Optimization of Cotton Mesophyll Protoplast Transient Expression System
LI Qing,, YU HaiPeng, ZHANG ZiHao, SUN ZhengWen, ZHANG Yan, ZHANG DongMei, WANG XingFen, MA ZhiYing, YAN YuanYuan,*College of Agronomy, Hebei Agricultural University/State Key Laboratory of North China Crop Improvement and Regulation/Key Laboratory for Crop Germplasm Resources of Hebei, Baoding 071001, Hebei
Abstract 【Objective】Cells of true leaves can well mimic the plant endogenous situation. It is an efficient way for expediting cotton functional study to establish an effective transient expression system using cotton protoplasts obtained from true leaves.【Method】The enzyme combination of cellulose and macerozyme were used to isolate protoplasts from true leaves of Gossypium hirsutum L. acc. TM-1. The effects of osmotic pressure, components of digestion buffer and digestion time on protoplast yield were studied and the validity of protoplasts were compared under different mannitol concentration and digestion time. To improve the transformation efficiency of cotton protoplast, the effects of mannitol and PEG concentration and buffers for protoplast culture were subsequently studied. In order to verify the optimized transient expression system, the vector 35S:LTP-GFP was constructed and transformed into protoplasts of Arabidopsis and cotton and tobacco epidermal cells followed by observation of fusion protein localization.【Result】High concentration of CaCl2 in the digestion buffer significantly inhibited the isolation of protoplast from cotton true leaves, which was opposite of that using cotyledon. 10 mmol·L -1 CaCl2 was employable for digestion buffer to isolate cotton protoplasts from true leaves. Mannitol concentration significantly affected protoplast yield that peaked under mannitol concentration of 0.5 mol·L-1, and protoplast validity decreased moiety under 0.4 mol·L-1 mannitol, suggesting that 0.5 mol·L-1 mannitol was most suitable to maintain the osmotic pressure of cotton protoplasts. Cotton protoplasts displayed suitable size when isolated from newly flattened true leaves, while protoplast enlarged and yield decreased when produced from young leaves flattened 5 days. The protoplasts dissociate slowly until being digested 9 h when the yield reached the peak. The transformation efficiency was greatly improved under isotonic condition of 40% PEG buffer. While hypotonic condition that is commonly applied to facilitate transformation was against the entrance of exogenous DNA into cotton protoplasts. After transformation, the protoplast ruptured abundantly in WI buffer,whereas the shape maintained well in W5 buffer adding 0.5 mol·L-1 mannitol. The transformation efficiency was improved to 90% using the optimized transient expression system. The subcellular location analysis results showed consistent GFP signal in protoplasts of cotton and Arabidopsis true leaf and epidermal cells of tobacco leaf.