0 引言
【研究意义】硫代葡萄糖苷(glucosinolatcs,简称硫苷)是一类含氮、含硫的植物次生代谢产物[1],广泛存在于食用油料作物、蔬菜和拟南芥等十字花科植物中,是该科植物产生的一类重要次生代谢产物[2-3],其降解产物具有多种生物学功能[4-5]。例如,硫苷降解产物异硫氰酸酯类(isothiocyanates,ITCs)、硫氰酸酯类(thiocyanates,TCs)、腈类(nitrilcs)、恶唑烷酮类(oxazolidinones,ODs)等具有较强的防腐和抗菌作用,可作为杀虫剂、杀菌剂、抗微生物制剂和天然的生物熏蒸消毒剂[6-7];硫苷具有芳香或刺激性气味,既可作为十字花科专食性昆虫(crucifer specialists)的引诱剂,刺激或抑制某些昆虫产卵与摄食[8],又可作为杂食性昆虫(generalist herbivores)的驱散剂[9];同时硫苷还赋予食物特殊的风味,如萝卜、甘蓝等的辛辣味[10];萝卜硫素则被证明是迄今发现的抗癌活性最强的植物活性物质,可预防前列腺癌、肺癌、食道癌等[10-11]。可见,硫苷及其降解产物对植物抗性和人类健康具有重要作用。拟南芥是重要的十字花科模式植物,相对于其他植物,具有较为完整的基因组信息和丰富的表达谱芯片数据资源。因此,在拟南芥中对硫苷的动态合成规律展开系统研究,不仅具有重要的理论研究意义,还可以为进一步在油菜、白菜等作物中开展相关研究提供重要的参考。【前人研究进展】硫苷类化合物均由β-D-硫葡萄糖基、硫化肟基团以及来源于氨基酸的侧链R基组成。通常根据侧链R基的来源将硫苷分为脂肪族硫苷、吲哚族硫苷和芳香族硫苷三大类[12]。目前硫苷生物合成途径较清楚,主要酶基因均已被证实[13]。硫苷的生物合成途径大致可以分为前体氨基酸侧链延长、核心结构合成和次级修饰3个主要阶段[14]。其中,核心结构合成过程是所有硫苷合成的共同部分。核心结构形成后再经羧基化、甲基化、葡萄糖基化等次级修饰形成不同的硫苷类化合物。硫苷侧链修饰具有重要的作用,硫苷及其降解产物的理化性质很大程度上取决于硫苷的侧链结构[15]。近年来,硫苷调控机理方面的研究也取得了一定的进展,bHLH、Dof和MYB类转录因子均被证实参与调控硫苷的生物合成[15-19]。其中,MYB类转录因子对硫苷的生物合成尤为重要[15-16,20-22]。【本研究切入点】虽然硫苷的生物合成和调控途径研究较清楚,但是对其合成规律、生物学功能仍缺乏系统的研究。鉴于硫苷的含量在不同材料和组织、不同发育时期和环境下均不同[3,23],因此,研究硫苷合成相关基因的表达模式的动态变化规律,将有助于揭示硫苷的生物合成和代谢规律、筛选控制硫苷合成或调控的关键基因。【拟解决的关键问题】本研究基于AtGenExpress[24-25]和PLEXdb[26]中的表达谱数据,结合Real-time PCR技术,在全基因组水平对拟南芥硫苷生物合成途径主要酶基因和调控基因的时空表达特性和胁迫诱导表达谱进行了系统的研究,以解析不同硫苷组分的生物合成规律和胁迫诱导表达模式。1 材料与方法
1.1 硫苷生物合成相关基因的组织表达特性分析
本研究分析的硫苷生物合成相关基因如表1所示。拟南芥组织表达谱芯片数据来源于AtGenExpress (http://www.weigelworld.org/resources/microarray/AtGenExpress),共包含不同发育时期的79个组织或器官[24]。均一化的芯片数据利用Cluster v3.0软件[27]采用层级聚类法分析基因的表达谱,并用Java TreeView软件[27]图形化基因表达谱热力图(heatmaps)。拟南芥哥伦比亚野生型(Arabidopsis thaliana ecotype Columbia-0)种子购自于ARBC(拟南芥种质资源中心),由重庆市油菜工程研究中心实验室保存。2016年1月进行组培室种植,培养条件为16 h光照/8 h黑暗,昼夜温度22℃/18℃,相对湿度70%。分别于苗期和花期取植株的根、茎、叶、花和种子等组织,液氮中速冻,-80℃保存备用。1.2 硫苷生物合成相关基因的胁迫诱导表达谱分析
拟南芥生物(病原菌和虫害)和非生物(激素和环境因子)胁迫诱导的基因芯片数据来自于AtGenExpress[25]和植物表达谱数据库(plant expression database, PLEXdb)[26]。所采用的9组芯片数据的编号分别为GSE5525(AT49)、GSE5640(AT50)、GSE5684(AT51)、AT59、GSE6516(AT63)、GSE17193 (AT84)、GSE18960(AT90)、GSE20188(AT110)和GSE24552(AT112)。病原菌和虫害芯片数据的试验处理详见PLEXdb数据库中相关芯片数据的试验设计(http://www.plexdb.org/plex.php?database= Arabidopsis)[26,28-30];激素和环境因子胁迫诱导芯片数据的实验处理详见AtGenExpress[24-25]。所有芯片数据均为已进行标准化处理的数据。Table 1
表1
表1拟南芥硫苷生物合成相关基因
Table 1Glucosinolate biosynthetic genes in Arabidopsis
| 全称 Full name | 简称 Abbreviation | 基因组编号 Genome number |
|---|---|---|
| 脂肪族硫苷合成 Aliphatic glucosinolates | ||
| 侧链延伸 Side-chain elongation | ||
| 支链氨基酸氨基转移酶3 Branched-chain amino acid aminotransferase 3 | BCAT3-AGSL | AT3G49680 |
| 支链氨基酸氨基转移酶4 Branched-chain amino acid aminotransferase 4 | BCAT4-AGSL | AT3G19710 |
| 胆汁酸转运蛋白5 Bile acid transporter 5 | BAT5-AGSL | AT4G12030 |
| 甲硫烷基苹果酸异构酶1 Methylthioalkylmalate synthase 1 | MAM1-AGSL | AT5G23010 |
| 甲硫烷基苹果酸异构酶3 Methylthioalkylmalate synthase 3 | MAM3-AGSL | AT5G23020 |
| 异丙基苹果酸异构酶大亚基1 Isopropylmalate isomerase large subunit 1 | LeuC1-AGSL | AT4G13430 |
