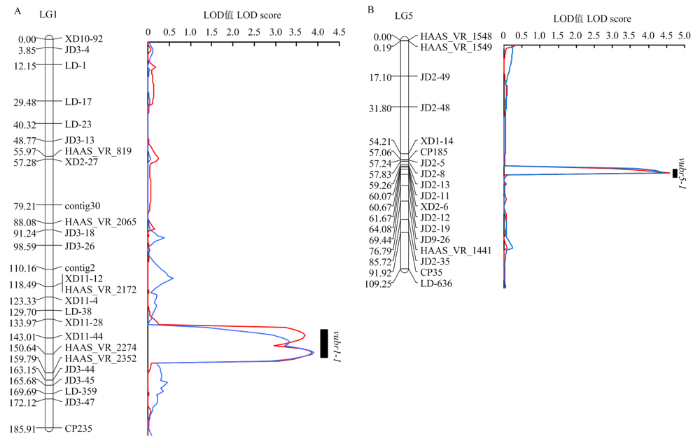 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}