【Conclusion】Our study has optimized the cotton mesophyll protoplast transient expression system, which could produce 8.10×10 6·mL-1 fine protoplasts with validity above 95% and transformation efficiency reached to 90%. This system is applicable for analysis of subcellular location, protein interaction and research on metabolism and regulation network. Keywords:cotton;TM-1;isolation of protoplasts;transient expression
PDF (4819KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 李青, 鱼海鹏, 张子豪, 孙正文, 张艳, 张冬梅, 王省芬, 马峙英, 阎媛媛. 棉花真叶原生质体分离及瞬时表达体系的优化. 中国农业科学, 2021, 54(21): 4514-4524 doi:10.3864/j.issn.0578-1752.2021.21.003 LI Qing, YU HaiPeng, ZHANG ZiHao, SUN ZhengWen, ZHANG Yan, ZHANG DongMei, WANG XingFen, MA ZhiYing, YAN YuanYuan. Optimization of Cotton Mesophyll Protoplast Transient Expression System. Scientia Agricultura Sinica, 2021, 54(21): 4514-4524 doi:10.3864/j.issn.0578-1752.2021.21.003
开放科学(资源服务)标识码(OSID):
0 引言
【研究意义】棉花是世界上最重要的经济作物之一,在国民经济中占据重要地位。随着基因组测序技术的发展,多个野生棉和栽培棉(Gossypium)基因组陆续完成测序与组装,并被升级[1,2,3,4,5]。这些高质量的基因组序列为解析棉花许多重要的进化事件与机制提供了便利,更加快了重要农艺性状相关位点的挖掘。然而,四倍体栽培棉种基因组较大,陆地棉遗传标准系TM-1(Gossypium hirsutum L. acc. TM-1)基因组包含74 350个编码蛋白的基因[5],大量重要基因的功能急需解析。但棉花突变体资源稀少,遗传转化周期长且转基因效率不高,制约着基因功能研究。利用基因的时空表达特征和调控网络,建立一套快速高通量验证棉花基因功能的方法,是加速棉花功能基因组学研究进程,并能快速获得目标性状的候选功能基因,从而进行基因组精准改良的基础[6]。原生质体无细胞壁障碍,是进行基因遗传转化操作、研究基因表达调控和功能的优良材料,为进行高通量的基因功能研究提供了一种经济、有效的方法。【前人研究进展】原生质体瞬时表达体系因其快速、高效的优势,被广泛应用于亚细胞定位、分子互作及代谢调控网络等研究。目前,已在拟南芥[7]、烟草[8]、小麦[9]、水稻[10,11]等植物中建立了高效的原生质体瞬时表达体系,并广泛应用于基础研究。XIONG等[12]利用大豆叶肉原生质体瞬时表达体系揭示了细胞质类光体结构的形成。结合叶肉原生质体瞬时转化和HPLC-MS技术,GAO等[13]鉴定了调控和合成玉米苯丙噁嗪的相关基因,为改良玉米抗虫能力提供了理论依据。原生质体还被用于挖掘植物抗病相关基因及识别病原菌的效应子[14,15]。棉花原生质体首先从陆地棉纤维游离出,之后从多个棉种的多种外植体分离出有活力的原生质体[16]。孙玉强[17]详细分析了影响棉花原生质体分离的酶液组合,从下胚轴、幼根和叶片中分离出原生质体。李妮娜等[18]进一步分析了叶龄、甘露醇浓度和酶解时间对棉花幼嫩子叶原生质体分离效率的影响,并利用PEG介导的棉花原生质体转化技术,对棉花锌指蛋白GhZFP2进行了亚细胞定位研究。【本研究切入点】尽管已有较为高效的分离棉花子叶原生质体的方法[18],但子叶细胞以薄壁组织为主,不同于真叶的海绵组织,真叶才是植物与环境发生互作的主要部位。因此,利用真叶原生质体进行基因的同源瞬时表达,避免了异源表达可能造成的差异结果,提高基因功能研究的准确性。但目前未见分离棉花真叶原生质体的报道,且已报道的棉花原生质体遗传转化效率较低,尚不能满足高通量研究基因功能的需要。【拟解决的关键问题】本研究运用酶解法分离与纯化棉花真叶叶肉原生质体,对比分析影响原生质体分离与目标基因瞬时转化的因素,优化棉花真叶原生质体瞬时表达体系,为开展高通量基因功能研究提供可靠方法。
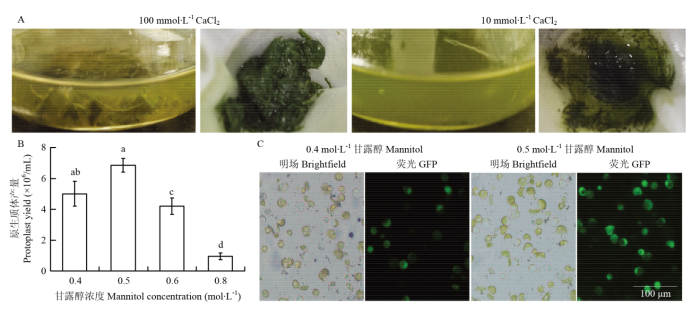
A:不同浓度CaCl2酶解液酶解效果;B:甘露醇对原生质体得率的影响(不同小写字母表示在P<0.05水平差异显著。下同);C:甘露醇对原生质体活力的影响 Fig. 1Effect of CaCl2 and mannitol concentration on protoplast isolation