| 异丙基苹果酸异构酶小亚基1 Isopropylmalate isomerase small subunit 1 | LeuD1-AGSL | AT2G43100 |
| 异丙基苹果酸异构酶小亚基2 Isopropylmalate isomerase small subunit 2 | LeuD2-AGSL | AT3G58990 |
| 异丙基苹果酸脱氢酶1 Isopropylmalate dehydrogenase 1 | IPMDH1-AGSL | AT5G14200 |
| 异丙基苹果酸脱氢酶2 Isopropylmalate dehydrogenase 2 | IPMDH2-AGSL | AT1G80560 |
| 异丙基苹果酸脱氢酶3 Isopropylmalate dehydrogenase 3 | IPMDH3-AGSL | AT1G31180 |
| 核心结构合成 Core structure | ||
| 细胞色素P450同族物CYP79F1 Cytochrome P450 79F1 | CYP79F1-AGSL | AT1G16410 |
| 细胞色素P450同族物CYP79F2 Cytochrome P450 79F2 | CYP79F2-AGSL | AT1G16400 |
| 细胞色素P450同族物CYP83A1 Cytochrome P450 81A1 | CYP83A1-AGSL | AT4G13770 |
| 谷胱苷肽硫转移酶 F11 Glutathione S-transferase F11 | GSTF11-AGSL | AT3G03190 |
| 谷胱苷肽硫转移酶 TAU20 Glutathione S-transferase TAU 20 | GSTU20-AGSL | AT1G78370 |
| 葡糖基转移酶 74C1 UDP-glucosyl transferase 74C1 | UGT74C1-AGSL | AT2G31790 |
| 脱硫硫苷磺基转移酶5b Sulfotransferase 5b | ST5b-AGSL | AT1G74090 |
| 脱硫硫苷磺基转移酶5c Sulfotransferase 5c | ST5c-AGSL | AT1G18590 |
| 次级结构修饰 Secondary modification | ||
| 黄素单氧化酶1 Flavin-monooxygenase glucosinolate S-oxygenase 1 | FMOGS-OX1-AGSL | AT1G65860 |
| 黄素单氧化酶2 Flavin-monooxygenase glucosinolate S-oxygenase 2 | FMOGS-OX2-AGSL | AT1G62540 |
| 黄素单氧化酶3 Flavin-monooxygenase glucosinolate S-oxygenase 3 | FMOGS-OX3-AGSL | AT1G62560 |
| 黄素单氧化酶4 Flavin-monooxygenase glucosinolate S-oxygenase 4 | FMOGS-OX4-AGSL | AT1G62570 |
| 黄素单氧化酶5 Flavin-monooxygenase glucosinolate S-oxygenase 5 | FMOGS-OX5-AGSL | AT1G12140 |
| 2-含氧戊二酸加氧酶2 Alkenyl hydroxalkyl producing 2 | AOP2-AGSL | AT4G03060 |
| 2-含氧戊二酸加氧酶3 Alkenyl hydroxalkyl producing 3 | AOP3-AGSL | AT4G03050 |
| 2-含氧戊二酸盐加双氧酶 2-Oxoglutarate-dependent dioxygenase | GS-OH-AGSL | AT2G25450 |
| 转录因子 Transcription factor | ||
| MYB28转录因子 Myb family transcription factor 28 | MYB28-AGSL | AT5G61420 |
| MYB29转录因子 Myb family transcription factor 29 | MYB29-AGSL | AT5G07690 |
| MYB76转录因子 Myb family transcription factor 76 | MYB76-AGSL | AT5G07700 |
| 吲哚和苄基硫苷合成 Indolic and benzylic glucosinolates | ||
| 色氨酸合成酶β亚基1 Tryptophan synthase beta subunit 1 | TSB1-IGSL | AT5G54810 |
| 邻氨基苯甲酸合酶α亚基1 A nthranilate synthase, alpha subunit, component I-1 | ASA1-IGSL | AT5G05730 |
| 核心结构合成 Core structure | ||
| 细胞色素P450同族物CYP79A2 Cytochrome P450 79A2 | CYP79A2-IGSL | AT5G05260 |
| 细胞色素P450同族物CYP79B2 Cytochrome P450 79B2 | CYP79B2-IGSL | AT4G39950 |
| 细胞色素P450同族物CYP79B3 Cytochrome P450 79B3 | CYP79B3-IGSL | AT2G22330 |
| 细胞色素P450同族物CYP83B1 Cytochrome P450 83B1 | CYP83B1-IGSL | AT4G31500 |
| 谷胱苷肽硫转移酶 F9 Glutathione S-transferase F9 | GSTF9-IGSL | AT2G30860 |
| 谷胱苷肽硫转移酶 F10 Glutathione S-transferase F10 | GSTF10-IGSL | AT2G30870 |
| 葡糖基转移酶 74B1 UDP-glucosyl transferase 74B1 | UGT74B1-IGSL | AT1G24100 |
| 脱硫硫苷磺基转移酶5a Sulfotransferase 5a | ST5a-IGSL | AT1G74100 |
| 次级结构修饰 Secondary modification | ||
| 细胞色素P450同族物CYP81F1 Cytochrome P450 81F1 | CYP81F1-IGSL | AT4G37430 |
| 细胞色素P450同族物CYP81F2 Cytochrome P450 81F2 | CYP81F2-IGSL | AT5G57220 |
| 细胞色素P450同族物CYP81F3 Cytochrome P450 81F3 | CYP81F3-IGSL | AT4G37400 |
| 细胞色素P450同族物CYP81F4 Cytochrome P450 81F4 | CYP81F4-IGSL | AT4G37410 |
| 吲哚硫苷甲基转移酶1 Indole glucosinolate methyltransferase 1 | IGMT1-IGSL | AT1G21100 |
| 吲哚硫苷甲基转移酶2 Indole glucosinolate methyltransferase 2 | IGMT2-IGSL | AT1G21120 |
| 转录因子 Transcription factor | ||
| MYB34转录因子 Myb transcription factor 34 | MYB34-IGSL | AT5G60890 |
| MYB51转录因子 Myb transcription factor 51 | MYB51-IGSL | AT1G18570 |
| MYB122转录因子 Myb transcription factor 122 | MYB122-IGSL | AT1G74080 |
| bHLH转录因子 bHLH | MYC4 | AT4G17880 |
| bHLH转录因子 bHLH | FAMA | AT3G24140 |
| bHLH转录因子 bHLH | bHLH05 | AT5G46760 |
| Dof转录因子 Dof | Dof1.1 | AT1G07640 |
| 钙调素结合蛋白1 Calmodulin-binding transcription factor | IQD1 | AT3G09710 |
| 异染色质蛋白1 Terminal flower 2 | TFL2 | AT5G17690 |
| 核心结构合成 Core structure | ||
| γ-谷氨酰多肽合成酶1 Gamma-glutamyl peptidase 1 | GGP1 | AT4G30530 |
| C-S裂解酶1 Supper root 1 | SUR1 | AT2G20610 |
| 硫苷降解途径 Glucosinolate (GSL) Breakdown | ||
| β-葡萄糖硫苷酶1β-thioglucoside glucohydrolase 1 | TGG1-AGSL | AT5G26000 |
| β-葡萄糖硫苷酶2β-thioglucoside glucohydrolase 2 | TGG2-AGSL | AT5G25980 |
| β-葡萄糖硫苷酶4β-thioglucoside glucohydrolase 4 | TGG4-AGSL | AT1G47600 |
| β-葡萄糖硫苷酶5 β-thioglucoside glucohydrolase 5 | TGG5-AGSL | AT1G51470 |
| β-O-葡萄糖基水解酶2 Penetration 2 | PEN2-IGSL | AT2G44490 |
| 黑芥子酶结合蛋白1 Myrosinase binding protein | MBP1 | AT1G52040 |
| 表皮特异硫蛋白 epithiospecifier protein | ESP | AT1G54040 |
| 腈特异硫蛋白1 nitrile-specifier protein 1 | NSP1 | AT3G16400 |
| 腈特异硫蛋白2 nitrile-specifier protein 2 | NSP2 | AT2G33070 |
| 腈特异硫蛋白3 nitrile-specifier protein 3 | NSP3 | AT3G16390 |
| 腈特异硫蛋白5 nitrile-specifier protein 5 | NSP5 | AT5G48180 |
| 腈水解酶1 Thaliana nitrilase 1 | NIT1 | AT3G44310 |
| β-O-葡萄糖基水解酶3 Penetration 3 | PEN3 | AT1G59870 |
| 蛋白转运的黑芥子酶相关蛋白1 Modified vacuole phenotype 1 | MVP1 | AT1G54030 |
| 表皮特异硫调控蛋白1 Epithiospecifier modifier 1 | ESM1 | AT3G14210 |
| 黑芥子酶β-glucosidase/myrosinase | PYK10 | AT3G09260 |
| 共同底物途径 Co-substrate pathways | ||
| 腺苷- 5'-磷酰硫酸激酶1 APS-kinase | APK1 | AT2G14750 |
| 腺苷- 5'-磷酰硫酸激酶2 APS-kinase | APK2 | AT4G39940 |
| 谷氨酸-半胱氨酸连接酶 Glutamate-cysteine ligase | GSH1,RML1 | AT4G23100 |
| β-羟异丁酰辅酶A水解酶1 Beta-hydroxyisobutyryl-CoA hydrolase 1 | CHY1 | AT5G65940 |
| 醛氧化酶4 Aldehyde oxidase 4 | AAO4 | AT1G04580 |
| 苯甲酰基氧化酶硫苷1 Benzoyloxyglucosinolate 1 | BZO1 | AT1G65880 |
| 腺苷- 5'-磷酰硫酸还原酶1 APS reductase 1 | APR-1 | AT4G04610 |
| 腺苷- 5'-磷酰硫酸还原酶2 APS reductase 2 | APR-2 | AT1G62180 |
| 腺苷- 5'-磷酰硫酸还原酶3 APS reductase 3 | APR-3 | AT4G21990 |
| ATP 硫酸化酶1 ATP sulfurylase 1 | ATPS1 | AT3G22890 |
| ATP 硫酸化酶3 ATP sulfurylase 3 | ATPS3 | AT4G14680 |
新窗口打开
根据拟南芥芯片信息,共找到78个探针,分别对应78个基因。由于高度同源,有4个探针对应2个基因(IPMDH1和IPMDH3,CYP79F1和CYP79F2),有2个基因(TSB1和NIT1)没有找到对应的探针。芯片数据用Cluster v3.0软件进行层级聚类法分析相关基因的表达谱[27],并利用Java TreeView软件可视化相应的表达谱热力图(heatmaps)[31]。利用STRING v10(http://string-db.org/newstring_cgi/show_input_page.pl? UserId=JnIkwsVs3Jb_&sessionId=U_KvLy4CMUs4)[32]软件分析硫苷生物合成相关基因的共表达和互作调控网络图。