A: Digestion effect of enzyme buffer with different CaCl2 concentration; B: Effect of mannitol concentration on protoplast yield (different letters show significant difference at P<0.05. The same as below); C: Effect of mannitol on protoplast viability
A:取样真叶;B:不同叶龄原生质体产量;C:老叶原生质体;D:幼叶原生质体 Fig. 2Effect of leave developmental stage on protoplast isolation
A: True leaves for protoplast isolation; B: Protoplast yields using the true leaves at different developmental stages; C: Protoplasts produced from elder leaves; D: Protoplasts produced from newly flattened young leaves
A、C、D:培养液类型、甘露醇和PEG浓度对转化效率的影响;B:荧光显微镜下的转化结果 Fig. 4Optimization of transformation conditions
A, C and D: Effect of protoplast culture solution, mannitol concentration and PEG concentration on transformation efficiency; B: Protoplasts after transformation under fluorescence microscope
ZHANG TZ, HUY, JIANG WK, FANGL, GUAN XY, CHEN JD, ZHANG JB, SASKI CA, SCHEFFLER BE, STELLY DM, HULSE-KEMPA M, WANQ, LIUB L, LIUC X, WANGS, PANM Q, WANGY K, WANGD W, YEW X, CHANGL J, ZHANGW P, SONGQ X, KIRKBRIDER C, CHENX Y, DENNISE, LLEWELLYND J, PETERSOND G, THAXTONP, JONESD C, WANGQ, XUX Y, ZHANGH, WUH T, ZHOUL, MEIG F, CHENS Q, TIANY. Sequencing of allotetraploid cotton (Gossypium hirsutum L. acc. TM-1) provides a resource for fiber improvement Nature Biotechnology, 2015, 33(5):531-537. DOI:10.1038/nbt.3207URL [本文引用: 1]
WANG MJ, TU LL, YUAN DJ, ZHUD, SHENC, LI JY, LIU FY, PEI LL, WANG PC, ZHAO GN, YE ZX, HUANGH, YAN FL, MA YZ, ZHANGL, LIUM, YOU JQ, YANG YC, LIU ZP, HUANGF, LI BQ, QIUP, ZHANG QH, ZHU LF, JIN SX, YANG XY, MINL, LI GL, CHEN LL, ZHENG HK, LINDSEYK, LIN ZX, UDALL JA, ZHANG XL. Reference genome sequences of two cultivated allotetraploid cottons, Gossypium hirsutum and Gossypium barbadense Nature Genetics, 2019, 51(2):224-229. DOI:10.1038/s41588-018-0282-xURL [本文引用: 1]
HUANGG, WU ZG, RICHARDG. P, BAIM Z, LIY, JAMESE. F, HUJ, WANGK, JOHNZ. Y, ZHUY X. Genome sequence of Gossypium herbaceum and genome updates of Gossypium arboreum and Gossypium hirsutum provide insights into cotton A-genome evolution Nature Genetics, 2020, 52(5):516-524. DOI:10.1038/s41588-020-0607-4URL [本文引用: 2]
JASON GW, ELIR, EDWARD SB. On the road to breeding 4.0: Unraveling the good, the bad and the boring of crop quantitative genomics Annual Reveiew of Genetics, 2018, 52(26):1-24. [本文引用: 1]
ZOU JJ, WEI FJ, WANGC, WU JJ, RATNASEKERAD, LIU WX, WU WH. Arabidopsis calcium dependent protein kinase CPK10 functions in abscisic acid and Ca2+ mediated stomata regulation in response to drought stress Plant Physiology, 2010, 154:1232-1243. DOI:10.1104/pp.110.157545URL [本文引用: 1]
JIANGL, WANGJ, LIUZ, WANGL, ZHANGF, LIU GC, ZHONGQ. Silencing induced by inverted repeat constructs in protoplasts of Nicotiana benthamiana Plant Cell Tissue and Organ Culture, 2010, 100:139-148. DOI:10.1007/s11240-009-9629-4URL [本文引用: 1]
SUNH, LANG ZH, ZHUL, HUANG DF. Optimized condition for protoplast isolation from maize, wheat and rice leaves Chinese Journal of Biotechnology, 2013, 29(2):224-234. (in Chinese) [本文引用: 1]
ZHANGH, NIL, LIU YP, WANG YF, ZHANG AY, TAN MP, JIANG MY. The C2H2-type Zinc finger protein ZFP182 is involved in abscisic acid-induced antioxidant defense in rice Journal of Integrative Plant Biology, 2012, 54:500-510. DOI:10.1111/jipb.2012.54.issue-7URL [本文引用: 1]
ZHANGY, XIAO WK, LUO LJ, PANG JH, RONGW, HE CZ. Down regulation of OsPK1, a cytosolic pyruvate kinase, by T-DNA in sertion causes dwarfism and panicle enclosure in rice Planta, 2012, 235:25-38. DOI:10.1007/s00425-011-1471-3URL [本文引用: 1]
XIONGL, LIC, LI HY, LYU XG, ZHAOT, LIUJ, ZUO ZC, LIUB. A transient expression system in soybean mesophyll protoplasts reveals the formation of cytoplasmic GmCRY1 photobody-like structures Science China Life Sciences, 2019, 62(8):1070-1077. DOI:10.1007/s11427-018-9496-5URL [本文引用: 1]
GAOL, SHEN GJ, ZHANG LD, QI JF, CZHANG C P, MAC R, LIJ, WANGL, SAIFU M, WUJ Q. An efficient system composed of maize protoplast transfection and HPLC-MS for studying the biosynthesis and regulation of maize benzoxazinoids Plant Methods, 2019, 15(1):144-157. DOI:10.1186/s13007-019-0529-2URL [本文引用: 3]
PATIENCEC, MARIEE C R. A cassava protoplast system for screening genes associated with the response to South African Cassava Mosaic Virus Virology Journal, 2020, 17(1):184-199. DOI:10.1186/s12985-020-01453-4URL [本文引用: 1]
SU YY, CHEN YE, CHENJ, ZHANG ZJ, GUO JY, CAIY, ZHU CY, LI ZY, ZHANG HY. Effectors of Puccinia striiformis f. sp. tritici suppressing the pathogenic-associated molecular pattern-triggered immune response were screened by transient expression of wheat protoplasts International Journal of Molecular Sciences, 2021, 22(9):4985-5003. DOI:10.3390/ijms22094985URL [本文引用: 1]
SUNY Q. Research on cotton protoplasts culture and fusion Journal of Huazhong Agricultural University 2011, 30(6):784-786. In Chinese. (in Chinese) [本文引用: 1]
LI NN, DING LY, ZHANG ZY. Establishment of isolation of cotton mesophll protoplast and transient expression system of target gene The Crop Journal, 2014, 40(2):231-239. (in Chinese) [本文引用: 8]
LI JY, LIU LB, DINGB, YANG HX, WUQ, ZHANGC. Research progress on plant protoplasts isolation and culture Molecular PlantBreeding, 2021: 1-20. https://kns.cnki.net/kcms/detail/46.1068.S.20210308.1639.026.html . (in Chinese) URL [本文引用: 3]
LI XY. A transient expression assay using Arabidopsis mesophyll protoplasts Bio-Protocol, 2011, 1(10):e70. [本文引用: 1]