1.3 试剂、药品和仪器
植物RNApure超纯总RNA快速提取试剂盒购自于北京博迈德基因技术有限公司;RQ1 RNase-Free DNase购于美国Promega公司;反转录试剂盒(PrimeScript 1st Strand cDNA Synthesis Kit)购于TaKaRa公司;实时荧光定量PCR试剂盒(SsoAdvanced Universal超混合液)、耗材和仪器(CFX96 Touch)均购于BIO-RAD公司。1.4 实时荧光定量PCR分析
按照RNApure超纯总RNA快速提取试剂盒的操作说明分别提取野生型拟南芥根、茎、叶、花、角果和成熟种子的总RNA;经DNaseⅠ(RQ1)处理去除总RNA中残留的痕量基因组DNA;根据TaKaRa公司的反转录试剂盒说明书进行第一链cDNA的合成。使用SsoAdvanced Universal Mix(SYBR Green)试剂盒和BIO-RAD公司的CFX96 Touch荧光定量PCR仪进行Real-time PCR(qPCR)扩增。以反转录获得的cDNA为模板,以拟南芥Actin 7(AT5G09810.1)为内标基因,分别用10个硫苷合成相关基因的特异引物(表2)进行qPCR扩增。采用2步法反应程序,95℃ 5 min;95℃ 15 s,58℃ 20 s,40个循环;绘制熔解曲线。设3次重复,取平均值,采用2-ΔΔCt法计算基因在不同样品中的相对表达量。Table 2
表2
表2拟南芥硫苷生物合成途径10个代表基因的qPCR引物
Table 2Primers of 10 representive genes involved in glucosinalate biosynthesis pathway in Arabidopsis
| 引物编号 Number of primer | 上游引物序列 Forward primer sequence (5°-3°) | 下游引物序列 Reverse primer sequence (5°-3°) |
|---|---|---|
| MYB28 | GGAGGAGATAACCACAATGAGGA | CATCCCTCGAAGTTTCCTACCA |
| MYB34 | CTCCGATATTTACACGACTGTCGAT | CATATTGTCATCTTCGTTCCAGGA |
| CYP83A1 | AGTGAGCTTATGTTGACCTTCACG | CGCTTTGAGTCCCATAAAGAATC |
| CYP83B1 | TGAGATGAGGAAGATGTGTATGGTG | TTAGATCAACGGTGCCTGATTG |
| IGMT1 | TTCCTATGGTTCTCAAAGCCG | TGCTATTTCATAGGGTGAGAGGAAT |
| AOP2 | ACCGAGAAAAAGAGGACAAGATACA | AATTTTGCGACCAGCTTCTGA |
| MAM1 | CTCCGCCTGACCCATTCG | ACATAGTTTCCGTCGGGAAGC |
| ASA1 | CCCTTCTGTTGCTTCTCGTTATC | CATCTGCCAACTTCTTTGTATCAC |
| CYP79F1 | AAGCGGATAATCTCATAGCTTACG | TAGTCTTCCATCATCAGAAAACACG |
| CYP79B2 | GTTTACCTTCGCTTTTTGCATC | TCTTTCCTTCTCTCCACATCTTG |
| Actin 7 | ATCTCTATGGAAACATCGTTCTCAG | ATCCTCCGATCCAGACACTGTA |
新窗口打开
2 结果
2.1 硫苷生物合成相关基因的组织表达特性
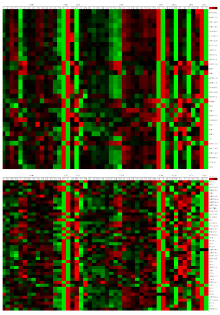
对硫苷生物合成和分解途径的70个酶基因和12个调控基因(表1)在拟南芥不同发育时期79个组织或器官中的组织表达特性分析结果表明(图1),大部分基因在不同组织和器官中的表达模式不同,且硫苷合成相关基因在根、茎、叶等营养组织中的表达量较高,在花和种子中的表达量普遍较低,说明硫苷可能主要在营养器官中合成。其中,脂肪族硫苷合成相关基因在茎、叶等地上营养器官中的表达量明显高于根中的表达量;吲哚族硫苷合成相关基因则在根、茎和叶中有较高的表达量,推测脂肪族硫苷主要在地上营养组织中合成,而吲哚族硫苷在地下和地上营养组织中均有合成。另外,同一基因在相同组织和器官不同发育时期的表达量通常有一定的差异,说明相关基因的表达可能具有时期特异性。脂肪族硫苷氨基酸侧链延伸基因MAM1在地上营养组织中的表达量较高,MAM3在根中的表达量较高;次级结构修饰基因FMOGSL-OX1、FMOGSL-OX3和FMOGSL-OX5在地上组织中的表达量较高,FMOGSL- OX2在花中的表达量较高,FMOGSL-OX4则在根中的表达量较高。而吲哚族硫苷侧链修饰相关基因(CYP81F2、CYP81F3、CYP81F4、IGMT1和IGMT2)在根中的表达量较高。说明脂肪族硫苷的结构修饰可能在植物发育不同时期和组织中均有发生,而吲哚族硫苷的结构修饰可能主要发生在根中。
此外,脂肪族硫苷合成的调控基因MYB28和MYB29的表达量较高,且与脂肪族硫苷合成途径酶基因的表达模式相似,而MYB76的表达量较低,说明该基因的功能可能相对较弱。同理,吲哚族硫苷合成的调控基因MYB34和MYB51的表达量相对较高,而MYB122的表达量则较低。其中MYB34在根和地上组织中均表达,而MYB51主要在地上组织中表达,表明MYB34调控地上和地下组织中硫苷的合成,而MYB51主要调控地上组织中硫苷的合成。在硫苷降解途径中,黑芥子酶基因TGG4在根中的表达量较高,PEN2和PEN3在大部分营养组织中的表达量较高,NSP2则在种子中的表达量较高(图1),说明不同硫苷降解酶基因在植物防御系统中可能负责相应组织中硫苷的水解过程。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1拟南芥硫苷代谢途径相关基因表达谱