FU LL, YANG XY, ZHANG XL, WANG ZW, FENG CH, LIU ZX, JIANG PY, ZHANG JL. Regeneration and identification of inter-specific asymmetric somatic hybrids obtained by donor-recipient fusion in cotton Chinese Science Bulletin, 2009, 54:2219-2227. [本文引用: 1]
EECKHAUTT, VAN HW, BRUZNICANS, LEUSL, VAN HJ. Somaclonal variation in chrysanthemum × morifolium protoplast regenerants Frontiers in Plant Science, 2020, 11:607171-607187. DOI:10.3389/fpls.2020.607171URL [本文引用: 1]
ELŻBIETAJ, ALEKSANDRAN, ANNAM. Progress towards sugar beet improvement through somatic hybridization: I. Inactivation of nuclei and cytoplasm in donor and recipient protoplasts Acta Societatis Botanicorum Poloniae, 1995, 64(4):341-347. DOI:10.5586/asbp.1995.044URL [本文引用: 1]
XIEX, JIANG JM, WANGY, REN MJ. Study on the method of protoplast isolation and transformation of Sorghum bicolor Seed, 2019, 30(8):43-46. (in Chinese) [本文引用: 1]
SANDHYAD, JOGAMP, ALLINI VR, ABBAGANIS, ALOKA. The present and potential future methods for delivering CRISPR/Cas9 components in plants Biotechnology & Genetic Engineering Reviews, 2020, 18(1):25-36. [本文引用: 1]
NICOLIAA, FÄLTA, HOFVANDERP, ANDERSSONM. Protoplast-based method for genome editing in tetraploid potato Methods in Molecular Biology, 2021, 2264(24):177-186. [本文引用: 1]
WU SP, ZHU HC, LIU JX, YANG QS, SHAO XH, BI FC, HU CH, HUO HQ, CHEN KL, YI GJ. Establishment of a PEG-mediated protoplast transformation system based on DNA and CRISPR/Cas9 ribonucleoprotein complexes for banana Plant Biology, 2020, 20(1):1-10. DOI:10.1111/plb.12676URL [本文引用: 1]
JIN DM, CHOI SH, LEE MH, JIE EY, AHN WS, JOO SJ, AHN JW, JO YD, AHN SJ, KIM SW. Development of a rapid selection system for salt-resistant mutants of nicotiana benthamiana through protoplast culture after gamma irradiation Plants, 2020, 9(12):1720-1733. DOI:10.3390/plants9121720URL [本文引用: 1]
HUANG MK, ZHANGL, ZHOU LM, WANG MZ, YUNG WS, WANG ZL, DUAN SW, XIAO ZX, WANG QW, WANGX, LI MW, LAM HM. An expedient survey and characterization of the soybean JAGGED 1 (GmJAG1) transcription factor binding preference in the soybean genome by modified ChIP mentation on soybean protoplasts Genomics, 2021, 113(1):344-355. DOI:10.1016/j.ygeno.2020.12.026URL [本文引用: 1]
SAHABS, HAYDEN MJ, MASONJ, SPANGENBERGG. Mesophyll protoplasts and PEG-mediated transfections: Transient assays and generation of stable transgenic canola plants Methods in Molecular Biology, 2019, 1864:131-152. [本文引用: 1]
ZHOU QY, JIANG ZH, LI YM, ZHANGT, ZHU HL, ZHAOF, ZHAOZ. Mesophyll protoplast isolation technique and flow cytometry analysis of ancient Platycladus orientalis Turkish Journal of Agriculture and Forestry, 2019(3):275-287. [本文引用: 2]
HU YF, SONG DL, GAOL, BABATOPE SA, WANG YB, HUANG HH, ZHANG JJ, LIU HM, LIU YH, YU GW, LIU YJ, LI YP, HUANG YB. Optimization of isolation and transfection conditions of maize endosperm protoplasts Plant Methods, 2020, 16(1):1-15. DOI:10.1186/s13007-019-0534-5URL [本文引用: 1]
ZHAO YW, HUANG ZG, LI HS. EffectS of washing solution on protoplast isolation from Arabidopsis thaliana leave Chinese Agricultural Science Bulletin, 2011, 27(12):187-190. (in Chinese) [本文引用: 2]
ZHUJ, NIEQ, YANG CL, CHENX. Effects of different plasma membrane stabilizers on cell wall regeneration of tobacco protoplasts Journal of Mountain Agriculture and Biology, 2012, 31(3):222-227. (in Chinese) [本文引用: 2]
WANG ZZ, ZHANG SF, HU ZH. Production, culture and plant regeneration of protoplasts from embryogenic callus of Gossypium hirsutum Journal of Integrative Plant Biology, 1998, 40:234-240. (in Chinese) [本文引用: 1]
YANG XY, ZHANG XL, JIN SX, FU LL, WANG LG. Production and characterization of asymmetric hybrids between upland cotton Coker 201 (Gossypium hirsutum) and wild cotton (G. klozschianum Anderss) Plant Cell Tissue and Organ Culture, 2007, 89(2/3):225-235. DOI:10.1007/s11240-007-9245-0URL [本文引用: 1]
ZENG GJ, CHENG YW, HAN SP, LÜY, LU YL, ZHOUC, ZHANG DC, SHEN XL. The establishment of protoplast isolation and transient expresion system in sorghum cultivar BTx623 Biotic Resources, 2021, 43(1):42-49. (in Chinese) [本文引用: 2]
YOO SD, CHOY, SHEENJ. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis Nature Protocols, 2007(2):1565-1572. [本文引用: 1]
ZHANGY, SU JB, DUANS, AOY, DAI JR, LIUJ, WANGP, LI YG, LIUB, FENG DR, WANG JF, WANG HB. A highly efficient rice green tissue protoplast system for transient gene expression and studying light/chloroplast-related processes Plant Methods, 2011, 7(1):30-44. DOI:10.1186/1746-4811-7-30URL [本文引用: 1]
GUO JJ, MORRELL-FALVEYJ L, LABBÉJ L, MUCHEROW, KALLURIU C, TUSKANG A, CHENJ G. Highly efficient isolation of Populus mesophyll protoplasts and its application in transient expression assays PLoS ONE, 2012, 7(9):44908-44917. [本文引用: 1]
 ,, 鱼海鹏, 张子豪, 孙正文, 张艳, 张冬梅, 王省芬, 马峙英, 阎媛媛
,, 鱼海鹏, 张子豪, 孙正文, 张艳, 张冬梅, 王省芬, 马峙英, 阎媛媛

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT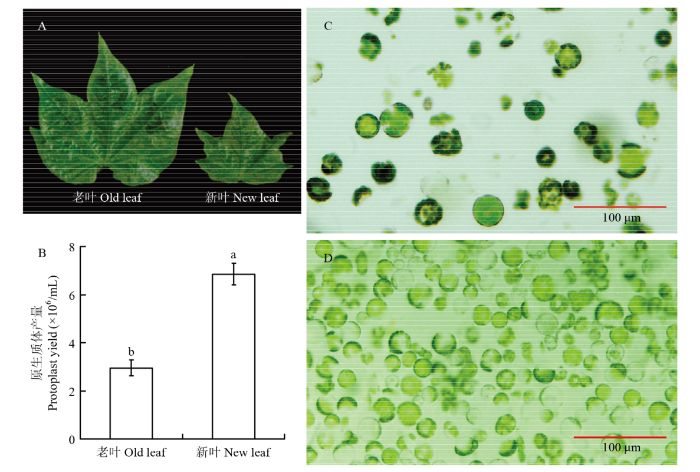 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT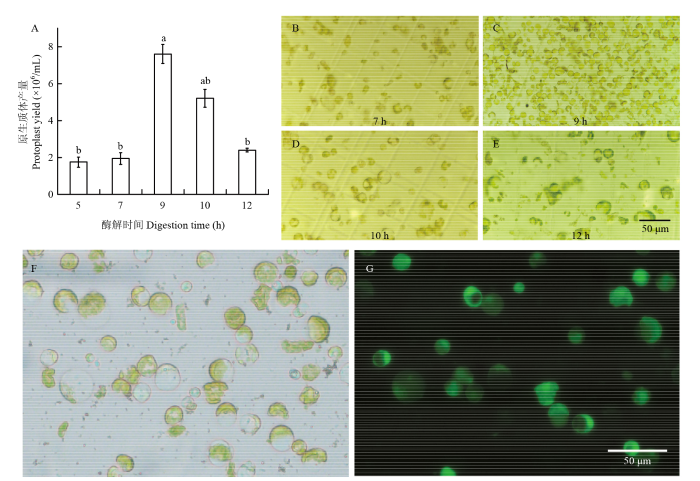 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT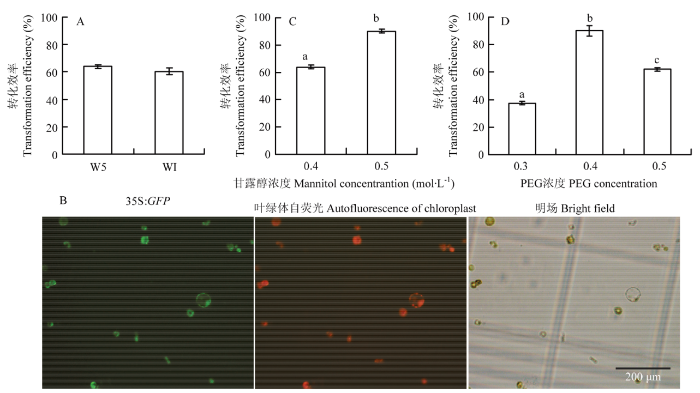 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT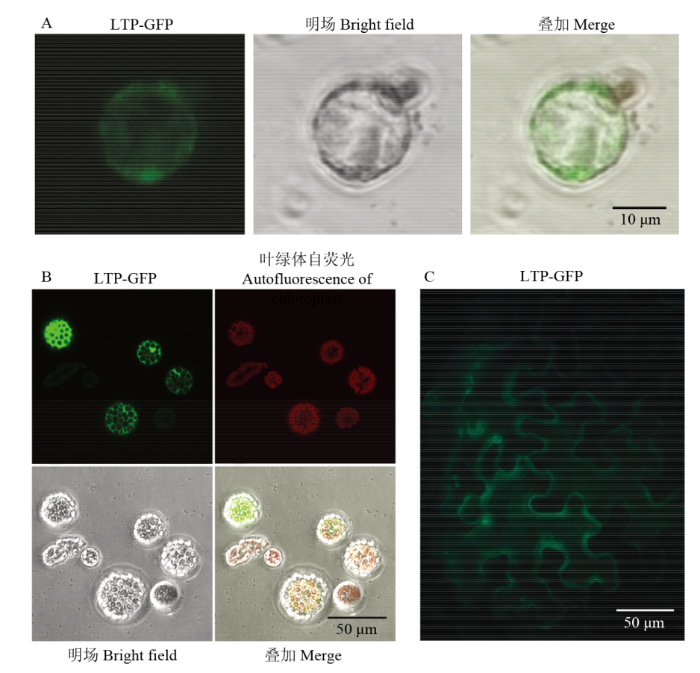 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}