顶部横坐标为79个拟南芥不同发育时期的组织。1—7:分别为野生型拟南芥(Wt)7、17、15、8、和21 d的根系;8—10:分别为Wt 7 d的子叶下胚轴、Wt 21 d第1和第2节茎;11—27:分别为Wt 7 d的子叶、Wt 7 d的第1和第2片叶、Wt 10 d的第4片叶、gl1-T突变体10 d的第4片叶、Wt 17 d的第2、4、6、8、12片叶、gl1-T突变体17 d的第12片叶、Wt 17 d的叶柄、叶片中部和叶片上半部分、Wt 15 d的叶、Wt 35 d的老叶、Wt 21 d的茎生叶;28—38:分别为Wt 7、8、21、22、23 d的地上部分组织、Wt 7、14、21 d的营养莲座叶;39—49:分别为Wt 7 d的营养组织和幼叶、营养组织、Wt 14 d抽薹前的茎尖、Wt和突变体clv3-7、lfy-12、ap1-15、ap2-6、ufo-1、ap3-6、ag-12 21 d抽薹后的花序;50—61:分别Wt为21 d发育时期9、10/11、12的花、突变体clv3-7、lfy-12、ap1-15、ap2-6、ufo-1、ap3-6、ag-12 21 d发育时期12的花、Wt 21 d发育时期15的花和28 d的花;62—71:分别为Wt 21 d发育时期15的花梗、Wt 21 d发育时期12、15的花萼、花瓣和雄蕊、Wt 6周的成熟花粉、Wt 21 d发育时期12、15的心皮;72—79:分别为Wt 8周种子发育时期3、4、5的角果和种子发育时期6、7、8、9、10的种子。右边纵坐标为硫苷合成相关基因。基因名后缀AGSL表示脂肪族硫苷合成相关基因,IGSL表示吲哚族硫苷合成相关基因,无标注的表示共有途径的基因
-->Fig. 1Expression analysis of glucosinolate biosynthetic genes in Arabidopsis
The top abscissa indicates 79 Arabidopsis tissues across different developmental stages. 1-7: Indicates the roots of Wild type (Wt) at 7, 17, 15, 8 and 21 days, respectively; 8-10: Indicates the hypocotyl of Wt at 7 days, the 1st node and 2nd internode of Wt at 21 days, respectively; 11-27: Indicates the cotyledons, leaves 1 and 2 of Wt at 7 days, the 4th rosette leaf of Wt and gl1-T at 10 days, the 2nd, 4th, 6th, 8th, 10th, 12th rosette leaf of Wt at 21 days, the 12th rosette leaf of gl1-T at 21 days, the petiole, proximal half and distal half of leave of Wt at 17 days, the leaf of Wt at 15 days, the senescing leaves of Wt at 35 days, the cauline leaves of Wt at 21 days, respectively; 28-38: Indicates the green parts of Wt at 7, 8, 21, 22 and 23 days, the vegetative rosette of Wt at 7, 14 and 21 days, respectively; 39-49: Indicates the shoot apex of vegetative and young leaves, and vegetative of Wt at 7 days, the shoot apex of Wt at 14 days, the inflorescence after bolting of Wt, clv3-7, lfy-12, ap1-15, ap2-6, ufo-1, ap3-6, ag-12 at 21 days, respectively; 50-61: Indicates the flowers of Wt at stage 9 and 10/11 at 21 days, the flowers at stage 12 of Wt, clv3-7, lfy-12, ap1-15 , ap2-6, ufo-1, ap3-6, ag-12 at 21 days, the flowers of Wt at stage 15 at 21 and 28 days; 62-71: Indicates the pedicels of flowers of Wt at stage 15, the sepals, petals and stamens of flowers of Wt at stage 12 and 15 at 21 days, the mature pollen of Wt at 6 weeks, the carpels of flowers of Wt at stage 12 and 15 at 21 days; 72-79: Indicates the siliques of Wt at seeds stage 3, 4 and 5 at 8 weeks, the seeds at stage of Wt 6, 7, 8, 9 and 10 at 8 weeks, respectively. The right ordinate indicates the genes involved in glucosinolate biosynthesis pathway, where the genes with AGSL indicate aliphatic glucosinolate biosynthetic genes, the genes with IGSL indolic glucosinolate biosynthetic genes, the others indicate the genes involved in the common pathway
-->
为验证芯片数据的分析结果,选择10个分别参与脂肪族和吲哚族硫苷合成(包括前体氨基酸侧链延长、核心结构合成和次级修饰3个阶段)(表1和表2)及调控的关键基因,采用qPCR法分析它们在拟南芥中的表达模式。结果表明,10个硫苷合成相关基因在营养组织中的表达量较高,在种子组织中的表达量则较低。其中,脂肪族硫苷合成相关基因MAM1、AOP2、CYP79F1、CYP83A1和MYB28主要在茎、叶或角果中优势表达,在根、花和种子中的表达量相对较低;吲哚族硫苷合成相关基因IGMT1、ASA1、CYP83B1、CYP79B2和MYB34在根、茎或叶中的表达量较高,在角果、花和种子中的表达量则相对较低(图2)。qPCR法分析的结果与芯片数据分析的结果一致性较好,均证明脂肪族硫苷合成相关基因主要在地上营养组织中表达,而吲哚族硫苷合成相关基因在地上和地下营养组织中均有较高的表达水平。仅AOP2、ASA1和MYB34的表达量略有差异,但是两组数据的总体表达模式仍一致。例如,AOP2在芯片数据的叶、茎和花器官中均有较高的表达量,在qPCR数据的叶和茎中也具有较高的表达量,但是在花中的表达量则相对较低(图1和图2)。这是因为两组数据的取样时期和组织不同所致,该基因在芯片数据的花中的表达量通常也较低,仅在特定时期的花器官中具有相对较高的表达量(图1)。
2.2 硫苷生物合成相关基因受生物胁迫的表达谱
对硫苷生物合成相关基因在9组生物胁迫芯片数据(AT49、AT50、AT51、AT59、AT63、AT84、AT90、AT110和AT112)中的表达谱分析结果表明,硫苷生物合成途径部分酶基因的表达明显受到病原细菌、真菌和虫害的胁迫诱导(图3)。在4组病原菌胁迫数据中,硫苷合成和降解相关基因明显受到细菌性病原菌的抑制表达。其中,假单胞杆菌Pseudomonas G62(AT112)浸染植株后,多数硫苷合成相关基因的表达受到抑制,仅有部分下游基因的表达未受影响(如MVP1、NSP2、NSP5、PEN3、BZO1、CHY1、TGG2和GSH1)。在丁香假单胞杆菌(Pseudomonas syringae)的胁迫下,除了部分次级修饰基因(FMOGS-OX2、FMOGS-OX4、FMOGS-OX5和GS-OH),脂肪族硫苷合成相关基因的表达量较低,而吲哚族硫苷合成和降解相关基因的表达量增加(图3)。说明脂肪族硫苷合成相关基因的表达受到该病原菌的抑制,吲哚族硫苷合成相关基因的表达受到诱导。在真菌胁迫数据中,仅木霉菌(Trichoderma spp,AT50)能够同时诱导脂肪族和吲哚族硫苷合成相关基因上调表达,其他真菌胁迫下脂肪族和吲哚族硫苷合成相关基因的表达模式通常相反(图3)。例如,当植株受到灰霉病菌(Botrytis cinerea)、甘蓝链格孢菌(Alternaria brassicicola)和白粉菌(Erysiphe orontii)(AT51)侵染后,多数脂肪族硫苷合成相关基因的表达受到抑制,而吲哚族硫苷合成和降解相关基因的表达受到诱导。相似地,疫霉菌(Phytophthora)侵染拟南芥叶片后,吲哚族硫苷合成相关基因的表达量迅速增加,脂肪族硫苷合成相关基因的表达量则较低。但随着浸染时间的增加,吲哚族硫苷合成相关基因的表达量随之下降,而脂肪族硫苷合成相关基因的表达量则上升。此外,发现部分脂肪族和多数吲哚族硫苷合成相关基因的表达还受到壳聚糖的诱导(AT84)。以上结果表明,吲哚族硫苷合成相关基因的表达更容易受到病原菌的诱导。
通过对多组抗虫芯片数据的分析发现,脂肪族硫苷合成和降解相关基因的表达受到菜青虫的胁迫诱导,而吲哚族硫苷合成和降解相关基因的表达明显受到西花蓟马、桃蚜(Myzus persicae,AT49)和粉虱(AT63)的胁迫诱导(图3)。说明不同硫苷组分的抗虫害作用可能不同,其中吲哚族硫苷可能对粉虱、西花蓟马和桃蚜有一定的防御作用。此外,本研究发现水杨酸(salicylic acid,SA)能够诱导吲哚硫苷合成途径大部分基因高表达(图3),说明吲哚族硫苷在植物防御系统中的作用依赖SA信号途径。
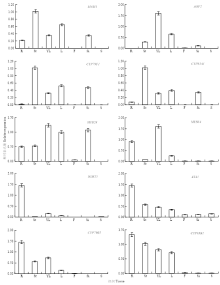 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2硫苷合成相关基因在拟南芥中的表达模式
R、St、YL、L、F、Si和S分别表示拟南芥的根、茎、幼叶、成熟叶、花、角果和成熟种子
-->Fig. 2Expression patterns analyses of glucosinolate biosynthesis genes in Arabidopsis by qPCR
R: Root; St: Stem; YL: Young leaf; L: Leaf; F: Flower; Si: Silique and S: Mature seed
-->
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3拟南芥硫苷生物合成基因受病、虫害胁迫诱导的表达谱
顶部横坐标为芯片探针号,AT112、AT49、AT50、AT51、AT59、AT84、AT110、AT63和AT90分别为拟南芥受假单胞杆菌)、西花蓟马、桃蚜、木霉菌、灰霉病菌、甘蓝链格孢菌、白粉菌、粉虱、吡虫啉、噻虫胺和水杨酸诱导的芯片数据;右边纵坐标为硫苷合成相关基因,基因名后缀AGSL表示脂肪族硫苷合成相关基因,IGSL表示吲哚族硫苷合成相关基因,无标注的表示共有途径的基因
-->Fig. 3The expression profiles of Arabidopsis glucosinolates biosynthesis genes induced by pathogen and insect stresses
The top abscissa indicates probes, where AT112, AT49, AT50, AT51, AT59, AT84, AT110, AT63 and AT90 are the Arabidopsis probes induced by Pseudomonas G62, Frankliniella occidentalis, Myzus persicae, Trichoderma spp, Botrytis cinerea, Alternaria brassicicola, Erysiphe orontii, Bemisia tabaci type B, imidacloprid, clothianidin and salicylic acid, respectively. The right ordinate indicates the glucosinolate biosynthetic genes. The genes with AGSL indicate aliphatic glucosinolate biosynthetic genes, the genes with IGSL indolic glucosinolate biosynthetic genes, the others indicate the genes involved in the common pathway
-->
2.3 硫苷生物合成相关基因受非生物胁迫诱导的表达谱
如图4所示,脂肪族硫苷合成相关基因主要在地上组织中表达,吲哚族硫苷合成相关基因主要在根中表达,仅有少数基因在悬浮细胞中表达,说明硫苷主要在地上和地下组织中合成。而且大部分上游基因(包括吲哚族和脂肪族)在地上组织和根中的表达受到多种非生物环境因子的胁迫诱导。例如,脂肪族硫苷合成上游基因BCAT4、LeuD1、LeuD2、BAT5、ST5b、GSTF11和IPMDH3在地上组织和根中的表达均受到低温(4℃)、渗透胁迫、高盐、UV-B和高温(38℃)的抑制,但是受到干旱和伤害的诱导而上升,说明这些环境因子对脂肪族硫苷的合成有影响。同理,UGT74B1、GSTF9、CYP79B2、CYP79B3、CYP81B1等基因在地上组织中的表达量较低,但是在根中的表达量相对较高且明显受到低温、高盐、渗透胁迫和高温的抑制,证明吲哚族硫苷的合成受到这些非生物因子的胁迫诱导。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4硫苷生物合成相关基因受非生物胁迫诱导的表达情况
顶部横坐标分别为拟南芥苗期地上部分组织、根系和悬浮细胞受低温、高温、高盐、干旱、渗透胁迫、UV-B辐射和机械伤害等胁迫诱导处理;右边纵坐标为硫苷合成相关基因,其中基因名后缀AGSL表示脂肪族硫苷合成相关基因,IGSL表示吲哚族硫苷合成相关基因,无标注的表示共有途径的基因
-->Fig. 4Expression patterns of Arabidopsis glucosinolate genes under abiotic stresses
The top ordinate indicates the experimental treatments of Arabidopsis seedling green parts, roots, and cell culture under cold, heat, salt, drought, osmotic, UV-B, and wounding stresses, respectively. The right vertical axis is genes involved in glucosinolates biosynthesis pathway, where the genes with AGSL indicate aliphatic glucosinolate biosynthetic genes, the genes with IGSL indicate indolic glucosinolate biosynthetic genes, the others indicate the genes involved in the common pathway
-->
但是,次级修饰和降解相关基因没有明显受到上述非生物胁迫因子的诱导表达(图4)。其中,脂肪族硫苷次级修饰相关基因(FMOGS-OX1和FMOGS-OX1-3)在地上组织中有较高的表达水平,而吲哚族硫苷次级修饰相关基因(CYP81F1-4、IGMT1和IGMT1)在根中有较高的表达水平。同理,硫苷降解酶基因(TGG2、TGG4、PEN2和MBP1)和调控基因(MYB29、MYB76、MYB51、MYB122和bHLH05)在地上组织和根中也具有相对较高的表达量(图4)。以上结果进一步证明脂肪族和吲哚族硫苷可能分别在地上和地下组织中合成,并与相应的黑芥子酶基因组成一个紧密的防御体系,通过响应相关因子的胁迫信号来调控硫苷的合成以抵御植株受到的非生物逆境胁迫。
2.4 硫苷生物合成相关基因受激素诱导的表达模式
如图5所示,经过吲哚乙酸(IAA)和脱落酸(ABA)处理3 h后硫苷合成相关基因的表达量明显降低,说明它们对硫苷合成可能具有抑制作用。经过生长素极性运输抑制剂(2,4,6T、PCIB、TIBA和NPA)处理后,大部分硫苷合成相关基因的表达量无明显变化,说明生长素在低水平时对硫苷合成没有影响,但是积累到较高浓度时则会抑制硫苷的合成。经过油菜素内酯(brassinolide,BL)、乙烯合成酶 (1-aminocycropro-pane-1-carboxylic acid,ACC)、玉米素(zeatin)、GA和茉莉酸甲酯(methyl jasmonate,MJ)处理后,脂肪族硫苷合成相关基因的表达量没有明显的变化,少数吲哚族硫苷合成相关基因的表达量增加,表明这些激素对硫苷生物合成的影响相对较低。但是经过乙烯抑制因子AgNO3处理后,部分吲哚族硫苷合成相关基因(例如ST5a、UGT74B1、ASA1、CYP81F2、MYB51和MYB122)的表达量明显增加,表明内源性乙烯可能对吲哚族硫苷的合成具有调控作用。GA和BR抑制因子对硫苷合成相关基因的表达没有明显的影响,并且GA和BR信号通路关键突变体gal-3、gal-5和det2-1中硫苷合成相关基因的表达量也无明显变化(图5),说明这两种激素对硫苷的生物合成没有影响。此外,大多数脂肪族硫苷合成相关基因在植株缺硫24 h后表达量明显降低,而吲哚族硫苷合成相关基因的表达量无明显的变化,说明硫对脂肪族硫苷的生物合成具有重要的作用(图5)。还发现部分硫苷合成相关基因(例如CYP79B2、MYB51和TGG2)在22℃时的表达水平比4℃时高,说明温度对硫苷的生物合成也有影响(图5)。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5硫苷生物合成相关基因受激素诱导的表达情况
顶部横坐标分别为拟南芥幼苗和种子受细胞分裂素(CK)、赤霉素(GA)、吲哚乙酸(IAA)、脱落酸(ABA)、油菜素内酯(BL)、乙烯合成酶(ACC)、玉米素和茉莉酸甲酯(MJ)诱导的试验处理;右边纵坐标为硫苷合成相关基因,其中基因名后缀AGSL表示脂肪族硫苷合成相关基因,IGSL表示吲哚族硫苷合成相关基因,无标注的表示共有途径的基因
-->Fig. 5Expression analysis of Arabidopsis glucosinolate biosynthetic genes under hormone inductions
The top ordinate is the experimental treatments of Arabidopsis seedlings and seeds induced by CK, GA, IAA, ABA, BL, ACC, and MJ, respectively; The right vertical axis indicates the genes involved in glucosinolates biosynthesis pathway, where the genes with AGSL indicate aliphatic glucosinolate biosynthetic genes, the genes with IGSL indicate indolic glucosinolate biosynthetic genes, the others indicate the genes involved in the common pathway
-->
2.5 硫苷生物合成途径的互作调控网络
脂肪族和吲哚族硫苷代谢支路内相关基因的表达模式相关性较高(图6)。MYB28转录因子基因的表达模式与CYP79F1、CYP79F2、FMOG-OX3和UGT74C1酶基因的表达模式相关性较高,说明它们可能是MYB28调控的下游靶基因。同理,MYB29与FMOG-OX1和AOP2的表达模式也非常相似,说明它们可能为MYB29的共调控基因。以上结果进一步证明MYB转录因子对硫苷生物合成具有重要的调控作用,且存在共调控机制。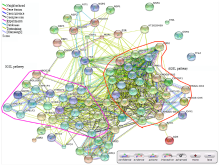 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6拟南芥硫苷生物合成途径相关基因的共表达网络
彩色的线代表不同数据来源的相关性分析结果;IGSL pathway和AGSL pathway分别表示吲哚族(紫色)和脂肪族(红色)硫苷合成途径相关基因;基因名称如
-->Fig. 6The connection network of Arabidopsis glucosinolate biosynthesis genes
Different lines with color represent the types of evidence for the association; IGSL pathway and AGSL pathway indicates indolic (purple) and aliphatic (red) glucosinolate biosynthesis pathway, respectively; the corresponding names of candidate genes were listed in
-->
3 讨论
硫苷广泛分布于十字花科植物的不同组织中,同一植物不同组织、不同生育期硫苷的组分和含量差异很大,具有动态变化的特征。通常,种子中硫苷的含量明显高于其他组织和器官。例如,甘蓝型油菜种子中硫苷的含量高达180 µmol·g-1,而叶片中硫苷的含量一般只有10—20 µmol·g-1[33]。但是种子自身合成硫苷的量却较少,说明植物体内存在一个将“源”组织和器官中合成的硫苷运输到“库”(种子)中的转运系统。该转运系统最近已经得到试验证实[34-36]。本研究基于大量芯片数据,系统分析了硫苷合成、降解、调控等82个相关基因在拟南芥79个不同发育时期和组织中的时空表达特性(图1)。结果显示硫苷合成相关基因在营养器官中高表达,在贮藏器官中低表达,证实了植物体内确实存在一个硫苷的转运系统,负责将营养组织中合成的硫苷转运到种子组织中积累。此外,硫苷代谢途径中同源基因的表达模式相似,但有一定的差别,说明同源基因间存在部分功能冗余。前人研究证明,R2R3-MYB类转录因子对吲哚族和脂肪族硫苷的生物合成具有重要的调控作用[14-16]。在拟南芥中有6个2R-MYB基因参与调控硫苷的生物合成(MYB28、MYB29、MYB34、MYB51、MYB76和MYB122)。其中,MYB28、MYB29和MYB67调控脂肪族硫苷的生物合成,MYB28为主效基因[37-38];而MYB34、MYB51和MYB122调控吲哚族硫苷的生物合成,MYB34为主效基因[16,39]。本研究表达谱分析结果表明,脂肪族硫苷合成的MYB调控基因和酶基因主要在地上部分营养组织中表达,而吲哚族硫苷合成的MYB调控基因和酶基因在地下和地上部分营养组织中均有表达(图1)。共表达分析结果表明,MYB调控基因与硫苷合成途径酶基因的表达模式相关性较高(图6)。例如,MYB28与CYP79F1、FMOG-OX3、UGT74C1和CYP79F1酶基因的表达模式相关性较高,说明它们可能是MYB28调控的下游靶基因。其中,CYP79F1和CYP79F2已经被证明是MYB28的靶基因[15],证明本研究结果的准确性。以上结果表明,调控基因和结构基因共同组成了一个复杂而精细的代谢调控网络,实现对硫苷生物合成的精细调控。
硫苷作为十字花科植物特有的一类次生代谢产物,其降解产物具有多种生化活性,对植物的防御系统和动物的健康均非常重要[2-7]。在生产中,硫苷及其降解产物具有双重作用。例如在油菜生产中,一方面硫苷的降解产物是菜籽饼粕中主要的抗营养因子,严重影响其作为动物饲料的食用安全性和适口性,限制了菜籽饼粕蛋白质资源的开发利用,同时也不利于油菜作为饲草或油-菜兼用的开发应用。另一方面,硫苷又对油菜的抗性和营养保健功能有重要作用,具有较强的防腐和抗菌作用,可作为杀虫剂、杀菌剂、抗微生物制剂和天然的生物熏蒸消毒剂等[40-42]。在植物的防御体系中完整的硫苷并不具有生物学活性,介导植物与昆虫互作的主要是经黑芥子酶水解的硫苷降解物,二者构成了熟知的“芥子油弹(mustard oil bomb)”,在植物防御系统中起重要作用[43]。本研究通过分析拟南芥中硫苷合成相关基因在不同组织和时期、不同生物和非生物胁迫下的表达特性,发现大多数情况下硫苷降解和合成相关基因的表达模式相似(图1),它们共同组成了一个紧密的生物防御体系,以抵御植株可能受到的逆境胁迫。硫苷降解相关基因在不同组织和时期、不同生物和非生物胁迫下的表达谱存在明显的差异,表明降解酶基因受不同胁迫因子的诱导,进而产生相应的硫苷降解产物以抵御各种逆境胁迫。其中,吲哚族硫苷合成相关基因更易受病、虫害的胁迫诱导表达(图3),推测该类物质是拟南芥中主要的化学防御物质,对其防御系统有重要的作用。另外,本研究还发现,部分基因的表达受到生物和非生物胁迫因子的共同诱导,例如脂肪族硫苷合成相关基因BCAT4、LeuD1、BAT5和吲哚族硫苷合成相关基因UGT74B1、CYP79B2、CYP81B1等同时受到生物和非生物的胁迫诱导表达(图3和图4)。
基因的表达模式对推断基因功能、基因间互作等具有重要的作用。虽然因为转录后修饰等机制,导致基因的转录水平与其蛋白水平可能不完全一致,但是表达谱是基因表达为相应蛋白、行使其生物学功能的重要前提,可以为揭示基因的时空表达规律和功能提供重要的线索。本研究结果为进一步在蛋白水平揭示硫苷的合成和积累规律,并在具有重要农艺性状的十字花科作物中开展相关研究提供了重要的参考。
4 结论
对拟南芥硫苷生物合成途径82个结构基因和调控基因的组织和生物及非生物胁迫诱导的表达谱进行系统的分析表明,硫苷合成相关基因主要在营养组织中高表达、在生殖器官中低表达。吲哚族硫苷生物合成相关基因的表达容易受到病原菌、虫害、高温等逆境胁迫的影响,并且硫苷合成途径相关基因的表达模式具有较高的相关性。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}